











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkCellulose is considered to be the most abundant organic compound derived from plant biomass. Perpetual renewal of plant biomass via the process of photosynthesis ensures an inexhaustible supply of such material. The worldwide production of this biopolymer is estimated to be between 1010 and 1011 tons each year (Lynd, Weimer, van Zyl, & Pretorius, 2002). Cellulose is a linear homopolysaccharide of β-1.4-linked anhydro-D-glucose units with a degree of polymerization of approximately 10 000 for cellulose chains in nature and 15 000 for native cellulose cotton (Habibi, Lucia, & Rojas, 2010; Siqueira, Bras, & Dufresne, 2010).
Cellulolytic enzymes degrade cellulose by cleaving the glycosidic bonds. Many fungi and bacteria are capable of producing multiple enzymes, collectively known as cellulases, that act in a synergistic manner to hydrolyze the β-1,4-D-glycosidic bonds within the cellulose molecule (Akiba, Kimura, Yamamoto, & Kumagai, 1995). Cellulases can be classified into three types: endoglucanases (endo-β-1,4-glucanase, EC 3.2.1.4), cellobiohydrolases (cellulose β-1.4-cellobiosidase, EC 3.2.1.91), and β-glucosidases (β-D-glucoside glucohydrolase, EC 3.2.1.21). Endoglucanases randomly hydrolyze internal β-1,4-D-glycosidic bonds in cellulose. As a result, the polymer rapidly decreases in length and the concentration of the reducing sugar increases slowly. Cellobiohydrolases hydrolyze cellulose by removing the cellobiose units from the nonreducing end of cellulose, and β-glucosidases, cleave glucose units from cellooligosaccharides (Siddiqui, Shemsi, Anwar, Rashid, & Rajoka, 1999).
In Argentina, subtropical and tropical rainforests are restricted to small areas that do not reach 2 % of the land area, and yet, are home to more than 50 % of the biodiversity (Brown, Grau, Lomáscolo, & Gasparri, 2002). Specifically these forests belong to two phytogeographic regions of the Amazon Domain: Yungas rainforests, located on the eastern slopes of the foothills in the Northwest of the country; and the Paranaense rainforest located in the province of Misiones and Northeast of Corrientes (Cabrera, 1994). Currently, studies of wood-decaying fungi diversity are in an active process of analysis and dissemination (Rajchenberg & Robledo, 2013; Gallo, Robledo, Romero, & Catania, 2014; Robledo, Giorgio, Franco, Popoff, & Decock, 2014).
However, knowledge about the cellulolytic enzyme production of these fungi is very much restricted. The forest environments in the province of Misiones in Northeast Argentina, are a rich reservoir of biodiversity (Zuloaga, Morrone, & Rodríguez, 1999). Exploration and description of new fungi from Paranaense rainforest can reveal innovative abilities for potential biotechnological applications such as plant biomass conversion (Saparrat, Martínez, Cabello, & Arambarri, 2002; Giorgio et al., 2012; Daâssi et al., 2016). Moreover, the relevance of screening of new isolates in addition to culture collection strains has been already highlighted (Tekere, Mswaka, Zvauya, & Read, 2001). In this sense, although most reports available are related to the Iguazú National Park (Wright, Lechner, & Popoff, 2008), there is no data for the rest of the region that remains still underexplored (Niveiro, Popoff, & Albertó, 2010).
A high forestry activity is found in the province of Misiones-Argentina due to availability of a lot of plant sources. Therefore, there is an increase of woody wastes (Teza et al., 2012). In spite of the huge amount of this plant material, there are still burning practices for its use, however the law XVI N°106 since 2012 prohibits this activity and promotes their utilization in sustainable processes (Sistema Argentino de Información Jurídica, 2012). This has lead to the search for innovative biotechnological strategies such as the production of second generation biofuels, which is also depending upon reactive and enzymes for the process. In this sense, the availability of enzyme cocktails from autochthonous fungal sources is also a priority (King, Donnelly, Bergstrom, Walker, & Gibson, 2008).
In the present study, we analyzed and compared the cellulolytic ability of 22 white-rot fungi, isolated from woody materials collected from the subtropical rainforest of the province of Misiones, Argentina, grown on agar medium supplemented with either sodium carboxy-methylcellulose (CMC) or crystalline cellulose (CC). Furthermore, submerged cultures of these fungi in a liquid medium with CMC were conducted to select the isolate with the highest level of filter paper assay (FPase) and endo-β-1,4-glucanase (EG) activities, which was subsequently identified at molecular level.
Materials and methods
Fungal isolates: Fungal isolates from different sources were selected for this study (Table 1). They belong to the following culture collections: 1. Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Buenos Aires (FCEyN-UBA); 2. Instituto Multidisciplinario de Biología Vegetal (IMBIV-CONICET) and 3. Instituto de Biotecnología Misiones, Universidad Nacional de Misiones (InBioMis-UNaM). Stock cultures of the Basidiomycetes isolates tested were kept at 4 °C on 20 (g L-1) agar slants supplemented with 12.7 (g L-1) malt extract (Saparrat et al., 2002).
Screening for fungal growth and cellulolytic ability on agar medium: Cellulolytic ability of each fungal isolate was determined by using plate screening medium containing Czapek’s mineral salt solution (NaNO3 2 g, KH2PO4 1 g, KCl 0.5 g, MgSO4.7H2O 0.5 g, FeSO4.7H2O 0.01 g, H2O 1 000 mL, adjusted to pH 5.0 with glacial acetic acid) and agar (20 g L-1) supplemented with 1 g L-1 of CMC (Sigma-Aldrich, USA) or with 1 g L-1 of CC (Sigma-Aldrich, USA) as sole carbon source. Agar block (6 mm in diameter) from five days old fungal colony grown on MEA plates was cut and inoculated in the center of the plates. Inoculated plates were incubated at 28 ± 2 °C for 4 days to estimate the growth extension of the colony (Guigón-López et al., 2010; Andrino, Morte, & Honrubia, 2011) and the degradation of carbon substrate supplemented through the procedure of Mangelli & Forchiassin (1999), calculated as the Cellulolytic Efficiency Index (CEI) according to Martínez, Chiocchio, & Godeas (2001). All analyzes were performed in duplicate.
Table 1: Fungal isolates used in this work and their source of conservation
| Culture Collection | Fungal isolates |
|---|---|
| Colección de Cultivos de Hongos, Departamento de Biodiversidad y Biología Experimental, FCEyN, UBA | Coriolus versicolor (L.) Quél. BAFC 266 |
| Irpex lacteus (Fr.) Fr. BAFC 1171 | |
| Lenzites elegans (Spreng.) Pat. BAFC 2127 | |
| Phlebia brevispora Nakasone BAFC 633 | |
| Pycnoporus sanguineus (L.) Murrill BAFC 2126 | |
| Trametes villosa (Sw.) Kreisel BAFC 2755 | |
| Laboratorio de Biotecnología Molecular, InBioMis | Trametes sp. LBM 029 |
| Trametes sp. LBM 030 | |
| Trametes sp. LBM 031 | |
| Trametes sp. LBM 033 | |
| Trametes villosa (Sw.) Kreisel LBM 109 | |
| Trametes villosa (Sw.) Kreisel LBM 110 | |
| Trametes villosa (Sw.) Kreisel LBM 111 | |
| Irpex sp. LBM 032 | |
| Irpex sp. LBM 034 | |
| Pleurotus sajor-caju (Fr.) Singer LBM 105 | |
| Pycnoporus sanguineus (L.) Murrill LBM 106 | |
| Pycnoporus sanguineus (L.) Murrill LBM 107 | |
| Pycnoporus sanguineus (L.) Murrill LBM 108 | |
| Laboratorio de Micología, IMBIV-CONICET | Perenniporia martius (Berk.) Ryvarden Cu 40 |
| Perenniporia martius (Berk.) Ryvarden Cu 57 | |
| Perenniporia martius (Berk.) Ryvarden Cu 58 |
Fungal screening for selected cellulolytic enzymatic activities under submerged conditions: Thirty mL Czapek’s mineral salt solution (adjusted to pH 5.0) supplemented with 1 g L-1 of peptone and 5 g L-1 of CMC in 250 mL Erlenmeyer flask was inoculated with each fungus, and incubated at 28 ± 2 ºC under static condition for 14 days. After 4, 9 and 14 days of incubation the culture medium was collected, centrifuged at 10 000 xg for 15 min, and the clarified supernatant was used to estimate total cellulose and endo-β-1,4-glucanase activities. FPase and EG activities were determined according to Ghose (1987) using Whatman no.1 filter paper and CMC as substrates. The released reducing sugars were estimated as glucose by the DNS method (Miller, 1959). One unit of FPase and EG activities were defined as the amount of enzyme required to liberate 1 μmol/mL min of glucose from the particular substrate under the assay conditions.
Molecular identification of the fungal isolate LBM 034: DNA extractions and PCR amplifications: Genomic DNA was extracted from a three day old mycelium grown on 12.7 g L-1 malt extract and corn extract 0.5 % v v-1according to Fonseca et al. (2016). PCR was performed in 20 µL reactions, using 1 Unit of Taq DNA polymerase (Thermo Scientific, Fermentas), 2 µL of 10X Taq polymerase buffer, 2 µL of 25 mM MgCl2, 1.4 µL of 2.8 mM dNTPs, 1 µL of each 10 µM primer and 1 µL of 60 ng template DNA. The oligonucleotide primer set ITS1 and ITS4 was used to amplify the ITS region of the rDNA (White, Bruns, Lee, & Taylor, 1990). Amplifications were performed on a Labnet thermal cycler model MultiGene TM II Personal Thermal Cycler using the following parameters: 4 min step at 94 °C, followed by 35 cycles of 40 s at 94 °C, 40 s at 55 °C and 40 s at 72 °C, and a final 10 min extension step at 72 °C.
DNA sequencing was performed on both strands in triplicate using the primers ITS1 and ITS4. Sequencing assays were analyzed on an automated DNA Sequencer ABI3730XL by Macrogen Korea. The nucleotide sequences were aligned using the Bio-Edit software.
Phylogenetic analysis: For primary sequence analyses, the software tools on the Fungal Barcoding (http://www.fungalbarcoding.org) and NCBI (http://www.nih.ncbi.gov) servers were used. The ITS sequence with the highest identity was obtained by nucleotide BLAST searches (Altschul et al., 1997). For nucleotide sequence phylogeny, maximum-parsimony (MP) method was adopted. Phylogeny analysis was carried out using the T.N.T version 1.1 software (Goloboff, Farris, & Nixon, 2008). To assess the support for the identified groups, we performed a Bootstrap test. Bootstrap analyses included 1 000 resampled matrices. For each resampled matrix, we performed 100 Random Addition Sequences (RAS) + Tree Bisection Reconnection (TBR) cycles. Sequences were aligned using T-Coffee Multiple Sequence Alignment. All alignment gaps were treated as missing data. The alignments were manually trimmed (overhangs were removed and gaps were corrected) prior to phylogenetic calculations. The ITS region of 5.8S ribosomal DNA sequences was submitted to GenBank (accession LN606596).
All results are shown as mean ± standard deviation (SD). The differences in fungal growth and CEI between isolates were analyzed by a one-way analysis of variance (ANOVA) and means were contrasted by the Tukey Test, using the InfoStat version 2013 software (Grupo InfoStat, FCA, UNC, Argentina). To establish differences of extracellular total cellulase and endo-β-1,4-glucanase activities among isolates, ANOVA tests were performed.
Results
Screening for fungal growth and cellulolytic ability on agar medium: Screening for fungal growth and cellulolytic ability was carried out by the hydrolysis of CMC or CC supplemented to the basal medium. All fungi showed ability to grow and degrade both substrates.
The Phlebia brevispora BAFC 633 isolate showed the fastest radial growth rate (0.44 ± 0.01 mm h-1, P < 0.05) on CMC medium, whereas Trametes sp. LBM 033, Perenniporia martius Cu 40, P. martius Cu 57 and P. martius Cu 58 isolates showed the slowest growth rate on this substrate with a value of 0.05 ± 0.01 to 0.10 ± 0.004 mm h-1. The Irpex lacteus BAFC 1171, P. brevispora BAFC 633, Lenzites elegans BAFC 2127, and Trametes villosa LBM 110 isolates showed the fastest radial growth rate (between 0.39 ± 0.01, and 0.37 ± 0.01, P > 0.05) on CC medium, whereas P. martius Cu 40, P. martius Cu 57, and P. martius Cu 58 isolates showed the lowest ability to grow on this medium (0.07 ± 0.01 mm h-1, Figure 1a).
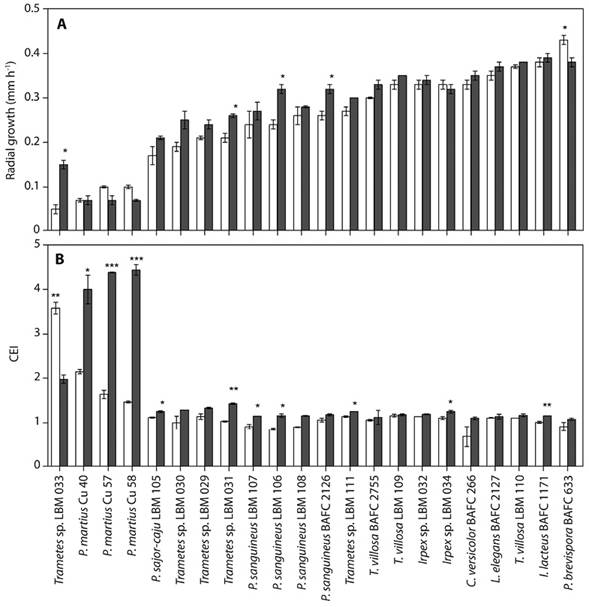
Figure 1 a. Radial growth of the colony under selective conditions in test plate. Clear (), and filled (■) bars represent culture medium containing CMC and CC, respectively. Asterisks (*) denote significant differences (P < 0.05, Tukey Test) in the growth extension of a given isolate on both types of substrates used. Data are mean ± SD of duplicates. b. Cellulolytic Efficiency Index (CEI) of the isolates. Clear (), and filled (■) bars represent culture medium containing CMC and CC, respectively. Asterisks (*) denote significant differences (P <0.05, Tukey test) in the CEI of a certain isolate on the two types of substrates used. Data are mean ± SD of duplicates.
The Pycnoporus sanguineus BAFC 2126, Trametes sp. LBM 031, Trametes sp. LBM 033, and P. sanguineus LBM 106 isolates showed differences in their radial growth ability. They were more efficient growing on CC medium (P < 0.05). On the other hand, P. brevispora BAFC 633 isolate was more efficient growing on CMC medium (P < 0.05, Figure 1a).
The cellulolytic ability of all fungi isolates was obtained by calculating the CEI. Trametes sp. LBM 033 isolate showed the most efficient degradation of CMC with a CEI of 3.58 ± 0.13 (P < 0.05), followed by P. martius Cu 40 (with a CEI of 2.15 ± 0.015; P < 0.05), P. martius Cu 57, and P. martius Cu 58 isolates (1.64 ± 0.09, and 1.47 ± 0.02, P < 0.05). CEI values lower than 1.16 were obtained for the rest of the filamentous fungi tested; BAFC 266 isolate presented the lowest CEI with a value of 0.68 ± 0.023.
Perenniporia martius Cu 58, and P. martius Cu 57 isolates exhibited the most efficient degradation of CC with a CEI of 4.44 ± 0.12, and 4.38 ± 0.01, respectively. These values were followed by P. martius Cu 40 isolate (with a CEI of 4.00 ± 0.32; P < 0.05, Figure 1b). CEI values lower than 1.98 were obtained for the other filamentous fungi tested.
Pycnoporus sanguineus LBM 107, T. villosa LBM 111, P. sanguineus LBM 106, Irpex sp. LBM 034, Pleurotus sajor-caju LBM 105, and P. martius Cu 40 isolates showed differences in their cellulolytic ability. They were more efficient in the degradation of the CC (P < 0.05), while Trametes sp. LBM 033 isolate was more efficient in the degradation of the CMC (P < 0.01, Figure 1b).
All isolates were selected for further studies related to cellulolytic enzymatic activities under submerged conditions.
Fungal screening for selected cellulolytic enzymatic activities under submerged conditions: At 4 days of incubation, cellulolytic activities (both FPase, and EG) were found in all the supernatants analyzed. I. lacteus BAFC 1171, P. sanguineus BAFC 2126, L. elegans BAFC 2127, P. sanguineus LBM 106, P. sanguineus LBM 107, P. sanguineus LBM 108, Trametes sp. LBM 029, Trametes sp. LBM 030, Trametes sp. LBM 031, Trametes sp. LBM 033, and Irpex sp. LBM 034 isolates presented high FPase activity (values between 33.70 and 43.73 U L-1), without significant differences (P > 0.05).
At 9 days, P. brevispora BAFC 633, P. sanguineus BAFC 2126, P. sanguineus LBM 106, P. sanguineus LBM 107, Trametes sp. LBM 030, and Irpex sp. LBM 034 isolates presented high FPase activity (between 37.33, and 43.88 FPase L-1), while P. brevispora BAFC 633, P. sanguineus LBM 106, and Irpex sp. LBM 034 isolates showed the highest FPase activity at 14 days of incubation (with values between 55.17, and 65.45 FPase L-1; Figure 2a).
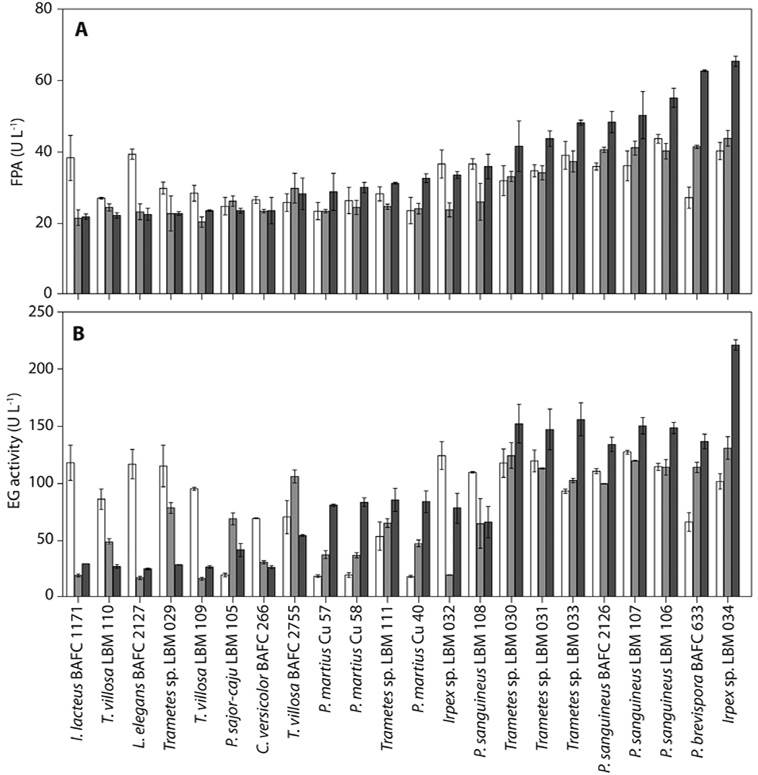
Figure 2 a. Extracellular cellulolytic activity of fungal isolates in submerged culture: Total cellulase activity (FPase). Clear (£), shadowed (¢), and filled (¢) bars represent 4, 9, and 14 days of incubation, respectively. Data are mean ± SD of duplicates. b. Endo-β-1,4-glucanase activity (EG). Clear (£), shadowed (¢), and filled (¢) bars represent 4, 9, and 14 days of incubation, respectively. Data are mean ± SD of duplicates.
At 4 days of incubation, I. lacteus BAFC 1171, P. sanguineus BAFC 2126, L. elegans BAFC 2127, P. sanguineus LBM 106, P. sanguineus LBM 107, T. villosa LBM 109, T. villosa LBM 110, Trametes sp. LBM 029, Trametes sp. LBM 030, Trametes sp. LBM 031, Irpex sp. LBM 033, and Irpex sp. LBM 034 isolates showed high EG activity (values between 86.36, and 127.53 U L-1), without significant differences (P > 0.05), and P. sajor-caju LBM 105, P. martius Cu 40, P. martius Cu 57, and P. martius Cu 58 isolate presented low EG activities. At 9 days, P. brevispora BAFC 633, T. villosa BAFC 2755, P. sanguineus LBM 106, P. sanguineus LBM 107, Trametes sp. LBM 030, Trametessp. LBM 031, and Irpex sp. LBM 034 isolates showed high EG activity (values between 106.15, and 130.97 U L-1), whereas T. villosa LBM109, I. lacteus BAFC 1171, and L. elegans BAFC 2127 isolates showed low EG activity.
Finally, at 14 days of incubation Irpex sp. LBM 034 isolate presented the highest EG activity (221.21 U L-1; P < 0.05) whereas P. brevispora BAFC 266, I. lacteus BAFC 1171, L. elegans BAFC 2127, T. villosa BAFC 2755, P. sajor-caju LBM 105, T. villosa LBM 109, T. villosa LBM 110, and Trametes sp. LBM 029 isolates showed low EG activity (Figure 2b).
Therefore, Irpex sp. LBM 034 isolate was selected for its cellulolytic ability showing the highest values of FPase and EG activities in submerged cultures analyzed after 14 days of incubation.
Molecular identification of the fungal isolate LBM 034: Irpex sp. LBM 034, isolated from the Misiones subtropical rainforest, was selected due to its outstanding potential for EG production. It was deposited at the Culture Collection of the Laboratorio de Biotecnología Molecular of the InBioMis. Because of these reasons, the fungus was explored at the molecular level for the first time.
The blast search indicated that LBM 034 isolate correspond to the specie Irpex lacteus (Fr.) Fr., as it showed the highest sequence similarities to I. lacteus strain KUC8958 (JX311924.1; 97 %), I. lacteus strain CB1 (KF318788.1; 97 %), and I. lacteus strain KUC9124 (JX290578.1; 97 %).
The phylogenetic tree was reconstructed by the maximum parsimony method, based on ITS1-5.8S-ITS2 rDNA region sequences. Alignment of all 38 ITS sequences from Basidiomycota yielded a consensus length of 503 characters, of which 338 (67 %) were parsimony informative. Parsimony analysis of combined nucleotide and gap data yielded a single most-parsimonious tree of 2163 steps (CI = 0.41, RI = 0.58). Phylogenetic tree showed that the LBM 034 isolate formed a cluster with species of I. lacteus with 100 % bootstrap of branch support (Figure 3). Therefore, the isolate selected in this study was assigned to the taxon I. lacteus.
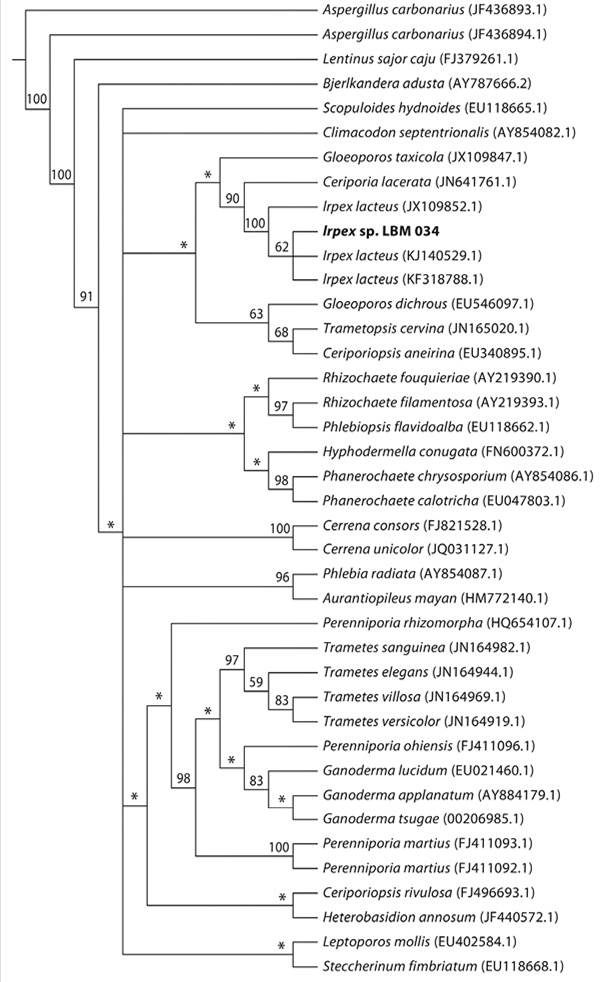
Figure 3 Phylogenetic tree performed using maximum parsimony method based on the sequence alignment of the ITS1-5.8S-ITS2 of the fungus under study with homologous sequences obtained from the GenBank (NCBI). The tree was constructed with T.N.T. program. Confidence levels were obtained by bootstrapping (1 000 replicates) and indicated at each node. *Represent the collapsed branches below a cutoff of 50.
Discussion
Although the diversity of wood-decaying fungi associated to the Paranaense rainforest region has been studied (Rajchenberg & Robledo, 2013; Gallo et al., 2014; Robledo et al., 2014), only a few researchers have explored the fungal biota of this environment, and the knowledge of cellulolytic enzyme production has been very much restricted. Therefore, the present study was carried out to analyze the cellulolytic ability of 22 white-rot fungi, isolated from woody materials collected from the subtropical rainforest of Misiones, Argentina.
First, the growing ability of filamentous fungi on agar medium supplemented either sodium carboxy-methylcellulose or crystalline cellulose as sole carbon source was scanned. Mycelia growth extension is indicative of the ability to synthesize and produce extracellular hydrolytic enzymes involved in cellulolysis, providing the nutritional requirements of these fungi of biotechnological importance (Nwodo-Chinedu, Okochi, Smith, & Omijidi, 2005).
All isolates showed ability to grow and degrade both substrates in spite of their different nature; one is a soluble derivative (CMC) and the other is an insoluble element (CC). Phlebia brevispora BAFC 633 isolate was the only fungus that showed the faster radial growth rate on CMC in comparison with CC medium, whereas P. sanguineus BAFC 2126, Trametes sp. LBM 031, Trametes sp. LBM 033, and P. sanguineus LBM 106 isolates showed the faster radial growth rate on CC in comparison with CMC medium.
Compared to our results, the Ascomycota Trichothecium roseum (Pers.) Link showed a similar radial growth rate to that P. brevispora BAFC 633, I. lacteus BAFC 1171, and T. villosa LBM 110. On the other hand, Shanmugam, Mani, & Narayanasamy (2008) reported a higher growth in this fungus on a medium using CMC compared to one with cellobiose or filter paper. Kudanga & Mewenje (2005) found a higher growth in some filamentous fungi on CMC medium due to the ability of CMC to act as an indirect inducer of cellulase production.
Trametes sp. LBM 033 isolate showed greater CEI on CMC medium, whereas I. lacteus BAFC 1171, Trametes sp. LBM 031, I. lacteus LBM 034, P. sajor-caju LBM 105, P. sanguineus LBM 106, P. sanguineus LBM 107, P. sanguineus LBM 108, T. villosa LBM 111, P. martius Cu 40, P. martius Cu 57, and P. martius Cu 58 isolates showed greater CEI on CC medium. On the other hand, no data are available on the physiology and the cellulolytic enzymatic system of P. martius, being this work the first report about its ability to degrade cellulose using extracellular hydrolytic enzymes.
The differences observed according to the cellulosic sources used may be indicative of a differential ability of these isolates to produce cellulolytic enzymes, possibly involving specific mechanisms of induction (Ganner et al., 2012). However, other characteristics of the enzymes produced in the culture medium, linked to their stability and activation can lead to differences in the extent of the degradation halo observed with regard to the diameter of the fungal colony (Llacza-Ladera, 2012).
To quantitatively evaluate the cellulolytic ability of the 22 isolates, they were scanned for the production of two extracellular hydrolytic enzymes under submerged fermentation.
The reduction in levels of cellulolytic enzyme activity increasing incubation time in some isolates could be attributed to several reasons such as the production and activity of proteases during growth (Ohnishi, Yoshida, & Sekiguchi, 1994), the acidification of the medium according to culture time (Gao & Breuil, 1995) and the low stability of cellulases (Janusz, Rogalski, Barwińska, & Szczodrak, 2006). However, it was found that only the Irpex sp. LBM 034 isolate showed a high cellulase activity towards filter paper and CMC substrates with increasing incubation time. Enzymatic activity values of Irpex sp. LBM 034 are higher than those previously reported by Llacza-Ladera (2012) in species of Paecilomyces sp. (48 U L-1) and Aspergillus sp. (48 U L-1). However, results here are somewhat lower than those reported by Vilches (2002) in species of Chrysosporium sp., and Fusarium sp. with activities of 120, and 112 U L-1, respectively. The difference of values obtained in this work with the previously reported values may be explained by the differences in the test conditions, such as incubation time, substrate concentration, temperature, pH and others, which are known to have influence on cellulases production (Li et al., 2013).
The production cost of cellulolytic enzymes has declined over the last decade (Stephanopoulos, 2007), but it is still high enough to seriously affect the lignocellulosic ethanol process (Klein-Marcuschamer, Oleskowicz-Popiel, Simmons, & Blanch, 2012). Most of the cellulolytic enzymes currently available in the market are from fungal sources. The primary industrial use of fungi is the production of enzymes and two species in particular are industrial workhorses: Trichoderma reesei E. G. Simmons and Aspergillus niger Tiegh. However, literally thousands of fungi occupy the same ecological niche as Trichoderma and Aspergillus as decomposers and recyclers of cellulosic biomass and as yet they have not been exploited commercially.
We also demonstrate that LBM 034 isolate produced high EG activity. Therefore, this isolate is a promising fungus for obtaining ethanol from cellulosic biomass.
ITS region sequences are highly used to infer phylogenetic relationships of closely related fungal species, as well as to evaluate the variability within a population and between geographically distant isolates (Hilden, Bortfeldt, Hofrichter, Hatakka, & Lundell, 2008). Based on our phylogenetic analysis (Figure 3), the LBM 034 isolate belongs to Irpex lacteus. Since there are previous reports about other isolates from this species (Hamada et al., 1999; 2001; Novotný, Cajthaml, Svobodová, Šušla, & Šašek, 2009), future studies are still needed using this fungus to evaluate its potential in regional development of bioethanol production.

