











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkWith a wide distribution in tropical areas, inselbergs feature a range of sizes and degrees of isolation associated with major biomes of the world, allowing experimental work, as well as being excellent elements to address different topics related to biodiversity and conservation (Porembski & Barthlott, 2000; Romer, 2005).
Due to its own characteristics, inselbergs form centers of diversity for certain functional groups of plants that are well adapted to extreme environmental conditions, and where the occurrence of seeds adapted to these conditions, becomes also greater in these formations than in the adjacent matrix (Hunter, 2003; Porembski, 2007). Its rocky structure is capable of forming xeric islands within a tropical rainforest matrix, resisting harsh soil edaphic and microclimate conditions (Sarthou, Larpinb, Fontyc, Pavoined, & Ponge, 2010).
Studies in outcrops of Brazilian semiarid region showed that, while the dominant climate is the semi-arid, the formation of semi-deciduous forests may happen, which characterizes the local occurrence of wetter environments, favoring the establishment of phanerophytes (Franga, Melo, & Santos, 1997). According to Araujo, Oliveira & Lima-Verde (2008), the vegetation that is established on rocky outcrops in the semi-arid region has a high species richness, demonstrating the relevance of studies related to the biology and ecology of these species, and the conservation actions for these areas (Scarano, 2007).
Due to the absence of large substrate accumulations, water storage becomes incipient and much of it is rapidly lost by runoff. Due to the little substrate available and extreme microclimate conditions, the vegetation that is established in these environments is very different from its surroundings, favoring even the development of ephemeral vegetation (Sarthou & Villiers, 1998). Environmental conditions inherent to these formations are capable of providing niches to different taxa, depending on the surrounding vegetation and environmental pressure. In such harsh environments, the presence of effective dispersive mechanisms is of paramount importance for the development and establishment of seedlings (Willson & Traveset, 2000).
Based on the premise that short distances associated with abiotic syndromes may influence the floristic composition in rocky environments, this study aimed to verify the composition, and to perform a floristic similarity analysis and detect dispersion syndromes of the species of a granite-gneiss outcrop, situated in the semi-arid tropic in Northeastern Brazil. This study sought to answer the following questions: a) what is the degree of similarity/ dissimilarity of this area in relation to other areas with rocky environments already studied in Northeastern Brazil?; b) do the dispersal syndromes corroborate the patterns found in outcrops of other arid or semiarid regions?
Material and Methods
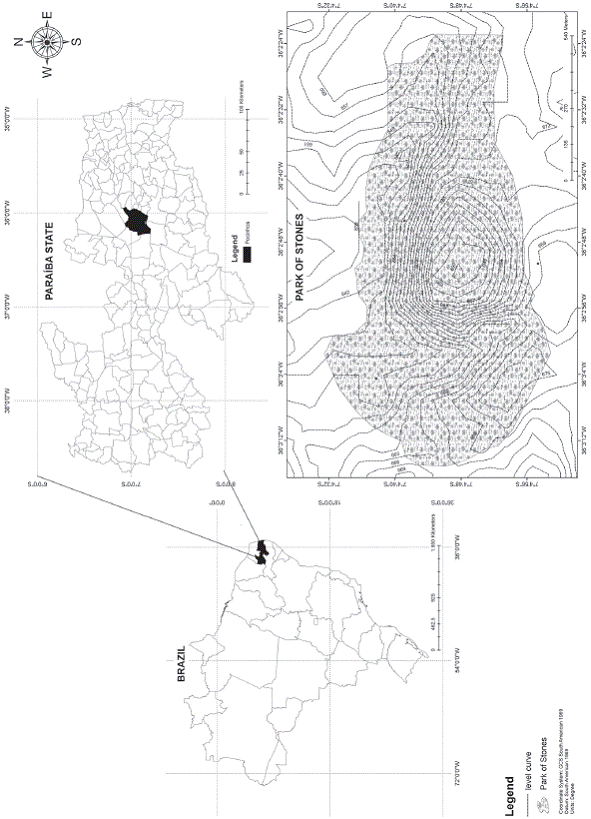
Study area: The outcrop selected for this study is located in the municipality of Pocinhos, Agreste mesorregion of Paraiba state, in the semi-arid Northeastern Brazil. It is located in a private estate with approximately 55 ha known as Parque das Pedras (07° 05' 14'' S - 36° 03' 56'' W), and, although it is open to tourism, it is visibly preserved (Fig. 1). The Agreste is one of mesorregion of the state of Paraiba. It is characterized as a climatic transition area where winds heated in the depression zone ascends and cools up, causing rainfall (Rodriguez, 2000), with annual average rates ranging from 600 to 1 000 mm (AESA, 2014). According to the new updated classification of Koppen-Geiger (Peel, Finlayson, & McMahon, 2007), the climate in this region is characterized as BSh, hot semi-arid, and the annual precipitation rates are close to littoral rates (Rodriguez, 2000). It presents well-developed soils with clayey characteristics (Oliveira & Oliveira, 2008). The vegetation of this unit consists of semidecidu-ous and deciduous forests, particular to areas of Agreste (Beltrao et al., 2005).
Floristic survey: For the record of the species that occur in the outcrop, 16 excursions were made in the period from December 2012 to November 2013, consisting of monthly collections in the dry season and bimonthly collections in the rainy season throughout the study area. Fertile branches (flowers and/or fruits) of individuals belonging to all strata of vegetation were obtained (Fig. 2 and Fig. 3). The specimens were processed according to the usual techniques (Judd, Campbel, Kellogg, Stevens, & Donoghue, 2009) and led to dry in the Herbarium Manuel de Arruda Camara (ACAM) of the State University of Paraiba (UEPB), Campina Grande, Paraiba state, Brazil. The taxonomic identification was based on the taxonomic literature or by comparison with specimens, identified by experts, belonging to the collections of herbaria in the state of Paraiba (EAN, JPB) and in the Virtual Herbarium of the Brazilian Flora (REFLORA). Acronyms of the herbaria were mentioned according to Holmgren, Holmgren and Barnett (1990). Specimens and scanned images also were sent to confirmation and/or identification by experts in different taxonomic groups in Brazil. Exsiccatae were incorporated into the collection of the Herbarium Manuel de Arruda Cámara (ACAM). The species were listed in alphabetical order by family, according to the 2). The spelling of the names of species and their respective authors were found in the database of the Missouri Botanical Garden (Tropicos, 2013) and in the online Species List of Brazilian Flora (Forzza et al., 2014). This work has been included exclusively the native species and for this reason, the only exotic species recorded in the study area was not included in the floristic list.

Fig. 2 Species recorded in the studied rocky outcrops: A) Jatropha mollissima. B) Tacinga inamoena. C) Cyrtopodium holstii. D) Lantana camara. E) Angelonia campestris. F) Lafoensia pacari.
Dispersal syndromes: To determine dispersal syndromes, young and mature fruits of almost all species recorded in outcrop were collected, except for Lippia grata Schauer (Verbenaceae), Justicia aequilabris (Nees) Lindau (Acanthaceae), Mitracarpus salzman-nianus DC., Staelia virgata (Link ex Roem. & Schult.) K. Schum. (Rubiaceae), and Evolvulus ovatus Fernald (Convolvulaceae) and their syndromes' determinations were conducted through taxonomic literature consulted or with expert assistance. For the other species, the syndromes were classified according to the work by Van der Pijl (1972), being classified into three categories: I-Anemochoric: when diaspores presented some sort of appendix with wings, feathers or in the form of dust; Il-Zoochoric: when some attractive element, such as, for example, strong colors and/or a food source in its diaspores, is presented, as well as those with adhesive structures in the form of hooks, bristles, spines or mucilage; III-Autochoric: when the diaspores showed no characteristic that allowed their classification in the previous two categories.

Fig. 3 Species recorded in the studied rocky outcrops: A) Jatropha mollissima. B) Pilosocereus pachycladus. C) Cyrtopodium holstii. D) Marsdenia caatingae. E) Angelonia campestris. F) Sapium argutum.
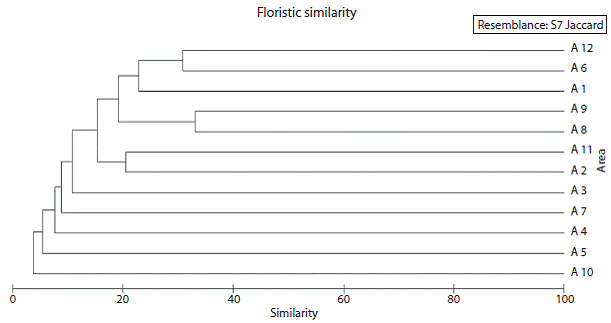
Fig. 4 Dendrogram of floristic similarity linking the study area with 11 studies on rocky outcrops and other areas of the surrounding matrix (Caatinga vegetation). A1 - Present study, A2 - Gomes et al. (2011), A3 - Porto et al. (2008), A4 -Araujo et al. (2008), A5 - Franga et al. (2005), A6 - Tolke et al. (2011), A7 - Gomes & Alves (2009), A8 - Machado-Filho (2011), A9 - Lima (2012), A10 - Araujo et al. (2005), A11 - Barbosa et al. (2005), A12 - Araujo et al. (Ined. 2013).
According to Van der Pijl (1972), the auto-choric dispersion may occur in two main forms: active, when the plant expels the diaspores with a ballistic action associated with dehiscence of the fruit; and passive, when they are carried by the movement of the sediment or by small animals. In the latter, species with explosive dispersion or by gravity (barochoric) were grouped. Of the species recorded in the area, two did not have their dispersal syndromes identified due to the absence of fruiting material or because they had immature fruits. All fruits collected in the study area were classified according to Spjut (1994), and, for species that did not fruit during fieldwork, the classification was determined based on the specific literature and with expert advice.
Similarity analysis: To calculate the flo-ristic similarity, originally a list consisting of the compilation of 11 floristic references was compiled using Excel software version 7.0, generating a presence/absence binary matrix with all elements identified at the species level. The selected studies included studies in rocky environments inserted in areas of Caatinga in states from the Northern and Southern portion of Northeast of Brazil (Table 1).
To avoid repetition and synonyms, the current nomenclature of each species was used. The dendrogram was obtained with the Primer 6.0 software, using the Jaccard index, based on the Unweighted Pair Group Method using Arithmetic averages (UPGMA) method.
Results
Floristic: In the study area, 161 species belonging to 127 genera and 50 families of angiosperms were recorded (Table 2, Fig. 2 and Fig. 3). Among the species, five were identified up to the genus level. The most representative family was Fabaceae (24 spp.), followed by Asteraceae and Convolvulaceae, with 12 and 11 species, respectively; together, they correspond to 29 % of all recorded species.
Table 1: Floristic studies in semi-arid Northeastern Brazil compiled from the elaboration of the matrix of presence/absence of species
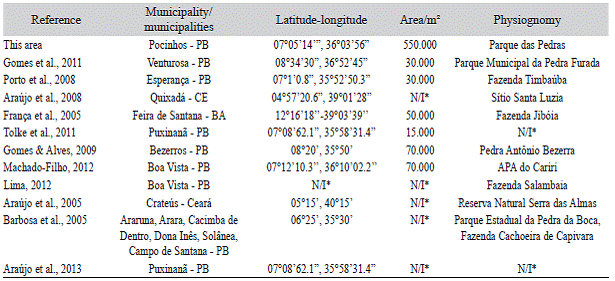
Table 2: Floristic list containing names of families and species, types of fruits and their dispersal syndromes of species recorded in the study area, Parque das Pedras, Pocinhos, Paraiba state, Brazil

Allophylus quercifolius (Mart.) Radlk (Sap-indaceae) and Lafoensia pacari A. St.-Hil. (Lythraceae) constituted new records for the state of Paraiba. Regarding monocots, the Bro-meliaceae family should be highlighted; it was represented by five genera and seven species, being Tillandsia the most representative genus, with three species. Within Orchidaceae, represented by five species and three genera, Cyrto-podium is the most representative genus with two species. Poaceae, Cyperaceae and Araceae totaled 3.1 % of the total species recorded.
With regard to habits, the herbaceous stratum was the most representative, with 54 % of the total composition of the outcrop. Shrub (15 %) and subshrubs (12 %) components make up together 27% of all recorded species, and vines and epiphytes total 12.5 %. Three species of hemiparasites belonging to two families were recorded: Phoradendron affine Pohl ex DC., Phoradendron piperoides (Kunth) Trel. (Santalaceae) and Struthanthus margin-atus (Desr.) Blume (Loranthaceae).
Similarity analysis: The similarity dendrogram showed the formation of well-defined clusters and a block whose areas appear forming similar subgroups among them (Fig. 4). The first group consists of rocky outcrops located in the municipality of Puxinana (Araujo et al., 2013, unpublished data- A12, Tolke, Silva, Pereira & Melo, 2011 - A6) with an index of floristic similarity between areas of J = 30. The studied area (A1) demonstrated greater affinity with this group, being presented, in the dendrogram, as its subgroup. This floristic relationship can be explained by the fact that the whole work was done in areas geographically close together. The second group comprises the works of Machado-Filho (2011) - A8 and Lima (2012) - A9 (both with unpublished data), showing the highest similarity index (J = 33.2), being these works developed in a region called Cariri, located in Northeastern Brazil. The third group comprises the studies of Barbosa, Lima, Agra, Cunha & Pessoa (2005) - A11, held at Eastern Curimatau of Paraiba state and Gomes, Costa, Rodal & Alves (2011) - A2, in the semi-arid region of the state of Pernambuco.
Some areas settled in the Caatinga domain formed subgroups apart from the rest. These areas include rocky habitats (Franga et al., 2005; Araújo et al., 2008; Porto, Almeida, Pessoa, Trovao, & Félix, 2008; Gomes & Alves, 2009), and one study was conducted in the backlands ("sertao") of Ceará State by Araújo, Costa, Figueiredo & Nunes (2005), being the most dissimilar (J = 0.6) among selected studies when compared to this study.
Dispersal syndromes: With respect to the dispersion mode of the diaspores, the results showed a predominance of abiotic syndromes, where authocory was the most representative totaling 51.5 % (83 spp.) of total species. This mode of dispersion is the result of the plant's own mechanisms through explosive dehiscence of dried fruits or by gravity. The second most representative syndrome is anemochory, with 26.7 % (43 spp.) of the recorded species, followed by zoochory, comprising 22.3 % (36 spp.). In this category, plants depend on animals to disperse as far as possible from the mother-plant.
Discussion
Bromeliaceae, Poaceae and Cyperaceae are very representative families in outcrops in South America (Sarthou & Villiers, 1998). However, Poaceae and Cyperaceae, in this study, were represented by two species each. Melinis minutiflora P. Beauv. (Poaceae) was one of the species found in the outcrop, but, because it is an exotic and invasive species, it was not included in the floristic list. Bro-meliaceae is represented by seven species, and the genera Encholirium and Tillandsia occur throughout the outcrop. This pattern of occurrence for the above mentioned genera was also observed by Franga, Melo and Gongalves (2006).
Corroborating the works of Araújo et al. (2005), Barbosa et al. (2005), Lima, Sampaio, Rodal and Araújo (2009) and Santos & Melo (2010), Fabaceae was the family best represented, with 24 species, corresponding to 15 % of species richness in the studied area. Considered the most taxonomically diverse of the Caatinga, this family is the third largest among angiosperms, whose economic importance makes it even more known (Judd et al., 2009).
Asteraceae was the second best represented family in number of species and its wide distribution, particularly common in open areas (Souza & Lorenzi, 2012), illustrates the predominance of its species in its different aspects, represented in the works of Lima (2012), Neves & Conceigao (2007) and Zappi et al. (2003).
Cactaceae, with five species, proved to be frequent in the study area, showing species with herbaceous to arborescent habits. Among the species found, Melocactus zehntneri (Britton & Rose) Luetzelburg is noteworthy because it is threatened by exploration and/or by anthropic pressure on their habitats (Fabricante, Andrade, & Marques, 2010). According to these authors, M. zehntneri plays a very important ecological role in the succession process, being able to colonize xeric environments and make them less harsh.
With regard to the habit, the tree component was the least representative, comprising 9 % of the total and, in that sense, Caiafa and Silva (2007) and Safford and Martinelli (2000) note that some outcrops showed shallow rock fractures, forming microenvironments characterized by the accumulation of 5 to 12 cm of soil, thus limiting the development of this stratum.
In the dendrogram, the highest value was found among the areas studied by Machado-Filho (2012) and Lima (2012), both settled in the Environmental Protection Area (EPA) of Cariri, in the semi-arid region of the state of Paraiba, Brazil. This close relation was expected, given that the studies were developed in the same geographical area between the municipalities of Cabaceiras and Boa Vista, Paraiba, whose climate and soil conditions are very similar and where rocky outcrops predominate.
The studies conducted in rocky environments settled in the city of Puxinana, in Agreste mesorregion of Paraíba (Tólke et al., 2011; Araújo et al., 2013, unpublished data), showed a floristic relation (J = 31) closer with each other rather than with the studied area (J = 19). However, it was expected, because they are geographically nearby areas, that the similarity value between them would be higher than the one recorded (Kunz, Ivanauskas, Martins, Silva, & Stefanello, 2009; Oliveira & Nelson, 2001). This dissimilarity can be explained partly by species recording, which occurred only in this study: Chloroleucon dumosum (Benth.) G.P. Lewis, Cuphea impatientifolia A. St.-Hil., Helicteres eichleri K. Schum., Jacquemontia corymbulosa Benth. andXimenia americana L.
The areas studied by Barbosa et al. (2005) and Gomes et al. (2011) formed a subgroup with a similarity value of J = 20, both located in the Agreste of the states of Paraíba and Pernambuco, respectively, showing weather conditions with erratic rainfall, varying from 664 mm to 1 054 mm, and similar frequency of rock formations. The most representative families in the two areas were Fabaceae and Euphorbiaceae, common in areas of caatinga (Cardoso & Queiroz, 2007; Sátiro & Roque, 2008), confirming the results obtained in the area under study.
The other areas had low similarity values (J < 20), in particular the study by Araújo et al. (2005), conducted in the Natural Reserve of Serra das Almas, CE, where three different formations were explored, contemplating areas from dense scrub vegetation to thorny deciduous trees and shrubs, with a dissimilarity between the flora of the surrounding matrix and the flora that is established on rocky outcrops. This vegetation is probably severely influenced by strict environmental aspects, demanding a greater adaptation of individuals, which contributed to this dissimilarity (Porembski & Barthlott, 2000).
Species such as Jatropha mollissima (Pohl) Baill. and Cnidosculus urens (L.) Arthur -Euphorbiaceae-, recorded in this study, have a passive authocory mechanism (Leal, 2003). According to the same author, seeds released by gravity may be secondarily carried to other areas by ants. Sapium argutum (Müll. Arg.) Huber (Euphorbiaceae) presents voluminous seeds, with vivid colors, and, although not mentioned in the study, it is possible that its seeds also present passive authocory by dispersion done by ants or even by sediment carrying along the outcrop.
The anemochoric syndrome includes 26.7 % of the recorded species' total, corroborating the data exposed by the literature, where areas of greatest exposure with low water levels show a predominance of abiotic vectors (Butler, Green, Lamb, McDonald, & Forster, 2007; Barbosa, Silva, & Barbosa, 2002; Griz & Machado, 2001; Machado, Barros, & Sampaio, 1997). Van der Pijl (1972) points out those taxa with wind dispersal present various structures that facilitate its spread. In addition, regarding anemochoric species collected in the study area, winged and feathery seeds were the most frequently observed.
The microclimate aridity of these rocky islands in relation to the surrounding area reflects the predominance of abiotic syndromes, as they represent habitats with a greater exposure to winds and sunlight (Araújo et al., 2008). The zoochorous syndrome amounted to 22.3 %, with most species that falls into this category being collected in wetlands surrounding the studied rocky outcrop. These conditions reflect what was found in the study by Silva and Rodal (2009), after the detection of dispersal syndromes in three areas with different rainfall regimes in the state of Pernambuco, Northeastern Brazil. It can be inferred that there is a gradual change in the spread spectrum of wetlands, dominated by species with zoochorous syndromes.
Thus, conducting floristic surveys associated with ecological aspects, such as the recognition of syndromes, provide a better understanding of the dynamics of species colonization, mainly with regard to the Caatinga vegetation in Northeastern Brazil. The results obtained in this study, compared to other similar studies (Araújo et al., 2008; Barbosa et al., 2002; Lima, 2012; Silva & Rodal, 2009), showed that abiotic syndromes (anemochory or authocory) are prevalent in outcrops and dry forests, supporting patterns found in outcrops in other regions of arid or semi-arid climates. Similar edaphic-climatic conditions are those that best explain the floristic similarity between the compared areas, especially those closer to each other. The combination of short distances and the predominance of abiotic syndromes suggest this affinity in the floristic composition, as observed in the works by Machado-Filho (2011) and Lima (2012).

