











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkLa infracomunidad es el conjunto de parásitos de diferentes especies que habitan en un individuo hospedero y constituye una unidad de estudio espacialmente discontinua en la que cada individuo hospedero es considerado una muestra aleatoria del hábitat (Holmes & Price, 1986; Bush, Lafferty, Lotz, & Shostak, 1997). Esto se debe a que hospederos de la misma especie adquieren parásitos de un conjunto local a partir de la ingestión de dietas similares y estar sometidos a similares condiciones y factores del hábitat.
El proceso de colonización de los parásitos a las infracomunidades puede ser secuencial si hay un patrón ontogenético de variación en la dieta de los hospederos. Esto puede modificar radicalmente la composición y abundancia de las especies de parásitos en hospederos que cohabitan (Díaz & George-Nascimento, 2002; Muñoz & Delorme, 2011). Por consiguiente, el tamaño de los hospederos se puede tomar como indicador de edad y composición de las infracomunidades; para encontrar evidencia de dichas variaciones ontogenéticas se requiere que el espectro de edad de los peces muestrea-dos sea lo suficientemente amplio como para que sean detectables los cambios (Rodríguez & George-Nascimento, 1996). Sin embargo, son escasos los estudios a nivel de población de hospederos en peces marinos, que abarquen todo el rango de edades o tallas, para así precisar el papel de la edad del hospedero en el proceso de colonización de la infracomunidad.
El cambio ontogénico de la dieta y/o hábi-tat por parte del huésped es un mecanismo que puede dar lugar a un patrón de anidamiento en la estructura de las infracomunidades (Poulin &Valtonen, 2001; Vidal-Martínez & Poulin, 2003) . No obstante, la adición de especies parásitas a una infracomunidad base formada por especies que no se ven afectadas por la talla del hospedero, podría crear también una estructura anidada, sin ocurrir un cambio estricto en la dieta (Zelmer & Arai, 2004). Si es importante identificar los patrones en la estructura de las comunidades parásitas, y detectar los procesos que los crean. Los procesos epidemiológicos, o capacidad de colonización/extinción de las especies parásitas, han sido sugeridos como causas de la estructura de ciertas comunidades parásitas (Morand, Rohde, & Hayward, 2002). Para explicar los patrones de distribución de las especies, Hanski (1982) planteó la hipótesis de especies núcleo (las cuales se encontrarán en muchas localidades en abundancia alta) y de especies raras (las cuales se encontrarán distribuidas en pocas localidades en abundancia baja).
La hipótesis de que el incremento del nivel de infección por parásitos aumenta con la edad del hospedero fue esbozada por Dogiel, Petrus-hevski and Polyanski (1958) y fue actualizada por Labropoulou, Machias, Tsimenides and Eleftheriou (1997). La hipótesis plantea que los cambios radicales en los niveles de para-sitación están supeditados a cambios estrictos en la dieta a partir de la talla de maduración y diferenciación sexual. Pero actualmente existen evidencias empíricas que sugieren la influencia de otros factores ecológicos del hospedero como la talla y edad (Johnson, Nelson, & Dock, 2004) , el tipo de hábitat (Muñoz, Grutter, & Cribb, 2006), la movilidad (Kennedy, 1990), y el comportamiento social y posible formación de cardúmenes (Luque, Mouillot, & Poulin, 2004). La exploración de estos factores (dieta, talla) en un contexto de análisis de la infra-comunidad a lo largo de la ontogenia puede indicar los procesos claves que determinan la formación de tales comunidades y su influencia sobre el hospedero.
Chromis cyanea (Poey 1860) es una de las especies marinas más comunes y abundantes de los arrecifes cubanos (Claro, Lindeman, & Parenti, 2001). La especie tiene un alto valor ornamental pero hasta la fecha no existen estudios que aborden el desarrollo de la infra-comunidad parásita a lo largo de su desarrollo ontogenético. Por este motivo nos proponemos como objetivos: (i) probar la hipótesis que las especies núcleo son responsables de la estructura en la infracomunidad y que por tanto aparecen antes que las especies raras y estocás-ticas; y (ii) determinar si existe relación de los descriptores de la infracomunidad parásita con el desarrollo ontogenético de C. cyanea inferido a partir de la longitud total.
Materiales y Métodos
Área de estudio y recolecta: El estudio se realizó en La Habana, costa noroccidental de Cuba, en las coordenadas (23°07'06.1" N - 082°26'56.4" W). Las colectas se realizaron durante el mes de abril del 2010 (estación de seca) con el objetivo de limitar el posible efecto de las variaciones estacionales en la infracomunidad parásita. Los ejemplares de C. cyanea fueron capturados procurando cubrir el máximo intervalo de talla descrito en la ontogenia de la especie; ello indica un valor de talla máxima de 12.5 cm según Bóhlke y Chaplin (1968). La captura se realizó mediante buceo autónomo, con el auxilio de una red de 0.3 mm de abertura, con un marco cuadrado de 40 cm. Los ejemplares capturados se colocaron en nailon de polietileno transparente para ser trasladados hasta el área de cuarentena del Acuario Nacional de Cuba.
Trabajo de laboratorio: Para evaluar la infracomunidad a lo largo de la ontogenia, los peces fueron divididos en clases de talla según dos criterios: (1) Se dividieron los peces en diez clases de talla de 1cm de largo total (LT) según Sturges (1926); y (2) Se dividieron los peces en dos clases de talla, menores y mayores de 6 cm de LT. Los peces fueron sacrificados mediante un corte en la región cervical (Fernández & Corrada, 2009). Se midió la LT con un pie de rey de 0.1 mm de precisión y el peso en una balanza técnica de 0.1 g de precisión. Los procedimientos parasitológicos aplicados fueron los descritos por Moravec et al., (1995). La identificación de los parásitos colectados se basó en: Yamaguti (1963) para monogé-neos; Vidal-Martínez, Aguirre-Macedo, Tomás, David y Edgar (2001) para tremátodos, nemá-todos y céstodos; Noga (1996) para turbelarios, y Kensley y Schotte (1989), y Cressey (1991) para crustáceos.
La magnitud de las parasitosis en los especímenes de C. cyanea fue descrita por tres variables (Bush et al., 1997): (i) prevalencia: porcentaje de individuos hospederos parasitados; (ii) abundancia: media de individuos parásitos por hospedero parasitado o no parasitado; e (iii) intensidad media: media de individuos parásitos por hospedero parasitado. El análisis de la infracomunidad consideró la riqueza como el número de taxa parásitos, la abundancia total como el número de individuos parásitos en cada infracomunidad (Holmes & Price, 1986) y la diversidad mediante el índice de Brillouin (Magurran, 1988) que se recomienda para comunidades completamente censadas.
Se aplicó la prueba no paramétrica de Kruskal-Wallis para detectar diferencias entre clases de talla. Para evaluar la existencia o no de patrones ontogenéticos en las infracomu-nidades a lo largo de las 10 clases de talla, se realizó un análisis de regresión simple. Las variables dependientes fueron la riqueza, abundancia total, diversidad y la composición de la infracomunidad, en tanto que el largo total fue la variable independiente. La composición de las infracomunidades fue evaluada como un vector de abundancia relativa expresada como porcentaje del total de individuos parásitos que cada taxón aporta en cada infracomunidad. Se realizó un análisis de correspondencia con efecto de arco corregido (DCA) que permitió ordenar las infracomunidades en una dimensión. La composición de las infracomunidades se representa a través de los puntajes de los taxa en el primer eje de ordenación, cuyas unidades son desviaciones estándar (x100) de la tasa de reemplazo a lo largo del gradiente. Para realizar el DCA solo se consideraron los taxa con más de cinco ocurrencias en el total de cada intervalo, debido a que este análisis es sensible a las especies raras (Jongman, Braak, & Tongeren, 1995).
Para establecer las posibles diferencias de la infracomunidad, antes y después de la talla de seis centímetros de largo total, se evaluó la riqueza, abundancia, y los índices de equita-tividad de Pielou, de diversidad de Brillouin y de dominancia numérica de Berger-Parker (Magurran, 1988). Las comparaciones se hicieron mediante la prueba U de Mann-Whitney (Jongman et al., 1995). Los ejemplares colectados e identificados, fueron depositados en las Colecciones Científicas del Acuario Nacional de Cuba.
Resultados
Análisis parasitológico: Se examinaron 278 ejemplares de C. cyanea y el largo total fluctuó entre 1.5 y 11.5 cm. Se obtuvo un total de 2 861 parásitos pertenecientes a 20 taxa distribuidos en siete clases. Crustacea y Nematoda fueron los mejores representados con cinco taxa cada uno, Trematoda con cuatro, Cestoda y Monogenea con dos, y Turbellaria y Ciliophora con uno. De los taxa colectados, seis se encontraron en estadío larval y 14 en fase adulto.
El 60% del total de hospederos presentó algún taxón parásito y en cada clase de talla el 53% o más estuvo parasitado por al menos un taxón. El 85% de las especies parásitas mostraron prevalencia por debajo del 20%. Más del 60% de las especies mostraron una intensidad de parasitación por debajo de cinco individuos por hospedero (Cuadro 1). La entidad Tetra-phyllidea mostró la mayor prevalencia en todas las clases de talla con un rango de valores entre 20 y 52%. Le siguen en orden de prevalencia Anilocra chromis, Eutetrarhynchus sp. y Caligus ocyurus. Tetraphyllidea tuvo los mayores valores de intensidad promedio con un máximo de 260 individuos en el primer intervalo de talla (Cuadro 1).
Cuadro 1: Parámetros de infección de la fauna parásita de Chromis cyanea para 10 intervalos de talla definidos por la longitud total
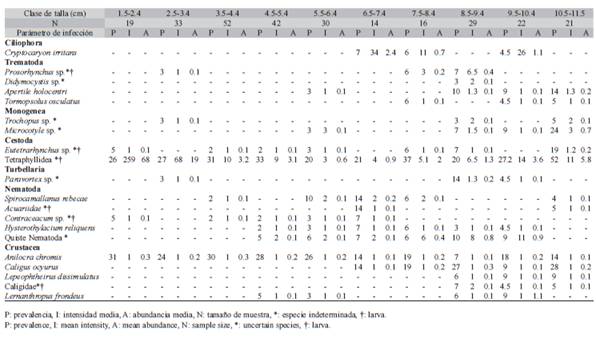
El taxón más prevalente y abundante fue Tetraphyllidea que no presentó diferencias estadísticas significativas entre intervalos de tallas para la abundancia (H(9; 278) = 11.61, p = 0.23) y la intensidad media (H(9 82) = 13.02, p = 0.16). Anilocra chromis no presentó diferencias estadísticas significativas en la abundancia (H(9; 278) = 9.71, p = 0.37) ni en la intensidad promedio (H(9; 65) =4, p = 0.91). El taxón dominante fue Tetraphyllidea en cada intervalo de talla, con un valor máximo de 0.42 ± 0.43.
Análisis ontogenético: El análisis de regresión entre el LT y los descriptores de la infracomunidad, indicó una relación lineal pobre aunque la pendiente de la regresión para cada uno de los descriptores fue similar y estadísticamente significativa (Fig. 1A, Fig. 1B, Fig. 1C). La posición en el gradiente de composición de los taxa más abundantes se indica en la figura 1D. La correspondencia en el primer eje del DCA de los taxa parásitos mostró un valor de 0.67 y la composición de la infracomunidad presentó un ligero incremento con el desarrollo ontogenético de C. cyanea. (Fig. 1D). Los taxa Tetraphyllidea y A. chromis parasitan al hospedero a partir de la talla de 1.5cm de LT y persisten durante toda la ontogenia (Fig. 1D).
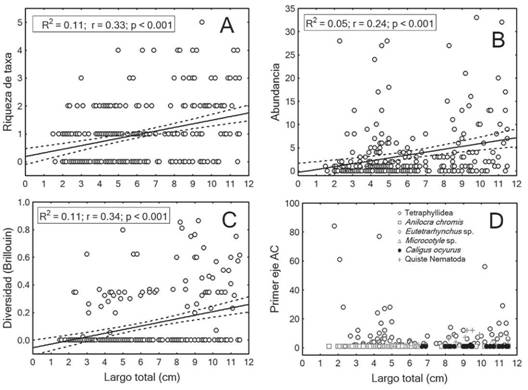
Fig. 1: Asociación de descriptores de la infracomunidad parásita con la longitud de 278 especímenes de Chromis cyanea. A) Riqueza de taxa parásitos; B) Abundancia; C) Índice de diversidad de Brillouin; y D) Composición de la comunidad medida por los puntajes del primer eje de un análisis de correspondencia (AC). Se indican los valores del coeficiente de determinación (R2), coeficiente de correlación (r) y la probabilidad (p) de cometer error de tipo I en un análisis de regresión.
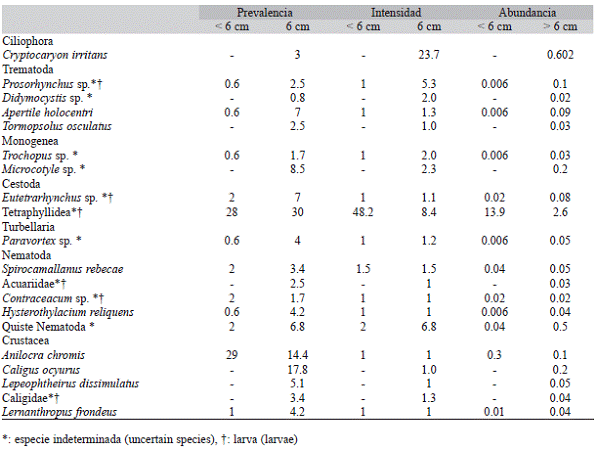
El análisis realizado de la composición de especies antes y después de la talla de maduración sexual de C. cyanea, reveló un incremento notable de los índices cuantitativos del parasitismo, para la mayoría de los taxa hacia la segunda clase de talla. La composición por grupos taxonómicos se comportó de forma heterogénea dentro de ambos intervalos. El número de entidades parásitas incrementó en la segunda clase de talla. Tetraphyllidea disminuyó la intensidad y la abundancia en el segundo intervalo, aunque incrementó ligeramente la prevalencia (Cuadro 2).
Los descriptores de la infracomunidad mostraron un comportamiento diferente antes y después de la talla de 6cm de LT. El promedio de especies por hospedero fue de 0.65 ± 0.78 para la clase < 6 cm y 1.32 ± 1.24 para la clase > 6 cm. El promedio de parásitos por hospedero resultó mayor en la primera clase de talla (14.0 ± 93.2) y menor que en la segunda (5.0 ± 8.5); esto se debe a intensidades de parasitación extremas por Tetraphyllidae que oscilaron entre 3 y 260 parásitos. La diversidad de Brillouin mostró un incremento notable en los especímenes > 6 cm de LT (Cuadro 3).
Cuadro 2: Parámetros de infección de la fauna parásita de Chromis cyanea para dos intervalos de talla definidos por la longitud total: < 6 cm (n = 165) y > 6 cm (n = 113)

Discusión
Este estudio es el primero en caracterizar la fauna parásita de Chromis cyanea a lo largo de la ontogenia de la especie. Otros estudios (Fernández & Corrada, 2009) han analizado la infracomunidad de esta especie hospedera, pero solo en estadios adultos.
La mayor parte de los parásitos de Chro-mis cyanea son especies generalistas que se transmiten por mediación de otras especies de peces simpátricas. El 35% de las especies parásitas son especialistas y solo la entidad quiste de nemátodo se definió con estatus desconocido. Resultados similares en el predominio de larvas de céstodos en C. cyanea fueron obtenidos por Fernández y Corrada (2009). Otros estudios realizados en peces marinos han descrito estos parásitos con altos indices de infección (Sánchez-Ramírez & Vidal-Martínez, 2002; Pérez de Olmo, 2008; Muñoz & Delorme, 2011). Gonzáles y Vidal-Martínez (2008) caracterizaron las comunidades parásitas de Symphurus plagiusa y detectaron una larva Trypanorhyncha con alta intensidad de parasitación y prevalencia. La presencia de larvas con valor de prevalencia elevado puede ser explicada por los hábitos zooplanctófagos de C. cyanea que permiten incorporar por vía trófica especies de crustáceos que constituyen hospederos intermediarios en el ciclo de vida de los céstodos.
La especie generalista Anilocra chromis infestó a Chromis cyanea desde las primeras clases de talla pero siempre con una intensidad de un individuo. Fernández y Corrada (2009) señalan que este parásito siempre se fija en la parte superior del opérculo y que luego de la infestación, se podrían activar mecanismos de exclusión que unidos a la limitada disponibilidad del sitio de fijación impidan la entrada a otros individuos de la misma especie. Tetraphy-llidea también parasitó a C. cyanea desde las primeras clases de talla y persistió durante todo desarrollo ontogenético. Esto puede explicarse por una elevada tasa de residencia que se favorece por la dieta de C. cyanea; constituida fundamentalmente por copépodos, quetognatos, anfípodos, moluscos y poliquetos que pueden ser portadores de nuevos estadíos infectivos (Pastor & Báez-Hidalgo, 2003).
La longitud total del hospedero, como indicador de la ontogenia, tiene un efecto débil sobre la infracomunidad parásita. Pero ha resultado una variable relevante para explicar las variaciones en la infracomunidad parásita de otras especies de peces marinos (Iannacone & Alvariño, 2009; Muñoz & Delorme, 2011; Muñoz & Zamora, 2011). Sin embargo, en estudios realizados en las familias de peces Serranidae y Sciaenidae la talla es irrelevante. Esta ausencia de efecto de la talla podría deberse a dos aspectos: (i) el espectro de tallas analizado en los otros estudios ha sido reducido y/o (ii) los hospederos durante su ontogenia no migran para ocupar otros hábitats (Oliva & Luque, 2002; Iannacone, Morón, & Guizado, 2010).
El incremento en la riqueza y abundancia de parásitos ocurre en los individuos de mayor talla. Esto podría deberse al efecto combinado de: (i) la acumulación de parásitos a lo largo de la ontogenia; (ii) una tasa mayor de ingesta o (iii) la ingesta de presas mayores más parasitadas. La conducta alimentaria de C. cya-nea incluye migraciones diarias que permiten explorar nuevos hábitats con mayor disponibilidad de alimento dentro del arrecife (Claro et al., 2001). Esto facilita la incorporación de parásitos por vía trófica que afectan la composición y abundancia de la infracomunidad (Luque & Poulin, 2007). Adicionalmente, las variaciones en la composición de la infraco-munidad con la talla pueden ser resultado del incremento en la disponibilidad y tamaño de los nichos que ofrecen una mayor heterogeneidad del hábitat y mayor superficie en los órganos (Luque, Porrozzi, & Alves, 2002). Sin embargo, otros estudios han encontrado que el tiempo ontogenético puede generar una estabilidad numérica en la infracomunidad a través de bajas tasas de colonización y altos tiempos de residencia (Díaz & George-Nascimento, 2002).
Las entidades parásitas Tetraphyllidea y Anilocra chromis son consideradas especies núcleo de la infracomunidad, con un efecto notable sobre la estructura comunitaria. Nuestros resultados apoyan la hipótesis de las especies núcleo que parasitan la población de hospederos antes que las especies raras y estocásticas y persisten durante la ontogenia (Vidal-Martínez, Aguirre-Macedo, Vivas-Rodríguez, & Moravec, 1998; Pérez de Olmo, 2008). La persistencia de las especies parásitas en la infracomunidad de C. cyanea a lo largo de las clases de talla reflejan un patrón anidado. Esto puede deberse a una acumulación de parásitos en el tiempo, que se mantiene para las especies transmitidas por vía trófica.
Los cambios obtenidos en los descriptores ecológicos antes y después de la talla de 6 cm en C. cyanea, podrían responder a un efecto acumulativo unido a los cambios en la dieta. Pastor & Báez-Hidalgo (2003) determinaron que a los 6.5 cm de LT ocurre la maduración sexual de C. cyanea en ambos sexos; esta maduración es acompañada de un cambio radical en la dieta. Estos cambios en la dieta suplen las necesidades energéticas de la especie y consisten en la incorporación de un espectro y volumen mayor de presas. Los requerimientos energéticos de los hospederos podrían explicar la persistencia de las características cuantitativas, en tanto que la variación de la composición de las infracomunidades podría resultar de cambios en la oferta de presas.
Se concluye que el desarrollo ontogenético de Chromis cyanea constituye una variable importante en la formación de la infracomu-nidad parásita. Los taxa Tetraphyllidea y Ani-locra chromis constituyen el núcleo de la infracomunidad parásita y la secuencia de aparición y persistencia de estas entidades durante el desarrollo ontogenético, apoyan la hipótesis de las especies núcleo. Los cambios en la infra-comunidad a partir de la talla de 6 cm podrían ser el resultado de un efecto acumulativo unido a cambios en la dieta que aporta nuevos esta-díos infectivos.

