










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.2 San José Apr./Jun. 2015
Effect of temperature on the expression of IFN-1 (α), STAT-1 and Mx-1 genes in Oncorhynchus mykiss (Salmoniformes: Salmonidae) exposed with the virus of the infectious pancreatic necrosis (IPNV)
Efecto de la temperatura sobre la expresión de genes IFN-1 (α), STAT-1 and Mx-1 en alevines de trucha arcoíris Oncorhynchus mykiss (Salmoniformes: Salmonidae) expuestos al el virus de la necrosis pancreática infecciosa (IPNV)
Efecto de la temperatura sobre la expresión de genes IFN-1 (α), STAT-1 and Mx-1 en alevines de trucha arcoíris Oncorhynchus mykiss (Salmoniformes: Salmonidae) expuestos al el virus de la necrosis pancreática infecciosa (IPNV)
Donald Arguedas Cortés1*,4*, Alex P. Romero Zuñiga2*,3*, Ricardo Enriquez Sais2, José S. Martínez Castañeda1 & César Ortega Santana1
Abstract
The infectious pancreatic necrosis (IPNV) is the causative agent of an acute illness well characterized in salmonids worldwide. Clinical signs and mortality rates are dependent on several factors such as the viral dose, the age of the fish, the water temperature, among others. An experimental study was conducted to measure the effect of temperature on the gene expression profile of IFN-1(α), STAT-1 and Mx-1 in rainbow trout fry, exposed to IPNV. Fry (n=198) were exposed at 8, 12 and 16°C, and samples were taken for 21 days to determine the virus titer and gene expression. In the first 11 days the greatest viral titer was recorded at 8°C compared with the values obtained at 12 and 16°C. At 8°C, there was a significant increase on day 4 of mRNA Mx-1 (t-test, p<0.05), time in which the viral titer began to decrease. Furthermore, as the viral titer increased, STAT-1 and Mx-1 (r=0.91) and (r=0.96) increased, respectively. The animals were able to recover from day 4 from some of the symptoms of IPN. Clinical disease was developed only in fish exposed to 12°C and all died between days 6 and 14, despite the highly significant increase shown in the average expression level of Mx-1, compared with the values recorded at 8°C and 16°C (Tukey, p<0.0001). Additionally, the expression profiles of IFN-1(α) and STAT-1 decreased completely (~0.016) and (~0.020 times) on day 7. The highest expression level of IFN-1(α), occurred at 16°C (Tukey, p<0.0005). Fry exposed at 16°C were normal during the experiment. IFN-1(α) possibly generated a protector effect from day 2 when they showed a significant expression increase compared with the results at 8°C and 12°C (t-student, p<0.0001); however, STAT-1 was not significantly affected by temperature, although the highest average expression value was recorded at 16°C. Our research supports the expression of relevant anti-viral response genes as IFN-1(α), STAT-1 and Mx-1 are physiologically modulated by the water temperature, directly influencing the development of the IPN disease in rainbow trout.
Key words: IPNV, temperature, rainbow trout, fry, ISGs, real-time PCR, mortality.
Resumen
El virus de la necrosis pancreática infecciosa (IPNV) es el agente etiológico de una enfermedad aguda bien caracterizada en salmónidos alrededor del mundo. Los signos clínicos y la tasa de mortalidad dependen de varios factores tales como la dosis viral, la edad del pez y la temperatura del agua, entre otros. Un estudio experimental se llevó a cabo para medir el efecto de la temperatura sobre el perfil de expresión génica de IFN-1(α), STAT-1 y Mx-1 en alevines de trucha arcoíris expuestos con IPNV. Los alevines (n=198) fueron expuestos a 8, 12 y 16°C, y se tomaron muestras durante 21 días para determinar el título viral y la expresión génica. En los primeros 11 días el mayor titulo viral se registró a 8ºC en comparación con 12 y 16. A 8°C, existió un incremento significativo en el día 4 del ARNm de Mx-1 (t-test, p<0.05), momento en que el título viral empezó a disminuir. Además conforme el título viral aumentaba, también STAT-1 y Mx-1 aumentaron (r=0.91) y (r=0.96) respectivamente. Los animales fueron capaces de recuperarse desde el día 4 de algunos de los síntomas de IPN. La enfermedad clínica se desarrolló únicamente en peces expuestos a 12°C y todos murieron entre el día 6 y 14, a pesar del incremento altamente significativo mostrado en el nivel promedio de expresión de Mx-1 a 12°C, comparados con los valores registrados a 8 y 16°C (Tukey, p<0.0001). Además los perfiles de expresión de IFN-1(α) y STAT-1 decrecieron el día 7 completamente (~0.016) y (~0.020) veces, respectivamente. El nivel de expresión promedio más alto de IFN-1(α) se registró a 16°C (Tukey, p<0.0005). Los alevines expuestos a 16°C se mostraron normales durante el experimento. IFN-1(α) posiblemente generó un efecto protector desde el día 2 cuando mostró un aumento significativo en comparación con los resultados a 8 y 12°C (t-student, p<0.0001); sin embargo, STAT-1 no fue afectado de manera significativa por la temperatura, aunque el más alto valor de expresión promedio se registró a 16°C. Nuestra investigación confirma que la expresión de genes relevantes de respuesta antiviral como IFN-1(α), STAT-1 y Mx-1 son fisiológicamente modulados por la temperatura del agua, influyendo directamente en el desarrollo de la enfermedad de IPN en trucha arcoíris.
Palabras clave: IPNV, temperatura, trucha arcoíris, alevines, ISGs, PCR tiempo- real, mortalidad.
The virus of infectious pancreatic necrosis (IPNV) is an aquabirnavirus member of the Birnaviridae family that causes infectious pancreatic necrosis (IPN) (OIE, 2006), an emerging disease that affects predominantly salmonids (Roberts, & Pearson, 2005; Smail et al., 2006). It was firstly recorded in brook trout (Salvelinus fontinalis) in North America (M’gonigle, 1941), however, the agent was not isolated from clinically ill animals until 1957 (Wolf, Snieszko, Dunbar, & Pyle, 1960).
IPNV has a cosmopolitan distribution that causes variable mortality rates worldwide (Ortega et al., 2002; García, Galiana, Falcó, Estepa, & Pérez, 2011), and because of its high antigenicity and genetic variability, it has been reported in other families of fish, mollusks, crawfish of fresh and seawater (Dobos, 1995; Hill, & Way, 1995; Saint-Jean, Borrego, & Pérez-Prieto, 2003; Ortega, & Enriquez, 2007).
The clinical illness episode and the mortality percentage have been reported to depend on various factors like water temperature, virulence level and animal’s age, and the latter can range from 6 to 90% (Santi, Vakharia, & Evensen, 2004). According to Wolf (1988), under experimental conditions at 12°C, clinical disease occurs in salmonids, but possible explanations are unknown. As fish gain weight, they begin to be resistant to this disease and become asymptomatic carriers (Imajoh, Hirayama, & Oshima, 2005), representing a risk to wild fish populations and the environment (Wolf, 1988).
The IPNV genome is composed of two segments of double-stranded RNA (dsRNA) that encodes five viral proteins. The segment A encodes VP2, VP3, VP4 and VP5; while B encodes only VP1. These proteins can activate or inhibit certain defense mechanisms of the immune system including the expression of Interferon-stimulated genes (ISGs) type1, considered the first line of defense against viral infection (Robertsen, Bergan, Rokenes, Larsen, & Albuquerque, 2003; Zhou et al., 2007).
The IFN-1(α) activation triggers the signaling pathway JAK-STAT by inducing genes such as Mx, 2-5 oligo adenylate synthetase (2-5 OAS), viperine antiviral protein (Vig-1), kinase protein RNA (PKR), among others (Sen, 2001; Platanias, 2005; Skjesol, Toril, Hegseth, Børre, & Jørgensen, 2009). JAK kinases pathway JAK-STAT (JAK1 and TYK2) and transcription factors associated with the family of STAT proteins (STAT-1, 2, 3 and 4) (Zhou et al., 2007) have been well studied in mammals (Sen, 2001) but poorly in fish (Jørgensen et al., 2007).
Innate immunity is developed from embryogenesis as a faster response, unlike the adaptive response that is performed four to six weeks after fertilization (Herbomel, Thisse, & Thisse, 2001). It is independent of temperature and the most important defense mechanism in aquatic organisms (Robertsen, 2006). Rubio-Godoy (2010) prefers to refer to it as relatively independent of temperature, although others have determined that the expression of antiviral and inflammatory genes is affected by changes in temperature in zebra fish. Specifically, antiviral expression genes was almost completely inhibited at 15°C, however, inflammatory genes such as IL-1β, TNF-α and INOS showed no obvious difference between 15 and 28°C (Dios, Romero, Chamorro, Figueras, & Novoa, 2010). Temperature not only influences the immune system of teleost but also the physiology of poikilothermic animals (Ruiz, & De Blas, 2003).
It is well known that change in water temperature has an impact on rainbow trout physiology (Guderley, 2004; Kraffe, Marty, & Guderley, 2007; Haverinen, & Vornanen, 2007) including their metabolic rate (Battersby, & Moyes, 1998). Additionally, immune-related functions such as endocytosis (Padron, Bizeau, & Hazel, 2000), immune gene transcription (Raida, & Buchmann, 2007), immunoglobulins concentration (Suzuki, Otaka, Sato, Hou, & Aida, 1997; Nikoskelainen, Bylund, & Lilius, 2004), and C-reactive protein (Kodama et al., 2004) have been reported to be dependent on the ambient temperature.
The present work contributes to our understanding of temperature effects on the expression of relevant antiviral genes of innate response in rainbow trout following bath exposed with IPNV.
Materials and Methods
Fish: A total of 198 rainbow trout fry (Oncorhynchus mykiss) (average weight of 1.0g±0.18 and a length of 3.92cm±0.14) were obtained from a farm with an IPNV- free history and transported to the Virology Laboratory, Research Center in Animal Health (CIESA),Toluca, Mexico. Before the experiment started, the cytopathogenic virus and bacterial agents–free condition was confirmed. Three groups of 66 fish were randomly distributed into three glass tanks of approximately 20L of water (with different water temperature of 8, 12 and 16°C) and were provided with artificial aeration. Mini-thermo-heaters of 25watts (Hidom HT-2025) were used to maintain the three specific water temperatures. Fish were acclimated during a period of 20 days. Control tanks (n=12 fish) were placed together for each experimental temperature to monitor mortality.
Virus: An isolated IPNV (strain Buhl), similar to the one obtained by Ortega, et al. (2002) and supplied by CIESA, was replicated by inoculation in bottles of 500mL in Chinook salmon embryo cells (CHSE-214) with 90% confluence maintained in (MEM), supplemented with 100µg/mL of streptomycin, 60µg/of penicillin and 2% of (FBS). Once reaching an extensive cytopathic effect (EPC), the virus was titered using the microplate method (Reed & Muench, 1938).
Infection: Fish (average weight of 1.0g±0.18 and a length of 3.92cm±0.14) were exposed during 30 minutes with a viral solution of 1x106.16 TCDI50/mL of MEM (endpoint dilution assay quantifies the amount of virus required to kill 50% of infected to produce a cytopathic effect in 50% of inoculated tissue culture cells). Afterwards, they were transferred into three other glass tanks (with water temperatures at 8, 12 and 16°C). Each temperature was monitored daily with a thermometer inserted into each tank. The animals were fed ad libitum (12% of body mass/day) with a commercial feed for trout (El Pedregal TM). The water used in each of the glass tanks was chlorinated after the experiment to destroy viruses (OIE, 2006).
Samples: At days 2, 4, 7, 11, 14 and 21 post infection (dpi), animals from the different tanks were euthanized and spleen and kidney (n=5) were obtained aseptically and mixed for viral quantification using microplate method (Reed, & Muench, 1938). Additionally, kidneys (n=6) with the same infection period were obtained and transferred to 300µL of RNA-later (Invitrogen®) for conservation for 24h. Afterwards they were stored at -80°C, until RNA isolation for gene expression studies could be completed.
Extracting total RNA, Retrotranscription (RT): Total RNA was extracted from kidney using the commercial kit (E.Z.N.ATM Total RNA Kit I, Omega-Biotek) and according to the manufacturer’s instructions. A sample of 9.4 µL of RNA was incubated using a thermocycler (Labnet Multigene Gradient) at 37°C for 30minutes, and was treated with DNase (Promega Cat.#M6101), mixed with 1µL of RQ1 RNase-Free DNase and 1µL RQ1 DNase 10X Reaction Buffer per sample. Immediately, 1 µL of Stop Solution was added, and the samples were incubated at 65°C for 10 minutes. Immediately, a first mix containing 1µL dNTPs 10mM, 1.6µL Oligo dT15mer was added to the samples, and were incubated at 60°C for 10 minutes. Subsequently, a second mix containing 4 µL of M-MLV Reverse Transcriptase 5X Reaction Buffer (Promega M531A), 0.5 µL of RNasin 40U/µL (Promega) and 0.5 µL M-MLV Reverse Transcriptase 200U/µL (Promega) was added, and the samples were incubated in a thermocycler, programmed to complete a cycle of 42°C for 60 minutes, and finally at 70°C for 10 minutes. The total final volume was 20µL for each sample. The protocol used was standardized by the Laboratory of Biotechnology and Aquatic Pathology, Austral University of Chile.
Real-Time PCR (qRT-PCR): The cDNA was amplified with a Step OneTM Real Time PCR system Thermal Cycling Block (Applied Biosystems) using the SYBR®Green method. PCR amplification was performed in individual wells of a 48-well optical plate, mixing 2µL of cDNA, 1µL primer Forward, 1µL primer Reverse, 1µL DEPC water and 5µL of SYBR®Green PCR Master Mix (Cat.#4344463), for a final volume of 10µL per sample. The standard cycling conditions were 50°C for 2 minutes and 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute. All samples were analyzed in triplicate. The efficiency of the primers was tested using serial dilutions of a known initial template (101-1010) to produce a standard curve. Relative quantification of the amplified gene products was calculated by 2-ΔΔCt method (Livak, & Schmittgen, 2001). The housekeeping gene elongation factor 1 alpha (ELF-1(α)) was used as the endogenous reference gene. Data are expressed as fold difference of mRNA expression normalized to the housekeeping gene (ELF-1(α)), relative to the values obtained for uninfected fry. Specific primer sequences are documented in Table 1.
We performed Pearson’s correlation (r) test to determine the strength of association between viral titer (n=5) and time (dpi), and viral titer with gene expressions (fold). Analysis of variance (one-way Anova) was used to evaluate the effect of temperature on the expression of each gene and Tukey’s test for multiple comparisons (n=36). We compared expression between genes at each temperature (between days post infection) and expression for each gene between temperatures using a Student t-tests (n=6). Anderson-Darlington normality test was used to determine if the variables were normally distributed. Statistical analyses were performed using GraphPad Prism® (GraphPad Software, Chicago, Inc, USA). The main differences of the group were considered statistically significant if the p-value was ≤0.05.
Results
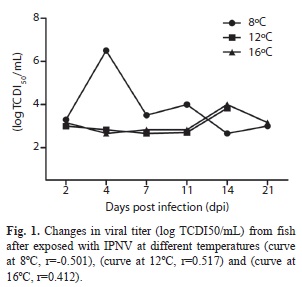
Viral kinetics: At day 2, infected fish showed the highest viral concentration (3.3 log10TCDI50/mL) (Fig. 1, curve at 8°C). Besides, a negative correlation between the viral concentration and time (dpi) (r=-0.501) was found. On the other hand fish exposed at 12ºC, showed a positive correlation (r=0.54) and the virus showed a replication curve of two phases: a decrease (from day 2 to 7) and an increase (from day 7 to 14); while fish exposed at 16°C showed a positive correlation (r=0.412) with a decrease (from day 2 to 4), and an increase (from day 4 to 14).
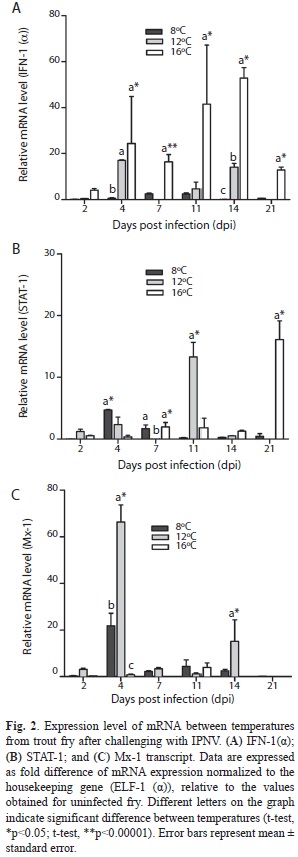
Effect of the temperature on average gene expression in exposed fish along the study: On average, expression profile of IFN-1(α) was significantly highest in fish exposed at 16ºC (Tukey, p<0.0005), while the temperature did not exert significant effect on STAT-1 expression average (Tukey, p>0.05); although, the highest average level expression was recorded at 16°C. At 12ºC, Mx-1 showed a highly significant increase in average level of expression (Tukey, p<0.0001).
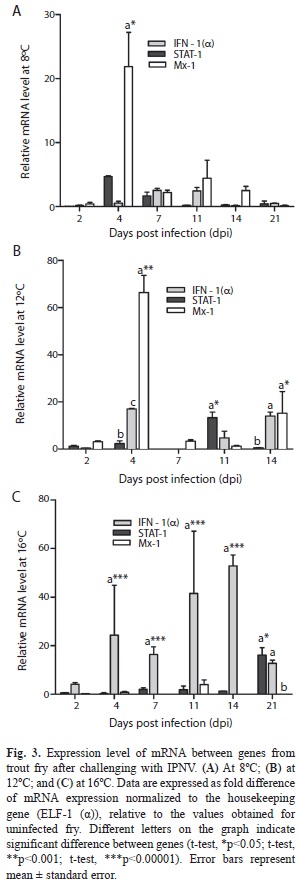
Gene expression at 8°C: The IFN-1(α) gene expression was lower when compared with values obtained at 16ºC (Fig. 2A), but was higher at days 7 and 21, when compared with those obtained at 12 and 16ºC. STAT-1 showed a significant up-regulation on day 4 when compared with the values registered at 12 and 16ºC, and on day 7 compared with those registered at 12°C (Fig. 2B) (t-student, p<0.05). Besides, on day 14, a down-regulation was registered compared with the values obtained at 12°C. It was evident that with the increase of viral titer, the expression of this gene also increased (r=0.91). The Mx-1 expression profile showed a significant up-regulation (~ 21.8 fold) at days 4, 11 and 14 when compared with IFN-1 (α) and STAT-1 values (Fig. 3A) (t-student, p<0.05). A highly positive association between viral titer values and Mx-1 expression profile (r=0.96) was observed. The animals showed some symptoms of IPN disease, however recovered from the symptoms presented from day 4.
Gene expressions at 12°C: The detected IFN-1(α) showed lower expression when compared with the values detected at 16ºC, but this transcript was practically inhibited at day 7 (Fig. 2A) (t-student, p<0.001). The STAT-1 expression value on day 7 was significantly inhibited at 12°C as compared to the values observed at 8 and 16°C (Fig. 2B) (t-student, p<0.05). Additionally, we observed that the expression profile of STAT-1 was lower on day 2 and 4 when compared with Mx-1 value, and completely down-regulated on day 7 (~ 0.020 fold). Furthermore, we observed that STAT-1 expression profile was lower on days 4 and 14 compared to the value registered for IFN-1(α) (Fig. 3B) (t-student, p<0.0001). It was also estimated that with an increase of viral titer, the profile expression of STAT-1 decreased (r=-0.71). At days 2, 4, 7 and 14, Mx-1 transcript showed the highest value when compared with the value registered at 8 and 16°C, displaying significant increasing at days 4 and 14 (Fig. 3C) (t-student, p<0.05). The Mx-1 expression profile was highest at days 2, 4 and 7 when compared with IFN-1(α) and STAT-1, showing a highly substantial up-regulation of transcript on day 4 (Fig. 3B) (t-student, p<0.0001) and day 14 (t-student, p<0.05). We found a negative correlation between viral titer and expression profile of Mx-1 (r=-0.15). Mortality records began on day 6. All animals died by day 14. Fish showed typical lesions of IPN disease: a whirling behavior and internal and external signs, such as darkening, pigmentation, mild to moderate exophthalmia, abdominal distention and accumulation of ascites.
Gene expression 16°C: On day 2, IFN-1(α) was increased when compared with the values obtained at 8ºC and 12ºC, with significant difference (Fig. 2A) (t-student, p<0.05), and showing a highly significant up-regulation on day 7 (t-student, p<0.001). Additionally, IFN-1(α) expression observed an increase when compared with STAT-1 and Mx-1, except for the last day (Fig. 3C) (t-student, p<0.00001). Furthermore, we observed that with an increase of viral titer, the expression profile of IFN-1(α) was increased as well (r=0.42). There was lower expression profile of STAT-1 on day 4 (Fig. 2B) (t-student, p<0.05) compared with the value registered at 8ºC, and on day 11 compared with the value registered at 12ºC (t-student, p<0.05). In addition STAT-1 was significantly up-regulated when compared with IFN-1(α) and Mx-1 on day 21 (Fig. 3C) (t-student, p<0.05). We found that viral titer decreased, and the profile expression of STAT-1 increased (r=-0.76). The mRNA of Mx-1 was lower compared with the value registered at 12ºC (Fig. 2C) except on day 11, the time when it showed a similar value with the data detected at 8ºC. Mx-1 was lower during the experiment when compared with the value of IFN-1(α), the latter being highly significant (Fig. 3C) (t-student, p<0.00001). We observed a negative correlation between expression profile of Mx-1 and viral titer (r=-0.71). No mortality was presented in the tanks and fish showed normal during experiment. Clinical signs of IPN disease were not presented.
Discussion
The temperature influences biological processes, mainly for fish immune responses (Padron et al., 2000; Raida, & Buchmann, 2007). In this study, we analyzed the effect of temperature changes on the expression of antiviral genes against IPNV.
Moss & Gravell (1969) found that IPNV can optimally replicate at 20°C using FHM and CHSE-214 cell lines. The same was reported by Roberts & Dobos (1983) however the viral multiplication results at 20°C were better using BF-2 cell line (Lepomis machrochirus) (Saint-Jean et al., 2003). Our results showed viral replication variations in infected fish in day 2 at different tested temperatures. That was caused by the innate response displayed against the virus and not by temperature alterations. It was observed that the virus can replicate at 4°C (Wolf, 1966), although the lowest temperature tested was 8°C.
Since IFN-1(α) activity was discovered for the first time in FMH cells infected with IPNV (Gravell, & Malsberger, 1965), several authors have demonstrated antiviral activity of cytokine and interferon-induced genes (ISGs) (Roberts, & Pearson, 2005; García et al., 2011) against viral infections in teleost. We agreed with the in vitro and in vivo studies (Kinkelin, & Dorson, 1973; Eaton, 1990; Robertsen, 2006; Verrier, Langevina, Benmansoura, & Boudinota, 2011) which described significant antiviral activity of Mx-1. In our investigation it was evident that at 8°C the viral titer observed was down-regulated by an expression increase of Mx-1 and STAT-1. Additionally, Mx-1 exerted its antiviral activity (Saint-Jean, & Pérez-Prieto, 2007) from the 4th experimental day, possibly interfering with the viral proteins transport (Kochs, Reichelt, Danino, Hinshaw, & Haller, 2005; Wu, Lu, & Chi, 2010). That explains the disappearance of some IPN symptoms at 8°C.
The results obtained at 12°C drew special interests depending on the mortality observed, experimental studies in 60ths (Wolf, 1988) showed high infected salmonids mortality with IPNV at 12°C but still with no explanation. Our possible explanations focus on the implementation of viral strategies for the evasion of the interferon system (Levy, & García, 2001). The first explanation was the significant decrease in the expression of STAT-1, before day 11 compared at 8ºC and the decrease of the IFN-1(α) expression during all infection period compared with 16ºC. Possibly on the first days of the infection, the virus would block the STAT-1 promoter or exerts a block in the interferon stimulated response elements (ISRE) or perhaps an interference in the signaling of IFN-1(α). Additionally, the potential antiviral actions could be the interaction of IPNV with proteins related to the recognition of viral components in the cytosol, as RIG-I and cytoplasmic molecules involved in signaling pathways MAVS (antiviral mitochondrial signaling protein): TRIF, TRAF3, TRAF6 and TBK1 (Xu et al., 2005; Feng et al., 2011). Although, an antiviral action against adapter molecule TICAM-1(a receptor for recognizing RNA duplex in teleost) for TLR-22 can occur (Matsuo et al., 2008). Our second explanation is a possible antagonistic effect by IPNV in the process of translation of the Mx-1 protein that was highest induced in the day 2, 4, 7 and 14 compared to the values obtained at 8°C and 16°C. Similar results were obtained for the expression kinetics of Mx-1 (Collet, Boudinot, Benmansour, & Secombes, 2004), the unique isoform in trout directly induced by IFN-1(α) as obtained by Saint-Jean & Pérez-Prieto (2007) but they injected brown trout (S. trutta) with IPNV virulent serotype (Sp) at 13±1°C. Although in our study Mx-1 after day 7 was not able to exploit the biological function reported in CHSE-214 cells (Larsen, Rokenes, & Robertsen, 2004) and in vivo studies (Saint-Jean, & Pérez-Prieto, 2007).
Skjesol et al. (2009) have referred and pointed to VP4 and VP5 proteins like IPNV virulence factors, however is unknown until now. Although, Dobos (1995) found that IPNV replicates in the cytoplasm, and virus and proteins never enter the cell nucleus, we believe that this issue needs to get the focus. Cellular localization studies of viral proteins like VP4 and VP5 might elucidate these approaches. Jørgensen et al. (2007) proposed a possible promoter activation blockage of Mx-1 in the infection of CHSE-MX10 cells with IPNV at 20°C temperature. In the contrary, at 12°C, we found clinical disease, but the average gene expression of Mx-1 was significantly higher compared to the obtained at 8 and 16°C. Clearly the situation in vivo involves different antiviral and cellular mechanisms.
The protector effect of IFN-1(α) in fish exposed to 16°C during first day of infection, possibly inhibited viral replication (Kinkelin, & Dorson, 1973). Dios et al. (2010), stimulating larvae of zebra fish (Danio rerio) with Poly-IC observed an inhibition in the expression profile of Mx at 15°C. Same results were obtained at 16°C in rainbow trout fry. Although, the expression inhibition of Mx observed by Dios et al. (2010) occurred at a low physiological temperature of a tropical fish (Danio rerio), as a contrary to our work which shows inhibition of Mx at a maximum physiological temperature (16°C). Therefore, the extreme temperature values for each species are critical physiological points to consider for interferon system immune response genes. Our results support that the expression of IFN-1(α), STAT-1 and Mx-1 are physiologically modulated by water temperature, directly influencing the development of IPN disease.
Acknowledgments
This document was funded by the research project No. 99736 (CONACYT) and would not have been possible without the allocation of the scholarship agreement: CONACYT-IICA, Registration No. 283454. We are thankfully with FONDAP 15110027: Interdisciplinary Center for Aquaculture Research (INCAR). Special thanks, also, to the excellent technical assistance of Alicia López Reyes in handling cell line.
References
Battersby, B. J., & Moyes, C. D. (1998). Influence of acclimation temperature on mitochondrial DNA, RNA, and enzymes in skeletal muscle. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 275, 905-912. [ Links ]
Collet, B., Boudinot, P., Benmansour, A., & Secombes, C. J. (2004). An Mx1 promoter–reporter system to study interferon pathways in rainbow trout. Developmental & Comparative Immunology, 28, 793-801. [ Links ]
Dios, S., Romero, A., Chamorro, R., Figueras, A., & Novoa, B. (2010). Effect of the temperature during antiviral immune response ontogeny in teleosts. Fish & Shellfish Immunology, 29, 1019-1027. [ Links ]
Dobos, P. (1995). The molecular biology of infectious pancreatic necrosis virus (IPNV). Annual Review of Fish Diseases, 5, 25-54. [ Links ]
Eaton, W. D. (1990). Anti-viral activity in four species of salmonids following exposure to poly inosinic: cytidylic acid. Diseases of Aquatic Organisms, 9, 193-198. [ Links ]
Feng, H., Liu, H., Kong, R., Wang, L., Wang, Y., Hu, W., & Guo, Q. (2011). Expression profiles of carp IRF-3/-7 correlate with the up-regulation of RIG-I/MAVS/TRAF3/TBK1, four pivotal molecules in RIG-I signaling pathway. Fish & Shellfish Immunology, 30, 1159-1169. [ Links ]
García, I., Galiana, A., Falcó, A., Estepa, A., & Perez, L. (2011). Characterization of an infectious pancreatic necrosis (IPN) virus carrier cell culture with resistance to superinfection with heterologous viruses. Veterinary Microbiology, 149, 48-55. [ Links ]
Gravell, M., & Malsberger, R. G. (1965). A permanent cell line from the fathead minnow (Pimephales promelas). Annals of the New York Academy of Sciences, 126, 555-565. [ Links ]
Guderley, H. (2004). Metabolic responses to low temperature in fish muscle. Biological Reviews, 79, 409-427. [ Links ]
Haverinen, J., & Vornanen, M. (2007). Temperature acclimation modifies sinoatrial pacemaker mechanism of the rainbow trout heart. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 292, 1023-1032. [ Links ]
Herbomel, P., Thisse, B., & Thisse, C. (2001). Zebrafish early macrophages colonize cephalic mesenchyme and developing brain, retina, and epidermis through an M-CSF receptor-dependent invasive process. Developmental Biology, 238, 274-288. [ Links ]
Hill, B. J., & Way, K. (1995). Serological classification of infectious pancreatic necrosis (IPN) virus and other aquatic birnaviruses. Annual Review of Fish Diseases, 5, 55-77. [ Links ]
Imajoh, M., Hirayama, T., & Oshima, S. I. (2005). Frequent occurrence of apoptosis is not associated with pathogenic infectious pancreatic necrosis virus (IPNV) during persistent infection. Fish & Shellfish Immunology, 18, 163-177. [ Links ]
Jorgensen, J. B., Johansen, A., Hegseth, M. N., Zou, J., Robertsen, B., Collet, B., & Secombes, C. J. (2007). A recombinant CHSE-214 cell line expressing an Mx1 promoter–reporter system responds to both interferon type I and type II from salmonids and represents a versatile tool to study the IFN-system in teleost fish. Fish & Shellfish Immunology, 23, 1294-1303. [ Links ]
Kinkelin, P., & Dorson, M. (1973). Interferon production in rainbow trout (Salmo gairdneri) experimentally infected with Egtved virus. Journal of General Virology, 19, 125-127. [ Links ]
Kochs, G., Reichelt, M., Danino, D., Hinshaw, J. E., & Haller, O. (2005). Assay and Functional Analysis of Dynamin-Like Mx Proteins. Methods in Enzymology, 404, 632-643. [ Links ]
Kodama, H., Matsuoka, Y., Tanaka, Y., Liu, Y., Iwasaki, T., & Watarai, S. (2004). Changes of C-reactive protein levels in rainbow trout (Oncorhynchus mykiss) sera after exposure to anti-ectoparasitic chemicals used in aquaculture. Fish & Shellfish Immunology, 16, 589-597. [ Links ]
Kraffe, E., Marty, Y., & Guderley, H. (2007). Changes in mitochondrial oxidative capacities during thermal acclimation of rainbow trout Oncorhynchus mykiss: roles of membrane proteins, phospholipids and their fatty acid compositions. Journal of Experimental Biology, 210, 149-165. [ Links ]
Larsen, R., Rokenes, T. P., & Robertsen, B. (2004). Inhibition of infectious pancreatic necrosis virus replication by Atlantic salmon Mx1 protein. Journal of Virology, 78, 7938-7944. [ Links ]
Levy, D. E., & Garcı́a, A. (2001). The virus battles: IFN induction of the antiviral state and mechanisms of viral evasion. Cytokine & Growth Factor Reviews, 12, 143-156. [ Links ]
Livak, K. J., & Schmittgen, T. D. (2001). Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods, 25, 402-408. [ Links ]
Matsuo, A., Oshiumi, H., Tsujita, T., Mitani, H., Kasai, H., Yoshimizu, M., & Seya, T. (2008). Teleost TLR22 recognizes RNA duplex to induce IFN and protect cells from birnaviruses. The Journal of Immunology, 181, 3474-3485. [ Links ]
M’gonigle, R. H. (1941). Acute catarrhal enteritis of salmonid fingerlings. Transactions of the American Fisheries Society, 70, 297-303. [ Links ]
Moss, L. H., & Gravell, M. (1969). Ultrastructure and sequential development of infectious pancreatic necrosis virus. Journal of Virology, 3, 52-58. [ Links ]
Nikoskelainen, S., Bylund, G., & Lilius, E. M. (2004). Effect of environmental temperature on rainbow trout (Oncorhynchus mykiss) innate immunity. Developmental & Comparative Immunology, 28, 581-592. [ Links ]
OIE. (2006). Manual of diagnostic test for aquatic animals. Paris, France: World Organization for Animal Health. [ Links ]
Ortega, C., & Enríquez, R. (2007). Factores asociados a la infección celular por el virus de la necrosis pancreática infecciosa (IPNV). Archivos de Medicina Veterinaria, 39, 7-18. [ Links ]
Ortega, C., Montes de Oca, R. M., Groman, D., Yason, C., Nicholson, B., & Blake, S. (2002). Case report: viral infectious pancreatic necrosis in farmed rainbow trout from Mexico. Journal of Aquatic Animal Health, 14, 305-310. [ Links ]
Padron, D., Bizeau, M. E., & Hazel, J. R. (2000). Is fluid-phase endocytosis conserved in hepatocytes of species acclimated and adapted to different temperatures? American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 278, 529-536. [ Links ]
Pérez-Prieto, S. I. (2003). Infectious pancreatic necrosis virus: biology, pathogenesis, and diagnostic methods. Advances in Virus Research, 62, 113-135. [ Links ]
Platanias, L. C. (2005). Mechanisms of type-I-and type-II-interferon-mediated signalling. Nature Reviews Immunology, 5, 375-386. [ Links ]
Raida, M. K., & Buchmann, K. (2007). Temperature-dependent expression of immune-relevant genes in rainbow trout following Yersinia ruckeri vaccination. Diseases of Aquatic Organisms, 77, 41-52. [ Links ]
Reed, L. J., & Muench, H. (1938). A simple method of estimating fifty per cent endpoints. American Journal of Epidemiology, 27, 493-497. [ Links ]
Roberts, R. J., & Pearson, M. D. (2005). Infectious pancreatic necrosis in Atlantic salmon, Salmo salar L. Journal of Fish Diseases, 28, 383-390. [ Links ]
Roberts, T. E., & Dobos, P. (1983). Studies on the mechanism of temperature sensitivity of infectious pancreatic necrosis virus replication. Journal of General Virology, 64, 331-339. [ Links ]
Robertsen, B. (2006). The interferon system of teleost fish. Fish & Shellfish Immunology, 20, 172-191. [ Links ]
Robertsen, B., Bergan, V., Rokenes, T., Larsen, R., & Albuquerque, A. (2003). Atlantic salmon interferon genes: cloning, sequence analysis, expression, and biological activity. Journal of Interferon & Cytokine Research, 23, 601-612. [ Links ]
Rubio-Godoy, M. (2010). Teleost fish immunology. Revista Mexicana de Ciencias Pecuarias, 1, 43-57. [ Links ]
Ruiz, F. A., & De Blas, I. (2003). Factors affecting the immune response in fish virus infections. AquaTIC, 19, 1-7. [ Links ]
Saint-Jean, S. R., Borrego, J. J., & Pérez-Prieto, S. I. (2003). Infectious Pancreatic Necrosis Virus: Biology, pathogenesis and diagnostic methods. Advances in Virus Research, 62,113-165. [ Links ]
Saint-Jean, S. R., & Pérez-Prieto, S. I. (2007). Effects of salmonid fish viruses on Mx gene expression and resistance to single or dual viral infections. Fish & Shellfish Immunology, 23, 390-400. [ Links ]
Santi, N., Vakharia, V. N., & Evensen, O. (2004). Identification of putative motifs involved in the virulence of infectious pancreatic necrosis virus. Virology, 322, 31-40. [ Links ]
Sen, G. C. (2001). Viruses and interferons. Annual Reviews in Microbiology, 55, 255-281. [ Links ]
Skjesol, A., Aamo, T., Hegseth, M. N., Robertsen, B., & Jørgensen, J. B. (2009). The interplay between infectious pancreatic necrosis virus (IPNV) and the IFN system: IFN signaling is inhibited by IPNV infection. Virus Research, 143, 53-60. [ Links ]
Smail, D. A., Bain. N., Bruno, D. W., King, J. A., Thompson, F., Pendrey, D. J., & Cunningham, C. O. (2006). Infectious pancreatic necrosis virus in Atlantic salmon (Salmo salar) post-smolts in the Shetland Isles, Scotland: virus identification, histopathology, immunohistochemistry and genetic comparison with Scottish mainland isolates. Journal of Fish Diseases, 29, 31-41. [ Links ]
Suzuki, Y., Otaka, T., Sato, S., Hou, Y. Y., & Aida, K. (1997). Reproduction related immunoglobulin changes in rainbow trout. Fish Physiology and Biochemistry, 17, 415-421. [ Links ]
Verrier, E. R., Langevin, C., Benmansour, A., & Boudinot, P. (2011). Early antiviral response and virus-induced genes in fish. Developmental & Comparative Immunology, 3, 1204-1214. [ Links ]
Wolf, K. (1988). Fish viruses and fish viral diseases. US: Comstock Publishing Associates. [ Links ]
Wolf, K. (1966). Infectious pancreatic necrosis (IPN) of salmonid fishes. Fish, 1, 1-8. [ Links ]
Wolf, K., Snieszko, S. F., Dunbar, C. E., & Pyle, E. (1960). Virus nature of infectious pancreatic necrosis in trout. Experimental Biology and Medicine, 104, 105-108. [ Links ]
Wu, Y. C., Lu, Y. F., & Chi, S. C. (2010). Anti-viral mechanism of barramundi Mx against betanodavirus involves the inhibition of viral RNA synthesis through the interference of RdRp. Fish & Shellfish Immunology, 28, 467-475. [ Links ]
Xu, L. G., Wang, Y. Y., Han, K. J., Li, L. Y., Zhai, Z., & Shu, H. B. (2005). VISA is an adapter protein required for virus-triggered IFN-β signaling. Molecular Cell, 19, 727-740. [ Links ]
Zhou, Z., Hamming, O. J., Ank, N., Paludan, S. R., Nielsen, A. L., & Hartmann, R. (2007). Type III interferon (IFN) induces a type I IFN-like response in a restricted subset of cells through signaling pathways involving both the Jak-STAT pathway and the mitogen-activated protein kinases. Journal of Virology, 81, 7749-7758. [ Links ]
Collet, B., Boudinot, P., Benmansour, A., & Secombes, C. J. (2004). An Mx1 promoter–reporter system to study interferon pathways in rainbow trout. Developmental & Comparative Immunology, 28, 793-801. [ Links ]
Dios, S., Romero, A., Chamorro, R., Figueras, A., & Novoa, B. (2010). Effect of the temperature during antiviral immune response ontogeny in teleosts. Fish & Shellfish Immunology, 29, 1019-1027. [ Links ]
Dobos, P. (1995). The molecular biology of infectious pancreatic necrosis virus (IPNV). Annual Review of Fish Diseases, 5, 25-54. [ Links ]
Eaton, W. D. (1990). Anti-viral activity in four species of salmonids following exposure to poly inosinic: cytidylic acid. Diseases of Aquatic Organisms, 9, 193-198. [ Links ]
Feng, H., Liu, H., Kong, R., Wang, L., Wang, Y., Hu, W., & Guo, Q. (2011). Expression profiles of carp IRF-3/-7 correlate with the up-regulation of RIG-I/MAVS/TRAF3/TBK1, four pivotal molecules in RIG-I signaling pathway. Fish & Shellfish Immunology, 30, 1159-1169. [ Links ]
García, I., Galiana, A., Falcó, A., Estepa, A., & Perez, L. (2011). Characterization of an infectious pancreatic necrosis (IPN) virus carrier cell culture with resistance to superinfection with heterologous viruses. Veterinary Microbiology, 149, 48-55. [ Links ]
Gravell, M., & Malsberger, R. G. (1965). A permanent cell line from the fathead minnow (Pimephales promelas). Annals of the New York Academy of Sciences, 126, 555-565. [ Links ]
Guderley, H. (2004). Metabolic responses to low temperature in fish muscle. Biological Reviews, 79, 409-427. [ Links ]
Haverinen, J., & Vornanen, M. (2007). Temperature acclimation modifies sinoatrial pacemaker mechanism of the rainbow trout heart. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 292, 1023-1032. [ Links ]
Herbomel, P., Thisse, B., & Thisse, C. (2001). Zebrafish early macrophages colonize cephalic mesenchyme and developing brain, retina, and epidermis through an M-CSF receptor-dependent invasive process. Developmental Biology, 238, 274-288. [ Links ]
Hill, B. J., & Way, K. (1995). Serological classification of infectious pancreatic necrosis (IPN) virus and other aquatic birnaviruses. Annual Review of Fish Diseases, 5, 55-77. [ Links ]
Imajoh, M., Hirayama, T., & Oshima, S. I. (2005). Frequent occurrence of apoptosis is not associated with pathogenic infectious pancreatic necrosis virus (IPNV) during persistent infection. Fish & Shellfish Immunology, 18, 163-177. [ Links ]
Jorgensen, J. B., Johansen, A., Hegseth, M. N., Zou, J., Robertsen, B., Collet, B., & Secombes, C. J. (2007). A recombinant CHSE-214 cell line expressing an Mx1 promoter–reporter system responds to both interferon type I and type II from salmonids and represents a versatile tool to study the IFN-system in teleost fish. Fish & Shellfish Immunology, 23, 1294-1303. [ Links ]
Kinkelin, P., & Dorson, M. (1973). Interferon production in rainbow trout (Salmo gairdneri) experimentally infected with Egtved virus. Journal of General Virology, 19, 125-127. [ Links ]
Kochs, G., Reichelt, M., Danino, D., Hinshaw, J. E., & Haller, O. (2005). Assay and Functional Analysis of Dynamin-Like Mx Proteins. Methods in Enzymology, 404, 632-643. [ Links ]
Kodama, H., Matsuoka, Y., Tanaka, Y., Liu, Y., Iwasaki, T., & Watarai, S. (2004). Changes of C-reactive protein levels in rainbow trout (Oncorhynchus mykiss) sera after exposure to anti-ectoparasitic chemicals used in aquaculture. Fish & Shellfish Immunology, 16, 589-597. [ Links ]
Kraffe, E., Marty, Y., & Guderley, H. (2007). Changes in mitochondrial oxidative capacities during thermal acclimation of rainbow trout Oncorhynchus mykiss: roles of membrane proteins, phospholipids and their fatty acid compositions. Journal of Experimental Biology, 210, 149-165. [ Links ]
Larsen, R., Rokenes, T. P., & Robertsen, B. (2004). Inhibition of infectious pancreatic necrosis virus replication by Atlantic salmon Mx1 protein. Journal of Virology, 78, 7938-7944. [ Links ]
Levy, D. E., & Garcı́a, A. (2001). The virus battles: IFN induction of the antiviral state and mechanisms of viral evasion. Cytokine & Growth Factor Reviews, 12, 143-156. [ Links ]
Livak, K. J., & Schmittgen, T. D. (2001). Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods, 25, 402-408. [ Links ]
Matsuo, A., Oshiumi, H., Tsujita, T., Mitani, H., Kasai, H., Yoshimizu, M., & Seya, T. (2008). Teleost TLR22 recognizes RNA duplex to induce IFN and protect cells from birnaviruses. The Journal of Immunology, 181, 3474-3485. [ Links ]
M’gonigle, R. H. (1941). Acute catarrhal enteritis of salmonid fingerlings. Transactions of the American Fisheries Society, 70, 297-303. [ Links ]
Moss, L. H., & Gravell, M. (1969). Ultrastructure and sequential development of infectious pancreatic necrosis virus. Journal of Virology, 3, 52-58. [ Links ]
Nikoskelainen, S., Bylund, G., & Lilius, E. M. (2004). Effect of environmental temperature on rainbow trout (Oncorhynchus mykiss) innate immunity. Developmental & Comparative Immunology, 28, 581-592. [ Links ]
OIE. (2006). Manual of diagnostic test for aquatic animals. Paris, France: World Organization for Animal Health. [ Links ]
Ortega, C., & Enríquez, R. (2007). Factores asociados a la infección celular por el virus de la necrosis pancreática infecciosa (IPNV). Archivos de Medicina Veterinaria, 39, 7-18. [ Links ]
Ortega, C., Montes de Oca, R. M., Groman, D., Yason, C., Nicholson, B., & Blake, S. (2002). Case report: viral infectious pancreatic necrosis in farmed rainbow trout from Mexico. Journal of Aquatic Animal Health, 14, 305-310. [ Links ]
Padron, D., Bizeau, M. E., & Hazel, J. R. (2000). Is fluid-phase endocytosis conserved in hepatocytes of species acclimated and adapted to different temperatures? American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 278, 529-536. [ Links ]
Pérez-Prieto, S. I. (2003). Infectious pancreatic necrosis virus: biology, pathogenesis, and diagnostic methods. Advances in Virus Research, 62, 113-135. [ Links ]
Platanias, L. C. (2005). Mechanisms of type-I-and type-II-interferon-mediated signalling. Nature Reviews Immunology, 5, 375-386. [ Links ]
Raida, M. K., & Buchmann, K. (2007). Temperature-dependent expression of immune-relevant genes in rainbow trout following Yersinia ruckeri vaccination. Diseases of Aquatic Organisms, 77, 41-52. [ Links ]
Reed, L. J., & Muench, H. (1938). A simple method of estimating fifty per cent endpoints. American Journal of Epidemiology, 27, 493-497. [ Links ]
Roberts, R. J., & Pearson, M. D. (2005). Infectious pancreatic necrosis in Atlantic salmon, Salmo salar L. Journal of Fish Diseases, 28, 383-390. [ Links ]
Roberts, T. E., & Dobos, P. (1983). Studies on the mechanism of temperature sensitivity of infectious pancreatic necrosis virus replication. Journal of General Virology, 64, 331-339. [ Links ]
Robertsen, B. (2006). The interferon system of teleost fish. Fish & Shellfish Immunology, 20, 172-191. [ Links ]
Robertsen, B., Bergan, V., Rokenes, T., Larsen, R., & Albuquerque, A. (2003). Atlantic salmon interferon genes: cloning, sequence analysis, expression, and biological activity. Journal of Interferon & Cytokine Research, 23, 601-612. [ Links ]
Rubio-Godoy, M. (2010). Teleost fish immunology. Revista Mexicana de Ciencias Pecuarias, 1, 43-57. [ Links ]
Ruiz, F. A., & De Blas, I. (2003). Factors affecting the immune response in fish virus infections. AquaTIC, 19, 1-7. [ Links ]
Saint-Jean, S. R., Borrego, J. J., & Pérez-Prieto, S. I. (2003). Infectious Pancreatic Necrosis Virus: Biology, pathogenesis and diagnostic methods. Advances in Virus Research, 62,113-165. [ Links ]
Saint-Jean, S. R., & Pérez-Prieto, S. I. (2007). Effects of salmonid fish viruses on Mx gene expression and resistance to single or dual viral infections. Fish & Shellfish Immunology, 23, 390-400. [ Links ]
Santi, N., Vakharia, V. N., & Evensen, O. (2004). Identification of putative motifs involved in the virulence of infectious pancreatic necrosis virus. Virology, 322, 31-40. [ Links ]
Sen, G. C. (2001). Viruses and interferons. Annual Reviews in Microbiology, 55, 255-281. [ Links ]
Skjesol, A., Aamo, T., Hegseth, M. N., Robertsen, B., & Jørgensen, J. B. (2009). The interplay between infectious pancreatic necrosis virus (IPNV) and the IFN system: IFN signaling is inhibited by IPNV infection. Virus Research, 143, 53-60. [ Links ]
Smail, D. A., Bain. N., Bruno, D. W., King, J. A., Thompson, F., Pendrey, D. J., & Cunningham, C. O. (2006). Infectious pancreatic necrosis virus in Atlantic salmon (Salmo salar) post-smolts in the Shetland Isles, Scotland: virus identification, histopathology, immunohistochemistry and genetic comparison with Scottish mainland isolates. Journal of Fish Diseases, 29, 31-41. [ Links ]
Suzuki, Y., Otaka, T., Sato, S., Hou, Y. Y., & Aida, K. (1997). Reproduction related immunoglobulin changes in rainbow trout. Fish Physiology and Biochemistry, 17, 415-421. [ Links ]
Verrier, E. R., Langevin, C., Benmansour, A., & Boudinot, P. (2011). Early antiviral response and virus-induced genes in fish. Developmental & Comparative Immunology, 3, 1204-1214. [ Links ]
Wolf, K. (1988). Fish viruses and fish viral diseases. US: Comstock Publishing Associates. [ Links ]
Wolf, K. (1966). Infectious pancreatic necrosis (IPN) of salmonid fishes. Fish, 1, 1-8. [ Links ]
Wolf, K., Snieszko, S. F., Dunbar, C. E., & Pyle, E. (1960). Virus nature of infectious pancreatic necrosis in trout. Experimental Biology and Medicine, 104, 105-108. [ Links ]
Wu, Y. C., Lu, Y. F., & Chi, S. C. (2010). Anti-viral mechanism of barramundi Mx against betanodavirus involves the inhibition of viral RNA synthesis through the interference of RdRp. Fish & Shellfish Immunology, 28, 467-475. [ Links ]
Xu, L. G., Wang, Y. Y., Han, K. J., Li, L. Y., Zhai, Z., & Shu, H. B. (2005). VISA is an adapter protein required for virus-triggered IFN-β signaling. Molecular Cell, 19, 727-740. [ Links ]
Zhou, Z., Hamming, O. J., Ank, N., Paludan, S. R., Nielsen, A. L., & Hartmann, R. (2007). Type III interferon (IFN) induces a type I IFN-like response in a restricted subset of cells through signaling pathways involving both the Jak-STAT pathway and the mitogen-activated protein kinases. Journal of Virology, 81, 7749-7758. [ Links ]
1. Autonomous University of the State of Mexico, Faculty of Veterinary Medicine and Animal Husbandry, Advanced Animal Health Research and Study Center, Toluca, AP.4-56, México; darguedas@utn.ac.cr, cortegas@uaemex.mx, simonmc@uaemex.mx
2. Laboratory of Biotechnology and Aquatic Pathology, Austral University of Chile, Valdivia, AP.50-9000, Chile; renrique@uach.cl, alexromero@uach.cl
3. Center INCAR: Interdisciplinary Center for Aquaculture Research, Valdivia, AP.50-9000, Chile; alexromero@uach.cl
4. Laboratorio LARED, Universidad Técnica Nacional, Guanacaste, AP Cañas 60 5700, Costa Rica; darguedas@utn.ac.cr
Received 01-IX-2014. Corrected 10-I-2015. Accepted 11-II-2015.


{kind=link}
{kind=link}
{kind=link}