











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkEstudiar la dieta de los anuros ayuda a reconocer parte de las funciones de este grupo biológico en los ecosistemas (Norval, Huang, Mao, Goldberg, & Yang, 2014), principalmente disminuyendo la abundancia de algunos grupos de invertebrados y la herbivoría foliar (Beard, Eschtruth, Vogt, Vogt, & Scatena, 2003). Además, amplía el conocimiento sobre sus historias de vida y la manera en que los organismos utilizan los recursos para asignar energía al crecimiento, al mantenimiento y a la reproducción (Zug, Vitt, & Caldwell, 2001). El estudio de los hábitos alimenticios de los anuros puede resultar complejo, ya que los sitios (Bonansea & Vaira, 2007) y la estacionalidad (Berazategui, Camargo, & Maneyro, 2007) pueden ofrecer recursos tróficos distintos. Asimismo, debido a la ontogenia (Whitfield & Donnelly, 2006) y el sexo (Maneyro, Naya, da Rosa, Canavero, & Camargo, 2004), los anuros pueden presentar diferentes requerimientos alimenticios. Estos factores pueden provocar variaciones intraespecíficas en la composición de la dieta, e influir en la estrategia de forrajeo (Lima & Magnusson, 2000).
La variación intersexual en la dieta de las especies se ha relacionado con la evolución del dimorfismo sexual, específicamente con el tamaño de la cabeza (Houston & Shine, 1993), el período y el comportamiento reproductivo (Lamb, 1984; Donnelly, 1991), los requerimientos energéticos para la producción de gametos (Bull, 2003) y la divergencia en la selección de hábitat (Plummer & Farrar, 1981). La variación intersexual en la dieta de anuros es conocida para pocas especies, entre las cuales se encuentran ránidos (Lamb, 1984), leptodactílidos (Maneyro et al., 2004), hílidos (Miranda, Ebner, Solé, & Kwet, 2006) y bufónidos (Quiroga, Sanabria, & Acosta, 2009); no existiendo casos documentados para la familia Craugastoridae, pese a que es la familia con el mayor número de especies a nivel mundial (Frost, 2018).
Dentro de la familia Craugastoridae, el género Craugastor está compuesto por 115 especies y se distribuye desde el sur de Estados Unidos hasta el noroeste de Colombia y Ecuador, a través de hábitats tropicales y subtropicales (Frost, 2018). Todos los miembros de este género se caracterizan por tener desarrollo directo y varias especies se encuentran entre las más abundantes en los ecosistemas donde habitan (Lieberman, 1986; Urbina-Cardona, Olivares-Pérez, & Reynoso, 2006; Rodríguez-Mendoza & Pineda, 2010), por lo que podrían tener un papel importante en la red trófica y la estructura de las comunidades (Urbina-Cardona & Reynoso, 2009). Los estudios de dieta en especies de la familia Craugastoridae se han centrado en el género Pristimantis (Arroyo, Serrano-Cardozo, & Ramírez-Pinilla, 2008; Garcia, Posso-Gómez, & Cárdenas-Henao, 2015), mientras que otros como Craugastor han recibido menor atención, a pesar de su amplia distribución geográfica y el elevado número de especies que lo componen.
Craugastor rhodopis (Cope, 1867) es una especie que alcanza los 42.3 mm de longitud hocico-cloaca (LHC) y presenta dimorfismo sexual en la talla, siendo las hembras (LHC = 36.7 - 42.3 mm) más grandes que los machos (LHC = 23.8 - 30.4 mm) (Streicher et al., 2014). Esta especie es endémica a México y se distribuye aproximadamente entre los 1 260 y 1 860 msnm, en bosques de encino-pino y bosques de niebla en los estados de San Luis Potosí, Hidalgo, Puebla y Veracruz (Streicher et al., 2014), en la Sierra Madre Oriental de México. Craugastor rhodopis tiene hábitos terrestres y está asociada a sitios con abundante cobertura arbórea y hojarasca. A pesar de ser una especie con alta abundancia en los ensambles de anfibios de los remanentes de bosque donde habita (Murrieta-Galindo, López-Barrera, Gónzalez-Romero, & Parra-Olea, 2013; Meza-Parral & Pineda, 2015), existe poca información acerca de diferentes aspectos de su historia natural e interacción con otras especies (Aguilar-López & Pineda, 2013). Dado que C. rhodopis presenta dimorfismo sexual, es posible que exista variación intersexual en su dieta.
En este estudio se evaluó la composición de la dieta consumida por adultos de C. rhodopis, analizando específicamente si existen diferencias entre sexos en el volumen, la frecuencia y la abundancia de las presas consumidas, y la combinación de estos tres atributos (índice de importancia relativa). Además, se evaluó la relación de la talla y el sexo de las ranas con el volumen, la cantidad de presas y el número de categorías de presa consumidas.
Material y métodos
Área y sitios de estudio: El área de estudio se localiza en la región montañosa central de Veracruz (19°30’12”- 19°02’22” N & 96°48’15”- 97°08’10” W) en un intervalo altitudinal entre 1 000-1 950 msnm, en la Sierra Madre Oriental. La topografía del área de estudio es heterogénea, el clima templado-húmedo con una temperatura media anual entre 12 y 18 °C y una precipitación total anual que varía entre 1 350 y 2 000 mm (Williams-Linera, 2007). El área, originalmente cubierta por bosque de niebla, actualmente está conformada por algunos fragmentos de bosque inmersos en una matriz de ambientes modificados (Williams-Linera, Manson, & Isunza, 2002). Para llevar a cabo las búsquedas de las ranas, se seleccionaron ocho fragmentos de bosque de niebla, con un área entre 0.09 y 3.53 km2, en los cuales la presencia de la especie ya ha sido confirmada (Meza-Parral & Pineda, 2015).
Recolecta de los ejemplares y análisis del contenido estomacal: Para la captura de los individuos de C. rhodopis se realizaron muestreos nocturnos (20:00 - 03:00 h) entre junio y octubre de 2012 (época de lluvias), utilizando la técnica de búsqueda libre con encuentros visuales (Crump & Scott, 1994). Todos los individuos fueron sacrificados con pentobarbital sódico, en un periodo máximo de 4 horas después de su captura (Garcia et al., 2015). Los ejemplares fueron depositados en la Colección de Anfibios y Reptiles del Instituto de Ecología A. C. (CARIE). De cada ejemplar se midió la longitud hocico-cloaca (LHC) y se extrajeron las gónadas para determinar su sexo, y el estómago. No se consideraron las presas del intestino, para permitir comparar con la mayoría de los estudios de dieta en craugastóridos, que solo analizan las presas del estómago. Este estudio se limitó a individuos adultos, a los cuales se les determinó el sexo con base en la presencia de testículos bien desarrollados en machos, y folículos u oviductos convolutos en hembras. Los ejemplares fueron recolectados con los permisos otorgados por la Dirección General de Vida Silvestre de la Secretaría de Medio Ambiente y Recursos Naturales (SGPA/DGVS/03665/06 y SGPA/DGVS/03444/15).
El contenido estomacal fue analizado empleando un microscopio estereoscópico, las presas encontradas fueron identificadas a nivel de orden con ayuda de las claves de Triplehorn & Johnson (2005). En insectos holometábolos, adultos y larvas fueron considerados categorías de presa distintas, y los himenópteros fueron separados en formícidos y no formícidos (Whitfield & Donnelly, 2006). Se contó el número de presas presentes en cada estómago, y se midió su largo y ancho con un calibrador digital (0.01 mm), para posteriormente calcular el volumen de cada una de estas con la fórmula del elipsoide (Crawford, Shepard, & Conner, 2009).
Análisis de datos: Para determinar la importancia de cada categoría de presa i en la dieta de la especie (en conjunto y por sexo), se tomaron en cuenta tres variables: 1) el porcentaje volumétrico de las presas (% V); 2) el porcentaje de frecuencia de ocurrencia (% FO); 3) el porcentaje numérico de las presas consumidas (% N). Posteriormente, se calculó el índice de importancia relativa (IIR) propuesto por Biavati, Wiederhecker y Colli (2004). Para comparar cada variable y el IIR de las categorías de presas entre hembras y machos, se utilizaron gráficas de rango-abundancia (Magurran, 2004).Con el fin de comparar el número de categorías de presas entre machos y hembras se generaron curvas de rarefacción para cada sexo, basándonos en el número de individuos de cada categoría de presa, empleando el programa EstimateS 9.0 (Colwell, 2013). Debido a la diferencia en el número de individuos entre sexos, se realizó una extrapolación de la curva de las hembras (48 individuos), al nivel de la muestra de machos (133 individuos) con mayor número de individuos (Colwell et al., 2012) y posteriormente se recalcularon los intervalos de confianza al 84 % que mimetizan una prueba estadística con un valor de alfa de 0.05 (MacGregor-Fors & Payton, 2013). Se analizó la sobreposición de la dieta entre sexos con el índice de Pianka (1973), considerando los datos de abundancia y el volumen de las presas por separado. Adicionalmente, se evaluó la variación en la composición de la dieta entre sexos con un análisis de similitud (ANOSIM), basado en una matriz de disimilitud de Bray-Curtis (Clarke, 1993).
Para determinar si existe relación entre el tamaño (LHC), el sexo de los ejemplares, o una interacción entres estos factores con el volumen, el número de presas y el número de categorías de las presas fue empleado un modelo lineal mixto, en el caso del volumen, y modelos lineales generalizados para el número de presas y número de categorías de presas. Para el modelo lineal mixto se empleó una función de liga raíz cuadrada, considerando los efectos fijos de la identidad de cada individuo (Crawley, 2007). Para el número de categorías de presas y el número de presas, los modelos lineales generalizados (GLM) fueron generados empleando una distribución Gamma, con la función de liga raíz cuadrada. Todos los análisis fueron realizados en R versión 3.1.3 (R Development Core Team, 2015) y sus extensiones gmodels (Warnes, 2006) y lme4 (Bates, Maechler, Bolker, & Walker, 2015).
Resultados
En total se recolectaron 77 ejemplares adultos (56 machos y 21 hembras) de C. rhodopis, con talla entre 26.3 y 44.3 mm, siendo las hembras en promedio 30 % más grandes ( ± ee = 37.2 ± 3.7 mm, intervalo = 26 - 45.89 mm) que los machos ( ± ee = 28.7 ± 4.8 mm, intervalo = 20.15 - 37.51 mm). Se encontró contenido estomacal en 66 individuos (47 machos y 19 hembras). En total se registraron 181 presas pertenecientes a 20 categorías (18 en machos y 12 en hembras). Fue posible determinar el volumen sólo de 101 presas contenidas en estómagos de 52 individuos (16 hembras y 36 machos). Orthoptera fue la categoría predominante en cuanto a volumen, frecuencia de ocurrencia, número e importancia relativa, seguida por Coleoptera y Araneae. Este patrón se observó al analizar toda la muestra en conjunto, así como al separarlas por sexo (Figura 1). El número de categorías de presas no difirió entre sexos, ya que se observa sobrelapamiento entre los intervalos de confianza al 84 % de las curvas de acumulación (Figura 2). En los machos se encontraron ocho categorías de presa que no se encontraron en las hembras: Hemiptera, Dermaptera, Diptera (adultos y larvas), Lepidoptera (larvas), Opilionida, Homoptera y Decapoda. Sin embargo, ninguna de estas estuvo entre las cinco presas más importantes en machos, por lo que se sugiere que su presencia se debe al mayor tamaño de muestra en estos. Las presas del orden Neuroptera (adultos y larvas) fueron encontradas únicamente en los estómagos de las hembras. Sin embargo, no se encontraron diferencias significativas en la composición de la dieta entre sexos, considerando los datos de abundancia (R = -0.01; P = 0.52) y volumen (R = 0.02; P = 0.16).
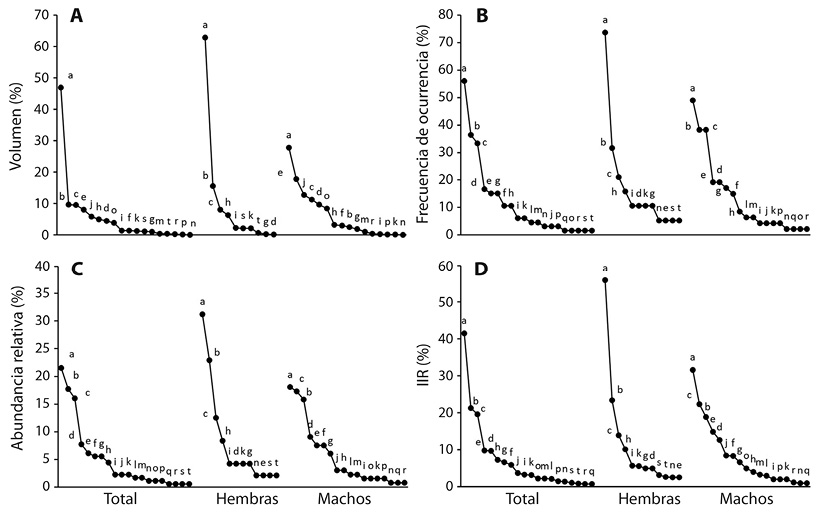
Figura 1 Rango-abundancia con el porcentaje de volumen (A), de frecuencia de ocurrencia (B), de abundancia (C), e índice de importancia relativa (D), del total de la muestra (T) y de cada sexo (H = hembras, M = machos). a = Orthoptera, b = Araneae, c = Coleoptera, d = Diplopoda, e = Chilopoda, f = Dermaptera, g = Hymenoptera (Formicidae), h = Isopoda, i = Coleoptera (larva), j = Hemiptera, k = Hymenoptera (no Formicidae), l = Diptera, m =Lepidoptera (larva), n = Acari, o = Diptera (larva), p = Opilion, q = Decapoda, r = Homoptera, s = Neuroptera y t = Neuroptera (larva).
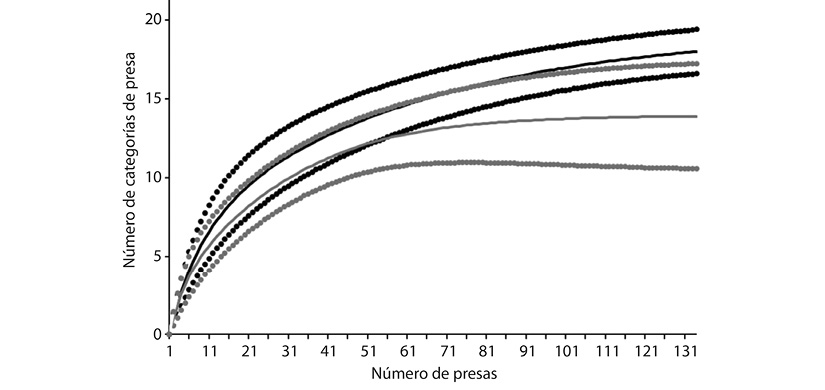
Figura 2 Curvas de acumulación que muestran el número de categorías de presa esperado e intervalos de confianza al 84 % para machos (líneas continua y punteadas de color negro) y hembras (líneas continua y punteadas de color gris).
El volumen promedio de las presas fue de 31.6 mm3 (ee = 62.7 mm3, intervalo = 0.1 - 365 mm3), el volumen de las presas encontradas en hembras osciló entre 0.1 y 365 mm3 ( ± ee = 53.54 ± 90.2 mm3) y en machos éste varió entre 0.1 y 226 mm3 ( ± ee = 22 ± 43 mm3). Se encontró una relación positiva entre la talla (LHC) de los individuos y el volumen de las presas (coef ± ee = 0.32 ± 0.06; c2 = 18.2, g.l. = 4; P < 0.001) y se observaron diferencias entre sexos. En los estómagos de las hembras las presas encontradas fueron de mayor volumen (c2 = 9.5, g.l. = 4; P = 0.002). Pero no se observó un efecto de la interacción entre la talla y el sexo de las ranas (c2 = 4.67; gl = 6; P = 0.1; Figura 3A).
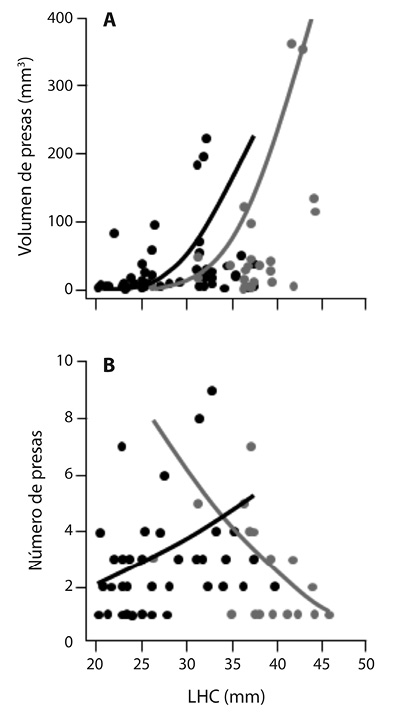
Figura 3 Relación entre la LHC de individuos adultos de Craugastor rhodopis y A) volumen de cada presa consumida, y B) número de presas consumidas. Puntos y líneas de tendencia, gris representa a las hembras y negro a los machos.
El número de presas consumidas por los ejemplares de C. rhodopis varió entre uno y nueve ( ± ee = 2.8 ± 1.7), en hembras osciló entre uno y siete ( ± ee = 2.5 ± 1.7) y en machos entre uno y nueve ( ± ee = 2.8 ± 1.7). No se encontró una relación entre la talla de las ranas y el número de presas consumidas (c2 = 8.05E-5; g.l. = 64; P = 0.98), ni diferencias entre machos y hembras (c2 =0.21; gl = 63; P = 0.46), pero sí se encontró un efecto de la interacción del sexo y la talla (c2 = 4; g.l. = 62; P = 0.01). En hembras se observó una asociación negativa entre estas variables (coef ± ee = -0.07 ± 0.02), siendo las hembras grandes las que consumen menor número de presas, mientras que en los machos la respuesta fue inversa (coef ± ee = 0.1 ± 0.02) siendo los machos grandes los que consumen más presas que los machos pequeños (Figura 3B).
Finalmente, el número de categorías de presas consumidas por C. rhodopis osciló entre uno y siete ( ± ee = 2.3 ± 1.3), los machos tuvieron entre uno y siete categorías ( ± ee = 2.5±1.4) y las hembras entre uno y cinco categorías ( ± ee = 2 ± 1.2). No se encontró un efecto de la talla de los individuos sobre el número de categorías de presas (c2 = 0.9; g.l. = 64; P = 0.6), tampoco diferencias entre sexos (c2 = 0.48; g.l. = 63; P = 0.22), ni respuesta significativa de la interacción de la talla con el sexo (c2 = 1.9; g.l. = 62; P = 0.1).
Discusión
Los resultados de este estudio indican que la dieta de C. rhodopis en el centro de Veracruz está compuesta por un amplio número de categorías de presas, dentro de las cuales predominan los ortópteros. Esto sugiere que esta especie emplea principalmente una estrategia de forrajeo de acecho. Debido a su amplia composición dietaria, al bajo porcentaje de individuos sin alimento y su alta abundancia en los fragmentos del bosque de niebla, C. rhodopis podría estar desempeñando un papel importante en la dinámica de las comunidades de artrópodos. Debido a que la dieta de los machos estuvo compuesta por presas más pequeñas que en las hembras, los individuos de cada sexo podrían estar impactando de manera diferencial en las poblaciones de artrópodos.
Un amplio número de categorías de presas ha sido reportado en otras especies del mismo género (e.g. Craugastor lineatus en Chiapas, México; Martínez-Coronel & Pérez-Gutiérrez, 2011) y de otros géneros de craugastóridos (e.g. Pristimantis labiosus; Gutiérrez-Cárdenas, Castillo, Martínez, Rocha, & Rojas-Rivera, 2016). Adicionalmente, la predominancia de ortópteros en la dieta de C. rhodopis, parece ser un fenómeno común en los craugastóridos, pues en varios de ellos se encuentran entre las tres principales presas consumidas (Parmelee, 1999; Martínez-Coronel & Pérez-Gutiérrez, 2011). Esto puede deberse a que los ortópteros son artrópodos móviles (Toft, 1980), presentan un cuerpo blando, con alto contenido energético y proteico (Anderson & Smith, 1998). Además, son de mayor tamaño en comparación a otros artrópodos, por lo que tienen una menor proporción superficie-volumen, y por lo tanto, una menor cantidad relativa de quitina (Hirai, 2002).
Aunque la determinación del modo de forrajeo de C. rhodopis no estuvo dentro de los objetivos de este estudio, pues para esto sería necesario cuantificar la oferta de alimento, ciertas características morfológicas y de la dieta sugieren que esta especie emplea principalmente un modo de forrajeo de acecho, al igual que otras especies de craugastóridos (Toft, 1981; Parmelee, 1999). Dentro de sus características morfológicas se encuentran un cuerpo robusto con un hocico ancho y una coloración críptica, mientras que las relacionadas a la dieta incluyen un elevado número de categorías de presa y una predominancia de presas grandes de gran movilidad y de hábitos solitarios (e.g. ortópteros y coleópteros; Toft, 1981). Parmelee (1999) sugirió que un alto porcentaje de estómagos vacíos también está relacionado con una estrategia de acecho; sin embargo, C. rhodopis en el centro de Veracruz parece no ajustarse a esta predicción, pues presentó un bajo porcentaje de estómagos vacíos en comparación con otras especies de craugastóridos de bosques de niebla de México y selvas lluviosas premontanas de Colombia (Martínez-Coronel & Pérez-Gutiérrez, 2011; Gutiérrez-Cárdenas et al., 2016). Alternativamente, es probable que esto se deba a que los machos no estaban involucrados en actividades reproductivas al momento de encontrarlos, o a una gran abundancia de presas en la época (nuestro estudio se llevó a cabo durante la época de lluvias) y sitios muestreados, tal como lo sugieren Biavati et al. (2004).
La composición de la dieta en C. rhodopis resultó ser similar entre sexos, al igual que en muchas otras especies de distintas familias de anuros (Biavati et al. 2004; Crnobrnja-Isailović et al., 2012; Rodrigues & dos Santos-Costa, 2014). Sin embargo, se observaron algunas presas exclusivas en machos. Es probable que esto se deba a la marcada inequidad del tamaño de muestra entre sexos. A pesar de que una dieta similar puede sugerir la presencia de competencia intraespecífica por los recursos tróficos (Crnobrnja-Isailović et al., 2012), es probable que en C. rhodopis sea mínima, dadas las diferencias intersexuales en el tamaño de las presas; además del bajo número de individuos sin comida. El efecto de la talla de las ranas en el tamaño de las presas, en el que ranas más grandes consumieron presas de mayor tamaño, en comparación con las más pequeñas, podría ser resultado de una estrategia para cubrir la inversión energética designada a la reproducción, mayor en hembras y machos de mayor talla, como se ha reportado para otras especies (Chen, Tang, Fan, Wang, & Pike, 2013; Finkler, Hayes, & Rifai, 2014). El efecto significativo de la interacción del sexo y el tamaño de las ranas sobre el número de presas consumidas, sugiere que la competencia intraespecífica entre adultos podría ser mayor entre machos grandes y hembras pequeñas, ya que son de tamaño similar y consumieron mayor número de presas que las hembras grandes y los machos pequeños.
Este trabajo sienta las bases para estudios posteriores relacionados con otros aspectos de la ecología trófica de esta especie, como: 1) el efecto de la ontogenia, que es un factor determinante en la composición de la dieta de varios craugástoridos (Whitfield & Donnelly, 2006), y 2) la composición de la dieta en función de la disponibilidad de las presas, con el fin de determinar la estrategia de forrajeo de la especie, y el grado de oportunismo y selección que tiene sobre los distintos tipos de presa. Asimismo, sería de gran relevancia 1) evaluar el grado de competencia intraespecífica que existe entre sexos y clases de edad, y 2) el efecto que tiene esta especie sobre las poblaciones de invertebrados que habitan la hojarasca, debido al número de categorías de presa que forman parte de su dieta.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de cualquier tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. El documento firmado se encuentra en los archivos de la revista

