











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkEntre los recursos presentes en los bosques neotropicales, las caobas (Swieteniaspp.) y los cedros (Cedrelaspp.) han sobresalido históricamente por la alta calidad de su madera, tanto para ebanistería como para construcción, lo que explica la sobreexplotación a que han sido sometidas estas especies. Por tanto, para suplir las necesidades de la industria, se ha intentado establecer plantaciones extensas, pero sin éxito, debido al daño causado por Hypsipyla grandella(Zeller) (Lepidoptera: Pyralidae), especie nativa del Nuevo Mundo. En efecto, la larva de este insecto barrena los brotes nuevos y, si el brote terminal es atacado, el árbol se bifurca y ramifica, de modo que se malogra su valor comercial (Schabel et al.,1999).
Desde hace medio siglo, en América Latina y el Caribe se han realizado numerosas investigaciones orientadas al manejo integrado de esta plaga, pero esto no se ha logrado, debido sobre todo al nivel de tolerancia tan bajo, pues basta con apenas una larva por árbol para provocar un daño irreversible (Hilje & Cornelius, 2001). No obstante, este fracaso reafirma la necesidad de procurar un enfoque preventivo, dentro del cual podrían ser útiles algunas sustancias que interfieran con el comportamiento de búsqueda o escogencia del hospedante por parte de H. grandella, las cuales podrían incorporarse como ingredientes activos de bioinsecticidas, dentro de una estrategia preventiva para el manejo de dicha plaga.
En tal sentido, hasta ahora se han detectado sustancias fagodisuasivas o represoras de la alimentación de la larva en extractos de madera y follaje del hombre grande (Quassia amara L. ex Blom, Simaroubaceae) (Mancebo et al.,2000), y en el follaje de la ruda (Ruta chalepensis L., Rutaceae) (Mancebo et al.,2001; Soto et al., 2007; Barboza et al. 2010a, 2010b).
Asimismo, con base en evidencias anecdóticas de agricultores acerca del posible efecto de la reina de la noche,Brugmansia candidaPers. (Solanaceae) sobre los adultos deH. grandella, se efectuó una evaluación preliminar con un extracto alcohólico de flores de dicha especie sobre la larva del insecto, la cual reveló que causa efectos análogos a los extractos de las citadas plantas; cabe anotar que el antiguo nombre de dicha especie era Datura arboreaRuiz y Pavón.
Es oportuno mencionar que varias especies de Brugmansiatienen importancia farmacológica, ya que producen alcaloides del tropano. Sin embargo, el fuerte interés comercial en estos compuestos ha provocado un sesgo en la investigación, lo que se refleja en la dificultad de hallar referencias sobre otras familias de compuestos que podrían cumplir otras funciones, incluyendo la defensa contra insectos herbívoros, ya sea mediante toxicidad o fagodisuasión.
Por tanto, el objetivo de esta investigación fue evaluar la posible actividad fagodisuasiva del extracto crudo y de varias particiones fitoquímicas de B. candidasobre la larva de H. grandella, así como determinar las familias de metabolitos secundarios presentes en la flor de dicha planta.
Materiales y métodos
Localización: La preparación del extracto crudo y las particiones fitoquímicas, más las pruebas químicas, se realizaron en los laboratorios de la Escuela de Química de la Universidad Nacional (UNA), en Heredia, Costa Rica, en tanto que los bioensayos se efectuaron en el Laboratorio de Entomología del Centro Agronómico Tropical de Investigación y Enseñanza (CATIE), en Turrialba, Costa Rica. Asimismo, las pruebas cromatográficas para la determinación de la presencia de escopolamina en las particiones más promisorias se realizaron en el Laboratorio Nacional de Servicios Veterinarios (LANASEVE), del Servicio Nacional de Salud Animal (SENASA), del Ministerio de Agricultura y Ganadería (MAG), en Heredia, Costa Rica.
Para lograr mayor uniformidad en las características genéticas y la composición química de las flores requeridas para la preparación del extracto crudo, se recolectaron en un solo sitio y en una sola fecha, al mediodía del 11 de marzo del 2012. El sitio fue elegido debido a la abundancia de plantas en floración, y correspondió a una finca cafetalera ubicada en San José de la Montaña, Barva, Heredia; sus coordenadas son 1 536 m de altitud, y 10°03’21’’N & 84°06’37’’O. Pertenece a una zona con estacionalidad climática, donde hay cinco meses secos, y la precipitación promedio anual es de 2 500-3 000 mm. Asimismo, ahí se presentan los suelos Typic Dystrandept y Lithic Dystrandept, que corresponden con un suelo oscuro y profundo, con buen contenido de materia orgánica y bajo en bases, derivado de cenizas volcánicas (Gómez, 1996).
Obtención del extracto crudo y de las particiones de la flor de B. candida: Se evaluó el efecto fagodisuasivo del extracto crudo de B. candida, siguiendo la metodología empleada por Barboza et al. (2010a). Los detalles de la preparación del extracto crudo en acetona y las particiones (agua, hexano, diclorometano y acetato de etilo), así como de los procedimientos de los bioensayos se encuentran en el Apéndice Digital 1.
Resultados
Efecto del extracto crudo:En relación con el consumo por parte de las larvas, hubo diferencias significativas entre los tratamientos (F= 22.48, g.l.= 6, p < 0.0001) (Figura 1A). Los discos impregnados con las soluciones correspondientes a los tres tratamientos de mayor concentración, tuvieron los menores porcentajes de consumo, y no hubo diferencias entre ellos (p > 0.05); sin embargo, mostraron diferencias con el tratamiento inmediatamente inferior (p < 0.5). Los testigos absoluto y relativo no mostraron diferencias entre sí, y tampoco fueron estadísticamente diferentes del tratamiento de menor concentración (p > 0.05). Los tratamientos a las concentraciones de 0.1 % y 0.3 % m/v no difirieron del testigo absoluto.
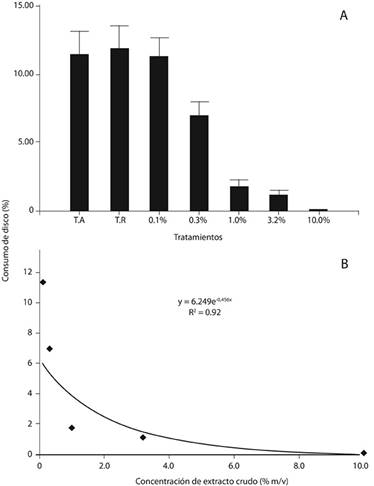
Figura 1. Consumo de discos foliares de cedro tratados con concentraciones crecientes del extracto crudo de la flor de B. candida, en comparación con los testigos absoluto (T.A.) y relativo (T.R.), al exponer larvas de instar III de H. grandella por 24 h (A), así como la relación entre ambas variables (B). En B, la línea continua muestra la tendencia esperada (estimada).
El mejor ajuste de la curva se encontró con un modelo potencial (R2= 0.92) (Figura 1B). En esta, así como en las demás curvas trazadas, se relacionó el porcentaje de consumo como variable dependiente, con las concentraciones de los tratamientos aplicados, tanto de extracto crudo como de sus particiones en cada caso respectivo, las cuales corresponden a la variable independiente. La tendencia revela que al aumentar la concentración del extracto, el porcentaje promedio de consumo foliar disminuye.
Estos resultados muestran que el extracto crudo de la flor de B. Candida posee efecto fagodisuasivo sobre la larva de H. grandella, y que ello se manifiesta a una concentración tan baja como 1.0 % m/v.
Efecto de las particiones fitoquímicas: En el caso del consumo de discos foliares de cedro tratados con la partición de hexano, hubo diferencias significativas entre los tratamientos (Figura 2A). Los discos impregnados con los tres tratamientos de mayor concentración mostraron el menor porcentaje de consumo foliar y fueron semejantes entre sí (p > 0.05). Los dos tratamientos de menor concentración y los testigos (relativo y absoluto) fueron semejantes entre sí (p > 0.05), pero difirieron (p < 0.05) de los tres tratamientos de mayor concentración. Asimismo, al aumentar la concentración, el porcentaje de consumo disminuyó. El modelo que presentó la curva de mejor ajuste fue el potencial (R2= 0.92) (Figura 3A).
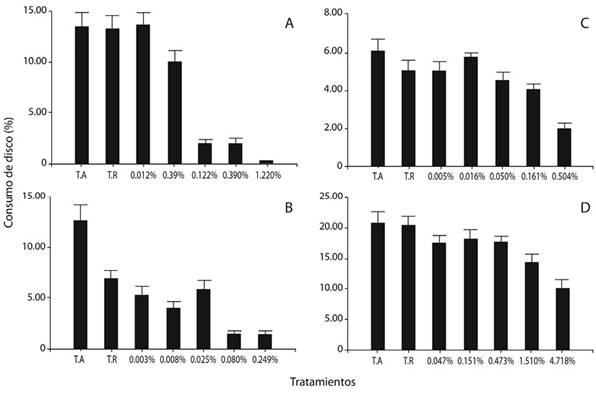
Figura 2. Consumo de discos foliares de cedro tratados con concentraciones crecientes de las particiones de hexano (A), diclorometano (B), acetato de etilo (C) y acuosa (D) de un extracto flores de B. candida, en comparación con los testigos absoluto (T.A.) y relativo (T.R.), al exponer larvas de instar III de H. grandellapor 24 h.
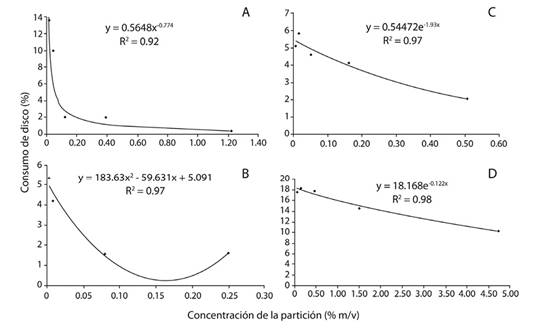
Figura 3 Relación entre el consumo de discos foliares de cedro tratados con concentraciones crecientes de las particiones de hexano (A), diclorometano (B), acetato de etilo (C) y acuosa (D) de un extracto flores de B. candida, al exponer larvas de instar III de H. grandella por 24 h. La línea continua muestra la tendencia esperada (estimada).
La fagodisuasión causada por la partición de hexano fue muy evidente, así como la más alta de los bioensayos específicos con particiones, a una concentración de apenas 0.122 % m/v, con una media de consumo del disco de 2.00 %, en contraste con el consumo mostrado por los testigos absoluto y relativo, que mostraron valores de 13.53 % y 13.25 %, respectivamente. El efecto fagodisuasivo se incrementó hasta alcanzar un mínimo de consumo de 0.35 %, a la mayor concentración.
Con respecto al porcentaje de consumo de los discos impregnados con los tratamientos preparados con la partición de diclorometano, hubo diferencias significativas (Figura 2B). Los testigos absoluto y relativo difirieron entre sí (p < 0.05). El testigo relativo fue semejante a los tres tratamientos de menor concentración (0.003 %; 0.008 % y 0.025 % m/v) (p > 0.05). El tratamiento de concentración de 0.008 % m/v fue similar a los dos tratamientos de mayor concentración que fueron aplicados (0.080 % y 0.249 % m/v) (p > 0.05), así como al tratamiento inferior y al testigo relativo. Además, la curva de mejor ajuste se halló utilizando un modelo polinómico (R2= 0.97), donde se observa una tendencia hacia la disminución del consumo conforme aumenta la concentración (Figura 3B).
Sin embargo, es oportuno resaltar que con dicha partición, los discos foliares sufrieron fitotoxicidad. En efecto, en el momento de la evaluación, una vez transcurridas 24 h después de impregnados los discos, se detectó su deterioro, con síntomas tales como la decoloración de los pigmentos propios de las hojas, así como el arrugamiento y la deshidratación del tejido.
En cuanto al consumo de discos tratados con la partición de acetato de etilo, hubo diferencias significativas entre los tratamientos (Figura 2C). Los testigos, tanto el absoluto como el relativo, así como los cuatro tratamientos de menor concentración, fueron semejantes entre sí (p > 0.05), pero difirieron del tratamiento de mayor concentración (p < 0.05). La curva que aportó mejor ajuste fue la del modelo exponencial (R2= 0.97) (Figura 3C). Se observó que al aumentar la concentración de los tratamientos, el consumo foliar tendió a disminuir.
Finalmente, hubo diferencias significativas (Figura 2D) entre el consumo de los discos impregnados con los tratamientos de la partición acuosa. Ambos testigos fueron similares a los tres tratamientos de menor concentración (p > 0.05), los cuales a su vez fueron semejantes al tratamiento que se aplicó al 1.510 % m/v (p > 0.05); sin embargo, este último no se asemejó a los testigos utilizados (p < 0.05). Los dos tratamientos de mayor concentración utilizados resultaron ser semejantes entre sí (p > 0.05), pero el inferior no difirió de los tratamientos de concentraciones menores a éste. Asimismo, al incrementarse la concentración de la partición se redujo el porcentaje de consumo. La curva que presentó el mejor ajuste fue la que arrojó el modelo exponencial (R2= 0.98) (Figura 3D).
Bioensayo con un patrón de escopolamina: Los resultados de los promedios de consumo foliar mostraron una marcada disminución en presencia del tratamiento que contenía el alcaloide, con valores de 9.08 y 8.00 % para los testigos absoluto (extracto crudo) y relativo (partición de hexano), respectivamente, y de apenas 0.90 % para el tratamiento de escopolamina (Figura 4).
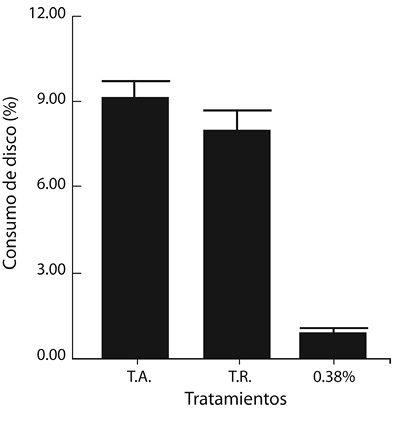
Figura 4. Comparación de los promedios de consumo en discos foliares de cedro impregnados con escopolamina (0.38% m/v) con los testigos absoluto (T.A.) y relativo (T.R.), al exponer larvas de instar III de H. grandella por 24 h.
En términos de mortalidad, no hubo diferencias significativas entre los tratamientos, así como tampoco difirieron del testigo absoluto (F = 0.20, g.l. = 3, p > 0.05). El porcentaje de mortalidad varió entre 25 y 33 % para el testigo absoluto y el extracto crudo al 1.0 % m/v, respectivamente. Todos los tratamientos mostraron un incremento en la mortalidad a partir del sétimo día después de la aplicación de los mismos, pero al completar 48 días después de la exposición, la mortalidad se mantuvo constante en todos los casos.
Por su parte, en cuanto a los efectos sobre el desarrollo, no hubo diferencias significativas entre los tratamientos, ni con el testigo absoluto, en la cantidad de individuos que consiguieron alcanzar el estado de pupa (F = 0.19, g.l. = 3, p > 0.05), ni tampoco en cuanto a los individuos que lograron emerger como adultos (F = 0.34, g.l. = 3, p > 0.05). De todas las pupas obtenidas, apenas dos no lograron convertirse en adultos, y correspondieron al tratamiento de la partición de hexano, pero ello quizá obedeció a otros factores, como su posible manipulación inadecuada. No hubo casos de adultos con malformaciones.
Tamizaje fitoquímico de la flor de B. candida: Los principales grupos de metabolitos secundarios presentes en la flor de B. Candida correspondieron a ácidos grasos, alcaloides, compuestos reductores, cumarinas, esteroles y/o triterpenos, polisacáridos y taninos (Cuadro 1, Apéndice Digital 2).
Identificación de la presencia de escopolamina en el extracto crudo de la flor de B. candida y en su partición más promisoria: El cromatograma mostró un pico con un tiempo de retención (tr) de 18.948 min, y su espectro de masas coincidió con el registrado para dicho compuesto, tanto en la biblioteca del espectrómetro de masas utilizado, como en la base de datos espectral para compuestos orgánicos SDBS (Spectral Database for Organic Compounds).
En cuanto al extracto crudo, el cromatograma mostró una señal con un trde 18.932 min (Figura 5, Apéndice Digital 3), cuyo espectro de masas, obtenido en modo de ión selectivo, mostró un ión molecular (M+) a 303 m/z (Figura 6, Apéndice Digital 3). Asimismo, el cromatograma de la partición de hexano reveló una señal con un trde 18.933 min (Figura 7, Apéndice Digital 3), y su espectro de masas también mostró un ión molecular (M+) a 303 m/z (Figura 8, Apéndice Digital 3). Tanto el extracto crudo como la partición de hexano exhibieron un pico base en 94 m/z.
Discusión
Hubo un claro y definido efecto fagodisuasivo del extracto crudo de la flor de B. candida sobre las larvas de H. grandella, como lo demuestra el hecho de que el consumo de discos foliares de cedro a una concentración de 1.0% m/v no fue significativo en términos estadísticos. Aunque a primera vista este dato podría sugerir que más bien hubo toxicidad, esto se descarta, por cuanto la mortalidad de las larvas no fue significativa una vez que fueron transferidas a dieta artificial para que completaran su desarrollo, además de que tampoco mostraron efectos subletales (disminución del tamaño o del peso, tiempo de desarrollo más prolongado, o malformaciones anatómicas).
El hecho de que H. grandella muestre especificidad o exclusividad por representantes de la subfamilia Swietenioideae (Meliaceae), de la que afecta 13 especies pertenecientes a varios géneros (Becker, 1976), sugiere que algunos principios químicos presentes en miembros de otras familias podrían actuar como disuasivos, repelentes, reguladores del crecimiento o insecticidas de dicho insecto. Sin embargo, en evaluaciones de especies pertenecientes a 16 familias (Hilje & Mora, 2006), se han detectado efectos adversos en apenas tres de ellas, de las familias Cucurbitaceae, Rutaceae y Simaroubaceae. Al respecto, se ha observado fagodisuasión en extractos de Q. amara y R. chalepensis(Mancebo et al.,2000; 2001; Soto et al.,2007;, Barboza et al., 2010a; 2010b), como se indicó previamente, a las cuales se suman los de frutos del tacaco cimarrón, Sechium pittieri(Cogn.) C. Jeffrey (Cucurbitaceae), que son tóxicos para las larvas (Mancebo et al.,2001).
Asimismo, de particular interés son tres especies exóticas de la familia Meliaceae, con las que H. grandella no ha tenido oportunidad de coevolucionar, por ser un insecto de origen neotropical. De ellas, algunos triterpenos (limonoides) presentes en la semilla del árbol de nim (Azadirachta indica A. Juss.) pueden ser tóxicos o reguladores del crecimiento de la larva de H. grandella, dependiendo de cómo se les formule (Mancebo et al. 2002). Asimismo, Pérez-Flores et al. (2012) demostraron la presencia de alcaloides, limonoides y fenoles en dos especies de meliáceas exóticas, los cuales pueden afectar el consumo foliar, el peso de la pupa y la duración del ciclo de vida de H. grandella; dichas especies son el cedro australiano, Toona ciliata M.J. Roem var. australis(F.V. Muell.) C. DC., y la caoba africana, Khaya senegalensis(Desr.) A. Juss.
Aunque se ignora el mecanismo específico de selección del alimento por parte de la larva de H. grandella, en sus maxilas hay receptores con función sensorial, denominados sensillas estilocónicas (Schoonhoven, 1980), lo que permite suponer que son ellas las que permiten detectar y evitar sabores desagradables.
Para retornar a los hallazgos con B. candida, su comparación con otros estudios se complica, por la escasez de investigaciones al respecto. En cuanto al efecto adverso de extractos de plantas del géneroBrugmansiasobre otros insectos, estos incluyen al coleóptero Tribolium castaneum(Abbasipour et al. 2011), plaga de granos almacenados, a la mosca Haematobia irritans(Cruz et al., 2011), de importancia médico-veterinaria, así como a las termitas Copotermes gestroi y Cryptotermes cynocephalus(Tarmadi et al., 2014). En estos casos los estudios se limitaron a evaluar la actividad biológica de los extractos, sin determinar la identidad de los compuestos causantes de dicha actividad.
Al respecto, en los bioensayos específicos, la partición de hexano fue la que causó mayores niveles de fagodisuasión. Bastó una concentración de 0.122 % m/v para que se expresara dicho efecto, con un consumo de apenas 2 %, en tanto que a la mayor concentración (1.220 %) éste fue de 0.35 %. Por tanto, es claro que los metabolitos secundarios presentes en la partición de hexano son los que poseen mayor actividad fagodisuasiva, y que dicha actividad se asemeja a la hallada para el extracto crudo. Este hallazgo coincide con el de Barboza et al.(2010a), al evaluar el efecto de un extracto crudo de ruda y de sus particiones sobre la larva de H. grandella.
Asimismo, al igual que con el extracto crudo, el modelo matemático utilizado para analizar los datos, reveló que a partir de cierta concentración, la curva tiene una tendencia constante y asintótica, lo cual significa que el consumo de los discos no varía significativamente en función de la concentración de la partición de hexano. Desde el punto de vista práctico, esta información podría ser valiosa para la eventual formulación de un producto comercial.
En cuanto a las demás particiones, la acuosa no contiene los metabolitos causantes de la actividad fagodisuasiva, en tanto que con la de acetato de etilo hubo fagodisuasión únicamente a la mayor concentración (0.504 % m/v). Por su parte, con la de diclorometano, es de suponer que la fitotoxicidad sufrida por los discos los tornó en menos atractivos para las larvas; en tales circunstancias, al igual que lo atestiguaron Barboza et al.(2010a), es difícil percibir el verdadero efecto de los compuestos separados en esta partición, no solo por esta fitotoxicidad, sino incluso por algún efecto tóxico que el propio solvente podría ejercer sobre las larvas. En todo caso, no debe desestimarse el aporte que algunas sustancias presentes en ellas pudiesen causar como sinergistas, y aproximarse al efecto causado por el extracto crudo.
A propósito de los solventes empleados en el proceso de extracción, este es un factor importante, ya que determina en gran parte la obtención de compuestos activos deseados, así como el grado de eficacia del proceso extractivo. En tal sentido, el uso de acetona para preparar el extracto crudo se basó en sus propiedades físicas, que son similares a las del metanol o el etanol, convencionalmente empleados en este proceso. Además, hay beneficios adicionales, como no formar azeótropos con el agua, como lo hace el etanol, y su bajo punto de ebullición; esta última propiedad facilita su eliminación en el proceso de concentración del crudo, lo que acorta dicha etapa y disminuye el consumo eléctrico asociado, sin alterar el perfil químico de los metabolitos obtenidos.
Los solventes utilizados en el proceso de particionamiento se distinguieron entre sí por poseer distintas propiedades físicas y químicas, de manera que permitieron la separación de los metabolitos secundarios según la afinidad de sus polaridades con cada uno de los solventes empleados. Durante el particionamiento del extracto crudo, se dieron pérdidas de masa asociadas a múltiples trasvases del material de un contenedor a otro, lo cual es un factor que, de manera inevitable, provoca una disminución en el porcentaje de rendimiento de cada una de las particiones fitoquímicas. Por su parte, la partición de mayor rendimiento fue la acuosa, lo que obedece a la elección de un solvente de naturaleza polar, que favorece mayormente la extracción de compuestos de polaridad semejante.
Ahora bien, el tamizaje fitoquímico de la flor de B. candidareveló la presencia metabolitos secundarios tales como ácidos grasos, alcaloides, compuestos reductores, cumarinas, esteroles y/o triterpenos, polisacáridos y taninos, como se indicó previamente.
De ellos, son de particular importancia los alcaloides, por su abundancia en B. candida, como lo demostraron Arteaga et al. (1993), al igual que en sus congéneres B. arborea, B. aurea y B. sanguinea; en las hojas y flores de estas cuatro especies predominan la escopolamina y la atropina, y la apoescopolamina y el 6-β-hidroxi-3-α-tigloiloxitropano, aparecen en menor cantidad. Por su parte, Otálvaro (2009) documentó que B. candidaes una rica fuente de alcaloides tropánicos, muy utilizados en formulaciones farmacológicas, como la atropina, hiosciamina, anisodamina y escopolamina. Por su parte, Eich (2008) indica que la escopolamina puede actuar como una neurotoxina, al atacar el receptor de acetilcolina.
Se ha descubierto que algunos alcaloides no poseen actividad farmacológica en mamíferos, y que varios de ellos son tóxicos o fagodisuasivos para insectos (Buchanan et al., 2000). Asimismo, Alves et al.(2007) observaron que, al ser atacadas por herbívoros, varias especies de Brugmansia incrementan la producción de alcaloides en el área afectada, lo que sugiere que actúan como compuestos defensivos; dichos autores también reportaron que las larvas del lepidóptero Spodoptera frugiperda(Noctuidae) sufrieron inhibición en su desarrollo, e inclusive la muerte, al consumir hojas de B. suaveolens.
En la presente investigación, las pruebas para alcaloides permitieron detectarlos únicamente en la partición de hexano. Este resultado no necesariamente demuestra su ausencia en los extractos etanólico y acuoso, ya que es posible que la concentración de alcaloides en ellos haya sido baja por lo que podrían haber pasado desapercibidos. Al respecto, a pesar de que los alcaloides en forma de base son poco solubles en agua (Arango, 2008), en la naturaleza los derivados del tropano se encuentran mayormente en forma de sales (Bruneton, 2001).
Asimismo, debe considerarse que la composición fitoquímica de las plantas varía dependiendo de múltiples factores, como el tipo de suelo, la época del año y el clima, así como de otros tan sutiles como la hora de recolección de la muestra. Estas variables pueden influir no solo en la composición química de la planta, sino que también en la distribución de los metabolitos en las distintas partes de la misma y la forma en que éstos se encuentran (como base, o bien como la sal correspondiente) dentro de ella, en determinado momento.
Además, resultó muy reveladora la presencia de alcaloides en la partición de hexano, puesto que en los bioensayos generales, dicha partición mostró la mayor actividad fagodisuasiva contra H. grandella. Esto sugiere fuertemente que este grupo de metabolitos tiene relación con tal actividad, pero la naturaleza de esta investigación no permite determinar si se debe especialmente a la presencia de alcaloides o a una acción sinérgica con otras familias de compuestos también presentes en la partición de hexano.
Al respecto, cabe destacar que en algunas familias detectadas mediante el tamizaje fitoquímico hay compuestos con funciones defensivas contra insectos, incluyendo algunas cumarinas, triterpenos y taninos (Buchanan et al., 2000). Pero para definir si la actividad detectada obedece sobre todo a la acción de esta familia de sustancias, es necesario realizar una evaluación más exhaustiva, la cual involucra procesos de fraccionamiento más finos. Aún así, el tamizaje fitoquímico realizado marca el inicio que permitiría orientar dichos procesos.
En todo caso, a propósito de alcaloides, fue posible confirmar la presencia de la escopolamina en el extracto crudo, así como en la partición de hexano. El ión molecular encontrado en ambos espectros de masas coincidió con la masa molecular de la escopolamina. Además, los iones característicos seleccionados en el modo de ión selectivo para el espectro de masas del extracto crudo, conservan la misma relación de intensidad que los que se hallaron en el espectro de masas de la partición de hexano. Esto indica que, durante el proceso de descomposición de las moléculas irradiadas, las fracciones más estables son las mismas en ambas muestras. Lo anterior valida que dichos fragmentos corresponden a la desintegración de la misma molécula.
Asimismo, el patrón de fraccionamiento hallado en ambos análisis mostró señales características que coinciden con el que se ha observado para la escopolamina. En ambas muestras se encontraron señales en 154 m/z, la cual corresponde a la fragmentación de la molécula en su enlace éster, al perder la unidad C9H9O2. Otra señal que destaca es la que se encuentra en 138 m/z, resultante de la pérdida de una unidad C9H9O3, que corresponde al ácido trópico. Ambos picos corresponden a pérdidas de fragmentación características de la escopolamina (Wang et al., 2012).
Es pertinente mencionar que Stashenko y Martínez (2012) determinaron la presencia de escopolamina en muestras de extractos obtenidos mediante fluidos supercríticos (CO2) provenientes de flores de B. suaveolens, y comprobaron que los resultados de sensibilidad obtenidos en este modo de detección pueden sobrepasar en hasta 100 veces la sensibilidad obtenida regularmente.
Ahora bien, los resultados obtenidos en la presente investigación con la escopolamina (0.38 % m/v) como fagodisuasivo, fueron muy claros y definidos, pues redujo en 90 % el consumo de discos foliares, en comparación con los testigos. Este hallazgo, además de aportar nuevo conocimiento en cuanto a la actividad de los alcaloides específicos en insectos, tiene gran valor práctico, pues la escopolamina, en combinación con coadyuvantes apropiados, podría ser formulada como un fagodisuasivo comercial.
Sin embargo, obviamente, antes sería necesario determinar la dosis mínima del alcaloide que debería contener el producto formulado, así como el posible efecto sinergista de algunos coadyuvantes, ya sea de otro origen, o de otras familias de compuestos también presentes en la flor de B. candida. Al respecto, es preciso indicar que aunque conocer de manera particular los distintos grupos funcionales causantes de fagodisuasión sería valioso, tanto en términos científicos como prácticos, se ha observado que al evaluar compuestos fagodisuasivos de manera individual, en la misma concentración en la que se encuentran en la planta, éstos tienden a ser menos activos que al ser aplicados de forma conjunta (Warthen, 1990).

