











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkLos bosques naturales y páramos andinos son considerados ecosistemas naturales de alta diversidad florística a nivel mundial y regional. En estos ambientes, los distintos estudios que han evaluado la diversidad taxonómica y funcional se han enfocado en evaluar biotipos de vegetación leñosa ≥ a 5 cm de diámetro (Báez et al., 2015; Homeier, Breckle, Günter, Rollenbeck, & Leuschner, 2010; Jadán et al., 2017). Sobre las existencias y dinámica de la regeneración natural en los dos ecosistemas el conocimiento es escaso, especialmente en la vegetación arbórea perenne ≤ 5 cm de diámetro. No se ha considerado que estas especies luego de un proceso de germinación, sobrevivencia y crecimiento natural perenne, marcarán el futuro florístico, estructural y funcional del ecosistema (Gardner, Barlow, Parry, & Peres, 2007; Pérez-Ramos, 2007). A la regeneración natural no se le ha considerado como la base fundamental para la renovación y continuidad de las especies, por tanto es uno de los procesos más importantes en el ciclo de vida de las plantas (Nathan & Muller, 2000; Wang & Smith, 2002).
En el paisaje andino del sur de Ecuador existen superficies indeterminadas de plantaciones forestales de Pinus patula (pino), implantadas hace algunas décadas para producir bienes y brindar servicios ecosistémicos (Pacheco, Díaz, Quizhpe, Asanza, & Jadán, 2017; Quichimbo et al., 2017). De estas plantaciones, varias están abandonadas, incluso desde su implantación y a las que no se les ha aplicado labores silviculturales, lo que ha implicado la existencia actual de superficies considerables de plantaciones abandonadas.
Del mismo modo, se reportan impactos negativos que han causado rupturas ecológicas inter-específicas y ha provocado la eliminación o desplazamiento de especies nativas (Bueno & Baruch, 2011; Calviño-Cancela, Rubido-Bará, & van Etten, 2012). Contrariamente, en otros contextos tropicales, se afirma que las plantaciones forestales implementadas con densidades adecuadas y especies bajas en alelopatía facilitan las condiciones del microhábitat lo que permite el crecimiento de la regeneración natural de leñosas perennes (Carnus et al., 2006; Feyera, Beck, & Lüttge, 2002; Hartley, 2002). La estructura arbórea tanto vertical como horizontal de las plantaciones ha permitido el desarrollo de especies pertenecientes a diferentes gremios ecológicos que contribuyen en mediano y largo plazo a la funcionalidad del ecosistema (Aguirre & Weber, 2007).
Investigaciones ecológicas han revelado que la abundancia de la regeneración natural arbórea es similar en plantaciones de diferentes especies exóticas (Lee, Hau, & Corlett, 2005), pero la riqueza fluctúa en mayor o menor número, respecto a la existente en bosques naturales cercanos (Barreto, Prieto, Duarte de Moraes, & Pena, 2011). Se afirma, que la cobertura del dosel arbóreo y entrada de luz facilita la germinación de semillas y establecimiento de especies leñosas nativas, en plantaciones mono específicas (Yirdaw & Luukkanen, 2004), y también en ecosistemas naturales (Harrington & Ewel, 1997; Palomeque et al., 2017) para consecuentemente, incrementar la diversidad florística en la regeneración natural.
En ecosistemas de vegetación sometidos a restauración ecológica pasiva a través de la regeneración natural, la riqueza de especies aumenta su número conforme disminuye la distancia hacia los bosques naturales (Günter, Weber, Erreis, & Aguirre, 2007). Esto permite inferir que la diversidad florística de la regeneración natural está asociada con la capacidad y mecanismos de dispersión de semillas que tienen las especies leñosas nativas (Cramer, Hobbs, & Standish, 2008). Consecuente se afirma que el crecimiento inicial de plántulas está condicionado por las propiedades químicas y físicas del suelo (Günter et al., 2009).
La afirmación sobre la eliminación de especies vegetales nativas consecuencia de la implementación de plantaciones forestales no puede ser generalizada, sino corroborada responsablemente mediante estudios locales que permitan conocer el impacto real. Estas indagaciones permitirán evaluar y monitorear sinergias e interacciones ecológicas dentro y fuera de las plantaciones forestales. Bajo estos antecedentes se desarrolló la presente investigación, considerando las siguientes preguntas: 1) ¿La diversidad, abundancia de la vegetación y composición florística difiere en ecosistemas naturales frente a la existente en plantaciones de pino? y; 2) ¿Cuáles variables predictoras de fisiografía, cobertura, estructura de la vegetación arbórea, edáficas y distancia horizontal hacia bosques naturales explican mayormente la variación en la riqueza, diversidad y abundancia de la regeneración natural arbórea en ecosistemas naturales y plantaciones forestales de pino?
Materiales y métodos
Área de estudio: La investigación se realizó en dos escenarios de estudio: 1) ecosistemas naturales que corresponden a parches o fragmentos de bosques naturales y páramos; 2) plantaciones forestales de pino P. patula. Se consideró a los páramos, ya que dentro de ellos se establecieron plantaciones forestales hace más de 30 años; actualmente se hace pertinente evaluar y determinar cuál es la capacidad de las plantaciones de pino para promover la regeneración natural al ser comparadas con el bosque nativo.
Los ecosistemas naturales están distribuidos en un gradiente altitudinal que va desde los 3 000 hasta los 3 870 m de altitud; aquí se instalaron 27 parcelas de muestreo. Las plantaciones forestales desde 2 500 hasta 3 800 m de altitud, en donde se instalaron 45 parcelas; estas parcelas fueron distribuidas en todo el gradiente con al menos 10 parcelas cada 500 m de diferencia altitudinal y a 500 m de distancia horizontal, de manera aleatoria. Estos escenarios están ubicados en el sur del Ecuador, provincia del Azuay; fueron identificados y ubicados luego de la generación de un mapa de cobertura vegetal a partir de orto-fotos del 2012. Ecológicamente están dentro de las zonas de vida de Bosques Húmedo Montano y Páramo (Holdridge, 1967).
Muestreo y toma de datos: En los parches de bosques naturales y páramos se instalaron las parcelas de 500 m2. En las plantaciones de pino parcelas 1 000 m2 cuya superficie fue superior a los ecosistemas naturales, para equilibrar la densidad arbórea representada por el número de individuos (N), que es superior en los bosques naturales (1 200 N/ha), frente a las plantaciones de pino (830 N/ha, bajo densidad en plantación de 3 x 4 m). En el interior de estas parcelas se ubicaron cinco sub-parcelas de 2 × 2 m (una concéntrica y cuatro al azar) obteniendo un área efectiva de muestreo de 20 m2; para ello se adoptó metodologías de investigaciones realizadas sobre la regeneración natural en bosques tropicales (Escobar et al., 1982; Muñoz & Muñoz, 2010). Aquí, todas las plantas arbóreas leñosas ≤ 5 cm de dap tomado sobre el nivel del suelo, fueron identificadas taxonómicamente in situ a nivel de especie; a las que no se la pudo identificar, fueron recolectadas y llevadas al herbario LOJA. Estas especies fueron diferenciadas en árboles según Jørgensen (1999); se las consideró como motivo del estudio por ser biotipos perennes que presentan mayor dificultad para germinar y desarrollarse frente a las plantas herbáceas. Además, estas especies luego de un proceso de crecimiento perenne forman parte de la estructura y funcionalidad del ecosistema. También se contabilizaron los individuos de cada especie.
En cada sub-parcela se midió indirectamente el Índice de Área Foliar (IAF) utilizando el equipo óptico CI-110 (Plant Canopy Imager, CID Bio-Science, Washington, EE. UU) a través de imágenes hemisféricas digitales (150 campos de visión) del dosel como una medida de la cubierta vegetal total por área de terreno. Aquí las imágenes fueron tomadas a 100 cm sobre el nivel del suelo para medir IAF de árboles y arbustos, excluyendo sus tallos (Paiva, Silva, & Haridasan, 2015). Se consideró condiciones de clima bajo cielo abierto (pocas nubes) pero con el lente sombreado para evitar la sub-estimación del IAF, la sobre-estimación de radiación del haz solar y la radiación difusa. Las medidas del IAF en las cinco sub-parcelas fueron promediadas para la parcela.
En cada parcela de 500 y 1 000 m2, se tomaron datos ecológicos de altitud y ubicación geográfica (Apéndice digital). También se tomó una muestra homogenizada de suelos proveniente de cuatro sub-parcelas para realizar análisis físicos y químicos de: densidad aparente, pH, Materia orgánica (MO), Nitrógeno (N), Fosforo (P), Potasio (K), Calcio (Ca), Magnesio (Mg), Hierro (Fe) y Manganeso (Mn). Estas muestras fueron enviadas al laboratorio de suelos de la Agencia Ecuatoriana de Aseguramiento de Calidad del Agro - Agro-calidad, Ecuador.
Análisis de la información - Diversidad y Abundancia: La diversidad fue analizada mediante las diferencias métricas en la riqueza de especies y el índice alfa de Shannon, entre los dos escenarios de estudio, mediante curvas de rarefacción y extrapolación de especies. Para ello se utilizó el método de distribución de probabilidad, con datos por parcela (área efectiva de muestreo - 20 m2) (Colwell & Elsensohn, 2014). La abundancia de la regeneración natural arbórea fue determinada con base al número de individuos (Ind) por superficie efectiva de muestreo (20 m2) y por especies. Adicionalmente se determinó el índice de equidad mediante la relación entre el índice de Shannon y el logaritmo del número de especies, para medir la distribución homogénea o heterogénea del número de individuos sobre el número de especies (escala de 0 - 1). Los promedios de estos parámetros fueron comparados entre los dos escenarios de estudio mediante la prueba t (P < 0.05).
Composición florística: La variación en este parámetro entre ecosistemas naturales y plantaciones de pino, fue determinada mediante análisis de similitudes ANOSIM y escalamiento multidimensional no métrico - NMS (Bray Curtis, P < 0.05). Este último permitió evidenciar diferentes grupos de vegetación bajo un ordenamiento espacial. Para estos análisis se utilizaron la composición de especies y sus abundancias registradas en las parcelas de muestreo.
Asociado al NMS se realizó un análisis de especies indicadoras para identificar cuáles de estas son características de los diferentes grupos de vegetación articuladas con sus características ambientales, con base a su relación significativa (P < 0.05) entre frecuencia y abundancia. Los análisis fueron realizados con el programa estadístico Queco (Di Rienzo et al., 2010).
Relación entre variables predictoras y parámetros de la vegetación: Se describió estadísticamente la variación en los parámetros de la vegetación: riqueza, diversidad alfa de Shannon y abundancia en el gradiente altitudinal. Luego se describieron las variables predictoras diferenciadas en: 1) cobertura del dosel: Índice de Área Foliar (IAF); 2) edáficas: físicas y químicas del suelo; 3) estructura arbórea: densidad o número de individuos arbóreos por hectárea (N/ha) y área basal (m2/ha) y 4) topográficas: altitud y pendiente. El área basal fue calculada a los biotipos leñosos adultos ≥ 5 cm de dap utilizando las ecuaciones propuestas en Prodan, Peters, Cox y Real (1997).
Posteriormente, se realizó un análisis de explicación a priori de partición de la varianza (Varpart), bajo procedimientos validados en bosques tropicales por Chain-Guadarrama, Finegan, Vilchez y Casanoves (2012) y Jadán et al. (2017). Este análisis permitió explicar la importancia relativa de las distintas variables predictoras sobre la variación en los parámetros de riqueza, diversidad y abundancia de la regeneración natural, comparada para los dos contextos de estudio. Previo este análisis se aplicó el procedimiento Fordware selection que permitió seleccionar las variables explicativas o predictoras (P < 0.05) sobre los parámetros de la regeneración natural. Se utilizó el programa estadístico Queco (Di Rienzo et al., 2010). En las plantaciones de pino se incorporó la distancia horizontal como variable predictora, que fue medida desde las parcelas hasta los remanentes de bosques naturales. Para ello se utilizaron orto-fotos a escala 1:30 000 y herramientas informáticas de gvSIG; cuando se verificó más de dos remanentes circundantes a las parcelas se calculó el promedio entre las diferentes distancias.
Finalmente, como un análisis a posteriori, se realizaron regresiones lineales entre las variables predictores (X) que mayormente explicaron los parámetros de la vegetación (Y) para conocer su relación positiva o negativa de interdependencia. Se utilizó el programa estadístico Infostat (Di Rienzo et al., 2017).
Resultados
Diversidad y abundancia: Las curvas de acumulación mostraron valores estimados similares (según los desvíos estándar; α < 0.05) para riqueza de especies e índice de Shannon, entre los ecosistemas naturales y plantaciones de pino en seis parcelas, con tendencia al incremento (Figura 1A, Figura 1B).
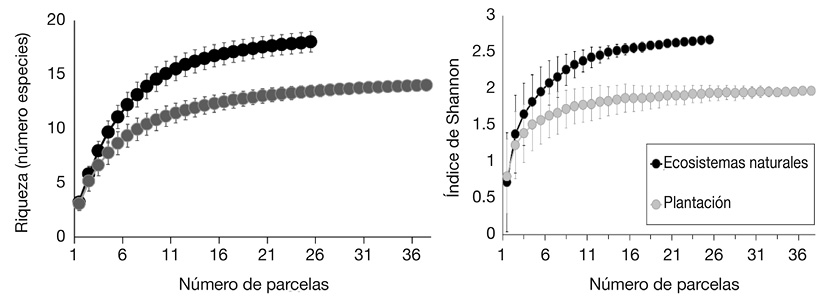
Figura 1 Curvas de acumulación de especies (± Desviación Estándar) (A), índice de Shannon (B) en relación al área muestreada - número de parcelas, en ecosistemas naturales y plantaciones de pino, provincia del Azuay - Ecuador.
No obstante, la riqueza de especies e índice de Shannon fueron superiores (α < 0.05) en los ecosistemas naturales cuando se incrementaron las parcelas y su tendencia llegó a estabilizarse. Los promedios de abundancia en la regeneración natural fueron similares estadísticamente entre los dos contextos de estudio (t; P = 0.5932).
La equidad no mostró diferencias estadísticas (t; P = 0.4629), pero sus valores fueron bajos (en escala de 0 a 1) tanto para los ecosistemas naturales y plantaciones (0.7, 0.68, respectivamente). Por lo tanto, se demostró una distribución heterogénea de individuos sobre las especies. Así, Miconiasp., registró 91 individuos en los ecosistemas naturales frente Viburnum triphyllum, Morella interrupta, Myrsine andina, Prumnopitys montana y Lomatia hirsuta que solo registraron un individuo (Cuadro 1). Lo mismo sucede en las plantaciones de pino; Miconia sp., registró 328 individuos frente a Myrciantes discolor que solo registró dos individuos.
Cuadro 1: Familias botánicas, especies y sus abundancias de la regeneración natural arbórea, registrada en los ecosistemas naturales y plantaciones de pino, provincia del Azuay, Ecuador. Table 1: Botanical families, species and abundances of natural tree regeneration, registered in natural ecosystems and pine plantations, Azuay province, Ecuador
| Familia | Nombre científico | Ecosistemas naturales | Plantaciones de pino |
| ARALIACEAE | Oreopanax rosei** | 109 | |
| ARALIACEAE | Oreopanax avicennifolia* | 18 | |
| ARALIACEAE | Oreopanax andreanu* | 8 | |
| ASTERACEAE | Gynoxys buxifolia** | 2 | 54 |
| CAPRIFOLIACEAE | Viburnum pichinchense | 1 | 43 |
| CAPRIFOLIACEAE | Viburnum triphyllum* | 5 | |
| CELASTRACEAE | Maytenus andicola* | 35 | 9 |
| CHLORANTHACEAE | Hedyosmum cumbalensis* | 21 | |
| CLETHRACEAE | Clethra ferruginea | 2 | |
| CLETHRACEAE | Clethra fimbriata | 8 | |
| CLETHRACEAE | Clethra sp. | 30 | 15 |
| CUNONIACEAE | Weinmannia fagaroides* | 16 | |
| CUNONIACEAE | Weinmannia rollottii | 3 | |
| ELAEOCARPACEAE | Vallea stipularis* | 78 | 29 |
| FABACEAE | Acacia dealbata | 32 | |
| MELASTOMATACEAE | Miconia sp. | 91 | 328 |
| MELASTOMATACEAE | Miconia theaezans | 9 | |
| MYRICACEAE | Morella interrupta | 1 | 16 |
| MYRSINACEAE | Geissanthus vanderwerffii** | 40 | |
| MYRSINACEAE | Myrsine dependens* | 27 | 8 |
| MYRSINACEAE | Myrsine andina | 1 | |
| MYRTACEAE | Myrcianthes discolor | 2 | |
| MYRTACEAE | Myrcianthes orthostemon | 11 | |
| PODOCARPACEAE | Podocarpus sprucei** | 52 | |
| PODOCARPACEAE | Prumnopitys montana | 1 | |
| PROTEACEAE | Lomatia hirsuta | 1 | 22 |
| PROTEACEAE | Oreocallis grandiflora* | 10 | |
| ROSACEAE | Hesperomeles ferruginea | 3 | |
| ROSACEAE | Polylepis incana | 2 | |
| ROSACEAE | Prunus ovalis | 7 | |
| TOTAL | 386 | 759 |
Especies indicadoras de características ambientales en ecosistemas naturales* y en plantaciones de pino** (P < 0.05).
Variación en la composición florística: La composición de especies mostró diferencias estadísticas significativas (ANOSIM; P = 0.001) entre los dos escenarios de estudio. El NMS (Stress = 0.2) según la distribución de las parcelas, mostró similitud de especies de la regeneración natural entre los ecosistemas naturales y las plantaciones de pino, a lo largo del primer eje de ordenación, bajo un 33 % de explicación (Figura 2). No obstante, con similar explicación (32 %) en el segundo eje, se separaron positivamente la mayor parte de parcelas de los ecosistemas naturales y negativamente las pertenecientes a las plantaciones de pino, lo que permitió observar disimilitud en la composición de especies. A esta disimilitud se adiciona las especies indicadoras que fueron diferentes en los dos escenarios de estudio (Cuadro 1, Figura 2).
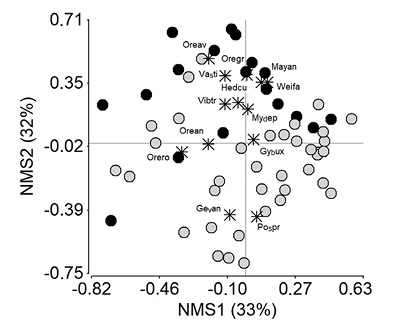
Figura 2 Escalamiento multidimensional no métrico para medición de similitud en la composición florística de la regeneración natural en ecosistemas naturales y plantaciones de pino, provincia del Azuay, Ecuador. Especies indicadoras ecosistemas naturales (círculos negros): Hedcu - Hedyosmun cumbalense, Mayan - Maytenus andicola, Mydep- Myrsine dependes, Oreav - Oreopanax avicennifolia, Oregr - Oreocallis grandiflora, Orean - Oreopanax andreanus, Vasti - Vallea stipularis, Vibtr - Viburnum triphyllum, Weifa - Weinmannia fagaroides. Plantaciones de pino (círculos gris claros): Pospr - Podocarpus sprucei, Orero - Oreopanax rosei, Gevan - Geissanthus vanderwerffii, Gybux - Gynoxis buxifolia.
Variación entre parámetros de la vegetación con variables predictoras: En los ecosistemas naturales la riqueza de especies, índice de diversidad de Shannon y abundancia presentaron coeficientes de variación ≥ al 90 % (Cuadro 2). En las plantaciones de pino, el coeficiente de variación mostró el mismo patrón que en los ecosistemas naturales, pero con menores valores mínimos ≥ al 69 % (Cuadro 2).
Cuadro 2: Variaciones en los parámetros de riqueza, índice de Shannon y abundancia de la regeneración natural arbórea registrada en ecosistemas naturales y plantaciones de pino, provincia del Azuay, Ecuador. Table 2: Variations in richness parameters, Shannon index and abundance of natural tree regeneration recorded in tree natural ecosystems and pine plantations, Azuay province, Ecuador
| Variable | Ecosistemas naturales | Plantación de pino | ||||||
| Media | CV | Mínimo | Máximo | Media | CV | Mínimo | Máximo | |
| Abundancia Rn (Ind/ha) | 14.5 | 106.2 | 0 | 51 | 16.9 | 117.2 | 0 | 97 |
| Riqueza (número de especies) | 3 | 95.2 | 0 | 9 | 2.5 | 69.2 | 0 | 6 |
| Shannon H | 0.7 | 99.9 | 0 | 2 | 0.7 | 83 | 0 | 1.6 |
CV: coeficiente de variación, Ind: individuos, Rn: regeneración natural.
Las variables predictoras dentro de los ecosistemas naturales registraron coeficientes de variación (CV) > 50 % (Cuadro 3); sin embargo, la altitud y densidad aparente registraron menores valores al 50 % en su CV. En las plantaciones de pino, los coeficientes de variación < 50 % se registraron para la mayor cantidad de variables predictoras (10 variables), en comparación con los ecosistemas naturales (Cuadro 3).
Cuadro 3: Variaciones en las variables predictoras de la regeneración natural arbórea en ecosistemas naturales y plantaciones de pino, provincia del Azuay, Ecuador. Table 3: Variations in the predictors variables of natural tree regeneration in natural ecosystems and pine plantations, Azuay province, Ecuador
| Variables predictoras | Ecosistemas naturales | Plantaciones de pino | ||||||||
| Media | CV | Mínimo | Máximo | Media | CV | Mínimo | Máximo | |||
| Cobertura arbórea | IAF | 27.2 | 76.5 | 0.8 | 59.2 | 2.7 | 42.9 | 0.2 | 5.5 | |
| Fisiográficas | Altitud (msnm) | 3 422.3 | 8.3 | 3 011 | 3 865 | 3 108 | 12.5 | 2 531 | 3 766 | |
| Pendiente (%) | 17.7 | 104.4 | 2 | 70 | 30.8 | 42.9 | 4 | 60 | ||
| Edáficas | Calcio (Ca) | 5.8 | 90.3 | 0.2 | 21.9 | 8.9 | 75.2 | 0.2 | 22.7 | |
| Densidad aparente (g/cm3) | 0.7 | 27.9 | 0.2 | 1 | 0.8 | 32.9 | 0.4 | 1.2 | ||
| Fosforo (P) | 10.3 | 93.4 | 3.5 | 41 | 3.5 | 0 | 3.5 | 3.5 | ||
| Hierro (Fe) | 939.1 | 82.8 | 15.4 | 3 113 | 750.9 | 48.5 | 194.2 | 1 761 | ||
| Magnesio (Mg) | 1.1 | 83.1 | 0.1 | 3.9 | 2 | 69.4 | 0.1 | 4.6 | ||
| Manganeso (Mn) | 28.7 | 93.5 | 2.7 | 110.2 | 21.7 | 57.7 | 3.9 | 55.8 | ||
| Materia Orgánica | 16.6 | 52.1 | 8.6 | 35.5 | 11.3 | 69.4 | 1.5 | 30.7 | ||
| Nitrógeno (N) | 0.8 | 52.1 | 0.4 | 1.8 | 0.6 | 69.2 | 0.1 | 1.5 | ||
| Potasio (K) | 0.4 | 97.3 | 0.1 | 1.9 | 0.6 | 92 | 0 | 2.3 | ||
| PH | 4.5 | 14.4 | 3.3 | 6.3 | 4.8 | 8.6 | 4.1 | 5.5 | ||
| Estructura arbórea | Densidad (N/ha) | 1 856.30 | 85.47 | 0 | 4 729 | 569.6 | 32.2 | 240 | 1 070 | |
| Área basal (m2/ha) | 19.8 | 96.65 | 0 | 60.1 | 30.9 | 45.2 | 2.7 | 62 | ||
CV: coeficiente de variación.
Partición de la variación: Este análisis demostró que ciertas variables predictoras como la estructura de la vegetación, fisiográficas y distancia hacia los bosques naturales en las plantaciones de pino, explicaron significativamente proporciones > al 60 % los parámetros de la regeneración natural arbórea (Cuadro 4). En los ecosistemas naturales la estructura arbórea (área basal y abundancia) y la altitud explicaron mayormente la variación en los parámetros evaluados (riqueza, diversidad y densidad) > 70 % (Cuadro 4). En las plantaciones de pino la distancia horizontal hacia los bosques naturales explicó con > 60 % la riqueza, diversidad y abundancia (Cuadro 4).
Cuadro 4: Valores de partición de la varianza (F; P < 0.05) de variables predictoras y su explicación sobre la riqueza, diversidad de especies y abundancia de la regeneración natural, en ecosistemas naturales (A) y plantaciones de pino (B), provincia del Azuay, Ecuador. Table 4: Partition values of the variance (F; P < 0.05) of predictors variables, and their explanation in the richness, diversity of species and abundance of natural regeneration, in natural ecosystems (A) and pine plantations (B), Azuay province, Ecuador
| Riqueza | Diversidad | Abundancia | |||||||
| A) Ecosistemas naturales | |||||||||
| Variables predictoras | % | F | P | % | F | P | % | F | P |
| Cobertura: IAF | 0.65 | 49.96 | 0.001 | 0.56 | 33.83 | 0.001 | 0.65 | 49.96 | 0.001 |
| Edáficas: Fosforo*, Calcio | 0.34 | 7.72 | 0.001 | 0.41* | 8.20 | 0.001 | 0.3 | 7.7 | 0.001 |
| Estructura arbórea: área basal y densidad* | 0.74 | 37.63 | 0.02 | 0.77* | 85.92 | 0.001 | 0.70* | 61.42 | 0.001 |
| Topográficas: altitud | 0.70 | 61.73 | 0.001 | 0.78 | 93.17 | 0.001 | 0.70 | 61.73 | 0.001 |
| Todas | 0.80 | 18.01 | 0.001 | 0.81 | 28.52 | 0.001 | 0.78 | 18.98 | 0.001 |
| Residuos | 0.20 | 0.19 | 0.22 | ||||||
| B) Plantaciones de pino | |||||||||
| Cobertura: IAF | 0.45 | 36.9 | 0.001 | 0.37 | 27.3 | 0.001 | |||
| Distancia horizontal | 0.6 | 67.2 | 0.001 | 0.88 | 324.7 | 0.001 | 0.6 | 67.2 | 0.001 |
| Estructura arbórea: área basal*; densidad | 0.33* | 22.5 | 0.001 | 0.49 | 21.9 | 0.001 | 0.33* | 22.5 | 0.001 |
| Topográficas: altitud | 0.33 | 22.9 | 0.001 | 0.52 | 48.3 | 0.001 | 0.33 | 22.9 | 0.001 |
| Todas | 0.65 | 18.8 | 0.001 | 0.9 | 66.6 | 0.001 | 0.6 | 22.5 | 0.001 |
| Residuos | 0.38 | 0.1 | 0,4 | ||||||
Variables predictoras vs parámetros de la vegetación: En los ecosistemas naturales, la riqueza se asoció significativa y positivamente con el área basal (P = 0.0116) y densidad arbórea (P < 0.0001) (Figura 3A). Esta variable registró valores de cero en las parcelas dentro de los páramos herbáceos ubicadas a mayor altitud (según los tamaños de los círculos, en donde el mayor tamaño representa mayor altitud). Contrariamente, se distribuyó con mayores valores en las parcelas donde el área basal oscila entre 10 y 25 m2/ha, y que están ubicadas en medianas y bajas altitudes. La riqueza de especies se distribuyó positivamente en las parcelas con mayor densidad de individuos arbóreos que están ubicados en menores altitudes (Figura 3B).
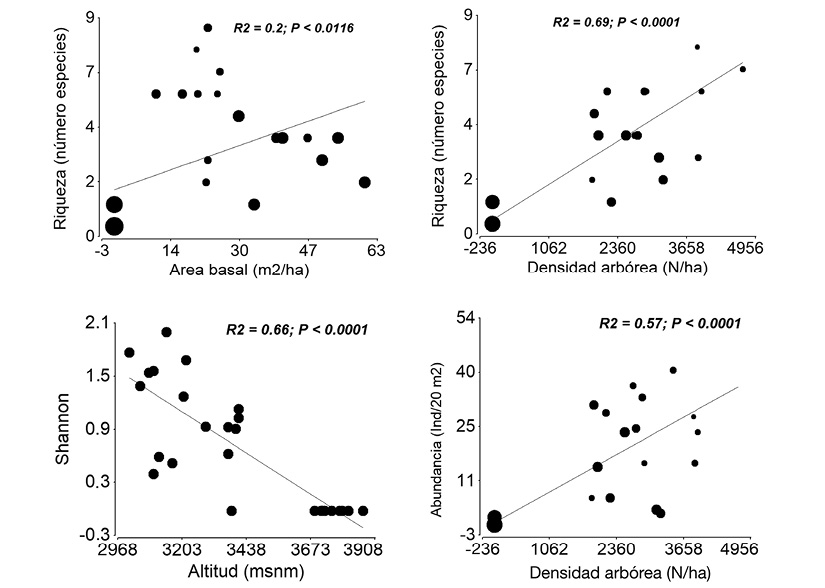
Figura 3 Relaciones entre riqueza, diversidad y abundancia con variables predictoras que explicaron con mayor porcentaje relativo su variación, en ecosistemas naturales, provincia del Azuay, Ecuador. Circulos de mayor tamaño representan parcelas ubicadas a mayor altitud.
El índice de Shannon se asoció negativamente con la altitud (P < 0.0001); aquí se registraron los mayores valores en las parcelas ubicadas en los sitios de menor altitud (Figura 3C). La densidad de la regeneración natural se asoció positivamente con la abundancia de la vegetación arbórea (P < 0.0001) (Figura 3D); los mayores valores se registraron en las parcelas con mayor número de individuos arbóreos, que están ubicados a menores altitudes.
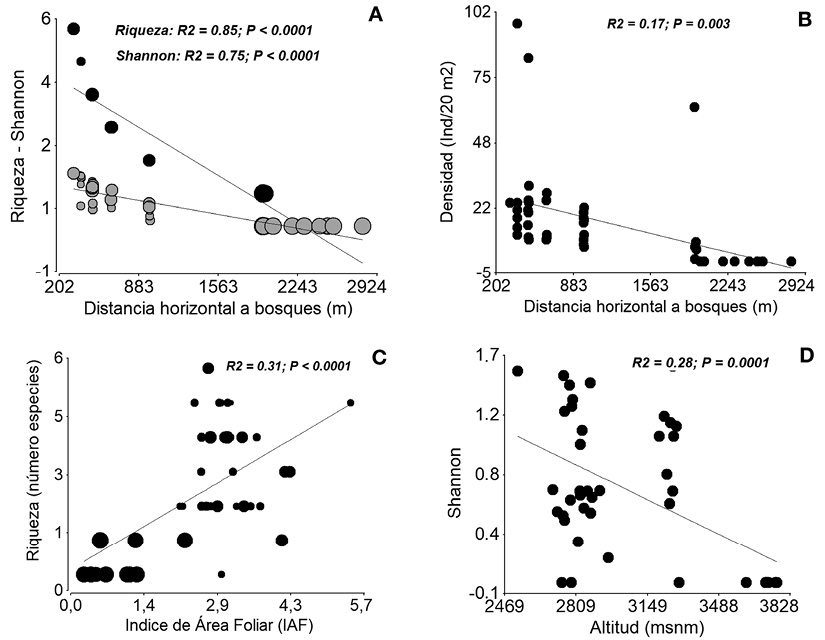
En las plantaciones de pino la riqueza de especies, índice de Shannon (Figura 4A) y abundancia de la regeneración natural (Figura 4B) se asociaron negativamente con la distancia horizontal hacia los bosques naturales (P < 0.05). El índice de área foliar que explicó relativamente en segundo lugar la riqueza de especies, se asoció positivamente (P < 0.05) (Figura 4C); los mayores valores de riqueza se registraron en las parcelas que tuvieron entre 1.6 y 4 del IAF. De igual manera, la altitud que explicó en segundo lugar la diversidad registró una asociación negativa (P < 0.05) (Figura 4D); aquí se registraron los menores valores en las parcelas ubicadas en mayores altitudes.
Discusión
Estudios sobre riqueza y abundancia de árboles en regeneración natural no han sido reportados para el área de estudio. En la presente investigación, los análisis de rarefacción con base a la intensidad de muestreo permitieron diferenciar mayor riqueza y diversidad florística de la regeneración natural arbórea en ecosistemas naturales. Estos mayores valores posiblemente estén asociados a la alta diversidad que poseen los bosques nativos andinos, lo cual ha sido registrado en árboles maduros (Barthlott, Mutke, Rafiqpoor, Kier, & Kreft, 2005). La diversidad florística de árboles maduros se enlaza con la producción de semillas y consecuentemente con el origen de la regeneración natural a nivel de hábitat y en zonas contiguas, según la intervención de agentes y mecanismos de dispersión. Resultados similares se han reportado por Barreto et al. (2011) y Lal, Singh y Bhardwaj (2015) quienes han registrado un mayor número de especies arbóreas de regeneración natural en bosques naturales, frente a la existente en plantaciones forestales en ecosistemas tropicales.
En las plantaciones de pino, en donde se registraron valores más bajos en riqueza y diversidad, la no disponibilidad de semillas de especies nativas dentro de la población de árboles de P. patula, se torna una fuerte limitante para dar origen a la regeneración natural. La presencia de árboles nativos y los procesos ecológicos de diseminación tanto anemófila y la realizada por animales silvestres resultan imprescindibles (Barreto et al., 2011; Telila, Hylander, & Nemomissa, 2015). La menor riqueza, también se debe posiblemente a factores de hábitat como: el porcentaje de luz, volumen de biomasa en el suelo y raíces que limitan los procesos de germinación y crecimiento inicial de las plántulas (Cavelier & Santos, 1999). Adicionalmente, la alta competencia intra-especifica entre individuos arbóreos de la misma especie posiblemente restringe la disponibilidad de macro y micro-nutrientes, especialmente Nitrógeno y Potasio, lo que incide negativamente en el crecimiento y desarrollo inicial de las plántulas (Vanclay, 2006).
Aunque la riqueza y diversidad de especies fue mayor en los ecosistemas naturales, la abundancia fue similar en los dos escenarios de estudio. Estos resultados son explicados por la baja equidad, demostrada con las altas variaciones de abundancia, por lo que se infiere una distribución heterogénea de individuos sobre las especies. Esto difiere con la vegetación arbórea (≥ 5 cm dap) de los bosques tropicales andinos, en donde se registró mayores valores más altos en la equidad con una distribución homogénea de individuos sobre las especies (Jadán et al., 2017).
La gráfica de escalamiento métrico multidimensional (NMS) mostró fuertes patrones de separación de composición de especies en el segundo eje de ordenación. Así se destaca disimilitud en la composición florística de la regeneración natural en los ecosistemas naturales frente a la registrada en las plantaciones de pino. Este resultado es validado mediante especies indicadoras, que fueron diferentes en los dos escenarios, cuya presencia permiten predecir la variación en la riqueza de especies (Lawton et al., 1998). Además, la variación o disimilitud florística de la regeneración natural puede especulativamente ser explicada por el bajo intercambio de especies (Chust et al., 2006); esto es demostrado en el presente estudio con la separación de la mayor parte de parcelas correspondientes a los diferentes escenarios de estudio.
La partición de la variación, muestra que la diversidad, riqueza y abundancia de la regeneración natural es influenciada principalmente por variables fisiográficas de altitud y estructurales de la vegetación arbórea como área basal y abundancia. Esta implicación es consistente con base a la relación positiva y significativa registrada en las regresiones lineales entre las variables predictoras frente a la riqueza y diversidad. La altitud ha sido un factor fisiográfico determinante en la variación de la riqueza y composición de especies, lo cual se ha registrado en varios estudios dentro de la región andina a nivel local y regional (Girardin et al., 2014; Homeier et al., 2010; Jadán et al, 2017). La relación positiva entre el área basal y la riqueza de especies, permite deducir que la mayor riqueza de especies se asocia con los sitios donde existen árboles de mayor tamaño.
La mayor explicación relativa de la distancia horizontal, sobre las variables evaluadas en las plantaciones de pino, se debe posiblemente a la disponibilidad de germoplasma o semillas en coberturas naturales circundantes (Cramer et al., 2008). Estos resultados se asocian con los postulados de la teoría de ensamblaje de dispersión, en donde la composición y abundancia de especies depende de la composición de las comunidades circundantes y de la capacidad de dispersión, asociado con la distancia geográfica, independientemente de las condiciones intrínsecas del hábitat (Rosindell, Hubbell, & Etienne, 2011). Estos postulados son consistentes con los resultados obtenidos, puesto que la menor riqueza de especies y su abundancia, se dan cuando se incrementa la distancia horizontal desde los bosques naturales hacia las plantaciones.
En las mismas plantaciones de pino, la explicación en segundo plano del índice de área foliar (IAF) sobre la riqueza de especies en la regeneración natural y su relación negativa se torna algo atípico con lo registrado por Palomeque et al. (2017), pero explicable al mismo tiempo. Así, los valores más bajos en riqueza se registraron en las parcelas ubicadas a mayor altitud y que poseen los menores valores del IAF. A estas parcelas circundan los páramos en donde los biotipos más importantes y dominantes son herbáceos. Además, hacia estas parcelas los remanentes de bosques naturales se encuentran a considerables distancias, por lo que la disponibilidad de semillas de biotipos arbóreos se torna una limitante para promover la riqueza de especies. Al mismo tiempo consolida a la distancia horizontal como la variable que explica la riqueza de especies, lo cual es consistente con lo registrado por Günter et al. (2007) quienes manifiestan que la riqueza florística en escenarios antropogénicos está asociada negativamente con la distancia hacia los bosques naturales.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de cualquier tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. El documento firmado se encuentra en los archivos de la revista.

