











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkSea urchins of the genus Diadema play perhaps the most important role in comparison to other herbivores inhabiting benthic communities, coral reefs, and tropical rocky shores in the world (Lessios, 1981; Carpenter, 1986; Hernández, 2006). Diadema mexicanum is an important algal consumer and similar to the rest of the species of this genus, maintains interaction with the coral community, which can be positive as a controller of algal growth (Edmunds & Carpenter, 2001; Lessios, kessing & pearse, 2001; Atrill & Kelmo, 2007), or negative as a bioeroder agent (Eakin, 1996, 2001; Herrera-Escalante, López-Pérez & Leyte-Morales, 2005). Diadema mexicanum is widely distributed along the tropical eastern Pacific, from the Gulf of California in Mexico (Caso, 1979; Cintra-Buenrostro & Reyes-Bonilla, 1998; Holguin-Quiñónez, Wrigth-López, & Solís-Marín, 2000) to the Lobos de Afuera Island in Peru (Hooker, SolísMarín, & Lleellish, 2005), including Clipperton Islands (Glynn, Vero, & Wellington, 1996), Archipelago of Revillagigedo (Reyes-Bonilla, 1995), del Coco and del Caño Islands in Costa Rica (Guzmán, 1988; Alvarado & Fernández, 2005; Alvarado & Chiriboga, 2008),Galapagos (Hickman, 1998; Edgar, Banks, Fariña & Martínez, 2004) in Ecuador, and Malpelo and Gorgona Islands in Colombia (Neira & Cantera, 2005).
Ecological studies of D. mexicanum have focused almost exclusively on population densities and the effect of the species as bioeroders of the Pacific reefs. Despite its wide distribution and ecological relevance, studies regarding population ecology of the species are scarce in Mexican coasts (Espino-Barr, Cibrián & García-Boa, 1996; Zamorano-de Haro, 2004). In our study, we analyzed population features such as density, abundance and, for the first time ever for this species, natural mortality and distribution pattern in two reef communities of the coast of Oaxaca from April 2008 to March 2009. This characterization of the two populations, besides being an important source of new data, becomes essential since a mass mortality event occurred in May 2009 (Benítez-Villalobos, Díaz-Martínez & Martínez-García 2009), which reduced the local population of this sea urchin to almost a complete disappearance in La Entrega, and had a lesser effect in other localities of the area. Therefore, the data we present in this work will be useful as a reference of the state of the populations just before the die-off and will serve to compare and evaluate the population dynamics as well as the possible recovery after the event.
Material and methods
Field work
The study area comprises the reef communities of La Entrega (15° 44’ 34” N - 96° 07’ 35” W) and Isla Montosa (15° 45’ 48” N - 96° 04’ 56” W), which belong to the reef complex of Bahias de Huatulco in the southern Mexican Pacific.
Field work was carried out monthly from April 2008 to March 2009. Ten 2 - m wide and 10 - m long belt transects were haphazardly laid over the reef and all the urchins that were included within every quadrant of 1 X 2 m along the transect were counted (CARICOMP, 2001). In every fieldtrip, environmental variables (pH, salinity) were recorded using a multiparameter metering device (Hannah HI9828) with a precision of ± 0.01 for both parameters. Bottom temperature was recorded every hour using Hobo Pendant Loggers (Onset Computer Corporation) permanently attached to the sea floor. Pluvial precipitation data were obtained from the meteorological station 0X01 of the National Water Commission, located in Puerto Angel, Oaxaca.
Data analyses
Density was estimated by square meter (ind m-2) every month in both study sites, and a two-way ANOVA was applied to identify statistical differences between localities and months. All results were analyzed with a significant level of 0.05 and values were reported as mean and standard deviation (SD).
Abundance was estimated using the Zippin (1958) model applied to the data resulting from the sum of all urchins counted in the ten transects every month, in order to obtain an estimation of the abundance of the population along the year in each locality. Abundances of seven months were chosen in order to carry out the calculation considering the largest number of trappings proposed by Zippin (1958), which are seven samples.
A step by step multiple linear regression analysis was performed to evaluate the relation between monthly variations of the density (dependent variable) and variations of bottom temperature (BT), salinity (S), pH, and pluvial precipitation (PP) (independent variables).
Rate of natural mortality of D. mexicanum in both localities was calculated using Berry’s model (1967), which has been used and modified for other marine invertebrates (Ramos-Cruz, Sánchez-Meraz, Carrasco-Ayuso, & Cervantes-Hernández, 2006). This model uses the monthly variation of abundance of an organism recorded in the field (Ramos-Cruz et al., 2006).
The Berry’s model assumes that following an event of recruitment occurred during (n) time intervals (considered as independent), abundance of the organisms (in this case monthly) diminishes due to the action of natural mortality (M) (Ramos-Cruz et al., 2006). In the case of D. mexicanum natural mortality was estimated in those intervals in which abundance recorded in the field showed three or more continuously descending values.
The mathematical algorithm used for calculation of mortality is the following:
Ln(Nt)= α+β(t)
Where:
α : origin ordinate of the regression pendant.
β : is the pendant of the regression or value corresponding to the total mortality (Zprtial).
t: is the interval in which there exist diminishing of abundance.
Spatial distribution pattern for both sites was estimated with the Morisita’s index (1962), which is based in the analysis of the variancemean relationship. The distribution pattern was corroborated using the Poisson’s and Binomial Negative models, obtaining the values of X2, calculated and from tables in order to execute the statistical confrontation (Mendenhall & Reinmuth, 1978). The K parameter obtained from the Binomial Negative model indicates the intensity of aggregation of individuals from a population, and two methods were used in order to calculate it. If the first method did not fulfill the assumptions, the second method was used.
First method for calculation of K
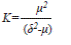
The value is satisfactory if it fulfills one of three assumptions:
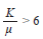
K > 3
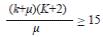
If K does not fulfill at least one of them, the second method is used:
Second method for calculation of K
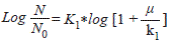
Where:
N = ∑ f (x)
No = the first date of f (x)
K 1 = Comes from the first method
Results
Density and abundance
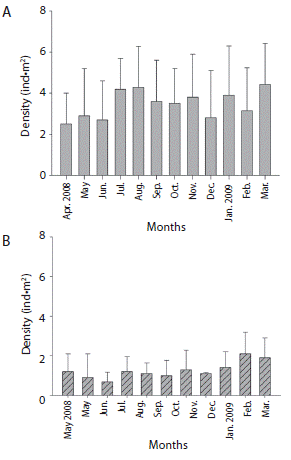
Mean monthly density in La Entrega was 3.4 ± 0.66 ind m-2, and it fluctuated between 2.5 and 4.4 ind m-2. An increase in density was recorded during June, July, August, and November 2008, reaching a peak in March 2009. Minimum values were observed in May, October, and December 2008, and February 2009 (Fig. 1A). In Isla Montosa, mean density was 1.2 ± 0.4 ind.m-2, fluctuating between 0.68 and 2.1 ind.m-2. During August and November 2008 we recorded high values, and a peak was recorded in February 2009, the minimum values corresponded to June and July 2008 (Fig. 1B). There were statistically significant differences between localities (F29.2 = 106.4, p < 0.05). According to Zippin’s model, abundance of D. mexicanum in La Entrega was 12 166 ± 25 individuals and 2 675 ± 33 individuals in Isla Montosa.
Environmental variables
Regarding environmental variables in La Entrega, mean (± SD) temperature during the sampling period was 26.58 ± 2.24 ºC, the lowest value (23.5 ºC) was recorded in March 2009 and the highest (29.8 ºC) in August 2008. Mean (± SD) salinity was 32.1 ± 1.5, the lowest value (29.3) was observed in June 2008 and the highest (34.4) in December 2008. Mean (± SD) value for pH was 8.7 ± 0.26 and it oscillated from 8.3 as the lowest during April and May 2008, and 9.1 as the highest in November 2008 (Fig. 2A).
In Isla Montosa mean value for temperature was 29.10 ± 2.26 ºC, the lowest value (23.1 ºC) was observed in November 2008 and the highest (29.7 ºC) in August 2008. Mean (± SD) salinity was 32.5 ± 2.6, the lowest value (28) was observed in July 2008 and the highest (35.3) in December 2008. Mean (± SD) value for pH was 8.9 ± 0.3, the lowest value (8.6) was recorded in October 2008, January and March 2009, and the highest (9.7) in February 2009 (Fig. 2B). The rainy season in the Bahías de Huatulco region during the sampling period occurred from March to September 2008, with a mean pluvial precipitation of 109.69 mm, and the highest precipitation during July (Fig. 2A, B).
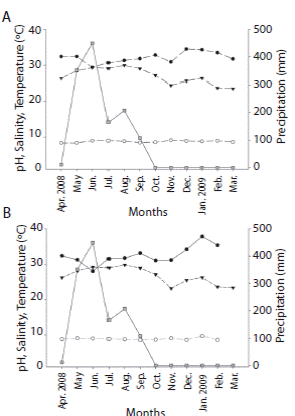
Fig. 2 Monthly variation of pluvial precipitation (solid line with grey squares), temperature (dashed line with black triangles), salinity (solid line with black dots) and pH (dashed line with white dots) in La Entrega (A) and Isla Montosa (B), from April 2008 to March 2009.
Multiple linear regression analysis of Isla Montosa was solved in two steps (Dn = - 2.002 + 0.109 S - 0.0508 TEMP, indicating that the total explained variance over Density, in terms of the selected environmental variables, was 82.67% (R2 = 0.894, R = 0.9, p multiple < 0.05). The first variable that went into the model was Salinity, providing 82.5% of the total explained variance. The partial contribution of variable Temperature to explain the variation of the abundance of D. mexicanum was 6.32 %. Regarding La Entrega,the analysis did not consider any variable to account for the ability to predict density of D. mexicanum.
Mortality rate
The natural mortality rate estimated, using the abundance recorded in the field for La Entrega, and considering one interval of continuous decreasing (August to October 2008) was 0.1. In Isla Montosa considering two intervals (May to July and August to October 2008) the average natural mortality rate was 0.15.
Spatial distribution
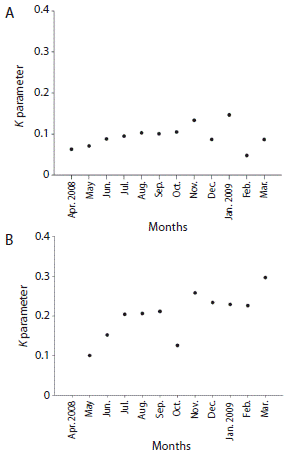
According to Morisita’s index, the spatial distribution pattern of D. mexicanum for both localities was patchy along the entire sampling period. The negative binomial distribution model confirmed the distribution pattern and Poisson’s model showed that the spatial distribution pattern was not random (Table 1). The aggregation parameter (k) provided by the negative binomial model for La Entrega showed that the months with higher levels of aggregation of D. mexicanum were April and December 2008, and February 2009, whereas the months when the urchin exhibited less aggregation were November 2008 and January 2009 (Fig. 3A). In Isla Montosa, the months when the intensity of aggregation was higher were May and October 2008, and December 2009. The lowest value of aggregation was recorded in February 2009 (Fig. 3B). Oscillations of k during the sampling period followed the fluctuation of the variance obtained with Morisita’s index in most of the months for both localities, and aggregation intensity was higher in La Entrega than in Isla Montosa.
Table 1 Monthly values of Morisita’s (σ² /µ), Poisson (λ), and negative binomial (k) models for La Entrega and Isla Montosa
| Locality | - | Entrega | - | - | Isla Montosa | - |
|---|---|---|---|---|---|---|
| Months | σ² /µ | λ | k | σ² / µ | λ | k |
| Apr-08 | 17.5 | 2.53 | 0.063 | - | - | - |
| May-08 | 24.7 | 2.97 | 0.071 | 7.79 | 0.96 | 0.100 |
| Jun-08 | 24.5 | 3.26 | 0.088 | 15.91 | 0.91 | 0.108 |
| Jul-08 | 20.7 | 4.19 | 0.095 | 6.42 | 1.36 | 0.204 |
| Aug-08 | 21.9 | 4.29 | 0.103 | 8.60 | 1.29 | 0.206 |
| Sep-08 | 16.9 | 3.65 | 0.101 | 7.60 | 1.10 | 0.211 |
| Oct-08 | 17.2 | 3.51 | 0.105 | 5.53 | 0.99 | 0.126 |
| Nov-08 | 17.3 | 3.88 | 0.134 | 7.87 | 1.29 | 0.258 |
| Dec-08 | 16.2 | 2.71 | 0.087 | 4.79 | 1.11 | 0.234 |
| Jan-09 | 18.2 | 3.90 | 0.146 | 7.96 | 1.44 | 0.229 |
| Feb-09 | 16.7 | 3.14 | 0.048 | 8.82 | 2.11 | 0.226 |
| Mar-09 | 27.5 | 4.42 | 0.087 | 6.47 | 1.93 | 0.297 |
Discussion
According to Zippin’s model, La Entrega is the locality with the highest abundance of D. mexicanum. An important difference between both sampled localities is the extension of the reef, which is less than a hectare in Isla Montosa and nearly seven hectares in La Entrega, making this latter one of the most extensive reefs in Bahías de Huatulco (Leyte-Morales, 2001), and is also the community with the largest proportion of death coral (around 25 %), as compared to Isla Montosa (around 10 %) (Leyte-Morales, 2001; Herrera-Escalante, López-Pérez, & Leyte-Morales, 2005). An important characteristic of this kind of substrate is that it is appropriate for proliferation of macro algae, which, in turn, constitute the most important source of food for D. mexicanum (Glynn & Leyte-Morales, 1997). Zamorano and Leyte-Morales (2009) proposed that in those communities where touristic activities are more intense and, consequently, possess more fragmented and death coral, populations of D. mexicanum are more abundant. Results of this study support that observation, since La Entrega is the community with the highest touristic impact in Bahías de Huatulco as well as the community with the most abundant population of D. mexicanum in the entire zone.
On the other hand, as a result of the strong influence of discharges from the Copalita River, especially during the summer, Isla Montosa shows the highest sedimentation rate (653.31 kg · m-2· yr-1), of the Bahías de Huatulco zone, whereas La Entrega is a relatively protected bay with minimal contribution of continental water and the sedimentation rate is low (14.33 kg · m-2· yr-1) (Granja-Fernández & LópezPérez, 2008).
Clemente & Hernández, (2008) proposed that the sedimentation rate and the presence of suspended sand are important factors that limit the population density of Diadema aff. Antillarum (now D. africanum) (Rodríguez, Hernández, Clemente & Coppard, 2013) in several localities of the Canary Islands. The authors observed that high sedimentation rates in rocky and coral communities inhibit algal recruitment and growth, producing adverse environments in terms of food source for the urchins, and suggest that the combined effect of intense waves and sediment covering the substrate may weaken adhesion of the urchins and restrain or, even more, hinder their motility.
Regarding temporal variations of density, monthly values in La Entrega showed the first increase between June and August 2008, which coincided with the rainy season (JuneOctober), which results from the increase in sea temperature, but in both localities the highest density was observed at the end of 2008 and beginning of 2009. This pattern observed in both localities suggests that the population density of D. mexicanum is indirectly influenced by the increase in primary production resulting from the effect of turbulence and mixture produced by the local winds known as “Nortes”. These winds reach their highest intensity from November to April, bringing nutrients from subsurface region and elevating substantially the availability of energy for primary producers and, in consequence, for planktonic and benthic herbivores, and this is reflected in an increase in population density (Fernández-Álamo & Färber-Lorda, 2006; Pennington et al., 2006).
Regarding other populations of Diadema mexicanum, the estimated average values calculated for both localities in this study were lower than those obtained previously by other authors (Hernández-Ballesteros, 2002; Herrera-Escalante et al., 2005; Zamorano & LeyteMorales, 2005; Benítez-Villalobos, Domínguez & López-Pérez, 2008), evidencing a continuous decrease in density since 2002, especially in La Entrega. Considering the Tropical Eastern Pacific, Isla Uva in the Gulf of Chiriquí in Panama is the locality with the highest reported population density (50 - 150 ind m-2), followed by Isla del Coco, Costa Rica (0.05 - 11.4 ind m-2), and La Entrega and Isla Montosa in Bahías de Huatulco, Mexico (0.36 - 7.5 ind m²), whereas the rest of studied localities show relatively low densities (0.1 - 2.7 ind m²). These differences in population densities do not seem to follow a defined latitudinal pattern, but result from particular characteristics inherent to each studied locality.
According to the statistical analysis, in Isla Montosa, the environmental variable that affected the population density of D. mexicanum in a negative way was temperature, whereas salinity showed to have a positive effect to a larger extent. The lowest estimations of density occurred during the raining season, coinciding with the highest values of sea temperature and the lowest values of salinity produced by the extensive supply of fresh water from the Copalita River. In general, the poor capacity of echinoids for ionic regulation makes this group of organisms extremely sensitive to low levels of salinity (Hyman, 1955; Shumway, 1977; Pia, Johnson & George, 2012) and this condition produces deficiency in consumption and utilization of energy, since a larger amount of energy is required to compensate the osmotic regulation at a cellular level, involving an important physiological cost (Forcucci & Lawrence, 1986). Long time exposures to low salinity conditions can be lethal for some echinoid species (Irlandi, Maciá & Serafy, 1997; Campbell & Russell, 2004).
The observed pattern in temperature and salinity reversed during the “Nortes” season (November-April) and coincided with the increase in population density, evidencing that despite the benefits of high temperature for tropical sea urchins, and the presence of this condition during the raining season in Isla Montosa, the combined effects of low levels of salinity, turbidity, and higher sedimentation rates create unfavorable conditions for D. mexicanum during that season.
In contrast, according to the statistical analysis, the environmental variables did not affect D. mexicanum population in La Entrega in the same way, as none of them showed a significant relationship with density of the urchin.
The apparent incongruent effect of salinity and temperature on the populations of both localities could be reflecting a complex interaction of various factors, such as reef geomorphology, currents, waves, sedimentation rates, and anthropogenic activity (López-Pérez, Hernández-Ballesteros & Herrera-Escalante, 2002), which, in synergy with complex interactions such as competence and predation, induce different scenarios on each community. Thus, probably the almost null influence of rivers in La Entrega did not allow evidencing clearly the effect of the environmental variables as occurred in Isla Montosa. In the latter, temperature had a negative effect, although it is directly related to population growth, it produces precipitation, which, in turn, affects negatively the population of D. mexicanum, as well as the low salinity.
The fact that Isla Montosa possesses a more stressful environment for D. mexicanum as a consequence of its high sedimentation rates and higher exposure to direct waves, is reflected also in a higher mortality rate as compared to the population of D. mexicanum living in La Entrega, and to other species inhabiting the tropical eastern Pacific, such as D. antillarum in the Virgin Islands (Randall, Schroeder, & Starck, 1964) and Barbados (Lewis, 1996), as well as D. paucispinum in Hawaii (Ebert, 1982).
The aggregated spatial pattern observed for D. mexicanum throughout the entire sampling period in both studied localities generally, occurs when the environment is heterogeneous or positive interactions exist between organisms (Ludwig & Reynolds, 1988; Krebs, 1999), this has been documented for a number of echinoderm species (Randall et al., 1964; Pearse & Arch, 1969; Bauer, 1976; Levitan, 1988; Hamel & Mercier, 1995; Campbell, Coppard, D’Abreo, & Tudor-Thomas, 2001; Alvarado, 2008).
Regarding populations of D. mexicanum, it has been proposed that the aggregation behavior does not simply correspond to a passive response to environmental limitations, since adjacent areas with similar substrata and feeding conditions show different levels of occupation and aggregation (Pearse & Arch, 1969). In La Entrega, this sea urchin shows a stratified pattern of aggregation, having densities of more than 11 individuals per square meter at higher depths in the reef (Herrera-Escalante et al., 2005), where conditions of large areas of rubble and death coral prevail.
Pearse & Arch (1969) suggest that large exposed aggregations of D. setosum and D. savignyi in unprotected areas, of the reefs of Papua New Guinea, result from populations with too large sizes, in which the number of individuals exceeds the number of available refuges, and the probability of encounter among organisms to form a patch is larger than the probability of finding an appropriate refuge in the reef. This seems to coincide with findings for the population of D. mexicanum in La Entrega, since this is the locality with the largest aggregations in the deep zone of the reef and also with the largest population density, significantly different from Isla Montosa and other localities of the Bahias de Huatulco area (Herrera-Escalante et al., 2005; BenítezVillalobos et al., 2008). Additionally, the low level of structural complexity, provided by the prevalence of corals of the genus Pocillopora in La Entrega, basically offers fewer crevices that could be used as refuge, propitiating that individuals of D. mexicanum compose large aggregations in open areas of the reef, constituted principally by rubble and death coral.
Despite that La Entrega and Isla Montosa are relatively near localities and, theoretically, their oceanic conditions are the same, population dynamics of D. mexicanum is different in terms of its population parameters characterized in this study. According to a lower mortality rate and larger densities and abundance, population of La Entrega inhabits an environment with more favorable conditions of food and space and relatively low levels of stress, whereas the population of Isla Montosa undergoes a major environmental stress in terms of high sedimentation rates and a smaller reef surface.

