











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkPlant-pollinator interaction had been focused at the species level (Duque-Buitra-go, Quintero, & Otero, 2014); nevertheless, more recently community level pollination studies had been explored on a broader perspective using interaction networks (Fontaine, Dajoz, Meriguet, & Loreau, 2005; Vázquez & Aizen, 2004). Plant-pollinator networks are now known to be asymmetric in their specialization affecting the community organization and ecosystem sustainability (Bascompte, Jor-dano, Melián, & Olesen, 2003; Fontaine et al., 2005; Vázquez & Aizen, 2004). In the tropics, the plants are more diverse than in temperate areas. For example, the cloud forest of La Planada on the Western slope of the Andes in the South of Colombia contain more than 890 vascular plant species (Vallejo, Samper, Memdoza, & Otero, 2004). This high plant diversity may be associated with a concordant requirement of pollinators producing complex plant-pollinator networks.
Male orchid bees (Apidae: Euglossini) are important pollinators of lowland Neotropics Angiosperm (Ackerman, 1985; Roubik & Hanson, 2004). The importance of male Euglossini bees has been documented for many plant families in different ecological and evolutionary contexts (Ramírez, Dressler, & Ospina, 2002); bees visited nine plant families as aromatic resources and 35 families for nectar, pollen and/or nesting materials. They also collect food resources such as nectar with their long tongues (a characteristic of the tribe) playing a determinant ecological and physiological role (Borrell, 2004; Borrell & Krenn, 2006). The proboscis length could have preponderant implications in niche segregation, due to its capacity for accessing deep corollas and the pressure generated across the mouth parts during nectar intake (Borrell, 2006; Parra-H & Nates-Parra, 2012).
Palynological studies of male euglossine floral preferences, could provide a more detailed assessment of plant species spectrum pollinated by these bees, since both males and females collect nectar from the same plant species and especially because there is no systematic methodology for female sampling (Ackerman, 1985). This kind of approaches should suggest general patterns in both, spatial and temporal use of resources.
Here we document bee-plant relationships by analysing pollen attached to male bodies and describe the intra and inter-specific variations in the use of resources by implementing an interaction network analysis. Our temporal and spatial gradient was founded in a primary forest region in the foothills of the Western Pacific cordillera of Colombia; a biodiversity hot-spot region highly rich in orchid bees (Otero, Chacón, Silverstone-Sopkins, & Giray, 2008; Otero & Sandino, 2003; Parra-H, Ospina-Torres, & Ramírez, 2006; Roubik & Hanson, 2004; Sandino, 2004).
Materials and Methods
Study site: This study was conducted in "Río Ñambí" natural reserve, located at the municipality of Barbacoas, "corregimiento" Altaquer, Nariño state - Southwest Colombia (1°18'00" N - 78°05'00" W). The reserve covers an area in the Pacific foothills that varies in altitude from 900 to 1 900 m.a.s.l, characterized predominantly by a sub-Andean cloud forest which is currently subjected to various deforestation processes. Precipitation in the region is the highest in Colombia (7 100 mm per year) with little seasonality, but a wetter season does occur between January and July. The annual average temperature is 24 °C with small variations across the altitudinal gradient (IDEAM, 2009; Gutiérrez, Carrillo, & Rojas, 2004; Solarte, 2005).
The reserve has an extremely high floris-tic diversity and also high level of endemism. Hilty (1980) and Salaman (2001) described it as an evergreen forest lacking predominant deciduous periods. Vegetation develops in slopes up to 35 degrees with a canopy that reaches 25 to 30 m and high level of epiphyt-ism. Franco-Rosselli, Betancur and Fernández-Alonso (1997) registered 205 species, 122 genera and 59 families. Nearly a third part of the plant families are represented by a single species; the most species-rich families in the region are: Rubiaceae, Araceae and Melasto-mataceae and the genera with most species were Anthurium, Psychotria, Philodendron, Piper, Inga and Miconia.
We chose different sampling points in two different areas of the reserve: "Ñambí" which corresponds to an area at approximately 1 440 m elevation and "La Tajada", which is at 980 m. These two areas are separated by no more than 10 km. Samples were collected on April, May, June, August and October 2005, excepting May in Ñambí due to excessive rainy weather conditions.
Pollen sampling: During collecting periods male euglossinae bees were sampled for three days of baiting with Cineole, Methyl Salicylate or Skatole (Otero & Sandino, 2003) the baits were placed on filter paper and bees were captured with an entomological net (Ack-erman, 1983). We obtained pollen samples from male bees by using the glycerinated gelatine method (Amaya, Gutierrez, Rojas Giraldo, & Villegas, 1999), which is widely used in bat and bird pollination studies. By rubbing the bee bodies with fuchsine-stained gelatine we obtained pollen loads that were then mounted on micro-slides. Each slide held the pollen load of a single male body. Pollen grains were observed using a LEICA(r) DMLS microscope with a 1 000x magnification, then identified with help of pollen catalogues (Colinvaux de Oliveira & Moreno, 1999; Palacios Chavez, Ludlow-Wiechers, & Villanueva, 1991; Roubik & Moreno, 1991) and the palynological collection of the Bee research Laboratory LABUN at UNAL Bogota.
Resulting data were analysed with the R, 2.14.1 software - Bipartite package (The R Foundation for Statistical Computing, version 2.14.1), for visualizing interaction networks and determining descriptive indexes such as connectance and nesting (Jordano, Vasquez, & Bascompte, 2009). Shannon diversity indexes for bees and plant host communities were estimated using the PAST(r) 1.74 software (Hammer, Harper, & Ryan, 2001). Species accumulation curves were also made with the Chao 2 and Jacknife 2 estimators using the EstimateSWin800(r) software.
To detect the relative importance of the different resources used by the sub-community of male euglossine bees and to represent the availability of resources throughout the year, we built an Arithmetic Index (AI) which considers three categories, with values from one to three to each category level as follows:
a. Resource use in relation to the number of specimens amount of visiting bees: 1 = scarcely visited (one to five specimens), 2 = moderately visited (6 to 30 specimens) and 3 = highly visited (more than 30 specimens).
b. Resource use in relation to visitor richness: 1 = restricted (a single visiting species), 2 = discrete (two to seven species) and 3 = generalize (more than seven species).
c. Resource frequency in relation to their appearance during the sampling periods (months): 1 = seasonal (one sampling period), 2 = semi-seasonal (two to three sampling periods) and 3 = non-seasonal (four to five sampling periods).
The resulting values of these three categories were summed to calculate the arithmetic index IA, which provided a numeric description of the resource relative importance for the euglossine bee community. Depending on the IA resulting value, plant species were classified as follows: Minor: IA = 3-4, scarcely visited by few species along all year.
Important: IA = 5-7, plant species of variable importance for the euglossine community. Predominant: IA =8-9, highly visited by many species along the year.
Results
Euglossine bee census and palinomorph documentation demonstrated that the male subcommunity is represented at least by 22 species, which interacts with 84 different plant species, from which five remain unidentified. In the upper area of Nambi we registered 15 bee species (134 specimens, H' = 1.40), which collectively were associated with 36 plant species (H' = 2.53). In the lower area of La Tajada we recorded 18 euglossine species (218 collected specimens, H' = 2.47), which had interactions with 72 plant species (H' = 3.54). The two areas had 12 orchid bee species and 24 host species in common.
Euglossine diversity was relatively low in both places. We captured more individuals and species of Euglossa at La Tajada (14 spp., 195 bees) than Nambi (10 spp., 121 bees), whereas Eulaema diversity was slightly higher at Nambi (4 spp., 12 bees) than La Tajada (2 spp., 14 bees). Euglossa maculilabris Moure was highly dominant in Nambi, whereas Eg. orellana Roubik Eg. championi Cheesman Eg. ignita Smith and Eg. mixta Friese were the major species in La Tajada (Table 1).
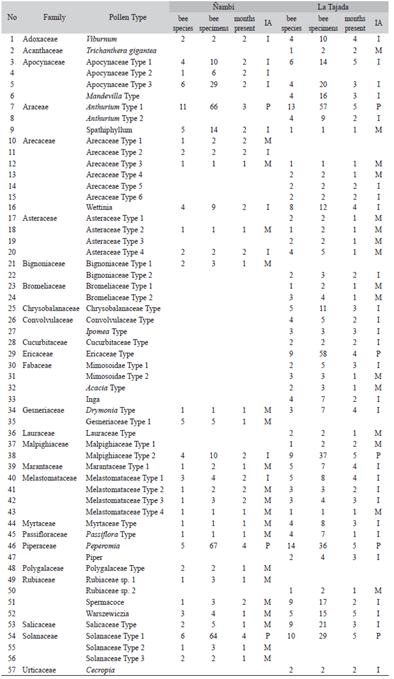
Among floral resources, Arecaceae and Melastomataceae were the major families in Nambi (4), followed by Rubiaceae, Apocyna-ceae, Solanaceae and Gesneriaceae (3). In La Tajada the most important families were Faba-ceae (7), Rubiaceae, Arecaceae (6), Melasto-mataceae (5) Asteraceae and Apocynaceae (5). In Nambi 8 % of the species were predominant, 29 % important and 63 % minor. In La Tajada 7 % were predominant, 39 % were important and 54 % were minor. The predominant species in Nambi were Anthurium Type 1, Peperomia and Solanaceae Type 1, while in La Tajada were Anthurium Type 1, Ericaceae Type, Malpighia-ceae Type 2, Peperomia and Solanaceae Type 1 (Table 2, Fig. 1). The two sites were also different in the composition of floral resources: only 22 of 84 species were shared between La Tajada and Nambi. Nevertheless this small group of shared plants included all but one of the predominant species. Ericaceae Type that only occurred at La Tajada (Table 2).
Table 1: Number of visited plant species and collected specimens per bee species in the two sampled areas
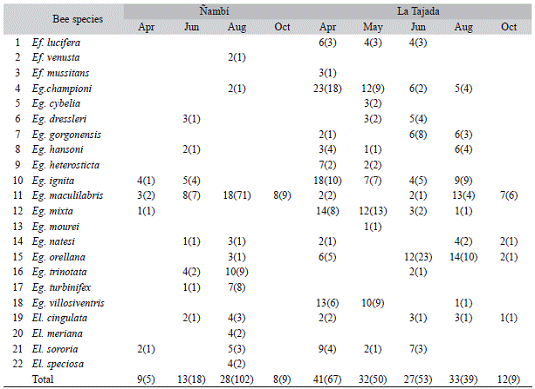
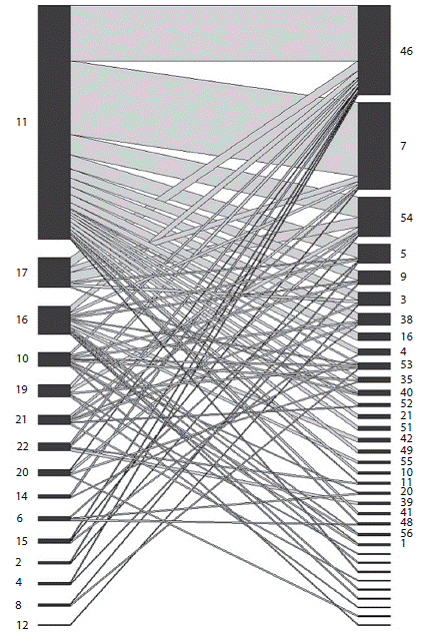
Figure 2 and figure 3 show interaction networks found in Nambi and La Tajada. Con-nectance values were 0.1629 and 0.1666 and nesting values were 14.16 and 14.93 respectively. Interactions (number of visited plant species) per bee species varied from 1 to 22 in Nambi and 1 to 31 in La Tajada, depending on the amount of collected specimens (species with more specimens interacted with more plant species). Interactions per specimen varied from 1 to 5 (mean = 2.5, sd = 1.03) and 0 to 6 (mean = 2.3, sd = 1.27) respectively, without finding significant differences between sampling sites or bee species.
Interactions through time were different between localities. More bees, plants and interactions occurred during August in Nambi while in La Tajada more interactions occurred during April (Table 1). Nevertheless it was a remarkable reduction in bee frequency, plant resources and interactions during the rainiest season (October) in both areas.
Regarding plant seasonality, most of the species were seasonal (only appeared on one sampling month) whereas non-seasonal plants (appeared in four or five sampling months) were relatively scarce (Ñambí: 50 % seasonal, 6 % non-seasonal; La Tajada: 51 % seasonal, 14 % non-seasonal). Non-seasonal plant species included those that were visited by more individuals and species of euglossines (Table 2).
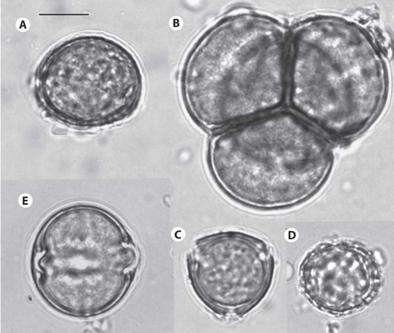
Figure 1: The most frequent pollen species: (A) Anthurium Type 1; (B) Ericaceae Type; (C) Malpighiaceae Type 2; (D) Peperomia; (E) Solanaceae Type 1. The bar at the upper right side measures ten micrometers.
Table 2: Identified plant species appearing in more than one bee specimen in the two sampled areas. M: Minor, I: Important, P: Predominant
Discussion
Bee studied communities frequently depend on a small group of highly visited nectar or aromatic species. This specificity is particularly reflected by the IA index and the high number of interactions found in Anthur-ium Type 1, Peperomia and Solanaceae Type 1 at Nambi, and Anthurium Type 1, Ericaceae Type, Malpighiaceae Type 2, Peperomia and Solanaceae Type 1 at La Tajada. Albeit few interactions were registered for most plant hosts (low-rate visited species) we quantitatively demonstrated that euglossines are polylectic perhaps facilitating their capacity to subsist in relatively adverse environments of high humidity and a drastic altitudinal variation. The high humidity may restrict nest development. For example, euglossinae nest at La Planada, a site nearby Ñambi with similar high humidity, were found with fungal infection (Otero, 1996).

Figure 3: Bee-plant interaction network in La Tajada. Left numbers correspond to euglossine species (Table 1) and right numbers correspond to plant species (Table 2).
By sampling pollen grains from specimens of male bees we discovered several plant taxa that have not been previously reported as floral resources of euglossines. Peperomia (Pipera-ceae) is known as a pollen resource for female Eg. augaspis (Arriaga & Hernández, 1998), yet we found a substantial number of male euglos-sines with Peperomia pollen. Since males do not forage pollen and Peperomia flowers do not produce nectar, these males likely become dusted with pollen when collecting aromatic compounds. Similarly, no Ericaceae have been reported as either aromatic or nectar hosts of orchid bees, yet we discover many euglos-sines with ericaceous pollen. Marantaceae, Rubiaceae, Fabaceae, Costaceae, Orchidaceae, Solanaceae, Apocynaceae, Araceae, Convol-vulaceae, Bignoniaceae and Gesneriaceae are families reported (Otero, Campuzano, Zuluaga, & Caetano, 2014; Ramírez, Dressler, & Ospina 2002) as aromatic or energetic resources. Here we add 16 more families to the list, some of which have many nectarless species such as Arecaceae and Melastomataceae. An alternative explanation for this apparent contradiction in the nature of resources is that pollen may be transferred from females to males when mating occurs.
The pollen registration of Asteraceae, Melastomataceae and Arecaceae, among others, corroborate of Ackerman (1985) findings that euglossines, despite the length of their tongues, are able of exploit shallow floral resources (Borrell, 2006). Male euglossines forage more actively for aromas than energetic resources (Zimmerman & Madriñán, 1988) and these new reports may correspond indeed to fragrances.
While both sites were similar in community richness they differed in composition and species abundance. The Ñambí site had a single dominant species: Eg. maculilabris (with the highest rank of interactions) accompanied by several species of Eulaema. At La Tajada four species were co-dominant: Eg. orellana, Eg. championi, Eg. ignita and Eg. mixta.
The interaction frequency distribution in both areas strongly fit a pattern typically found in plant-animal networks, where many species have few interactions and a few species are supergeneralists (Jordano et al., 2009), as for example Peperomia or Anthurium Type 1 in our case.
This pattern is less accentuated at La Tajada as a result of the availability of floral resources that are in fact more diverse than in Nambi (Solarte, 2005) and consequently, will have more possible interactions between rare species. The higher diversity index values and the total number of interactions support the idea of a more complex interaction network at La Tajada.
Nesting and connectance values were similar in both areas, indicating that the networks are in fact strongly nested. This means that there is a core of bees and plants interacting with each other (e.g. dominant species as Eg. maculilabris, Eg. championi or Eg. ignita interact with predominant species as Peperomia, Anthurium Type 1) as well as a group of rare species (e. g. Ef. lucifera Kimsey, Eg. gorgo-nensis Cheesman or Eg. turbinifex Dressler) interacting with generalist hosts (predominant species). The nested topology constitutes an asymmetric relationship between species that would supposedly provide greater stability for the total system if a species loss occurred (Jordano et al., 2009). The fact that nesting and connectance values were similar for both sites suggests that the euglossine bee community does not strongly differ in its resource usage patterns across the altitudinal gradient, although it does in its species composition.
The absence of the Ericaceae Type in Nambi may be related to variation in bee preference between sites, since this pollen mor-phospecies was well related to absent or very scarce bee species in this site. The euglossinae with ericaceous pollen at La Tajada included: Ef. lucifera, Eg. heterosticta Moure, Eg. villo-siventris Moure, Eg. cybelia Moure, Eg. cham-pioni, Eg. mixta.
Differences in bee community composition also may be due to human intervention events (Otero & Sallenave, 2003; Otero & Sandino, 2003), as well as natural factors such as altitudinal variation (Janzen, 1981; Parra-H & Nates-Parra, 2012), which in our case seems to be a consequence of the flight and thermoregulation bee capabilities (May & Casey, 1983).
Our results showed lower bee capture during the rainy season than the dryer periods. As euglossine bees have little activity on rainy days, the low capture rate on the wettest season may be related with both: a low euglos-sinae population size; or to diminution on the foraging activity associated with the rain. As euglossine bees population are stable (Otero & Sandino, 2003; Roubik & Ackerman, 1987), the second possibility is more likely. The pollination tracking hypothesis predict the abundance of pollinators, Euglossine bees in these case, are correlated with the flowering periods (Zimmerman, Roubik, & Ackerman, 1989), our data showed the most frequent pollen on bees, classified as predominant, were from plants that flower during most of the sampling periods suggesting a steady state phonological strategy (sensu Gentry, 1974). The male euglossine pollination of steady state plants could be favoured by their trap-liner foraging (sensu Ackerman, Mesler, Lu, & Montalvo, 1982).
This study share new light on euglos-sine bees- plant interactions showing new associations previously unknown by combining two sampling techniques: the capture of male euglossinae bees attracted to chemical fragrances and the sampling of pollen from the bee body. This combination allows us to explore a tropical interaction network in a biodiversity hotspot to develop the understanding of the complexity of tropical interactions.

