











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkThe conversion of land to coffee cultivation and the intensification of coffee production are among the major causes of changes in natural vegetation and other soil cover types in tropical areas (Perfecto & Vandermeer, 2002; Perfecto & Armbrecht, 2003; Guhl 2004; Jha et al., 2014). Crops such as coffee, occupy large areas at moderate elevations of the tropics, and their cultivation has led to a drastic reduction in biodiversity (Perfecto & Vandermeer, 2015). In Colombia, it is estimated that 70 % or more of the original extent of the Andean forest has been transformed (Etter & van Wyngaarden, 2000), largely as the result of the expansion of coffee cultivation. In 2010, Colombian coffee plantations accounted for approximately 3 000 000 hectares in 19 departments, along the slopes of the Andes between 1 000 and 2 000 meters above sea level (Federación Nacional de Cafeteros FNC, 2010), and 60 % of the land dedicated to the cultivation of this crop has experienced a reduction in shade cover and the application of chemical inputs such as pesticides and herbicides, among others (Guhl, 2004).
Although technology inputs yield greater production, the land is subjected to an accentuated process of intensification that results in soil degradation (Perfecto, Rice, Greenberg, & Van der Voort., 1996). Characteristics such as spatial heterogeneity or the degree of homogenization in agricultural landscapes, as well as management practices, can affect the diversity and distribution of local and regional biota, as well as the incidence of invasive species or pests (Armbrecht, Rivera, & Perfecto, 2005; Philpott et al., 2008). Because of technological advances, quantitative information is now available, and a compendium of images portrays the size, speed and extent of the patterns of deforestation; however, little is yet understood about the effect of fauna reduction on the functions and services of ecosystems (Dirzo et al., 2014).
Research on coffee ecosystems has received considerable attention in recent decades, particularly due to the effects that the intensification of coffee production has had on the associated biodiversity (Perfecto & Vandermeer, 2015). Studies of ants, in particular, have proved useful in analyzing diversity and conservation in these agro-ecosystems. For example, it has been documented that the diversity of ants may be reduced in monoculture coffee plantations under full sun conditions due to changes in light and leaf litter (Perfecto & Vandermeer, 2002; Armbrecht et al., 2005) as well as a reduction of nesting sites for several species (Philpott & Foster, 2005; Roberts et al., 2000). In this regard, the quality of the microhabitat’s structure is a determining factor for the establishment of many species of ants; this is the case for traditional crops of shade-grown coffee, which contain a greater diversity of ants because they ensure the minimal requirements for the survival of ant communities (Armbrecht & Perfecto, 2003). The negative impacts of full-sun plantations on guilds of predatory and omnivorous ground-dwelling ants has also been studied (García, Zabala, & Botero, 2008; Dias et al., 2013), emphasizing the role of this group of insects within the different habitats in predicting the ant communities’ response to disturbances (Andersen, 1997; Leal, Wirth, & Tabarelli, 2014).
Several studies in Colombia have associated the diversity of ant populations with the management of coffee plantations, especially in the area known as the coffee zone (Armbrecht et al., 2005; Rivera & Armbrecht, 2005; Gallego-Ropero, 2005,García et al., 2008; Zabala, Arango, & de Ulloa, 2013). Nevertheless, the country’s geographical location, its topography and different life stories provide great diversity in the structure and composition of communities, and the ecological context often affects the manner in which ants respond to changes in land use. This is especially true for Colombian coffee plantations, where rapid transformation has favored the establishment of plantations at higher altitudes and displaced the majority of production towards the southern part of the country (FNC, 2011). There are few studies that relate the diversity of ant populations to the management of coffee plantations in these areas (Urrutia-Escobar & Armbrecht, 2013). The present study describes the specific diversity of ground-dwelling ants and identifies functional groups in two contrasting coffee-cultivation systems (shaded and exposed) and a control system (forest patches) in the department of Cauca, in the southwestern Colombian Andes.
Materials and methods
Study site: The study was conducted in the municipalities of Popayán and Cajibío, located in the department of Cauca, in the Southwestern Colombian Andes, at an altitudinal strip between 1 450 and 1 800 meters above sea level; defined by Holdridge (Espinal, 1968) as premontane moist forest (bh-PM). The climate is temperate and humid, with a mean temperature of 27.9 °C; the region experiences a bimodal rainfall regime with a mean annual precipitation of 2 258 mm. The region is characterized by mixed coffee cultivation in small plots that has areas of exposed coffee monoculture and heterogeneous growth, including both exposed and shaded coffee plantations. Between May and September 2014, 20 research sites were chosen for the study, including eight shaded coffee plantations, eight unshaded coffeeplantations and four forest patches (Fig. 1).
Ant sampling: In general, the sampling protocol suggested by Alonso and Agosti (2000) was followed. Each site was demarcated by two transects 50 meters long and two meters wide, with 12 sampling points established at ten-meter intervals. An active search was conducted at each station for 20 minutes, followed by the placement of a pitfall trap with 95 % ethyl alcohol for 24 h. In addition, the entire layer of leaf litter from a 1-m2 area was collected and sifted using the mini-Winkler extraction technique for 48 h. A total of 36 samples were obtained from each site. The collected samples, preserved in 90 % alcohol, were initially separated by morphospecies. Subsequently, were organized at the lowest possible taxonomic level using Fernández (2003) generic keys and various specific keys according to genus. Comparisons were also made with material classified by specialists from a collection stored in the Museum of Entomology of the Universidad del Valle, Cali (MUSENUV, national registry of collections 077). The specimens, duly mounted on pins, labeled and classified, are preserved in the museum’s collection (Jiménez et al., 2016).
Habitat variables: In each study site, two separate sampling points at least 50 m from each other were chosen at random. At each point, a circle 24 m in diameter was drawn and bisected by two axes: one in a north-south orientation, the other in an east-west orientation. At points located every four meters along these axes, measurements were taken and analyzed for nine habitat variables distributed over three vertical levels (Armbrecht, 2003): 1) the arboreal level: canopy cover, tree richness, tree density, epiphytic richness, tree height and tree diameter; 2) coffee plant level: coffee plant density and mean coffee plant height; and 3) soil level: leaf litter depth.
Functional groups: The species were classified into functional groups based on the literature (Delabie, Agosti, & Nascimento, 2000; Silvestre, Brandão, & Da Silva, 2003; Silva & Brandão, 2010; García-Martínez et al., 2015). The classification of these groups allows a functional comparison of the ants to be made at the different intensity levels of the study sites (forest patches, exposed coffee plantations and shaded coffee plantations) using the Sørensen similarity index, adapted by Silvestre (2000), which includes the number of guilds at each sample point and the number of species present within the guilds. In cases where information is not reported or the specific epithet is not known, the classification is based on habits reported for the genus.
Data analysis: We used Hill’s numbers to analyze diversity, measured in units of effective number of species (Jost, 2006): 0D (species richness), 1D (exponential of the Shannon index) and 2D (inverse of the Simpson index). These measures comply with the principle of replication, are easy to interpret and are therefore recommended in comparative studies of diversity (Jost, 2010; Chao et al., 2014). The completeness of the sample was measured considering the sample coverage (Cs) at the management level and among different types of plantations and forest patches (sites). This measure makes it possible to compare the diversity of communities among samples of the same coverage, using extrapolation and interpolation of the coverage of their reference samples (Chao & Jost, 2012; Chao et al., 2014), with the subroutine R for data on frequency of capture, where 0 represents the minimum and 100 the maximum completeness (R Development Core Team, 2014). A visual comparison of the confidence intervals was performed to establish significant differences (no overlap) between land uses or management (Cumming Fidler, & Vaux, 2007), and analysis of variance and ANOVA post hoc comparisons were performed after confirming the normality of the data. The Jaccard similarity index was used to compare the composition of species between land use types. This index has values ranging from 0 (minimum similarity) to 100 (maximum similarity) and was represented using the un-weighted pair group method with an arithmetic mean (UPGMA). The statistical robustness of this result was based on the similarity profile test (SIMPROF), using the PRIMER software, version 6.1.16 (Clarke & Gorley, 2006). The relationship between the composition of species and sites, explained by environmental variables, was established through a canonical correlation analysis (CCA) and generalized linear models in accordance with the distribution of the data and the type of variable using the R-3.2.1 software (R Development Core Team, 2014).
Results
Overview: A total of 15 666 individuals from nine subfamilies were collected, distributed over 40 genera and 92 morphospecies (Table 1). Sixty percent of the individuals were identified to the species level. The remaining 40 % belonged to genera with difficult taxonomic identification such as Pheidole, Solenopsis, Hypoponera and Camponotus (Lyal & Weitzman, 2004). Myrmicinae accounted for the largest representation in the samples (44 species), followed by Dolichoderinae (six species) and Ponerinae (13 species). Pheidole was the genus with the largest number of species (14), followed by Solenopsis (8) and Strumigenys (7). The most abundant species (in individuals) were Solenopsis sp. 1 (2 871), Linepithema piliferum (2 231), Solenopsis sp. 2 (1 692) and Wasmannia auropunctata (1 617).
Table 1 List of the ant species found in each soil use. FOS: forests, SHC: coffee with shade and SUN: unshaded coffee
| Species | FOS | SUN | SHC | Total |
|---|---|---|---|---|
| Adelomyrmex tristani | 3 | - | - | 3 |
| Apterostigma gonoides | 4 | - | - | 4 |
| Atta cephalotes | 114 | 182 | 8 | 304 |
| Azteca velox rochai | - | - | 2 | 2 |
| Brachymyrmex heeri | 11 | 6 | 4 | 21 |
| Brachymyrmex longicornis | 20 | 79 | 118 | 217 |
| Camponotus sp. 2 | - | 2 | 22 | 24 |
| Camponotus sp. 4 | - | 1 | - | 1 |
| Camponotus sp. 7 | - | 1 | 8 | 9 |
| Camponotus sp. 9 | - | 1 | 1 | 2 |
| Cardiocondyla minutior | - | 33 | - | 33 |
| Cheliomyrmex andicola | - | 39 | 11 | 50 |
| Crematogaster crinosa | 12 | - | 17 | 29 |
| Crematogaster nigropilosa | 3 | - | 28 | 31 |
| Cyphomyrmex cornutus | 17 | 66 | - | 83 |
| Cyphomyrmex rimosus | 228 | 84 | 7 | 319 |
| Cyphomyrmex salvini | 11 | - | - | 11 |
| Discothyrea horni | 1 | - | 1 | 2 |
| Dorymyrmex biconis | - | 12 | - | 12 |
| Ectatomma ruidum | - | 84 | 17 | 99 |
| Gnamptogenys andina | - | - | 15 | 15 |
| Gnamptogenys bisulca | 55 | - | - | 55 |
| Gnamptogenys dichotoma | - | - | 1 | 1 |
| Gnamptogenys nigrivitrea | 22 | - | - | 22 |
| Heteroponera microps | 15 | 27 | 19 | 61 |
| Hypoponera sp. 1 | 8 | 7 | 11 | 26 |
| Hypoponera sp. 2 | - | 69 | 20 | 89 |
| Hypoponera sp. 3 | 5 | 46 | 22 | 73 |
| Hypoponera sp. 4 | 7 | 4 | 34 | 45 |
| Hypoponera sp. 5 | 1 | - | 2 | 3 |
| Labidus coecus | 13 | 1 | - | 14 |
| Labidus praedator | - | - | 31 | 31 |
| Leptogenys pucuna | 1 | - | - | 1 |
| Linepithema fuscum | - | 164 | 63 | 227 |
| Linepithema iniquium | 4 | 61 | 25 | 90 |
| Linepithema neotropicum | - | 75 | 171 | 246 |
| Linepithema piliferum | 95 | 1 075 | 1 061 | 2 231 |
| Mayaponera constricta | 2 | - | - | 2 |
| Myrmelachista zeledoni | 2 | 1 | - | 3 |
| Neivamyrmex punctaticeps | - | - | 27 | 27 |
| Neoponera aenescens | 9 | - | - | 9 |
| Neoponera verenae | 14 | - | 6 | 20 |
| Nesomyrmex asper | 1 | - | 45 | 46 |
| Nylanderia steinheili | 84 | - | - | 84 |
| Octostruma balzani | - | 2 | 7 | 9 |
| Octostruma stenoscapa | 3 | - | - | 3 |
| Species | FOS | SUN | SHC | Total |
| Odontomachus chelifer | 1 | 104 | 44 | 149 |
| Odontomachus eritrocephalus | - | - | 48 | 48 |
| Pachycondyla fuscoatra | 1 | - | - | 1 |
| Pheidole sp. 1 | 1 | 1 063 | 161 | 1 225 |
| Pheidole sp. 11 | 45 | - | 1 | 46 |
| Pheidole sp. 12 | 61 | - | - | 61 |
| Pheidole sp. 13 | 73 | 3 | 3 | 79 |
| Pheidole sp. 14 | 2 | - | - | 2 |
| Pheidole sp. 15 | 16 | 3 | 82 | 101 |
| Pheidole sp. 16 | 85 | - | - | 85 |
| Pheidole sp. 21 | 14 | - | 22 | 36 |
| Pheidole sp. 22 | 1 | - | - | 1 |
| Pheidole sp. 3 | 25 | - | - | 25 |
| Pheidole sp. 4 | 2 | 1 179 | 931 | 2 112 |
| Pheidole sp. 5 | - | 1 | 3 | 4 |
| Pheidole sp. 6 | - | 132 | 31 | 163 |
| Pheidole sp. 7 | 59 | - | 37 | 96 |
| Prionopelta amabilis | - | 1 | 20 | 21 |
| Pseudomyrmex ejectus | - | - | 1 | 1 |
| Pseudomyrmex gracilis gracilis | - | - | 1 | 1 |
| Pseudomyrmex sp. 3 | - | - | 1 | 1 |
| Pseudomyrmex sp. 4 | - | 1 | - | 1 |
| Pseudomyrmex sp. 5 | - | 5 | - | 5 |
| Pseudomyrmex sp. 7 | - | 1 | - | 1 |
| Rasopone ferruginea | 43 | 1 | 6 | 50 |
| Rhopalothrix sp. 1 | 5 | - | - | 5 |
| Rogeria belti | 5 | 1 | 1 | 7 |
| Solenopsis geminata | - | - | 38 | 38 |
| Solenopsis sp. 1 | 30 | 2 288 | 553 | 2 871 |
| Solenopsis sp. 2 | 768 | 535 | 389 | 1 692 |
| Solenopsis sp. 3 | - | 87 | 2 | 89 |
| Solenopsis sp. 4 | - | - | 2 | 2 |
| Solenopsis sp. 6 | 34 | 4 | 25 | 63 |
| Solenopsis sp. 7 | - | 90 | 5 | 95 |
| Solenopsis sp. 8 | - | - | 16 | 16 |
| Stigmatomma orizabanum | 3 | - | - | 3 |
| Strumigenys connectens | 3 | - | - | 3 |
| Strumigenys denticulata | 11 | 15 | - | 26 |
| Strumigenys louisianae | - | 14 | 1 | 15 |
| Strumigenys margaritae | - | - | 3 | 3 |
| Strumigenys nubila | 71 | - | 4 | 75 |
| Strumigenys raptans | 20 | - | - | 20 |
| Strumigenys skia | - | 16 | 2 | 18 |
| Typhlomyrmex pusillus | 2 | - | - | 2 |
| Wasmannia auropunctata | 35 | 147 | 1 435 | 1 617 |
| TOTAL | 2 181 | 7 767 | 5 672 | 15 666 |
The samples collected from the shaded coffee plantations (SHD) contained a greater number of species (61 species and 5 672 individuals), whereas those collected from the unshaded coffees (SUN) contained a greater number of individuals (7 813) but fewer species (48). In contrast, the forest patches (forests) had 56 species represented by only a few individuals (2 181); this coverage was also remarkable for containing the highest number of exclusive species (21), including Adelomyrmex tristani, Apterostigma goniodes, Cyphomyrmex salvini, Discothyrea horni, Gnamptogenys bisulca, Gnamptogenys nigrivitrea, Neoponera aenescens, Stigmatomma orizabanum, Strumigenys raptans and Typhlomyrmex pusillus.
Sample coverage and diversity: The sample coverage at the land use level exhibited a deficit of less than 5 % (range 95.8-97.7 %); therefore, to compare the diversity of species in their different orders (qD), the sample was interpolated to 95.8 % completeness with a confidence interval (CI) of 95 %. The total results for each type of land use reveal a zeroorder difference in diversity (species richness) between sites with canopy cover (forests and SHD) and sites exposed to direct sun (SUN), with very similar values for SHD and forests (Fig. 2). The other levels of diversity (1D and 2D) exhibited significantly higher diversity in the forests than in the two coffee plantations. The diversity of common species (1D) presented significant differences between shaded and exposed coffee plantations, but the diversity of dominant species (2D) exhibited no difference between SHD and SUN.
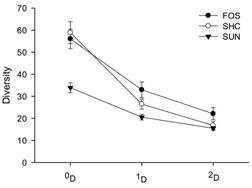
Fig. 2 Comparison of the three orders of diversity of soil ants in the three soil uses (FOS: forest patches, SHC: shade coffee and SUN: unshaded coffee). 0D: richness, 1D: typical diversity, 2D: Diversity of the most abundant species.
A comparison of the zero-order diversity among all the sites representing each land use (Fig. 3) reveals a higher mean value for the forest sites but no significant differences among the three types of land use (F = 1.81, P < 0.1931). The 1D diversity exhibited differences between management types (F = 4.58, P < 0.025), and the Tukey test indicated that the forest and SUN sites were significantly different (t = -3.25, P < 0.0196). The diversity of dominant species also exhibited variation among management systems (F = 3.7174, P < 0.025), and the Tukey test indicated that this difference occurred between forest and SUN sites (t = -2.578, P < 0.048). These results reveal a large variability within each land use type, and by some measures of diversity, the SHD sites resemble forests, while others resemble the SUN sites.
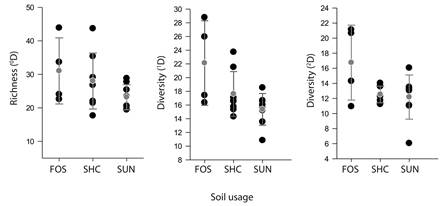
Fig. 3 Local diversity (α) of ants at sites grouped by soil use. The gray dots represent the mean and bars 95 % confidence intervals. FOS: forests, SHC: coffee with shade and SUN: unshaded coffee.
Composition of species among different land use types: The Whittaker curves present species distribution according to their abundance (frequency of capture) at sites with different land use types (Fig. 4). The curves with steeper slopes appear in the areas planted with coffee, especially in SUN, due to the presence of few species in great abundance. However, the forest sites also indicate the dominance of two species, Solenopsis sp. 2 and Cyphomyrmex rimosus, but the slope is more gradual towards the center of the curve, with a second group led by Atta cephalotes, which is distributed more equitably. Finally, the curve is tapering at those rare species with low frequency of capture. In the SHD sites, Wasmannia auropunctata was found to be the dominant species, whereas Solenopsis sp. 1 was dominant in the SUN sites. The abundance of L. piliferum and Pheidole sp. 4 is notable in both coffee plantation types.
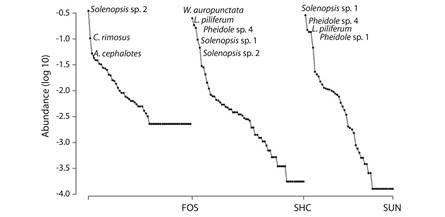
Fig. 4 Relative abundance curves for ant species in three contrasting soil usage. FOS: forests, SHC: coffee with shade and SUN: unshaded coffee.
A graphical and statistical comparison of the composition of ants among different sites (Fig. 5) reveals that at a low level of similarity (12.4 %), there is a significant separation (π = 18.64, P < 0.05) between the group of forested sites (FOR3, FOR1, FOR4, FOR2), together with three shaded coffee plantations (SHD3, SHD1, SHD2), and the remaining sites. At 23 % similarity, there is a significant separation between the forests and SHD3, SHD1 and SHD2 (π = 2.81, P < 0.05). At 33.6 % similarity, there is significant separation between the sun-grown coffee site 8 (SUN8) and the other SUN sites (π = 10.94, P < 0.05). At 53 % similarity, the sun-grown coffee site 4 (SUN4) is separated from the other SUN sites (π = 7.96, P < 0.05) and at 64.9 % similarity, it is apparent that the shaded coffee plantations constitute a significantly separate group from the exposed coffee plantations (π = 6.9, P < 0.05), but shaded coffee site 8 exhibits no differences from the exposed coffee plantations.
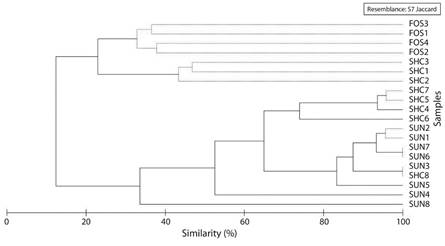
Fig. 5 Ant assemble associated to the different soil uses in the Cauca coffee area, based on Jaccard similarity index. Continuous lines show statistically different groups according to SIMPROF, while dotted lines the groups not statistically separated by SIMPROF. FOS: forests, SHC: coffee with shade and SUN: unshaded coffee.
Relationship between diversity and environmental variables: Environmental variables explain 81.34 % of the variability of species composition. Most of this variability was explained by axis 1 (28.6 %) and axis 2 (16.7 %). All of the unshaded coffee (SUN) and five of the eight shaded coffee plantations (SHD) exhibit high values for soil and air temperature variables and for coffee plant density. The other variables had higher values in the forest, especially relative humidity and the density of trees and epiphytes (Fig. 6).
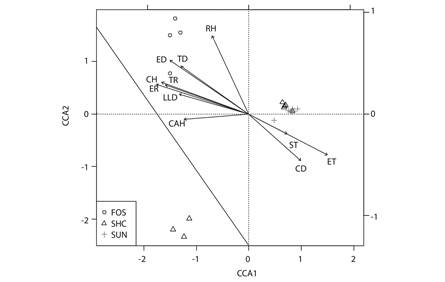
Fig. 6 Canonical correspondence analysis. CH: coffee height, CAH: canopy height, CC: Canopy cover, TD: tree density, CD: coffee density, ED: epiphyte density, LLD: leaf litter depth, RH: relative humidity, TR: tree richness, ER: epiphyte richness, ST: soil temperature ET: environmental temperature.
Functional groups: Ten functional groups of ants were found: nine in forests, ten in unshaded coffee plantations and nine in shaded coffee plantations. In general, the dominant guild of omnivorous ground-dwelling ants accounted for the greatest number of species (22), followed by the group of specialized predatory cryptic poneromorphs (15 species) and specialized predatory cryptic Myrmicinae (14 species); these were the most commonly found guilds in the three land use types. The first group consists predominantly of Pheidole and Solenopsis, which nest in the ground and are aggressive, wide-ranging foragers that are generalists in their eating habits. The second group includes the small poneromorphs, which, like the third guild, nest at ground level or in leaf litter. Their presence is thus associated with the amount of leaf litter, which explains why they are found in greater numbers in the forest. Guilds of Pseudomyrmecinae and minimal vegetation specialists appeared infrequently in the sample, because the method used was aimed at the soil’s myrmecofauna. Moreover, the nomadic habit of legionnaire ants makes it difficult to find them in the field; these three groups were therefore excluded from the analysis of functional similarity.
The functional similarity index as adapted for guilds by Sørensen reveals greater similarity between the shaded and exposed coffee plantations (71.58 %), as these land uses share 37 species in seven guilds, indicating a high degree of similarity in the structure of their ant communities, but less so in their composition. Therefore, ecological equivalence is considered present in several of these species. One of the guilds that best represents this is the guild of large predatory species, where species such as Neoponera verenae and Odontomachus erythrocephalus complement the predatory role in soils of shaded coffee plantations; in contrast, Ectatomma ruidum and Odontomachus chelifer are most abundant in soils of exposed coffee plantations. The other comparisons exhibit values approximately at or below 50 % for similarity among ant communities. Forests and shaded coffee plantations exhibit 54 % similarity; these two land use types share 29 species in seven guilds. Forests and unshaded coffee plantations exhibit a similarity index of 42.11 %, sharing 20 species in seven guilds.
Discussion
The use of complementary methods such as leaf litter extraction with Winkler bags, pitfall traps and manual collection, as suggested by Alonso and Agosti (2000), ensured a high sampling coverage (range 95.8 - 97.7 %) for a megadiverse family such as Formicidae. These methods have been used in several studies in Colombia, yielding very good results in heterogeneous landscapes (Jiménez, Lozano-Zambrano, & Álvarez-Saa, 2008; García, Zabala, & Botero, 2008).
The species richness of ants recorded for the area (92 morphospecies) is similar to that found by other authors in other coffee-growing areas of Andean Colombia. Including vegetation ants, Armbrecht et al. (2005), documented 115 morphospecies in Apía (Risaralda) and 65 morphospecies in Támesis (Antioquia), two locations of the central Andes. Using tuna bait, Gallego and Armbrecht (2005) reported 54 species of ants in coffee plantations in the former region. Applying a method similar to the one used in this study to three 2 500-ha sites, Sánchez-Clavijo, Durán, Vélez, García, and Botero (2008) reported 75 species in El Cairo (Valle del Cauca), 110 in Támesis (Antioquia) and 99 in Santander. Zabala et al. (2013) recorded 96 species of ants in the leaf litter of three representative samples of a coffee-growing landscape in the western Andes of Risaralda. Using Winkler sacs, Cepeda-Valencia, Pérez -Pedraza, L., and Fernández (2014) documented 96 morphospecies of ants associated with leaf litter in both forest patches and shaded coffee plantations in Cundinamarca. Based on this evidence, ant assemblages in coffee-growing areas of the Colombian Andes tend to fluctuate at approximately 100 species.
In the department of Cauca, the closest reports to the research site are those by Sinisterra Gallego-Ropero, and Armbrecht (2016), who identified a community of ants present in extrafloral nectaries of guamo trees (Inga edulis and I. densiflora) used for shade on coffee plantations and the myrmecofauna in the leaf litter on the ground; these authors reported 43 species of ants. In an area of Cauca located approximately 60 km North of the present research site, Urrutia-Escobar and Armbrecht (2013) reported 82 species of ants in the coffee farms of Caldono, Cauca, with the greatest species richness found in shaded coffee plantations (60), followed by forest patches (56) and finally the exposed coffee plantations (33 species). It is noteworthy that the composition of ants in the two aforementioned landscapes differs by 34 species from the results recorded in the present study, including several species of Strumigenys and Gnamptogenys. These agricultural ecosystems seem to be composed of significant ant fauna, considering that Fernández and Sendoya (2004) reported 700 species in Colombia.
The total number of species counted in the SHD sample was three species more than in the forest patches, but the latter contain the most exclusive species (21), with a smaller representation of total area in the region. However, the use of an effective number of species makes it possible to draw direct comparisons of the magnitude of the difference among communities (Jost, 2007; 2010), and the use of a fixed percentage of sample coverage makes it possible to adjust for the number of samples without losing information; this provides greater certainty to the finding that forests have more diverse ant communities than do other land use types. This difference suggests that forests have, on average, 1.25 and 1.6 times more diversity of ants than do SHD and SUN, respectively. In other words, SHD maintains 80.3 % and SUN 62.3 % of the diversity of these forests. Clearly, the SUN sites represent environments with the fewest number of species, differing statistically from other land uses, possibly due to factors related to the intensification of coffee cultivation. It is possible that the mechanisms that explain this difference are both physiological and ecological in nature, as reported by Philpott and Armbrecht (2006).
Our results are consistent with other studies in which forest patches, even when significantly disturbed, function as a reservoir of species or sometimes as a connector between cultivated areas and native forests (Zabala et al., 2013). Similarly, shaded coffee plantations can function as a favorable environment for ant fauna (Philpott, Perfecto, & Vandermeer, 2006; Perfecto & Vandermeer, 2015). Armbrecht, Rivera & Perfecto (2005) and Urrutia-Escobar and Armbrecht (2013) revealed a reduction of ant species richness associated with the intensification of coffee cultivation. García et al. (2008) also reported greater diversity of ants in areas with greater vegetation cover, such as forest patches and shaded coffee plantations, and a reduction of this diversity in exposed coffee plantations.
The composition of species found in our study partially coincides with records of other coffee-growing areas of the country, as well as expanding the distribution of several species in the department of Cauca. The composition of ant species in the region’s forests differed significantly from the majority of the coffee plantations, especially those that lacked shade. In fact, the exposed coffee plantation displayed notable differences in both composition and diversity from habitats that contain trees (forests and shaded coffee plantations). The biological value of forests is especially noteworthy for the species found exclusively in forested patches, such as A. tristani, A. goniodes, Fulakora orizabana, S. raptans, S. nubila, S. connectens and T. pusillus, among others; the presence of these species indicates that, although they have been significantly disturbed and not protected by any means, these small patches of forest still preserve fauna typical of native forest ecosystems. As in other studies, in the Colombian coffee zone, shaded coffee plantations have a richer and more equitable ant fauna than unshaded coffee plantations, reinforcing the idea that implementation of shade favors the establishment of ant fauna.
The rank abundance curves were constructed based on the frequency of capture along the sampling points, attempting to reduce the effect of these social insects’ foraging type. The species’ distribution according to their abundance varied for the three types of land use, i.e., each land use presented a different species as being most abundant; this is the result of differences in the structure of communities, in part a result of history, land use and the requirements of the species. It is interesting to note that C. rimosus is one of the dominant species in the soil of the forest patches. This species, which cultivates fungi, is characterized by small nests and prefers open, damp areas (Mehdiabadi & Schultz, 2010); however, this species was not abundant in areas of other land use types, possibly because the moisture and type of leaf litter in forest soils favored the species, whereas some agronomic practices in planted areas may have hindered the species’ establishment. Conversely, species such as L. piliferum and Pheidole sp. 4, which are generalists, were very abundant in coffee plantations but rare in forest patches. It is worth noting that L. piliferum has been reported to care for root aphids such as Aleyrodidae and Pseudococcidae (Wild, 2007), a relationship that has been reported in Colombia by Mera-Velasco, Gallego-Ropero, and Armbrecht, (2010) and by Ramirez, Herrera, and Armbrecht (2010) in the departments of Cauca and Valle del Cauca, respectively. This situation needs to be investigated in greater detail, given that several nests of this ant species were observed to contain aphids among the roots of coffee plants, especially in the exposed coffee sites.
The analysis of similarity based on the composition of species reinforces the aforementioned finding: the ant fauna found in forest patches differs from that found in coffee plantations, sharing only 12.4 % of its species with exposed coffee plantations. This is in part because 22 species found in SUN were not found in the forest patches, whereas the latter displayed 21 exclusive species. The shaded coffee plantations occupy an intermediate position between forest patches and SUN sites, with some SHD sites conveying greater resemblance to forest patches in their species composition, whereas others more closely resemble SUN sites. These results signify the importance of preserving the few remaining forest fragments embedded in the agricultural matrices, especially because they are not protected by conservation measures. Ideally, these forest patches will function as a source of species and proper shade management practices in coffee plantations will contribute to an increase of associated diversity in coffee production areas.
The classification of ants by functional groups or guilds and knowledge about the functionality and importance of ants in soil-related processes has been shown to facilitate a predictive understanding of how ant communities respond to disturbances (Silvestre et al., 2003). The fact that ten different groups were observed indicates the versatility of roles played by these ants in coffee farming systems. The dominant group of omnivorous ground-dwelling ants displayed the highest species richness, which is explained by the presence of the genera Pheidole and Solenopsis. The former is widely distributed globally, classified as hyperdiverse and one of the most common genera in the Neotropics, with approximately 500 species (Wilson, 2003); Solenopsis is also described as very common in leaf litter, with more than 90 species in the Neotropics (Fernández, 2003). These genera have been reported in farming systems of several countries (Pacheco, Vasconcelos, Groc, Camacho, & Frizzo; Saad, SouzaCampana, Bueno, & Morini, 2017; Perfecto & Vandermeer, 2015), and some of their species have been indicated as potential predators of the coffee berry borer (Hypothenemus hampei Ferrari) (Gallego & Armbrecht, 2005; Philpott & Armbrecht, 2006).
A comparison of guilds between the different land uses also reflects greater similarity between forest patches and shaded coffee plantations, but an analysis of the composition of species reveals marked differences among some groups. The large ground-dwelling predators are most affected. Only two of the eight species reported in this study were found with high frequency in the SUN sites: O. chelifer (104 records) and E. ruidum (84 records), with a single record of Rasopone ferruginea reflecting an isolated event. In contrast, R. ferruginea is the dominant large predator in forest patches (43 records), indicating that coffee plantations do not offer the minimal ecological requirements for this species, which remains isolated in the region’s few conserved patches of forest. Similar results were found by Dias et al. (2013) in unshaded coffee in Brazil and by Philpott et al. (2006) in Mexican coffee plantations. Conversely, E. ruidum was not found in the forests studied. Endemic in Central and South America and considered an opportunistic generalist found in abundance in disturbed sites (Fernández, 1991), this species has been described as a dominant species because it is present in a variety of habitats, commonly including coffee and cocoa plantations of Central America (Lachaud, Valenzuela, Corbara, & Dejean, 1990,Santamaria, Armbrecht, & Lachaud, 2009). Its opportunistic behavior may allow this species to exploit places where other predatory species cannot compete, which could explain its dominance in exposed coffee plantations.
The group of specialized predatory cryptic Myrmicinae also varies according to land use: among the nine species recorded in forest patches, five were exclusive, among which the genus Strumigenys was most represented. In addition, the dominant species as measured by frequency of capture differed for each land use: Strumigenys nubila was dominant in the forest patches, Nesomyrmex asper was dominant in the SHD sites, and Cardiocondyla minutior was dominant in the SUN sites. Species of small mass recruitment tree-dwellers were abundant in the coffee plantations (SHD and SUN sites) but exhibited low frequency of capture in forests. These ants were representative in the sample, although the methodology used was aimed at ground-dwelling ants. It is possible that the scarcity of trees in the coffee plantations pressures these species to forage on the ground; in fact, a study unrelated to the current study using tuna bait to collect ants in vegetation confirmed the presence of these species in forest vegetation (Jiménez et al., 2016).
Among the fungus-growing Attini group, C. rimosus were dominant in forests and A. cephalotes were dominant in the SUN sites. More fungus-growing species were established in the forest patches, the majority of which have cryptic coloration, are inconspicuous, dig and build small inconspicuous nests and use decomposing organic matter to cultivate their fungi (Della Lucia, 2011). Therefore, they need appropriate conditions of moisture, temperature and organic matter, which are more easily found in forests. In contrast, A. cephalotes, or leaf-cutter ants, strip fresh plant material in large quantities as a food resource to cultivate the symbiotic fungus that feeds their large colonies (Hölldobler & Wilson, 2010). Their nest-building and vegetation-cutting activities can drastically affect the functioning of ecosystems (Farji-Brener & Illes, 2000, Meyer, Leal, Tabarelli, & Wirth, 2011); their polyphagous habits make them among the most destructive pests of various crops in South America (Della Lucia, 2011, Montoya-Lerma, GiraldoEcheverri, Armbrecht, Farji-Brener, & Calle, 2012). It has been reported that leaf-cutter ants of the genus Atta show a preference for open or sparsely vegetated locations for nesting (Vasconcelos, Vieira-Neto, Mundim, & Bruna, 2006), suggesting a greater concentration of nests in places changed by human activity, such as unshaded coffee. In general, the species most abundant in coffee plantations are described as generalists of wide distribution, common in agro-ecosystems (Delabie et al., 2000; García et al., 2008).
An analysis of the environmental variables reveals differences among the conditions each land use has and how these differences can influence the establishment of ant communities, which is observable in the 81.34 % of variability in the composition of species found. Forest patches preserve very specific conditions that make them environmentally different from coffee plantations; however, no statistically significant differences were found between forest patches and SHD sites in terms of the leaf litter depth, canopy cover or canopy height. This result demonstrates, on the one hand, the effect of human intervention in the forest patches studied and, on the other hand, the presence of some trees with thick trunks and well-developed canopies capable of providing a thick layer of leaf litter in some shaded coffee plantations. The presence of shade trees in the plantations promotes biodiversity in agroecosystems (Majer & Delabie, 1999) and additional sources of income to farmers (Philpott & Dietsch, 2003). This finding underscores the dramatic change that accompanies the removal of tree vegetation from habitats managed by humans.

