











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkEl ámbito hogareño es el área dentro de la cual un individuo se mueve (Rose, 1982), con la finalidad de adquirir recursos que incrementen su supervivencia y reproducción, entre ellos: alimento, refugio, apareamiento, sitios de percha y anidación (Hirth, 1963; Gutiérrez & Ortega, 1985). Se ha documentado que, debido a factores bióticos y abióticos, el ámbito hogareño en lagartijas puede variar inter e intraespecíficamente.
Los estudios intraespecíficos indican que el tamaño del ámbito hogareño puede variar de acuerdo al sexo y a la condición reproductora. En este contexto los machos incrementan el tamaño de su ámbito hogareño durante la estación de reproducción (Ruby, 1978). El ámbito hogareño también puede disminuir si la densidad poblacional o la cantidad de alimento aumenta (Stamps, 1983; Ruby & Dunham, 1987). También se ha registrado variación interanual e interpoblacional en el tamaño del ámbito hogareño (Hulse, 1981; Ruby & Dunham, 1987). En este sentido, debido a restricciones fisiológicas, los individuos disminuyen su ámbito hogareño cuando el año o el hábitat son extremos (Hulse, 1981; Ruby & Dunham, 1987). Los estudios entre especies con el uso de métodos comparativos, sugieren que el tamaño del ámbito hogareño está relacionado con el tamaño corporal y el sexo (Perry & Garland, 2002), donde las especies de mayor tamaño y los machos tienen ámbitos hogareños más amplios (respecto a las especies pequeñas y a las hembras). El tamaño del ámbito hogareño también se puede relacionar con el nivel de actividad de los individuos, debido a que las especies más activas necesitan ámbitos hogareños más amplios para satisfacer demandas energéticas mayores (Verwaijen & Van Damme, 2008). Adicionalmente, diversos estudios han documentado que el ámbito hogareño también se relaciona con el modo de forrajeo. Se considera que las especies de forrajeo amplio tienen un ámbito hogareño amplio (McCoy, 1965; Hulse, 1981; Rowland, 1992; Verwaijen & Van Damme, 2008).
En México existen diez especies parte-nogenéticas (Flores-Villela, 1993) del género Aspidoscelis (Reeder, Cole, & Dessauer, 2002), tres de ellas son endémicas de la Península de Yucatán y son conocidas colectivamente como grupo A. cozumela (Wright, 1993). Un híbrido entre un macho de Aspidoscelis deppii Wiegmann, 1834 y una hembra de Aspidoscelis angusticeps Cope, 1878 originó a la lagartija partenogenética Aspidoscelis maslini Taylor & Cooley, 1995. Cambios morfológicos y citoge-néticos en una hembra de A. maslini originaron a la lagartija partenogenética Aspidoscelis cozumela Gadow, 1906 (Taylor & Cooley, 1995; Manríquez-Morán, Villagrán-Santa Cruz, & Méndez-de la Cruz, 2000; Manríquez-Morán, 2002; Elizalde-Rocha, 2007).
La lagartija A. cozumela es microendémica de Isla Cozumel, es ovípara, de forrajeo amplio, insectívora, con valores bajos en supervivencia, tamaño de nidada y variación genética (Lee, 1996; Hernández-Gallegos, 2004; Manríquez-Morán, Villagrán-Santa Cruz, & Méndez-de la Cruz, 2005; Manríquez-Morán & Méndez-de la Cruz, 2008; Manríquez-Morán, Méndez-de la Cruz, & Murphy, 2014). Diversos estudios han documentado que A. cozumela tiene elevada dependencia por las playas arenosas, donde se ha registrado el mayor número de individuos y poblaciones (Lee, 1996; Hernández-Gallegos, Manríquez-Morán, Méndez, Villagrán, & Cuellar, 1998; Manríquez-Morán, 2002; Hernández-Gallegos, 2004; Díaz de la Vega-Pérez, Jiménez-Arcos, Manríquez-Morán, & Méndez-de la Cruz, 2013). La IUCN cataloga a A. cozumela en la categoría de preocupación menor. Sin embargo, la información ecológica y biogeográfica disponible indica que esta lagartija se encuentra amenazada en la isla (López-González & González-Romero, 1997; Manríquez-Morán & Méndez-de la Cruz, 2008). En este trabajo se describe el ámbito hogareño de A. cozumela, con el objeto de: 1) saber si esta especie partenogenética presenta un ámbito hogareño amplio (típico en especies de forrajeo amplio), 2) evaluar el solapamiento intraespecífico, 3) comparar con otras especies de Aspidoscelis (gonocóricas y parteno-genéticas) y con otras lagartijas que ocupan hábitats costeros, y 4) discutir los factores bióticos y abióticos que moldean su tamaño. Adicionalmente, la información del ámbito hogareño de A. cozumela se incorpora desde una perspectiva de conservación, ya que los hábitats costeros en Isla Cozumel se encuentran severamente amenzados.
Material y Métodos
Zona de estudio: La zona de estudio para el análisis del ámbito hogareño de A. cozumela fue una playa que se localizó a 14 km al sureste de San Miguel en Isla Cozumel (20°24'33" N - 86°51'31" W). La isla está localizada a 17.5 km de la costa Noreste de Quintana Roo en la Península de Yucatán. La temperatura promedio de Isla Cozumel es de 25.5 oC y su precipitación anual acumulada es de 1 607.1 mm (García, 2004). Con base en los datos climatológicos de la zona de estudio y de acuerdo con trabajos previos (Yáñez-Arancibia & Day, 1988; Lee, 1996; Gómez-Ramírez & Reséndiz-Espinosa, 2002), en la Península de Yucatán pueden identificarse tres estaciones: sequía (finales de marzo-abril), lluvias (mayo-septiembre) y "nortes" (octubre-principios de marzo). Es común que de agosto a octubre se presenten huracanes de gran magnitud que modifican los ecosistemas y el entorno costero (Gómez-Ramírez, 1998). La vegetación en la playa es halófila típica de duna costera, la cual esta compuesta de plantas de forma de vida erecta o postrada (arbustivas y herbáceas), que se desarrollan en altos niveles de insolación y salinidad y fuertes vientos (Téllez-Valdez, Cabrera-Cano, Linares-Mazari, & Bye, 1989). Cabe señalar que en Isla Cozumel existen diferentes vertebrados endémicos, entre ellos mamíferos (Wilson & Reeder, 1993), aves (Howell & Webb, 1995) y la lagartija parteno-genética A. cozumela (Taylor & Cooley, 1995).
Dimensión del área de estudio y censos realizados: El área de estudio fue de 4 000 m2 (20 x 200 m) y se delimitó en subcuadrantes de 10 x 10 m a través de estacas marcadas con diferente número y letra cada una. El esfuerzo de muestreo consistió en nueve censos que involucraron un método de captura-marcaje-recaptura. Cada censo duró de dos a cuatro días y se realizaron entre las 09:00 h y las 18:00 h durante 1999 (marzo 15-16, mayo 16- 17, julio 28-29 y octubre 4-5), 2000 (marzo 18-20, julio 19-21, octubre 17-19) y 2001 (abril 17- 20 y septiembre 18-20). Los muestreos fueron realizados sistemáticamente (Sánchez, 2011) y la consideración de aspectos bióticos y abióticos relevantes al momento de evaluar el ámbito hogareño en lagartijas (Rose, 1982), contribuyeron en la detección de tendencias y denotaron un esfuerzo de muestreo adecuado (Sánchez, 2011). En este sentido, los censos realizados cubrieron las épocas de sequía, lluvias y "nortes" que existen en Isla Cozumel, la temporada de reproducción registrada en A. cozumela (marzo-agosto) y dos generaciones completas en la población.
Captura de individuos y registro de información: En cada censo, la zona de estudio fue recorrida sistemáticamente cuadrante por cuadrante durante varias ocasiones, hasta cubrirla por completo. Todas las lagartijas encontradas fueron capturadas con una trampa de malla de alambre, marcadas permanentemente por ectomización de falanges (Tinkle, 1967) y liberadas en el sitio de captura. Es importante resaltar que la ectomización no afecta el desempeño de los individuos (Paulissen & Meyer, 2000; Borges-Landáez & Shine, 2003). Con el fin de minimizar las capturas repetidas dentro de cada censo, cada individuo se marcó con un punto de pintura indeleble en la base de la cola. De cada hembra se registraron los siguientes datos: fecha, hora del día, longitud hocico-cloaca (LHC) al milímetro más cercano (utilizando una regla plástica) y su localización en el área de estudio por bi-coordenadas.
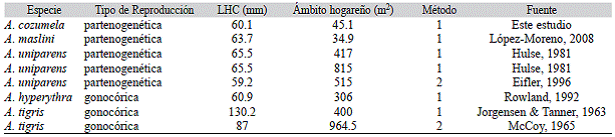
Ámbito hogareño: Para el cálculo del ámbito hogareño de A. cozumela se consideraron a las hembras con tres o más recapturas. El ámbito hogareño se calculó con el método del polígono convexo (Rose, 1982) mediante el programa McPaal (Stüwe & Blohowiak, 1985). El método del polígono convexo es uno de los métodos más utilizados para evaluar el ámbito hogareño en lagartijas (Rose, 1982) y se ha utilizado para obtener el ámbito hogareño de diferentes especies de Aspidoscelis (Perry & Garland, 2002). Para realizar comparaciones con el ámbito hogareño de A. cozumela, de estudios previos se obtuvieron los ámbitos hogareños de especies gonocóricas y parteno-genéticas de lagartijas del género Aspidoscelis.
Solapamiento del ámbito hogareño: Para el cálculo del solapamiento del ámbito hogareño (Haenel, Smith, & John Alder, 2003) se consideró: 1) el número de hembras que se solapaban, 2) el número de hembras cuyo ámbito hogareño se solapaba con determinado ámbito hogareño, 3) el área del ámbito hogareño que se solapaba con la(s) hembra(s) circundante(s) y 4) la proporción que se solapaba con respecto al ámbito hogareño total.
Adicionalmente, se correlacionó la LHC promedio de las hembras con su ámbito hogareño usando el coeficiente de correlación de Pearson. La LHC promedio se obtuvo al promediar las diferentes LHC registradas para cada hembra. Los análisis se realizaron en Stat-graphics Plus 5.0 y se utilizó un alfa de 0.05.
Resultados
Ámbito hogareño: Todas las hembras capturadas durante su último censo fueron adultas, lo anterior con base en la talla a la madurez sexual registrada previamente (56 mm de LHC). De acuerdo con el intervalo entre la primera y la última captura, 14 hembras se capturaron por un período no mayor a un año, cinco hembras por un período de 1-1.5 años y una hembra por un período de 1.5-2 años. El intervalo entre la primera y la última captura tuvo un promedio de 303.3 días (EE = 33.2 días, intervalo = 134 - 715 días, n = 20). Se obtuvieron un total de 20 ámbitos hogareños (Fig. 1), 14 de ellos fueron de tres recapturas, cinco de cuatro recapturas y uno de seis recapturas. No se encontró diferencia en el ámbito hogareño entre las hembras con tres y cuatro recapturas (t de student, p = 0.0578). El ámbito hogareño tuvo un promedio de 45.1 m2 (EE = 14 m2, intervalo = 1.5-287.5 m2, n = 20). No se encontró relación entre la LHC promedio de las hembras y el tamaño de su ámbito hogareño (p = 0.9229, n = 20). Sin embargo, se encontró relación cuando no se consideraron los individuos con valores extremos (i. e., hembras con ámbito hogareño > 100 m2, n = 2), las hembras más grandes tuvieron ámbitos hogareños más amplios (r = 0.61, p = 0.0072, n = 18, Fig. 2). Los resultados del ámbito hogareño de A. cozu-mela, indican que su área es la más pequeñas cuando se consideran trabajos publicados con especies partenogéneticas y gonocóricas de Aspidoscelis (Cuadro 1).
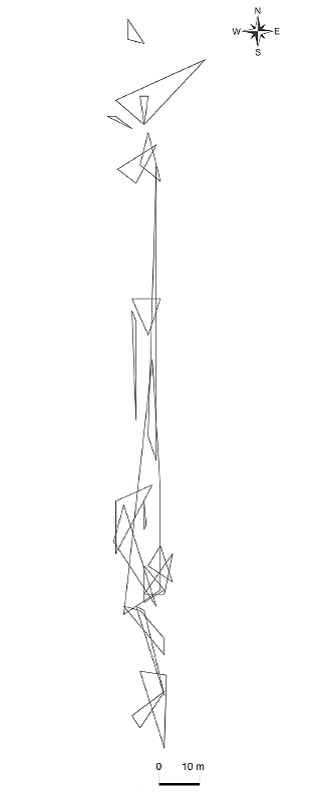
Fig. 1: Ambitos hogareños de la lagartija partenogenética Aspidoscelis cozumela en Isla Cozumel. Cada polígono representa el área utilizada por una hembra (n = 20).
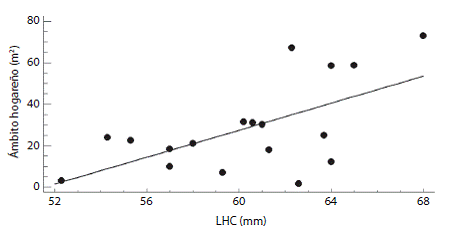
Fig. 2: Relación de la LHC con el tamaño del ámbito hogareño en la lagartija partenogenética Aspidoscelis cozumela en Isla Cozumel.
Solapamiento del ámbito hogareño: Durante los censos realizados se registró que 14 hembras mantuvieron un solapamiento espacial con al menos una hembra. El ámbito hogareño de estas hembras se solapó con 1.6 hembras (EE = 0.2 hembras, intervalo = 1 - 4 hembras, n = 14). Cada hembra se solapó en promedio 8.0 m2 (EE = 1.4 m2, intervalo = 1.1 - 19.7 m2, n = 22), que representó el 22.9% (EE = 5.7%, intervalo = 0.37 - 100%, n = 22) de su ámbito hogareño.
Discusión
La evidencia del estudio de captura-marca-je-recaptura indica que la lagartija partenoge-nética A. cozumela es marcadamente residente en los hábitats de playa de Isla Cozumel. Las hembras mantienen ámbitos hogareños significativamente reducidos, ya que el 90% de ellas presentaron ámbitos hogareños que fueron < 73 m2. El ámbito hogareño de A. cozumela es el más pequeño que se ha publicado en el género Aspidoscelis y difiere de lo registrado en otras especies partenogenéticas y gonocó-ricas de Aspidoscelis, donde se han registrado ámbitos hogareños relativamente más amplios.
Cuadro 1: Ámbito hogareño de hembras de especies partenogenéticas y gonocóricas del género Aspidoscelis
Ámbitos hogareños reducidos en hábitats de playa o costeros, también han sido registrados en especies de forrajeo amplio como la lagartija partenogénetica A. maslini (34.9 m2, López-Moreno, 2008) y en hembras de Ameiva qua-drilineata Hallowell, 1861 (187.7 m2, Teiidae, Hirth, 1963), así como en especies de forrajeo pasivo como en hembras de Liolaemus lutzae Mertens, 1938 (22.3 m2, Liolemidae, Rocha, 1999) y Liolaemus multimaculatus Duméril & Bibron, 1837 (21.31 m2, Liolemidae, Kaco-liris, Williams, Ruiz-De Arcaute, & Cassino, 2009). Un ámbito hogareño reducido en hábitats de playa o costeros, puede ser producto de una amplia similitud en las características ambientales que imperan en estos hábitats. En el caso particular de A. cozumela, puede ser adicionalmente una herencia de A. maslini, con quien también comparte amplia semejanza en características de historia de vida (Hernández- Gallegos, 2004).
El ámbito hogareño puede variar de acuerdo con diferentes características bióticas y abióticas (Ruby, 1978; Hulse, 1981; Stamps, 1983; Ruby & Dunham, 1987; Perry & Garland, 2002; Verwaijen & Van Damme, 2008), entre ellas el tamaño corporal y el modo de forrajeo han resultado buenos estimadores del tamaño del ámbito hogareño (Perry & Garland, 2002; Haenel et al., 2003; Verwaijen & Van Damme, 2008). Como se ha registrado en hembras de diversas especies de lagartijas (Schoener & Schoener, 1982; Morales-Mávil, Vogt, & Gadsden-Esparza, 2007), el ámbito hogareño de A. cozumela se incrementa con la LHC. Para el caso de A. cozumela, esta tendencia se presenta en la mayor parte de la población, excepto en las hembras con los dos ámbitos hogareños más grandes (101 m2 y 287.5 m2). Cabe señalar que, para obtener valores más precisos, la exclusión de datos extremos en ámbitos hogareños se ha realizado previamente (Kacoliris et al., 2009). Un tamaño corporal mayor involucra una demanda mayor de recursos que pueden cubrirse con un ámbito hogareño más grande (Haenel et al., 2003). Desde la concepción inicial de los dos modos de forrajeo (Pianka, 1966), las especies del género Aspidoscelis se han considerado como especies de forrajeo amplio (Pianka, 1966; Vitt & Breitenbach, 1993), que activamente buscan a sus presas y que presentan ámbitos hogareños relativamente amplios (McCoy, 1965; Hulse, 1981; Rowland, 1992). La lagartija parteno-genética A. cozumela es de forrajeo amplio (Lee, 1996) y los resultados de este estudio se contraponen con las predicciones teóricas que establecen que las especies de forrajeo amplio tendrán ámbitos hogareños amplios que cubran su alta energía metabólica, alto nivel de actividad y requerimentos nutricionales (Verwaijen & Van Damme, 2008).
El ámbito hogareño reducido de A. cozu-mela puede ser explicado por los beneficios de la calidad térmica de las playas que parecen favorecer la densidad poblacional elevada en esta especie. Se ha registrado que A. cozumela ocupa una variedad de hábitats en Isla Cozu-mel (López-González & González-Romero, 1997; Manríquez-Morán & Méndez-de la Cruz, 2008), entre ellos: playas, matorral costero, plantaciones de coco y bosque tropical semideciduo (López-González & González-Romero, 1997). Los datos actuales indican que el hábitat óptimo para A. cozumela son las playas, ya que es en donde se ha registrado la mayor densidad y el mayor número de poblaciones (Lee, 1996; Hernández-Gallegos et al., 1998; Manríquez-Morán, 2002; Hernández-Gallegos, 2004; Díaz de la Vega-Pérez et al., 2013). Algunos autores proponen que la presencia de una especie en un hábitat depende en gran medida de la adquisición de recursos (Rose, 1982; Gutiérrez & Ortega, 1985; Perry & Garland, 2002). Por ello, los atributos bióticos y abióticos de un hábitat son relevantes en la frecuencia de uso de un sitio. Las características bióticas y abióticas de las playas donde se encuentra A. cozumela son: vegetación arbustiva y herbácea (generalmente no más de un metro de altura) que crea hábitats abiertos con elevada insolación. Estos ambientes en Isla Cozumel presentan la mejor calidad térmica entre 13 ambientes (principalmente playas) de la Península de Yucatán, donde se encuentran las tres especies partenogenéticas del grupo A. cozumela y sus especies parentales (Díaz de la Vega-Pérez et al., 2013). Debido a estas características, posiblemente las playas ofrezcan a la lagartija A. cozumela los microhabitats más propicios para termorregular y obtener la temperatura corporal elevada necesaria para realizar las diferentes actividades (38 oC, Díaz de la Vega-Pérez et al., 2013). Una situación similar ha sido registrada en otras especies de la familia Teiidae, donde los individuos alcanzan una temperatura corporal elevada en las playas (Hirth, 1963; Vitt, Zani, Caldwell, & Durtsche, 1993). Es posible que al centro del continente o la isla los individuos no obtengan los requerimientos de termorregulación y alimento necesarios, por ello los individuos prefieren permanecer en las playas aún bajo condiciones climáticas severas (Hirth, 1963).
Los huracanes modifican (entre agosto y octubre) los ecosistemas y el entorno costero de la Península de Yucatán (Gómez-Ramírez, 1998). Se ha registrado que estos eventos tienen su mayor incidencia en la región noreste de la península (Escobar-Nava, 1986; Konrad, 1996), donde se encuentra Isla Cozumel. Estos fenómenos naturales remueven la arena y la vegetación de las playas (Hernández-Gallegos et al., 1998) y al ser recurrentes, pueden no sólo promover grandes cambios ambientales sino también cambios en la biota terrestre de la región. Se desconoce si A. cozumela usa otros hábitats como refugio durante los huracanes, si este fuera el caso, las hembras que ocupan playas en Isla Cozumel deberían modificar su ámbito hogareño significativamente, ya que en playas son altamente sedentarias.
Como se ha registrado en hembras de diferentes especies (Stamps, 1983), la densidad poblacional influye inversamente en el tamaño del ámbito hogareño (Ruby & Dunham, 1987; Haenel et al., 2003). Menezes y Rocha (2013) estudiaron diferentes poblaciones costeras de cinco especies de la familia Teiidae y propusieron tres tipos de poblaciones de acuerdo con su densidad: poblaciones con densidad pobla-cional baja (hasta cinco individuos/hectárea), poblaciones con densidad poblacional intermedia (entre cinco y diez individuos/hectárea) y poblaciones con densidad poblacional alta (más de diez individuos/hectárea). El grado de conservación del hábitat se relacionó directamente con la densidad poblacional. Mitchell (1979) estudió la densidad poblacional en cuatro especies de Aspidoscelis (dos partenogenéticas: Aspidoscelis uniparens Wright & Lowe, 1965 y Aspidoscelis sonorae Lowe & Wright, 1964 y dos gonocóricas: Aspidoscelis inornata Baird, 1858 y Aspidoscelis tigris Baird & Girard, 1852) y registró densidades poblacionales de menos de 20 individuos por hectárea. Cabe resaltar que las densidades registradas en A. uniparens y en A. tigris (Mitchell, 1979) y su ámbito hogareño relativamente amplio (de 415 a 815 m2 y de 400 a 964.5 m2, respectivamente), contrastan con los valores registrados en A. cozumela (45.1 m2 de ámbito hogareño y 122 hembras por hectárea, Hernández-Gallegos, 2004). En este sentido, la densidad poblacio-nal marcadamente elevada en una lagartija de forrajeo amplio como A. cozumela en las playas de Isla Cozumel, puede influir en la reducción significativa de su ámbito hogareño. Probablemente, las hembras de A. cozumela en un ámbito hogareño reducido (ligeramente solapado con otras hembras), encuentren los recursos bióticos y abióticos necesarios para sobrevivir y reproducirse.
De acuerdo con estudios previos el grado de solapamiento en lagartijas puede variar inter e intrasexualmente (Lewis & Saliva, 1987; Rocha, 1999; Haenel et al., 2003). El solapamiento registrado en A. cozumela (22.9%) fue similar al registrado en las hembras de Ameiva exsul Cope, 1862 (22.4%, Lewis & Saliva, 1987). Sin embargo, los resultados de solapamiento en A. cozumela difieren cuando se comparan con la lagartija partenogenética Darevskia armeniaca Meheli, 1909, donde se registró solapamiento marcadamente elevado del ámbito hogareño entre las hembras (sola-pamiento con 6-7 hembras, involucrando más del 93% del ámbito hogareño, Galoyan, 2013). Una agresividad y competencia bajas (producto de su origen monoclonal y similitud genética) pueden explicar el solapamiento elevado en D. armeniaca (Galoyan, 2013). Cabe señalar que en D. armeniaca las hembras adultas catalogadas como residentes temporales, usan el mismo ámbito hogareño (104.8 m2) por 1-2 años y posteriormente lo cambian (su longevidad es > a cinco años, Galoyan, 2013). En el caso de A. cozumela, a diferencia de lo que sucede en D. armeniaca, la información indica que las hembras usan el mismo ámbito hogareño durante toda su vida. Lo anterior, se apoya en el hecho de que A. cozumela es anual en Isla Cozumel (4.5% de las hembras sobrevive un segundo año, Hernández-Gallegos, 2004) y que las hembras tienen una longevidad no mayor a dos años.
Dirzo et al. (2014) propusieron que las siguientes características pueden ser indicadoras de riesgo de extinción en vertebrados: distribución geográfica restringida, tasa reproductora baja, ámbito hogareño amplio y tamaño corporal grande. En este sentido la lagartija partenogenética A. cozumela tendría un riesgo de extinción intermedio, ya que cumpliría con los primeros dos criterios: es microendémica de Isla Cozumel y presenta el tamaño de nidada más bajo del género Aspidoscelis (Lee, 1996; Manríquez-Morán et al., 2005). Bajo condiciones naturales, el ámbito hogareño y tamaño corporal pequeños y otras características biológicas (i. e., doble potencial reproductor, madurez sexual temprana y tiempos generacionales cortos, Hernández-Gallegos, 2004), pueden promover su prevalencia en las playas y cola-teralmente disminuirían su riesgo de extinción. Sin embargo, las actividades antropogénicas en Isla Cozumel pueden incrementar significativamente dicho riesgo. En Isla Cozumel, las playas y su vegetación nativa se encuentran altamente amenazadas por el desarrollo de la infraestructura turística, de hecho por esta actividad la mayoría de las poblaciones del lado Oeste de la isla han sido extirpadas (Manríquez-Morán & Méndez-de la Cruz, 2008). Cabe señalar que, por actividades humanas, otros grupos de vertebrados como mamíferos y aves enfrentan severas problemáticas de conservación en Isla Cozumel (Martínez-Morales, 1999; Cuarón, Martínez-Morales, McFadden, Valenzuela, & Gompper, 2004). Es importante mencionar que solo la infraestructura turística (que elimina el componente vegetal) ha provocado la extirpación local de poblaciones en las playas, ya que A. cozumela parece estar bien adaptada a fenómenos naturales de gran magnitud como son los huracanes (Hernández-Gallegos, 2004), los cuales no promueven extirpaciones locales (Hernández Gallegos et al., 1998; Hernández-Gallegos, 2004).
Una situación similar a la que experimenta A. cozumela se ha registrado en la lagartija partenogenética Aspidoscelis rodecki McCoy y Maslin, 1962, donde las actividades turísticas y el desarrollo urbano han provocado extirpación local de poblaciones (Hernández-Gallegos, 2004; Elizalde-Rocha, Méndez-de la Cruz, Méndez-Sánchez, Granados-González, & Hernández-Gallegos, 2008). Es necesaria y urgente una regulación estricta de las playas en Isla Cozumel (y en el resto de la Península de Yucatán), que si bien promueva su aprovechamiento también procure la conservación de la flora y fauna nativas (Hernández-Gallegos, 2004). Resulta importante proteger la costa Este de la isla, que presenta una diversidad importante de vertebrados y constituiría el único refugio para especies microendémicas (como A. cozumela) que habitan preferentemente hábitats costeros.

