











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkLos páramos son ecosistemas tropicales de alta montaña que se caracterizan por presentar condiciones drásticas a causa de la alta radiación, baja temperatura anual, frecuentes heladas nocturnas y alta humedad relativa (Monasterio, 1980; Hofstede, 1995; Castrillo, 2006). La alternancia de irradiación directa durante el día y congelamiento y gelifluxión durante la noche genera amplias variaciones de temperatura que superan los regímenes estacionales (Monasterio, & Sarmiento, 1991; Luteyn, 1992). En general, estas condiciones abióticas conllevan a que los páramos tengan una baja productividad primaria, una lenta descomposición de la biomasa, difícil absorción de nutrientes y retraso de los procesos sucesiona-les (Luteyn, 1999; Sánchez, 2003).
Colombia tiene una extensión de 14 400 km2 de páramo, que aunque corresponden a más de un 40 % de la extensión mundial, en realidad sólo representan el 1.3 % del territorio Colombiano (Hofstede, Segarra, & Mena, 2003) . Diversas actividades como la ganadería, agricultura, quema, deforestación y extracción de recursos biológicos son actualmente fuertes factores antrópicos que llevan a que el territorio de páramo sea cada vez más reducido, afectando la sobrevivencia de especies de fauna y flora específicas para este ecosistema (Vargas, Jaimes, Castaño, & Mora, 2004; Alonso-Amelot, 2008; Castaño, 2002).
Espeletia paipana S. Díaz & Pedraza es una roseta caulescente endémica del departamento de Boyacá, que se encuentra en la categoría En Peligro Crítico por la UICN, debido a su dramática reducción poblacional, ocupando un área menor a 0.007 km2 (López, 2004) . Para el año 2004 se reportó un total de 76 individuos entre adultos, jóvenes y plántulas, de los cuales, actualmente sobreviven 27 individuos en su mayoría adultos.
Con base en observaciones detalladas de las hojas de E. paipana, en el 2012 detectamos una enfermedad foliar que se caracteriza por la pérdida de pubescencia, lesión que en estados avanzados causa la formación de manchas necróticas, clorosis y deformación de la hoja. El descubrimiento de la enfermedad dio origen al interrogante sobre si la acelerada mortandad poblacional estaba relacionada con la afectación por dos vías simultáneas: el estrés abiótico a causa de la disminución de la regulación hídrica y térmica que representa la pérdida de pubescencia foliar (PPF), y el estrés biótico, generado por la acción patogénica de microorganismos asociados a la lesión en el tejido fotosintético.
La pubescencia foliar es una característica de gran valor adaptativo para las plantas del páramo, debido a que atenúa los efectos negativos de condiciones ambientales adversas sobre el estado fisiológico y el crecimiento. Es una modificación de la epidermis que varía tanto en longitud como en densidad de acuerdo a la altitud, con el fin de regular la alta radicación incidente, la economía hídrica y la difusión de gases (Meinzer & Goldstein, 1985, Molina-Montenegro, 2008). En espeletias, la pubescencia aumenta con el gradiente altitudi-nal permitiendo el desacople con la temperatura ambiente, es decir, actúa como regulador térmico que permite mantener los óptimos de temperatura para la fotosíntesis. Llega a reflectar más del 20 % de la radiación incidente, reduciendo cerca de un 10 % la cantidad de radiación asimilada por la hoja; aumenta el grosor de la capa límite, al disminuir la pérdida de calor por convección durante la transpiración, y retiene humedad, al reducir el efecto abrasivo y la desecación a causa del viento (Goldstein, Rada, Canales, & Zabala, 1989; Azócar & Rada, 1993; Levizou, Drilias, Psaras, & Manetas, 2004; Castrillo, 2006).
Con relación a la infección foliar por patógenos, se sabe que la incidencia de fitopa-tógenos altera la susceptibilidad y/o tolerancia de las plantas a las condiciones del ambiente y al ataque de insectos (Paul, 1982; Fisher & Petrini, 1991; Sempere & Santamarina, 2008). Aspectos que posiblemente determinan en gran medida el éxito reproductivo y la permanencia de las poblaciones vegetales en el tiempo. Sin embargo, es escaso el conocimiento acerca del tipo de interacciones bióticas que rodean a la gran mayoría de especies vegetales de páramo y más aún, se desconoce el efecto conjunto del estrés biótico y abiótico sobre el desempeño fisiológico de las mismas.
En este estudio se evaluó el estado sanitario foliar y el rendimiento fisiológico de E. paipana con el fin de identificar los factores que han acelerado la reducción de su población. Este trabajo se llevó a cabo mediante el análisis del comportamiento de la incidencia y la severidad de la PPF, la caracterización de la comunidad de hongos asociados a esta patología y el ensayo de la capacidad infectiva de hongos altamente agresivos, aislados del tejido foliar afectado. Para determinar el efecto de la PPF en la fisiología de la especie se valoró la conductancia estomática, la temperatura foliar y el contenido de clorofilas. La identificación de una actividad patogénica y la alteración fisiológica asociada a la PPF, permitirá determinar estrategias de control y prevención para esta y otras espeletias.
Materiales y Métodos
Área de estudio: El estudio se realizó durante el 2013 en el páramo La Cuchilla (5°51'11.4" N - 73°0.8'33.7" W) a una altitud entre los 3 509 y 3 567 m, dentro del Parque Natural Municipal Ranchería. El páramo es parte del Corredor Guantiva-La Rusia-Iguaque ubicado en el costado occidental de la Cordillera Oriental, en jurisdicción de los departamentos de Boyacá, Santander y Cundinamarca (Sánchez & Cely, 2003; Solano, Roa, & Calle, 2005). Este complejo presenta un régimen de lluvia bimodal comprendido entre Abril-Mayo y Octubre-Noviembre, y dos periodos secos en Diciembre-Marzo y Junio-Agosto con temperaturas promedio en época seca de 27.6 °C y en época húmeda de 25.7 °C. La precipitación anual fluctúa entre 700 mm y más de 3 000 mm (Solano, et al., 2005).
La población vegetal del páramo La Cuchilla esta conformada por pastizales de Calama-grostis, matorrales de Hypericum, Chusquea y helechos del género Blechnum. E. paipana se encuentra asociada a poblaciones de E. neme-kenei, E. incana, E. argentea y Espeletiopsis guacharaca (López, 2004).
Distribución de la población: La distribución de los 27 individuos de E. paipana en el sitio de estudio comprende tres zonas: cima (3 567 m), pendiente (3 542 m) y depresión (3 509 m). La zona fue considerada como un factor determinante de las diferentes variables morfo-fisiológicas y sanitarias, razón por la cual se analizó su efecto en este estudio.
Dado que la condición más estresante en los páramos es la alta radiación lumínica, se midió la radiación fotosintéticamente activa (PAR) en el rango diurno de mayor intensidad solar, en intervalos de 15 minutos desde las 11:00 h hasta las 14:00 h usando un medidor Apogeé MQ-100.
Evaluación de la incidencia y la severidad: Se diagnosticó la pérdida de pubescencia foliar (PPF) tanto en el haz como en el envés de las hojas, como una enfermedad que conlleva la presencia de manchas necróticas, quemazón, clorosis y deformación. La incidencia de PPF se evaluó en cada uno de los 27 individuos como el porcentaje de hojas que presentaron los síntomas mencionados. Como un factor colateral a la PPF, se estimó la incidencia de la herbivoría por plagas (porcentaje de hojas con signos de esqueletización y defoliación). Los datos fueron tomados a los largo de nueve meses y analizados por zonas mediante el test de Kruskal Wallis. Además, se realizó una correlación mediante el coeficiente de Spearman para determinar el comportamiento de la herbivoría respecto a la PPF.
Por otro lado, la severidad se estimó como el porcentaje de área afectada de la hoja, calculado en cinco hojas por individuo seleccionadas al azar, fotografiadas y analizadas en el programa Axio Vision LE 4.8 (2009). Los datos de severidad fueron transformados con Log10 (x+1) y analizados mediante ANOVAS a una vía para identificar diferencias entre zonas, y determinar la variación de la severidad entre el haz y el envés de las hojas.
A partir de los resultados de incidencia y severidad se estimó el porcentaje de biomasa foliar con PPF, con base en el cual se clasificó a los individuos en tres niveles de afectación: bajo (0-5 %), medio (5-10 %) y alto (> 10 %).
Mediciones morfo-fisiológicas: Durante cuatro días consecutivos en la época seca de enero y en la época húmeda de noviembre, se midió la conductancia estomática (Gs) y la temperatura foliar (Tfol) en dos hojas sanas y dos hojas con PPF de cada uno de los 27 individuos. Para mayor homogeneidad de los datos, los registros se realizaron durante las tres horas de mayor luminosidad simultáneamente a las mediciones de radiación PAR, utilizando un porómetro digital Decagon Devices SC-1. Así mismo, se midió el índice de contenido de clorofila (ICC): tres mediciones por hoja sana y tres mediciones en hojas afectadas, empleando un clorofilómetro OptiScience CCM-200.
Los datos fueron normalizados mediante la transformación tipo Log10 para la Gs y el ICC y A/X para la variable Tfol, y analizados mediante ANOVA a dos vías, para observar el efecto de la PPF, la zona y la época. Las variables Gs y Tfol fueron correlacionadas con el coeficiente de Spearman para establecer cómo se relaciona el grado de enfriamiento foliar con la conductancia estomática.
Dentro de las características morfológicas, se midió el área foliar (Afol) y la altura de los individuos. El Afol se obtuvo del promedio de cinco hojas maduras por roseta mediante mediciones con regla de las dimensiones de la hoja, fotografías y su posterior análisis en el programa Axio Vision LE 4.8.
Aislamiento e identificación de hongos: Se obtuvieron muestras del tejido foliar afectado a partir de 15 hojas de E. paipana. Las muestras se almacenaron en bolsas sellables a 4 °C y se procesaron dentro de las 24 horas siguientes. El material se cortó en secciones pequeñas (< 1 cm2) que comprendían el área afectada y parte del tejido sano colindante. La esterilización de los cortes se realizó por inmersión en alcohol 70 % (1 min), en hipoclorito de sodio 1 % (1 min) y cinco lavados de 1 min con agua destilada estéril de acuerdo con la metodología de French & Hebert (1980). Posteriormente, se sembraron tres cortes en cajas de Petri utilizando Agar Papa Dextrosa (PDA), Agar malta y Agar nutriente. Las siembras se incubaron a 27 °C durante ocho a 30 días. Los cultivos fueron purificados e identificados mediante microcultivos, empleando las claves taxonómicas de Barnett (1960), Hanlin (1990) y Castaño (1998).
Los resultados de los aislamientos se contrastaron con observaciones al microscopio de 20 improntas realizadas en campo. Para esto se tomaron muestras con cinta transparente que cubrieran el área afectada y sana de la hoja. Las muestras se colorearon con azul de metileno al colocar la cinta sobre la lámina portaobjetos.
Estudio de patogenicidad: Dado el estado de amenaza de E. paipana, es escaso el material foliar sano que puede extraerse de la zona de estudio. Para las pruebas de patogenicidad se realizaron tres pruebas de infección en hojas de E. paipana (4 hojas) y de E. nemekenei (20 hojas), empleando las cepas Botrytis sp., Fusarium sp. cepa F1 y Fusarium sp. cepa F2, aisladas del tejido foliar afectado. A partir del cultivo puro de cada cepa, se raspó el micelio superficialmente para obtener una suspensión de conidias en agua estéril y Tween al 0.5 %. La suspensión se observó al microscopio y se cuantificó mediante un hemocitómetro. Se preparó 5 mL de inóculo a una concentración de 106 esporas/mL, que fue suministrada en dosis de 0.5 mL en el ápice, base y en el centro de la vena principal de la hoja empleando jeringas de 5 mL.
Se evaluó el avance de la infección de cada cepa y la respuesta de un control (seis hojas sanas) cada dos a tres días durante un periodo de 42 días en el laboratorio, donde las hojas se mantuvieron en cámara húmeda bajo condiciones de luz difusa entre 18 a 68 ^mol/m2.s, temperatura de 19 °C y humedad relativa de 88 %.
Todos los análisis estadísticos se realizaron utilizando el programa STATISTICA 7.0 (StatSoft, Inc., 2004).
Resultados
Radiación fotosintéticamente activa: La radiación PAR varía significativamente entre épocas (MW-U (N = 312) Z = 3.57, P < 0.05), y fue altamente fluctuante en la cima, donde los valores máximos alcanzaron los 3 000 ^mol/ m2.s en época seca, el doble de los valores registrados para la época húmeda. En general, la radiación PAR supera los 2 000 ^mol/m2.s en las demás zonas.
Pérdida de pubescencia foliar (PPF): La PPF en E. paipana es una enfermedad que conlleva a la formación de manchas necróticas irregulares, clorosis y la deformación de las hojas. En el haz se observó la marchitez y desprendimiento del tejido intervenal, y en el envés se evidenciaron lesiones cafés sobre la nervadura central a manera de un "raspado" que asciende hacia el ápice de la hoja causando marchitez. La incidencia de la PPF varió entre zonas (KW-H (2.230) = 78.68, P < 0.05) con mayor repercusión en los individuos de la depresión, donde el porcentaje de hojas afectadas se mantuvo entre un 37 a 55 %. En cada zona, el porcentaje de incidencia fue constante a lo largo de los nueve meses de seguimiento y presentó una correlación negativa con la incidencia de la herbivoría por plagas (r = -0.62, P < 0.05), la cual sufrió un descenso marcado a partir de junio cuando inició la época de lluvias (Fig. 1).
Aunque la incidencia de la PPF fue constante, la severidad de la misma aumentó en promedio un 0.13 % por mes, y difirió entre zonas (F (2 420)=16.76, P < 0.05); esta fue mayor en la cima. El porcentaje de área afectada por la PPF fue mayor en el haz de las hojas (1.420) = 5.79, P = 0.01). Entre los meses de Mayo a junio se observó un rápido incremento de la severidad (0.33 %) en todas las zonas, que finalizó en julio, causando la muerte del 64.8 % de las hojas con porcentajes mayores al 50 % (Fig. 1). En general, el porcentaje de biomasa foliar total con PPF fue bajo en el 50 % de la población, medio en un 42 % y alto en el 8 % restante.
Efecto de la PPF en la fisiología de E. paipana: La PPF afectó significativamente la Gs (F (1.804) = 315.57, P < 0.05). El intercambio gaseoso de las hojas sanas (143.2 mmolH2O/m2.s) duplicó en promedio al de las hojas con la enfermedad (68.6 mmolH2O/ m2.s), principalmente durante la época seca. La respuesta fue distinta para los individuos de la pendiente (F (2.804) = 8.10, P < 0.05), los cuales mantuvieron valores de Gs similares durante las dos épocas, alrededor de 102.6 mmolH2O/m2.s (Fig. 2). La Gs promedio fue mayor en los individuos de la depresión (F (2.197) = 7.66, P < 0.05) (Cuadro 1).
Se encontró una alta variabilidad del contenido de clorofila entre hojas, por lo cual no se evidenció un patrón definido relacionado con la PPF. Con relación a la zona, el contenido de clorofila fue significativamente mayor en la pendiente (F (2.309) = 11.42, P < 0.05) (Fig. 2).
Efecto de la época y la zona en la fisiología de E. paipana: Las características morfo-fisiológicas de los individuos de E. paipana varían entre zonas. A medida que se desciende hacia la depresión, los individuos poseen tallas más pequeñas con hojas más grandes (F (2.97) = 14.75, P < 0.05), que sin importar la época logran mantener temperaturas cercanas a 19 °C, aproximadamente 4 °C por encima de las temperaturas registradas en la cima y la pendiente (F (2.197) = 35.21, P < 0.05) (Cuadro 1) . En contraste, durante la época seca, los individuos de la cima y la pendiente mostraron temperaturas foliares que superaron en unos 4 °C a las registradas para la época húmeda (Fig. 2) . Al analizar las correlaciones entre variables morfo-fisiológicas, se encontró una relación significativa entre Gs y Tfol (r = 0.44, P < 0.01) durante la época húmeda que no estuvo presente durante la época seca.
Cuadro 1: Variaciones morfo-fisiológicas de E. paipana entre zonas topográficas
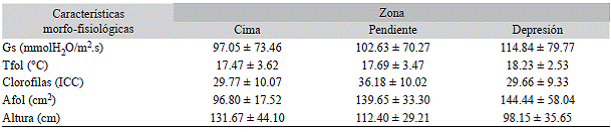
Media ± desviación estándar, Gs: Conducticia estomática, Tfol: Temperatura foliar, Afol: área foliar.
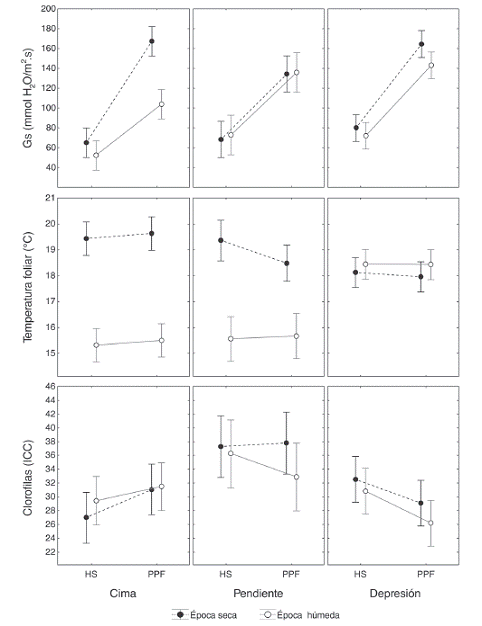
Fig. 1: Incidencia y severidad de la PPF (Pérdida de Pubescencia Foliar) en individuos de E. paipana ubicados en cima (A), pendiente (B) y depresión (C). En la izquierda se compara la incidencia de la PPF y de la herbivoría por insectos a lo largo de nueve meses. En la derecha se muestra el avance de la severidad en el haz y el envés de hojas de durante ocho meses de seguimiento. Las franjas sombreadas indican períodos de cambios significativos en los porcentajes de incidencia y severidad.
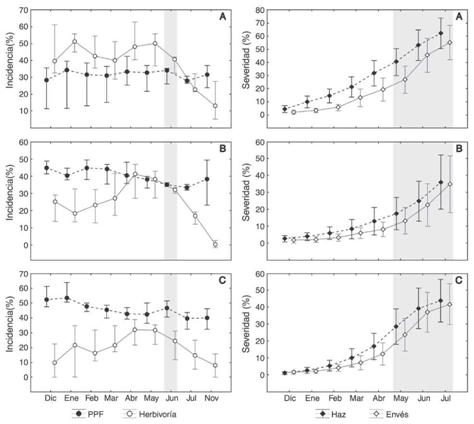
Fig. 2: Efecto de la pérdida de pubescencia foliar (PPF), zona y clima sobre las variables fisiológicas: Conductancia estomática (Gs), temperatura foliar (Tfol) e índice del contenido de clorofila (ICC). HS: Hojas sanas.
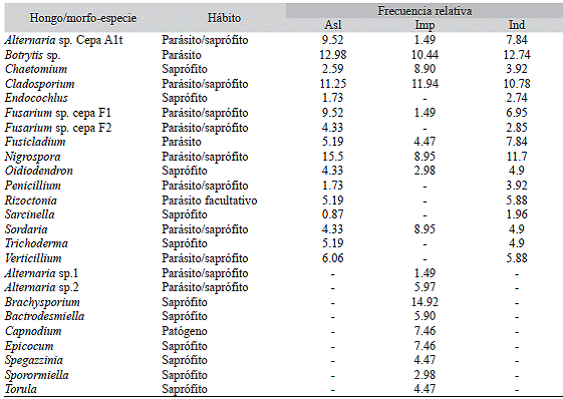
Aislamiento e identificación de hongos: A partir de las muestras del tejido foliar se aislaron 117 hongos y se logró identificar 16 géneros con una alta representación de Nigrospora (15.5 %), Botrytis (12.9 %), Cla-dosporium (11.25 %) y Fusarium (13.8 %), este último con dos morfotipos clasificados como cepa F1 y cepa F2. Dentro de los géneros menos frecuentes se encontró a Sarcinella (0.9 %) y Endocochlus (1.7 %) (Cuadro 2).
Algunas cepas no presentaron cuerpos de fructificación que permitieran su identificación y por tanto fueron agrupados como micelios estériles. Botrytis, Cladosporium y Alternaría fueron los géneros más frecuentes en los individuos con PPF a partir de los cuales se obtuvieron las muestras.
En las improntas realizadas en campo, se observaron esporas y algunos residuos mice-liares de hongos de los géneros Nigrospora, Epicoccum, Brachysporium, Alternaria, Fusi-cladium, Cladosporium, Fusarium, Spegazzi-nia, Sporormiella, Torula y Chaetomium. A partir de los ensayos tanto en campo como en laboratorio, se observaron tres morfotipos distintos para el género Alternaria, de los cuales se obtuvo la cepa Alt1 (Cuadro 2).
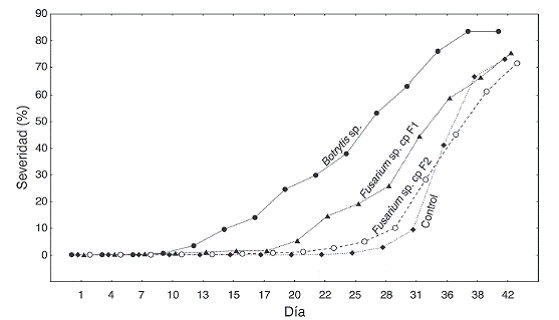
Pruebas de patogenicidad: Por su fácil aislamiento, purificación y replicación, se seleccionaron los hongos Botrytis sp, Fusa-rium sp. cepa Fl y Fusarium sp. cepa F2 para las pruebas de patogenicidad. A pesar de estar presentes persistentemente en las lesiones, no fue posible reproducir los síntomas observados en campo durante los 42 días de evaluación de la prueba. Sin embargo, se logró describir el comportamiento de los hongos seleccionados sobre el tejido foliar sano de E. paipana y de E. nemekenei.
Las hojas inoculadas con Botrytis sp. presentaron síntomas a los 10 días. Inicialmente se observaron manchas necróticas de color marrón distribuidas heterogéneamente sobre el envés. A los 20 días, las manchas crecieron y se unieron diseminándose desde el ápice hacia la base, dando a la hoja un aspecto blando y flexible. En algunos casos, el crecimiento del micelio sobrepasó la pubescencia de la hoja, exhibiendo cuerpos fructíferos y esclerocios de color pardo. De esta manera, a partir del día 28 las hojas comenzaron a morir con porcentajes de severidad superiores al 50 % (Fig. 3).
Fusarium sp. cepa F1 causó pudrición blanda en la base de la vena principal de la hoja, tornándose de color purpura-café, y en algunos casos produjo micelio en la superficie del envés. La incidencia de este patógeno fue baja, con porcentajes de severidad menores al 30 % hasta el día 28, cuando las hojas comenzaron a morir por pérdida de turgencia y necrosis. Por otro lado, las hojas inoculadas con Fusarium sp. cepa F2 presentaron manchas circulares negras cuyo diámetro aumentaba en el tiempo, generando grietas y marchitamiento, síntomas asociados con la invasión del hongo a través de los vasos conductores de la hoja, en este caso el porcentaje de severidad aumentó a partir del día 31, causando la muerte de la mayoría de las hojas (Fig. 3).
Botrytis Fusarium Fusarium E. paipana E. nemekenei
Discusión
Aunque la incidencia de la PPF en E. paipana tiende a ser constante en el tiempo, la severidad aumenta de manera intensiva iniciando con un porcentaje de área foliar afectada menor al 10 % y llegando al 45 % en la depresión y al 65 % en la cima al final de ocho meses de seguimiento. El efecto foto-oxidativo de la radiación diurna puede ser considerado un factor que intensifica la severidad de la enfermedad en el haz de las hojas, debido a que diariamente están expuestas a valores PAR superiores a 2 000 umol/m2.s. Esta radiación sobrepasa ampliamente los umbrales de foto-inhibición para la gran mayoría de las plantas, incluso para aquellas consideradas heliófitas (1 445.16 umol/m2.s, Fitter & Hay, 2002). El avance de la severidad de la PPF fue estimado en alrededor de 7.68 cm2 de área foliar/mes. Esta tasa de incremento fue similar entre zonas y conllevó a la pérdida de un importante número de hojas en toda la población.
La correlación negativa entre la incidencia de la herbivoría por plagas y la incidencia de la PPF, sugiere que existe una relación antagónica entre los agentes causantes de estos dos tipos de daño foliar. La herbivoría afecta en mayor proporción a los individuos ubicados en la cima, principalmente durante las épocas secas del año. Cuando el área foliar se reduce a causa del consumo por insectos, disminuye el transporte de agua y nutrientes en la planta, haciéndolas menos tolerantes a la depredación y más susceptibles al estrés hídrico (Tyree, Velez, & Dalling, 1998; Cavender & Bazzaz, 2000). Sin embargo, al iniciar la temporada de lluvias en el mes de junio la herbivoría se reduce y la PPF permanece constante.
La reducción del área foliar funcional, a causa de la PPF, limita significativamente la eficiencia de los procesos fisiológicos en E. paipana. En hojas sanas, la pubescencia abundante aumenta la capa límite, reduciendo el choque térmico con el ambiente y permitiendo la apertura estomática y la difusión de gases durante el día (Meinzer & Golstein, 1985; Gates, 1980). Se ha visto que con el incremento de la altitud, la pubescencia foliar sirve para asegurar el desacople térmico de la temperatura ambiente y reflejar un mayor porcentaje de radiación, así esto implique una menor fijación de carbono (Sandquist & Ehleringer, 1997).
La pubescencia es considerada la primera barrera frente a la alta radiación fotosintética-mente activa y UV-B. Llegando a reflectar más del 20 % de la radiación incidente, con el fin de evitar el daño en el fotosistema II y la degradación de otros pigmentos (Lang & Schindler, 1994; Molina-Montenegro, 2008; Ranjbarfor-doei, Samson, & Van Damme, 2011). Aunque no se comprobó un efecto negativo en los contenidos de clorofilas asociado a la PPF, se observó que las hojas afectadas presentan signos de fotoinhibición, evidentes por una reducción de la media de conductancia estomática de alrededor del 48 % durante la franja del mediodía. Esto conlleva a bajas tasas de asimilación de CO2 (Goldstein, et al., 1989; Ázocar & Rada, 1993; Castrillo, 2006) que a su vez afectarán el ritmo de crecimiento y la reposición de biomasa.
E. paipana presenta un promedio de conductancia estomática (115.27 mmolH2O/m2.s) menor al que se ha medido en Espeletias típicas de los páramos venezolanos como E. schultzii (150 y 100 mmolH2O/m2.s) y E. neriifolia (200-250 mmolH2O/m2.s) (Cavieres, Rada, Azócar, García-Núñez, & Cabrera, 2000; Azócar & Rada, 2006). Para poblaciones de E. schultzii se ha demostrado que la Gs se reduce en un 34 % al ascender de 2 800 a 4 200 m (Rada, Azócar, González, & Briseño, 1998), mientras que con E. neriifolia se ha establecido que la asimilación neta de CO2 durante días nublados se reduce a valores críticos, cercanos al punto de compensación (Cavieres, et al., 2000). Estas evidencias son solo una muestra de la complejidad de las respuestas funcionales de las rosetas suculentas frente a la variabilidad ambiental de alta montaña, aunque en general sugieren que las espeletias son plantas que apuestan a rendimientos fisiológicos bajos pero seguros, gracias a características morfo-anatómicas de alta adaptación al páramo.
La zona topográfica donde están ubicados los individuos de E. paipana es un factor decisivo en la morfo-fisiología de la especie, de igual o mayor importancia que la PPF. El contenido de clorofila fue en promedio mucho mayor en los individuos de la pendiente en comparación con la depresión y la cima, lo que puede indicar que existe una importante influencia del ángulo de orientación con relación a la irradiación. Los individuos de la cima y la depresión, al estar expuestos de forma perpendicular a la radiación solar, producirían una menor cantidad de pigmentos fotosintéticos y harían una mayor inversión en pigmentos foto-protectores (Manuel, et al., 1999; Kórner, 2003; Molina-Montenegro & Cavieres, 2010).
En la cima hubo una correlación altamente positiva entre la Tfol y la Gs durante la época húmeda, que sugiere que el desacoplamiento con el exterior se reduce significativamente por efecto de una mayor influencia directa del viento (Kozlowski & Pallardy, 1979; Allen & Ort, 2001), llevando a que la planta tenga una menor capacidad de regulación térmica y mayores limitaciones difusivas para hacer fotosíntesis. Estas plantas también presentan una menor área foliar para reducir la fotoinhibición por la alta radiación que se presenta en la cima durante la época seca (Valladares, et al., 2002; Colmenares, Rada, & Luque, 2005).
Por otra parte, en la depresión los individuos poseen hojas más grandes que en promedio mantienen una mayor temperatura, con valores cercanos a los óptimos para la fotosíntesis en E. neriifolia a 3 200 m (19.4 °C, Cavieres, et al., 2000). Teniendo en cuenta que en Espeletias los óptimos fisiológicos se reducen a medida que se asciende en altitud (Cavieres, et al., 2000; Azocar & Rada, 2006), puede afirmarse que a 3 509 m y con una Tfol media de 18.3 °C, E. paipana podría realizar una fotosíntesis óptima asociada a una alta Gs. La regulación de la temperatura foliar en la depresión persiste sin importar la época del año, y podría verse favorecida por la arquitectura densa y de bajo porte de la roseta, que minimiza la exposición al aire frío (Goldstein, Meinzer, & Monasterio, 1985).
El alto nivel de afectación del 8 % de la población, la pérdida de biomasa foliar, sumado a los efectos sobre el desempeño fisiológico de los individuos, sugieren que la PPF es un serio causante de mortalidad en la población de E. paipana. La comunidad de hongos asociados al tejido foliar sin pubescencia, comprende en su mayoría una gran variedad de hongos endófitos, de los cuales algunos géneros como Fusarium, Botrytis y Alternaria, poseen especies con alta capacidad patogénica (Fisher & Petrini, 1991; Agrios, 2005). Con base en la alta representatividad en los aislamientos y su poder de infección durante las pruebas de patogenicidad, la cepa Botrytis sp. es propuesta como el agente causal de la PPF.
Botrytis es un patógeno ubicuo que infecta hojas, flores y frutos, causa estrangulamiento del tallo, manchas y pudrición en tubérculos, bulbos y raíces de vegetales y frutales (Elad & Shtienberg, 1995; Agrios, 2005). Las condiciones de alta humedad relativa, un bajo déficit de presión de vapor en la superficie de las plantas y temperaturas bajas, características de los ambientes de páramo en época húmeda, promueven la infección por Botrytis y predisponen la susceptibilidad del hospedero (Elad, 1996; Widiastuti, et al., 2011). La acción necrotrófica de este hongo le permite infectar a su hospedero a partir de heridas superficiales, donde contamina el agua extracelular con sus esporas. Posteriormente estas germinan infectando el tejido ileso, proceso que lleva a cabo mediante la producción de enzimas como la lipasa y la cutinasa (Agrios, 2005). Desde el momento en el que inicia la infección, los contenidos celulares se tornan cafés, hasta que eventualmente se produce la muerte de las partes afectadas (H.M.R., 1899).
La participación de Botrytis en conjunto con otros patógenos, pone en evidencia la existencia de mecanismos de resistencia de la planta frente al efecto negativo de la infección. Esta podría ser una de las razones por las que no se ha hecho hincapié en estudiar el estado sanitario de estas plantas y sus interacciones bióticas. Por otro lado, en este estudio se demuestra que pese a la resistencia que genéticamente (por adaptación) estas especies poseen frente a la patogenicidad de muchos microorganismos, las alteraciones drásticas en las condiciones ambientales pueden promover su vulnerabilidad a todo tipo de estrés biótico.
Aunque no fue posible reproducir los síntomas de PPF en los ensayos de patogenicidad, se sugiere que en condiciones naturales el agente causal de la infección podría actuar de manera sinérgica con otros microorganismos como Nigrospora, Cladosporium y Alternaria. Géneros comunes en lesiones foliares de hábito saprófito facultativo que pueden actuar de forma patogénica bajo condiciones de estrés (Fisher & Petrini, 1991; Sempere & Santama-rina, 2008). Lamentablemente, en las condiciones poblacionales actuales de E. paipana, el material foliar sano es insuficiente para realizar nuevas pruebas de patogenicidad que corroboren este planteamiento.
En general, E. paipana es una especie con serios problemas que afectan su sobrevivencia y conservación. Estos incluyen las bajas posibilidades de regeneración in vitro; la reducción de la conductancia estomática que conllevarían a un bajo rendimiento fotosintético; una infección patogénica causante de la pérdida de pubescencia foliar que afecta en mayor o menor grado al 100 % de la población y de severidad creciente, que a su vez limita el desempeño fisiológico de la especie. Es evidente la necesidad inmediata de una intervención in situ que podría involucrar el estudio y uso de controladores biológicos como los hongos Thrichoderma, Chaetomium y la bacteria Bacillus licheniformis (datos no mostrados), aislados de la misma especie con el fin de evitar su extinción.

