











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkLike other invertebrates, sea urchins have an innate immune system. In this system, coelomocytes are fundamental in the innate immune response, acting as the main immune effectors. The coelomocytes mediate immune responses by phagocytosis and encapsulation of foreign particles, in conjunction with the release of antimicrobial molecules by degranulation (Smith et al., 2006).
In echinoderms, the classification of coelomocytes is based essentially on morphological criteria, commonly distinguishing four categories: phagocytes, vibratile cells, red spherule cells, and colourless spherule cells (Smith et al., 2010). The proportions of each type of cell in the celomic cavity may change considerably and this variability could be due to the nutritional, immune and homeostatic status of the individuals surveyed. However, phagocytes constitute the largest proportion of coelomocytes who are involved in graft rejection, chemotaxis, phagocytosis, encapsulation, immune gene expression, agglutination reactions and aggregation (Smith et al., 2010).
Seawater temperature is an important factor controlling the survival of marine invertebrates. Rise in temperatures could increase their susceptibility to pathogens; hence the emergence of different diseases. There is evidence of an increase in the frequency of epidemics as a result of heat stress in different ecosystems (Harvell et al., 1999). By studying the immune response in the Antarctic sea urchin (Sterechinus neumayeri) when exposed to increased seawater temperatures, it could give us an indication of the potential impacts that global warming could have on the immune mechanisms of benthic invertebrates and their degree of susceptibility to pathogens. Coelomocytes are known to be the key cells involved in defensive responses of the Antarctic sea urchin Sterechinus neumayeri (Borges et al., 2002; Branco et al., 2012) However, how the echinoderms immune system respond to pathogen aggression, tissue injury or stress conditions, still have not been addressed in Antarctic echinoderms. The information regarding immune mechanisms involved in Antarctic marine invertebrates is also limited. The few studies that do exist have been carried out mainly in the context of the inflammatory response and phagocytosis at low temperatures (Silva et al., 1998, 2001; Silva & Peck, 2000). For example, Borges et al. (2002) demonstrated different types of coelomocytes and their ability as phagocytes (S. neumayeri). A second study proposed different cellular biomarkers, to better comprehend the thermal stress effects on immune parameters of this Antarctic sea urchin (Branco et al., 2012). However, a study focusing on the immune parameters of this echinoid challenged with lipopolysaccharides is still lacking in the current literature.
In the last few years, the studies on the echinoderms immune system had taken a new perspective, related to the genomic data produced after sequencing the purple sea urchin genome. One of the most characteristic features of the sea urchin genome is that it possesses a large number and diversity of innate immunity genes, mainly at the receptor level, and is unique when compared with other species (Sea Urchin Genome Sequencing Consortium, 2006). The vast repertoire of receptors, such as Toll receptors, involved in the processes of pathogen recognition, is remarkable. A complementary system and transcription factors in immunity and hematopoiesis has also been identified, which is a novel finding for this echinoderm (Rast et al., 2006).
Insight into the immune response of Antarctic organisms has been poorly studied; we have little information about molecular processes related to immune responses at polar temperatures and gene patterns in Antarctic marine invertebrates subjected to a bacterial challenge. The current study reports coelomocyte characterization, phagocytic capacity and how these cellular parameters could be modified by environmental factors (Silva & Peck, 2000; Borges et al., 2002, 2010; Branco et al., 2012). In this study we primarily characterized the cellular and molecular response of coelomocytes in S. neumayeri, to exposure against a specific pattern of pathogens like lipopolysaccharides (LPS) or bacteria. In addition, we evaluated how this response could be modified by acute thermal stress.
Material and metthods
Sea urchin collection and maintenance
Adult males and females (n=110) of Antarctic sea urchin, were collected by SCUBA divers from depths of 6 to 8 m from Maxwell Bay (62°12’12.2” S - 58°56’41.7” W; King George Island, Antarctica) during the summer of 2010 and 2011. Sea urchins were kept in seawater at 1 °C in a cold chamber at the chilean Antarctic Scientific Base “Profesor Julio Escudero”. Specimens were acclimated for one week before experiments.
Coelomocytes extraction
Coelomic fluid was collected via the peristomial membrane with a needle 12.7 x 0.33 mm and a syringe of 1 ml. Coelomocytes were counted in a Neubauer chamber in order to obtain a total and differential coelomocyte count. Cells were fixed in 4 % of paraformaldehye in Filter Sea Water (FSW) and conserved in a RNA later (Ambion).
Immune challenge
Sea urchins were stimulated by injection of lipopolysaccharides (LPS) from Escherichia coli. Experimental challenges using 100 μg LPS of E. coli J5 026:B6 (Sigma) were conducted with a group of sea urchins (18 animals) to stimulate an immune response. A control group in an independent aquarium was injected with seawater and the coelomic fluid was collected via peristomial membrane; the fluid without dilution was placed in a Neubauer chamber for the counting of absolute and relative coelomocyte types. Coelomic fluid was collected after 0, 2, 6, 24, 36 and 48 h of exposure. The cells were immediately centrifuged at 700 g for 10 min (4°C) to separate the coelomocytes from coelomic liquid. The experiments were done using three animals for each condition and time.
Phagocytosis assay and Phagocytic Indexes
The phagocytosis assay was conducted by placing 150 μl of coelomic fluid on a glass slide for cell spreading. After one hour, a yeast Saccharomyces cerevisae suspension was added to the glass slide in a proportion of 10 yeasts per phagocyte. This suspension was prepared diluting approximately 100 µg of lyophilized yeast in 10mL of filtered marine seawater, resulting in a concentration of approximately 3 x 106 yeast cell/ml. Then this suspension was homogenized and stored at 4 °C. The glass slides were incubated for to 2 h in the same experimental temperatures which sea urchins were exposed in order to obtain phagocytic indexes. Phagocytic indexes were used according to Silva and Peck (2000) and were calculated based on the following:
Phagocytic capacity: PC = Number of phagocytic amoebocyte phagocytosing / Total number of phagocytic amoebocyte
Phagocytic index: PI = Total number of phagocytosed yeast/Number of phagocytic amoebocyte phagocytosing
Statistical analysis was executed using ANOVA and Tukey’s test (GraphPad Software Inc.). Statistical difference was considered significant when p < 0.05 and p < 0.01.
Acute thermal stress
The Antarctic sea urchins were exposed to 5 °C and 10 °C for 48 h. Twenty animals by each acute thermal stress treatments were directly transferred from control temperature (1 °C ± 0.5 °C) to two aquarium for acute thermal stress without an acclimation period at 5°C and 10°C. Ten animals composed the control group. Four sea urchins were randomly collected from each group at 1, 24 and 24 h post thermal stress, respectively. Coelomocyte samples were obtained as described above and stored in RNA later (Ambion).
RNA extraction, cDNA synthesis, PCR and cloning
Total RNA was extracted from S. neumayeri from coelomocytes using a Trizol reagent (Invitrogen) and treated after with DNAse Turbo (Ambion) according to the manufacturers instructions. Then, the quantity and the integrity of the total RNA were checked by spectrophotometric and agarose gel electrophoresis, respectively. 1 µg of the total RNA was reverse transcript, using M-MLV reverse transcription kits (Invitrogen) according to the manufacturers’ instructions in a final volume of 20 µl. cDNA has served as a matrix in PCR reactions on selected genes to isolate them or semi quantify their expressions. The cDNA was amplified by PCR, using 1 U of Taq polymerase (Invitrogen) and 1 µM of each primer (see Table 1), at a final volume of 25 µl. An amplification program for PCR consisted in 5 min at 94 °C, followed by 30 cycles of 94 °C for 45 s, 55 °C for 45 s, and 72 °C for 1 min, and finally an elongation step at 72 °C for 10 min. Amplified products were analyzed on 1% agarose gels, cloned into PCR 2.1 TOPO TA cloning vector (Invitrogen), and sequenced from both directions with T7 and T3 primers.
Table 1 Primers used in real-time PCR expression analysis
| Targeted genes | Sense | Primer sequences, 5’-3’ | Amplicon size (bp) |
|---|---|---|---|
| Metallothionein (Y08621) | Forward Reverse | GACGTGCTCTAAGCCCAGAC TCACAATGAGACCCACCGTA | 300 |
| Allograft inflammatory factor 1 (FJ824731) | Forward Reverse | ACGGAAGTGGAACCATCAAC CAGATGTGGAGCAGCGTAAA | 200 |
| LBP/BPI (JQ736681) | Forward Reverse | CTGCCCAACTCTGCCGGTTA CATCGCCATGTTCGGGTACA | 165 |
| 28S (KC218476) | Forward Reverse | GGGTATAGGGGCGAAAGACT GTCGGGCCTCTTACCAATTT | 174 |
| 18S | Forward Reverse | GAGCCTGCGCTTAATTTGAC GGCGCAACTATTTAGCAAGC | 200 |
Quantitative PCR analysis
Quantitative PCR (qPCR) analysis was done to determine whether there were acute changes in LBP/ BPI, AIF-1 and MT gene expression, sampled at 2, 6, 24, 36 and 48 h post-stimulation. The primers Forward and Reverse were used to amplify the immune related genes (Table 1). These qPCR reactions were composed by 1.0 µl of cDNA diluted at 1:10 ratio, 0.5 µl of both primers (5 µM) adding 25 µl of a ready to use solution of SYBR Green PCR Master Mix (Applied Biosystems) into a final volume of 50 µl complete with water. Amplification conditions, performed in ABI Thermocycler 7 500, consisted in 40 cycles, 94 ºC for 10 min; 94º C for 1 min; 55° C for 1 min; 72 °C for 1 min with a single fluorescence measurement; melting curve program and finally a cooling step. For further analysis of the expression level, the crossing points (CP) were determined for each transcript, using the 7 500 program version 2.0.1 (Applied Biosystems). Specificity of a qPCR product was determined by agarose gel electrophoresis and melting curve analysis. The copy ratio of each analyzed cDNA was determined as the mean of three technical replicates. The relative expression level of Sn-AIF-1 was calculated based on the 2-ΔΔCP method using the average of 28 S / 18 S as the reference gene (Livak & Schmittgen, 2001). Three independent experiments were performed, on a pool of three sea urchins from both defined groups and with the same timing. Data was subjected to ANOVA-test and followed by post-hoc Tukey tests (p < 0.05) using STATISTICA software.
Oxygen consumption during bacterial challenge
Twenty-four animals were used for the evaluation of the effect of bacterial inoculation on the metabolism of Sterechinus neumayeri. The experiment comprises two treatments (Challenge and Control group); six replicates per treatment. Two animals were incubated together in 1 000-ml hermetic chambers; at 0.5 °C (0.45 µm filtered seawater, Fildes Bay, King George Island, Antarctica). Each chamber was conssidered a replicate. Animals were starved 24 h before oxygen consumption determinations.
The microorganism used for the challenge was the heat-killed bacterial strain Vibrio anguillarum (ATCC), by injecting a mixture of 200 μl per urchin (1 x 106 bacteria / ml) into the coelomic cavity. Oxygen content was quantified before and after incubation (ca. 1.4 h, Control = 1.5 h; challenge group = 1.2 h) by an optic sensor connected to a Microx MX3 AOT oxygen meter (PreSens, GmbH, Germany), previously calibrated with saturated seawater (100 %) and 5 % sodium sulfite solution (0 %). Chambers were inverted carefully 3 times at the end of each trial to ensure adequate mixing. Six chambers without animals were used as a control group per treatment. Afterwards, animals were measured: test diameter, nearest 1 mm with a caliper; animal volume, to the nearest 5 ml with a graduated cylinder; dry mass (DM), nearest 10 mg after drying in a Memmert oven at 60°C for 48 h. This data was used for correcting water volume during the challenge period and for DM calculation for a weight-specific oxygen consumption estimation. Oxygen consumption rate was expressed in units of μg O2 h-1 g-1DM. Student t-test was performed for the comparison between Challenge and Control treatments since data was normaly distributed (Kolmogorov-Smirnov test) and variances were homogeneous (Bartelet test).
Results
A significant increase in number per ml of phagocytic amoebocyte (PA) and red spherule cells (RSC) in coelomocytes was found in sea urchins exposed to LPS for 2 h (p < 0.001). Furthermore, a significant increase in the total number of cells (p < 0.001) was observed in sea urchins induced by LPS in the same 2 h. The colorless sphere cells (CSC) increased their number after 48 h (p < 0.05) (Table 2).
Table 2 Effect of LPS injection in vitro in the number of coelomocytes and phagocytic indexes of Sterechinus neumayeri challenged at different periods of time
| Parameters | PA/ml (x 10 5 ) | CSC/ml (x 10 5 ) | RSC/ml (x 10 5 ) | VC/ml (x 10 5 ) | Total coelomocytes/ml (x 10 5 ) | PC (%) | PI | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Period afther injection (hours) | Control | LPS | Control | LPS | Control | LPS | Control | LPS | Control | LPS | Control | LPS | Control | LPS |
| 2 | 10.6±1.4 | 20.0±2.6** | 3.7±1.2 | 4.3±1.2 | 1.4±0.4 | 4.8±0.4** | 2.3±0.9 | 3.6±0.8 | 18.0±0.7 | 32.7±3.8** | 17.73±3.78 | 41.44±1.78** | 1.27±0.10 | 1.15±0.12 |
| 6 | 14.7±1.4 | 17.6±1.4 | 1.8±0.8 | 3.0±0.7 | 1.8±0.2 | 1.2±0.5 | 3.4±1.2 | 2.4±0.4 | 21.7±1.6 | 24.2±3.0 | 16.88±4.67 | 36.39±7.58** | 1.32±0.31 | 1.73±0.11 |
| 24 | 5.0±0.4 | 6.2±0.3 | 1.0±0.1 | 1.0±0.4 | 0.3±0.01 | 0.2±0.03 | 1.5±0.03 | 1.0±0.03 | 7.8±0.1 | 8.4±0.8 | 21.98±3.65 | 24.50±5.87 | 1.29±0.20 | 1.26±0.15 |
| 36 | 9.8±0.9 | 8.8±1.4 | 1.6±0.7 | 1.6±0.4 | 0.6±0.2 | 0.2±0.2 | 2.1±0.6 | 1.6±0.6 | 14.1±2.1 | 12.2±1.0 | 16.46±3.24 | 17.20±3.03 | 1.57±0.43 | 1.27±0.26 |
| 48 | 15.6±2.9 | 14.1±1.4 | 0.8±0.4 | 2.6±0.4* | 1.7±0.7 | 1.3±0.6 | 2.7±1.2 | 2.3±0.5 | 20.8±2.6 | 20.3±2.1 | 17.20±4.38 | 18.54±3.03 | 1.51±038 | 1.49±0.21 |
Values quoted are mean ± SD
PA Phagocytic Amoebocyte, CSC Colorless Sphere Cell, RSC Red Spherule Cell, VC Vibratile Cell, PC Phagocytic Capacity, PI Phagocytic Index
*Refers to statical defference between control and LPS challenged group.
*(P <0.05), **(P < 0.001)
Regarding phagocytic indexes, the only significant difference was an increase in Phagocytic capacity (PC) to 41.44 % (SD = 1.78), which was significantly higher than the control group value at 17.73 % (SD = 3.78), after 2 h of LPS injection and to 36.39 % (SD = 7.58) when compared to the control group at 16.88 % (SD = 4.67) after 6 h of LPS injection (Table 1).
Real time PCR was used to evaluate the expression of three transcripts in coelomocytes of the Antarctic sea urchin exposed to LPS from E. coli. The mRNA transcript of these genes could be detected in coelomocytes of control and challenged animals. The injection of LPS induces the expression of MT, AIF-1 and LPB/BPI genes. The injection of LPS produced a rapid increase level at 2 h of the MT transcript (Fig. 1). MT showed significantly higher gene expression levels at all times when compared to the control condition (p < 0.05). AIF-1 exhibited significantly higher expressions at 24 and 36 h (p < 0.05), but significantly lower gene expression (p < 0.05) at 6h. The expression after 48 h dropped close to the control levels (Fig. 1) Finally, the expression of LBP/BPI exhibited a significant increase at 2 and 6 h after the LPS challenge (p < 0.05). After 24 h, the LBP/BPI expression showed no significant variation with respect to the control group (Fig. 1).
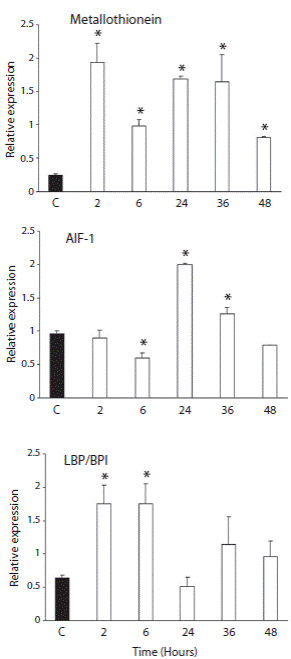
Fig. 1 Variation of gene expression during LPS challenge in Sterechinus neumayeri. The Antarctic sea urchins were exposed to 100 μg LPS of E. coli J5 026:B6 for 2, 6, 24, 36 and 48 h. Bar represent the mean of three independent experiments and error bars show SD. *p < 0.05. The amplicon obtained after qPCR were: 300 pb (MT); 200 pb (AIF-1) and 165 pb (LBP/BPI).
Acute thermal stress produced changes in profile gene expression in these same genes. MT increased their expression in coelomocytes from sea urchins exposed at 5 °C after 24 h (p < 0.05) and this pattern was similar when the animals were exposed at 10 °C during 48 h (Fig. 2). MT exhibited significantly higher gene expression levels at 24 h (p < 0.05). The expression of genes involved in the immune response (AIF-1 and LBP/BPI) showed a similar tendency to MT at 5 °C, because their expressions increase. A significant variation in their expression at 24 h by AIF-1 was shown (Fig. 2). On the other hand, LBP/BPI gene expression levels at 1 and 48 h were even significantly higher than in the control animals. However, the expression of AIF-1 showed lower gene expression levels at 1 and 48 h (p < 0.05). AIF-1 mRNA expression decreased significantly over an exposure time of 24 and 48 h to acute thermal stress at 10 °C. In the same way, the expression level of LBP/BPI was affected during thermal stress. The first hours during thermal stress the LBP/BPI gene expression increased significantly, but at 24 and 48 h the LBP/BPI expression did not show a significant difference in comparison to the control condition (Fig. 2). However, 48 h of exposure at 10 °C could produce a depression in these genes.
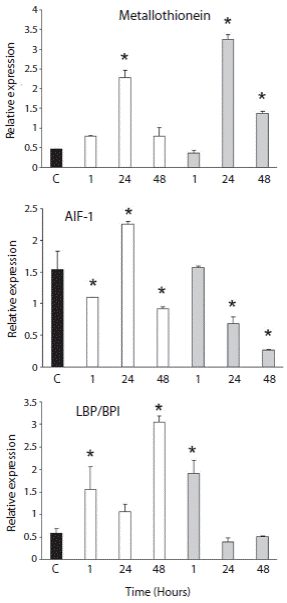
Fig. 2 Variation of gene expression during acute thermal stress in Sterechinus neumayeri. The Antarctic sea urchins were exposed to 5 °C and 10 °C for 1, 24 and 48 h. Black bars: control group; white bars: 5 °C and grey bars: 10 °C. Bar represent the mean of three independent experiments and error bars show SD. *p < 0.05. The amplicon obtained after qPCR were: 300 pb (MT); 200 pb (AIF-1) and 165 pb (LBP/BPI).
No difference was observed in sea urchin size, neither among experimental treatments or in inoculation experiments (mean 58.99 ± 3.83 mm test diameter Student t-test, t = 0.888; P = 0.395). Oxygen depletion during incubation remained between 4.5 and 10.8% of initial oxygen concentration, ensuring that no hypoxic conditions occurred during respiration rate quantifications. Inoculated individuals showed higher respiration rates after 1.2 h of inoculation, compared to the control group (17.32 ± 4.73 μg O2 h-1 g-1 and 11.16 ± 3.93 μg O2 h-1 g-1, respectively; Student t-test t = - 2.367; P = 0.042) (Fig. 3).
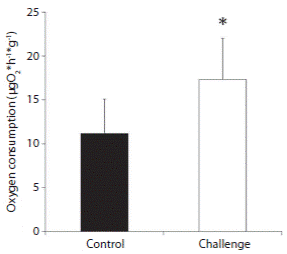
Discussion
The significant increase in the absolute number of red sphere cells observed in the acute challenge with LPS (2 h) demonstrated that such substance is capable of inducing an innate immune response in S. neumayeri. This LPS-inducted cell population seems to be involved in cellular stress, as previously demonstrated by Matranga et al. (2000), Pinsino et al. (2008), Borges et al. (2010) and Branco et al. (2012). In other previous studies of echinoderms S. purpuratus and Asterias rubens, bacterial challenge and LPS produced an increase in the number of coelomocytes after injection (Clow et al., 2004; Holm et al., 2008).
A red sphere cell is a cell type among the four different types of coelomocytes found in sea urchins. Its percentage in the coelomic fluid varies considerably among species. Smith et al. (2006) reported a range varying from 7 to 40 % of this cell type in physiological conditions. These are round shaped cells, containing red granules in their cytoplasm and an amoeboid manner of migration (Smith et al., 2010). Inside their granules, there is a substance named Echinochrome A. It is believed that such substance is released into the coelomic fluid after cell degranulation, which in turn happens after being stimulated with bacteria. This molecule has antimicrobial properties against Gram positive and negative bacteria (Gerardi et al., 1990; Haug et al., 2002). Red spherule cells express SpNFkB (Smith et al., 2006), a transcription factor described in the sea urchin Strongylocentrotus purpuratus, which seems to be only detected after coelomocytes are activated by LPS.
Sea urchins stimulated by LPS after being challenged for 2 h, demonstrated an increase in the number of phagocytic amoebocytes which coincides with a higher phagocytic capacity for the same period. Phagocytic amoebocytes are the most abundant cell type and the one involved in phagocytosis (Bertheusen & Seljelid, 1978). Similar to red sphere cell, phagocytic amoebocytes also express SpNFkB, as well as other immune genes, such as complement homologues and a C-type lectin. All factors are only expressed after being challenged (Smith et al., 2006).
A LPS injection in sea urchins induced an increase in mRNA of profilin and a complement component C3 in coelomocytes (Smith et al., 2006). Profilin is an actin-binding protein that mediates signal transduction involved in cytoskeleton modifications, assisting the correct actin filament polymerization. Such cytoskeleton rearrangement is mandatory for the pseudopodia extension, a fundamental step in the phagocytic response (Smith et al., 2010). It has been suggested that the C3 complement might be involved in augmented phagocytosis. The increased Phagocytic Capacity observed 2 h after LPS injection, can be explained by an immediate upregulation in the synthesis of profilin or complement molecules.
Sea urchin coelomocytes produce a typical gene activation response in the presence of bacteria, allogenic stimulation, and injury (Smith et al., 2010). Previous studies using LPS to activate the immune system have been done to determine changes in gene expression in sea urchin coelomocytes (Nair et al., 2005; Smith et al., 2006). After injecting LPS, S. purpuratus expresses a vast set of family genes related to an immune response, as do other marine invertebrates. Surprisingly, the purple sea urchin (S. purpuratus) also expresses a high number of these genes, such as 185 / 333, toll-like receptors, leucine-rich repeat-containing proteins, and multiple scavenger receptor cysteine-rich (Rast et al., 2006; Terwilliger et al., 2007).
The induction of MT, AIF-1 and LBP/ BPI mRNAs by LPS in coelomocytes could indicate an increase in immune related gene expression. The higher RNA expression of MT shows that this gene could be involved in the immune response. In general MT, a cysteine-rich stress response protein, has been shown to play numerous roles in the cell: it serves as a reservoir of essential heavy metals, it scavenges free radicals, and it can sequester heavy metals. Scudiero et al. in 1997 characterized for the first time the cDNA of MT from S. neumayeri gonad tissue. Normally, this gene is over expressed when the cells or tissues are exposed to a stress response. During the inflammatory reaction produced by LPS, this protein can be induced by several mediators and participate in cytoprotection (Inoue et al., 2009). The phagocytosis of yeast by coelomocytes could produce an oxidative stress by ROS production and LPS could trigger a similar situation, the expression of MT could prevent oxidative damage in coelomocytes and tissues. Further studies about the role of MT in immune cells could be made to understand their participation in modulating immune responses.
During a microbial challenge, coelomocytes could display great cellular activity, such as chemotaxis or proliferation. They migrate rapidly toward injured sites to discharge their molecular effectors content or to display phagocytosis. Recently, AIF-1 from S. neumayeri coelomocytes was cloned and characterized their expression (Ovando et al., 2012). The bacterial challenge like to LPS produces an increase of gene expression after 24h and was correlated with an increase of total coelomocyte numbers. During this bacterial stimulation, we also observed an increase of the total number of coelomocytes, which is in accordance with previous studies of echinoderms S. purpuratus and Asterias rubens, in which bacteria and LPS produced an increase in the number of coelomocytes after injection (Clow et al., 2004; Holm et al., 2008). In the case of the S. purpuratus, LPS produce an increase of the gene expression related to cell proliferation in coelomocytes. Express sequence tags related to proliferation or apoptosis were identified in three of these isolated sequences (Nair et al., 2005). Recently, in the mussel Mytilus galloprovincialis, the expression of AIF was overexpressed by Vibrio splendidus in association with several other genes implicated in cell proliferation (Venier et al., 2011). The overexpression of mammals AIF-1 promotes cell activation, proliferation, and survival of different cells implicated in the inflammatory reaction (Yang et al., 2005; Tian et al., 2009).
In invertebrates, LPS binding proteins (LBP) participate in the transduction of cellular signals from LPS and LBPs which have been characterized in several invertebrates (Gonzalez et al., 2007). In mammals, LBP is an acute phase plasma protein constitutively secreted by the liver that induces cellular responses (Thomas et al., 2002). Instead, the bactericidal/permeability-increasing protein (BPI) contributes to the elimination of Gram-negative bacteria by increasing the permeability of the bacterial membranes (Krasity et al., 2011). BPI is present from humans to invertebrates, and in the oyster (C. gigas) the functionality of this protein was demonstrated as an antibacterial molecule (Gonzalez et al., 2007). In S. purpuratus, three sequences have been obtained, that were assigned as Gram-negative bacterial proteins (GNBP) (Rast et al., 2006), even if in reality these sequences belong to the LBP/ BPI family proteins. This is the first report of gene expression of the LBP/BPI protein in a sea urchin after being challenged with LPS. In coelomocytes of S. neumayeri, the LBP/BPI gene was expressed in unchallenged Antarctic sea urchins, and its transcription appeared up regulated in response to a bacterial challenge to reach a maximum at 2 and 6 h. If LBP/BPI could be accumulated extracellularly, this protein could opsonize bacteria allowing enhances in phagocytosis by coelomocytes that occur in other animal models (Weiss, 2003). Further studies would be needed in functional genomics about AIF-1 and LPB/BPI of related echinoderms to demonstrate the function in cell proliferation and antibacterial activities.
During the thermal stress experiments, the increase in temperature produces in the Antarctic sea urchins a differential response related to three genes studies. The increase of MT has been observed in other marine organisms after thermal stress (Van Cleef-Toedt et al., 2001; Serafim et al., 2002; Piano et al., 2004). This suggests that a direct temperature effect on the MT gene expression must be taken into account, which would imply the presence of a heat-inducible element on the MT-gene promoters, similar to what happens in yeast (Van Cleef-Toedt et al., 2001). In S. neumayeri, heat and oxidative stress will possibly induce the gene expression by activation of the MT gene transcription. Similarly, at 5 °C, the expression gene of AIF-1 and LBP/BPI increase. We have little information about their promoter sequence; therefore more investigations are necessary in this direction. However, at 10 °C we observe a metabolic depression similar to that occurred in other organisms, as a response to stress conditions when heat triggers the decrease in mRNA expression in order to spare energy by fall the transcription and translates processes (Boutet et al., 2009). After higher temperature conditions the animals try to limit their high metabolic cost reaction, such us when immune effectors syntheses consume more ATP (Ardia et al., 2012).
Brockington and Peck (2001) for a standardized 2.3 g individual of S. neumayerii, reported a respiration rate of 1.44 µmol O2 h-1 ind-1 during summer. Our values, converted to the same units as reported by Brockington and Peck (2001), but at 0.5 °C, correspond to 0.99 µmol O2 h-1 ind-1, representing comparable responses. Low metabolic levels in Antarctic invertebrates have been observed (Clarke, 1991) and the studies with sea urchins showed that different factors could change their metabolism, such as the size of the animals or their reproductive state (Brockington & Peck, 2001). Because the immune response is a characteristic ATP-consuming process, we may show that the phagocytes of the innate immune system increase their consumption of O2 during phagocytosis of bacteria or high proliferation rates among coelomocyte populations. Our preliminary results suggest that the turning on the immune response during a short time demands a high energetic cost that we observed after a bacterial challenge. Additional research will be needed about gene expression in other tissues, including the anti-oxidant genes. Furthermore, the metabolic measurement, during a long period of bacterial challenge time to estimate the metabolic cost of immune response al low temperatures.

