











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkTermites are social insects present in tropical ecosystems. They play an important role in the environment in wood decomposition (Miranda, Vasconcellos, & Bandeira, 2004). They are also involved in soil aeration and fertilization due to the nature of their nests, constituted by organic matter, and to the tunnels they built, that facilitates the air exchange (Batalha, Da Silva Filho, & Martius, 1995). Despite their positive impact on ecosystems, they are rather considered as agricultural and urban pests.
Approximately 2 800 species of termites have been reported worldwide; most of them consume cellulose in any form, which may affect structures made of e.g. wood, cardboards, and books (Verma, Sharma, & Prasad, 2009). In agricultural systems, termites are considered harmful to many crops such as maize, rice, yams, cotton, sugarcane and eucalyptus (Miranda et al., 2004). From the economical perspective, these feeding preferences can cause big economic loss (Boulogne et al., 2017). In the U.S., for example, it is estimated to cause over 1 billion dollars of damage annually. In Mali alone, the damage caused by several species of termites is responsible of 10 to 30 % in loss of the total peanut crops production year (Verma et al., 2009). Meanwhile, in India they are responsible of 15 to 25 % of damages in corn cultivars; causing over 30 million dollars of loss per year (Verma et al., 2009). In South America, there were no found economical reports of damage caused by termites; however, several termite species have been reported as pest, e.g. Comatermes perfectus (Kalotermitidae), Coptotermes niger (Rhinotermitidae), C. testaceus (Rhinotermitidae), Heterotermes convexinotatus (Rhinotermitidae) and Amitermes foreli (Amitermes) affecting mainly cultivars of coffee (Coffea sp.), corn (Zea mays), eucaliptus (Eucalyptus sp.) and passion fruit (Passiflora edulis) (Abadía, Arcila, & Chacón, 2013).
Synthetic pesticides, on the other hand, remain as the main method to prevent termite attacks on wooden structures. However, the persistence of chemical insecticides along with the damage they cause to environment, resulted in the need to search new techniques for pest control (Verma et al., 2009). Ethological methods are among these “greener” ways of pest control and it consist on designing baited traps with very small quantities of chemical compounds called semiochemicals. These compounds are intermediates in insect communication and affect specifically those considered as pest having a reduced environmental impact (Howse, Stevens, & Jones, 1998).
Note that, from the 2 800 species of termites identified, only few of them have been studied in relation to their chemical communication. So far, this number is close to 60 species (Bordereau & Pasteels, 2010). Interestingly, in Isoptera there are many common compounds in different families and genera, regardless to be located in different geographical regions or even if they are not phylogenetically related, this is in fact something not common in other insect orders in which they present more specificity.
The objective of this study is to isolate and identify the trail-following pheromone of Microcerotermes exiguus (Isoptera: Termitidae). There is not very much information about M. exiguus only that is a phytophagous/xylophagous termite affecting plants from the family Poaceae, being identified as pest in sugarcane cultivars in Brazil (Ferreira et al., 2009). Understanding the chemical communication of this termite may provide a novel approach to control this termite negative impact.
Material and methods
Insect collection: Epigeous nests (up to 20 cm in height and 60 cm in diameter) of M. exiguus (Isoptera: Termitidae) composed of several thousands of individuals were collected at the Simón Bolívar University located in Caracas, Venezuela (10°24’38’’ N - 66°52’88’’ W) in 2010. Nests were taken to the laboratory and placed on a circular plastic container at room conditions (~70 % RH; ~26 ºC) and at an induced photoperiod of 12:12 L:D (Marins & DeSouza, 2008). Each nest was supplied with water and dry wood of Chusquea fendleri collected from the vicinity of the nest.
Body extracts: Foraging workers of M. exiguus were captured and immobilized by freezing at -2 °C for 10 min. Whole body extracts (WBE) were prepared by immersing approximately 20 000 termites on a beaker containing 100 mL of n-hexane (HPLC grade, Mallinckrodt). The extract was hermetically stored at -5 ºC for 24 h, then vigorously agitated and filtered. Subsequently, the extract was concentrated until approximately a volume of 1 mL with a flow of N2 at room temperature. The resulting extract was stored at -20 °C prior to use at different concentration for bioassays, which were done after proper dilutions with n-hexane to obtain the needed concentration for each experiment. These experiments were performed at “Laboratorio de Comportamiento y Control de Plagas” Simón Bolívar University, Caracas, Venezuela.
Chemical micro-reactions: A series of different organic micro-reactions were performed in vials containing WBE, in order to deactivate the pheromone, providing indications on the main functional groups that could be part of the compound (Schulz, 2005). These reactions were: reduction (with NaBH4), saponification, alkaline extraction, silyl-derivatization, bromination and acetylation (Millar & Haynes, 1998).
Solvent-less extraction with solid phase microextraction (SPME): Solid phase microextraction (SPME) was applied in cold-anesthetized workers of M. exiguus with the help of a stereomicroscope. To this purpose, 100 individuals were induced to expose their sternal gland stretching the abdominal sternite with a needle. Subsequently, a SPME fiber of Carboxen/Polydimethylsiloxane (CAR/PDMS) 85 μm (Supelco, Bellefonte, PA, USA) was gently rubbed on this region during 5 min in order to extract the trail following pheromone from their reservoir. The fiber was in perfect contact with this area, allowing it to adsorb any substance on this surface. This action was carefully performed to avoid any damage to the fiber. Controls were carried out by rubbing the dorsal region of workers under the same conditions. The fiber was then desorbed in the injection port of the gas chromatograph for three min under the conditions reported below. The intention of using this technique was having a different approach to isolate the active compound, but using few individuals (100), instead of thousands needed for liquid extraction. Additionally, with SPME analysis we compared the chromatographic patterns from both, dorsal and ventral areas (with the protocol mention above), in order to have more specific information about the zone of production and secretion of this semiochemical.
Gas chromatography-mass spectrometry analysis: Extracts (10) and the DTOH standard were analyzed using GC-MS in an Agilent 7890A GC coupled to a 5975C mass spectrometer instrument (Agilent, Palo Alto California, USA). A DB-5 column (30 m x 0.25 mm i.d., 0.25 μm film thickness; J & W Scientific) fused silica capillary column was used for analysis. Helium was used as carrier gas and injections in splitless mode were made at 250°C. The initial oven temperature of 40 °C was kept for 1 min, then increased to 240 °C at a rate of 5 °C/min, then finally kept for 1 min. Electron impact mass spectra (70 eV) were obtained in the m/z range of 40-250. We focused our efforts in a targeted analysis looking for fragments corresponding to those alcohols reported previously as pheromones in termites, i.e.: 5E,9E-2,6,10-trimethylundecadien-1-ol, 4,6-dimethylundecan-1-ol, (3Z)-dodec-3-en-1-ol, (3Z,6Z)-dodeca-3,6-dien-1-ol and finally (3Z,6Z,8E)-dodeca-3,6,8-trien-1-ol (DTOH).
Bioassays: Previous experiments carried out in our laboratory, where extracts of dissected thorax and abdomen from workers of M. exiguus were tested separately through a Y-shape bioassays, confirmed than the gland responsible for pheromone production is located in the abdominal region (data not published). Behavioral experiments were carried out following the method by Cristaldo et al. (2014). The experiment consisted in drawing a Y-shaped trail on a glass platform (20 cm x 20 cm). The Y-shaped was performed with a Hamilton syringe (10 μL), the first straight line of 5 cm were drawn using 5 μL of WBE or different concentrations of the synthetic pheromone. At the bifurcation point, one of the branches was drawn laying also 5 μL of WBE or synthetic pheromone for another 5 cm while the other branch (separated by an angle of 120°) was done with pure n-hexane as control. Each worker of M. exiguus was placed and kept on the beginning of the trail until the assay started. Responses were considered positive when termites walked at least 8 out of 10 cm of the Y-shape trail in less than 5 min. If any termite were still confused after five min, or did not follow the trail the result was considered negative. Platforms were changed once a new test with another worker was performed to prevent any trail reinforcement. A total of 30 replicates were done under light and room conditions. If an active compound is present in hexane extracts, we expected to observe a trail following behavior on the tested individual. If there is any change on the structure of the compound, this effect would be not possible to observe. A chi square test (a < 0.05) was used to analyze preferences of individuals. Recruitment time was determined also in a Y-shaped trail, using hexane WBE of 2, 20, 100, 200 and 450 individuals. Time spent by termites inside a small chamber (2 cm diameter, with a 2 mm open hole), just before the beginning of the trail, was recorded. Each termite was tested only once.
Synthetic pheromone: Synthetic standard of (3Z,6Z,8E)-dodeca-3,6,8-trien-1-ol (DTOH) was kindly provided by Prof. Jocelyn Millar (Riverside, California).
Chi-squared tests were performed with Statistica 10.0 (Statsoft, Tulsa, OK, USA).
Results
The effects of chemical micro-reactions on the whole body extracts of M. exiguus are shown in Table 1. In Table 1 can be seen that there is no statistically significant differences between the reactions of reduction, saponification and alkaline extraction with respect to whole body extracts of M. exiguus. However, after acetylation, silylation or bromination, the trail following behavior was markedly reduced, producing statistically significant differences from the original extract.
Table 1: Number of workers of M. exiguus that followed the path created during bioassays using whole body extracts (WBE) and after each micro-reaction (N = 30, a < 0.05)
| Type of test | Number of workers | X 2 | p |
| M. exiguus WBE (previous the reaction) | 28 | - | |
| WBE - Reduction with NaBH4 | 25 | 1.5 a | 0.227643 |
| WBE - Saponification | 26 | 0.7 a | 0.389424 |
| WBE - Alkaline extraction | 25 | 1.5 a | 0.227643 |
| WBE - Silylation | 11 | 21.1 b | 0.000004 |
| WBE - Acetylation (at 25 °C) | 16 | 12.2 b | 0.000460 |
| WBE - Acetylation (at 70 °C) | 3 | 41.7 b | 0.0000001 |
| WBE - Bromination | 6 | 32.8 b | 0.0000002 |
Different letters shows statistically significant differences.
In the total ion chromatogram (TIC) of whole body extracts, were observed the presence of a peak (Rt: 23.3 min; Retention index: 1483) which its mass spectra has a fragmentation pattern with m/z ions: 41; 55; 67; 79; 91; 105; 119; 135; 142; 180 (Figure 1), it strongly suggest the presence of (3Z,6Z,8E)-dodeca-3,6,8-trien-1-ol. Additionally, a perfect match between the retention time and mass spectra of this compound and a synthetic standard of DTOH was observed (Figure 1A). Moreover, SPME analysis (Figure 2) corroborated, once again, that the alcohol (DTOH) is extracted only from the abdominal region of the termite, specifically from the ventral sternites.
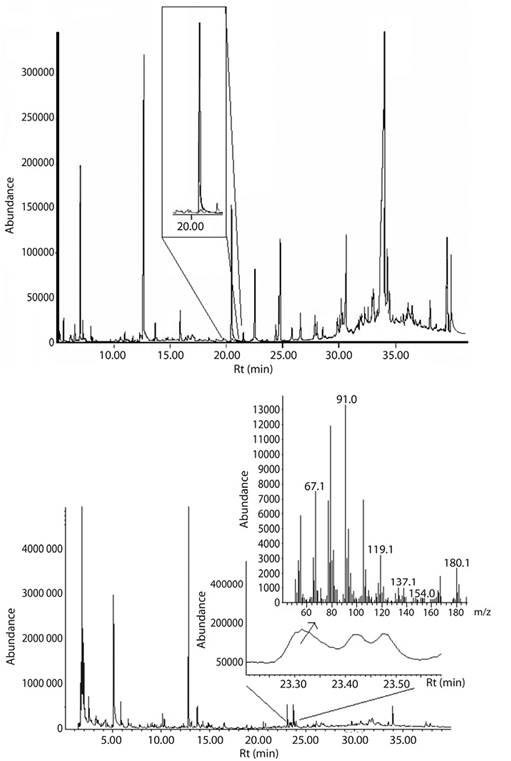
Figure 1 (A) Total ion chromatogram of whole body extracts of M. exiguus, and in the zoomed-in oval section coelution of DTOH (black trace) with WBE (gray trace). (B) Total ion chromatogram (TIC) of SPME analysis (CAR/PDMS 85 µm) from ventral region of 100 M. exiguus workers. Zoom inside the TIC shows a peak eluting at 23.3 min with a very similar mass spectrum of synthetic (3Z,6Z,8E)-dodeca-3,6,8-trien-1-ol (shown at the top right).
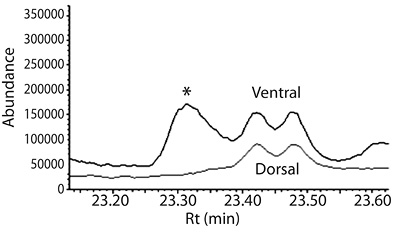
Figure 2 SPME analysis of ventral (glandular) and dorsal region (not glandular) of 100 M. exiguus workers (CAR/PDMS, 85µm). The peak (*) at 23.3 min is exclusive from the ventral region and its mass spectrum is very similar of synthetic (3Z,6Z,8E)-dodeca-3,6,8-trien-1-ol.
Behavioral studies, in a Y-shape assay, were carried out to confirm the activity of synthetic DTOH. The dose-response curve (Figure 3, left) showed that the maximal response of termites with the synthetic DTOH is at a concentration equivalent to 1 pg/cm. Additionally, the synthetic compound, at this concentration, did not show statistically significant differences (N = 30; a < 0.05) with WBE of M. exiguus. Furthermore, the measurement of the time that the termite spent to realize the behavioral assay decrease with the increment on the WBE concentration (Figure 3, right). The recruitment time at higher concentration (10.26 ± 6.05 s) is statistically similar to the recruitment time of the synthetic DTOH at 1pg/cm (13.67 ± 5.76 s). Can be also seen (in the Figure 3, right) that the statistical dispersion is even smaller with the increasing level on the extract concentration, they showed to be less confused (walking randomly) and recognize their own scent and the synthetic pheromone in a minor time. The average walking speed of termites goes around to 0.33 to 0.31 cm/s respectively, and the activity of trials done with WBE, as well as with the DTOH, start vanishing progressively lasting up to two hours (over a smooth-glass surface).
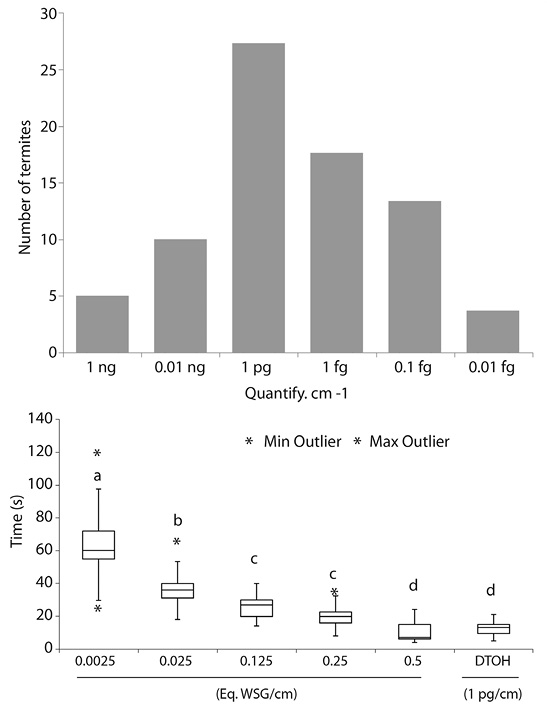
Figure 3 (A) Dose-response curve at different amount of DTOH, representing the number of termites following a trail marked with each value per cm (N = 30; a = 0.05; ng = nanogram, pg = pictogram; fg = femtogram). (B) Bar plots representing the time (s) that workers of M. exiguus are recruited on different concentrations of equivalent of worker sternal gland (Eq.WSG) and DTOH at 1pg/cm. Different letters shows statistically significant differences.
Discussion
A reduction on the trail-following activity, after a chemical micro-reaction, indicated that the molecule responsible for this behavior has been modified. Silylation and acetylation reacts over functional groups with acidic hydrogen such as: -OH; -SOH; -COOH; -SH; -NH2; -NH or -POH, while bromination reacts over double bonds. According to previous reports, and due to the fact that groups like -SOH; -COOH; -SH; -NH2; -NH or -POH are not very common in pheromones, a hydroxyl group is the most probably reacting one and, losing as consequence, its trail-following activity over the termite (Cristaldo et al., 2014). Note that some reactions, such as bromination and acetylation, could have also generated some aversion behavior toward the termites, and this could be an additional factor to considerer on the results. Thus, this study suggested that the pheromone is an “unsaturated alcohol”.
(3Z,6Z,8E)-dodeca-3, 6, 8-trien-1-ol was isolated in 1968 by Matsumura from the termite specie Reticulitermes virginicus and in wood infested from the fungus Lenzites trabea being the first unsaturated alcohol reported as termite pheromone (Matsumura, Coppel, & Tai, 1968). Since then, nine different classes of compounds (alcohols, aldehydes, ketones and terpenes) have been identified using SPME in at least 60 termite species (Bordereau & Pasteels, 2010; Cristaldo et al., 2014). Alcohols compounds reported in termites are: 5E,9E-2,6,10-trimethylundecadien-1-ol (in Mastotermes darwiniensis, Porotermes adamsoni and Stolotermes victoriensis), 4,6-dimethylundecan-1-ol (in Hodotermopsis sjostedti) (Bordereau & Pasteels, 2010), (3Z)-dodec-3-en-1-ol (inMacrotermes annandalei M. bellicosus, M. subhyalinus, M. barneyi, Odontotermes hainanensis, O. maesodensis) (Bordereau & Pasteels, 2010; Peppuy et al., 2001),(3Z,6Z)-dodeca-3, 3-dien-1-ol (in Odontotermes formosanus) (Bordereau & Pasteels, 2010; Du, Lo, Tang, & Liang, 1982), and again (3Z,6Z,8E)-dodeca-3, 6, 8-trien-1-ol isolated in Reticulitermes speratus, R. lucifugus grassei, R. santonensis, Nasutitermes exitiosus and Prorhinotermes simplex as well as in many other species of Rhinotermitidae (Tokoro, Takahashi, Tsunoda, Yamaoka, & Hayashiya, 1991; Wobst et al., 1999; Sillam-Dussès et al., 2010).
The effectiveness of (3Z,6Z,8E)-dodeca-3, 6, 8-trien-1-ol inducing trail following behavior is very high at extremely low concentrations. The maximum response in M. exiguus was obtained at a concentration equivalent to 1 pg of DTOH per cm. Concentrations above or below this value reduced the activity, probably due to the fact that the pheromonal activity occurs within a limited range of concentrations. The values below this range do not trigger alerts in termites. On the other hand, high concentrations can saturate their antennas producing a rejection to the pheromone (Bordereau & Pasteels, 2010). The optimum concentration of pheromone depends on the species. For example, in lower termites the threshold is between 10-1 and 10-2 ng pheromone/cm, but in higher termites, is between 10-2 and 10-3 pg/cm reported with DTOH (Sillam-Dussès et al., 2009; Saran, Millar, & Rust, 2007).
In Nasutitermes corniger, two compounds have been isolated and identified as the trail pheromone, and the induced following behavior is only optimal under an adequate mixing (similar to the naturally deposited amounts) of the components (Sillam-Dussès et al., 2010). The hypothesis proposed for a bicomponent pheromone is that, one of the compound (the more volatile) once is deposited in the substrate is released faster producing a quick recruitment of other individuals from the colony, while the second compound, as consequence of being less volatile, not really contribute to recruitment but provide a long-term effect of orientation to the resources for the colony (Bordereau & Pasteels, 2010; Sillam-Dussès et al., 2010).
During the bioassays, only slight differences have been noticed between the results using WBE or synthetic DTOH. Both, DTOH and WBE, were active for at least two hours, most probably due to the absence of another component in the extract that could lead to any synergistic effect, or that would make the signal to last longer. Likely the rate of release of the DTOH is not only influenced by the characteristic of the mixture, but also on the type of the surfaces or the substrates on which the signal was deposited. For example, Saran et al. (2007) determined that the recruitment effect of DTOH in Reticulitermes hesperus lasts about 24 hours. However, the differences between both results are attributed to the properties of the substrate on which the signal was released.
Despite (3Z,6Z,8E)-dodeca-3, 6, 8-trien-1-ol has several isomers, it is very well documented that changes in the structure reduces the behavioral activity in termites. For example, in Porotermes adamsoni, which secretes identical trail following pheromone to M. darwiniensis, the E isomer is clearly more active than the Z one (Sillam-Dussès et al., 2007). Tokoro, Takahashi, and Yamaoka (1992) isolated the isomer (3Z,6E,8E)-dodeca-3, 6, 8-trien-1-ol in termites, which was present in a very low proportion compared to (3Z,6Z,8E)-dodeca-3, 6, 8-trien-1-ol. However, this compound alone did not induce trail following behavior. Additionally, Tai, Matsumura, and Coppel (1971) demonstrated the isomer (Z,Z,E) was 10 000 times more active than the isomer (Z,Z,Z). Thus, these facts show how stereoisomerism plays a very important role in the chemical communication system in insects, mainly because the chemoreceptors located in the antennae of insects, act as the lock and key model of enzymatic reactions (Kaissling, 2014).
After performing several complementary chemical and behavioral experiments, can be concluded that the main compound responsible for the trail following behavior on Microcerotermes exiguus, is (3Z,6Z,8E)-dodeca-3,6,8-trien-1-ol (DTOH). This information could be used to design different strategies for termite control as an alternative to the excessive use of pesticides.

