










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.2 San José Apr./Jun. 2015
Reproductive success of Cabralea canjerana (Meliaceae) in Atlantic forest fragments, Brazil
Éxito reproductivo de Cabralea canjerana (Meliaceae) en fragmentos de bosque del Atlántico, Brasil
Éxito reproductivo de Cabralea canjerana (Meliaceae) en fragmentos de bosque del Atlántico, Brasil
Edivani Villaron Franceschinelli1*, Roselaini Mendes do Carmo2*, Carlos de Melo e Silva Neto1, Bruno Bastos Gonçalves1 & Leonardo Lima Bergamini3*
Abstract
In Brazil, the Atlantic forest remnants have high biological diversity and a high level of endemism, but very little is known about the reproductive success of native species. Cabralea canjerana is a common tree in the Montane Atlantic forest, and its reproduction is highly dependent on pollinators. In order to contribute with the particular knowledge on this species, we collected data in three fragmented and three continuous forest sites, where the effects of fragmentation on both mutualistic (pollination) and antagonistic (seed predation) interactions were analysed. We determined fruit production and weight of 25 trees per site. The number of seeds and the percentage of predated and aborted seeds were also accessed for seven fruits of 10 trees per site. Pollinator visitation frequencies to flowers were recorded in two forest fragments and in two sites of the continuous forest. Our data showed that plants of C. canjerana produced more fruits (z-value=-8.24; p<0.0001) and seeds per fruit (z-value=-6.58; p=0.002) in the continuous than in the fragmented sites. This was likely due to differences in pollination, because the number of pollinator visits was higher in the continuous forest than in the fragments. Seed abortion (z-value=4.08, p<0.001) and predation (z-value=3.72, p=0.0002), on the other hand, were higher in the fragmented than in the continuous sites. Then, mutualistic and antagonistic interactions were affected by fragmentation, decreasing the reproductive success of the study tree. This study was the first to show a decrease in the reproductive output in forest fragments in an Atlantic forest tree species. This decrease may threaten the population structure and viability of C. canjerana in forest fragments. Rev. Biol. Trop. 63 (2): 515-524. Epub 2015 June 01.
Key words: Cabralea canjerana, fruit production, habitat fragmentation, plant-pollinator interaction, moth pollination, reproductive success, pre-dispersal seed predation, seed set.
Resumen
En Brasil, los remanentes de bosque del Atlántico tienen una alta diversidad biológica y un alto nivel de endemismo, pero se conoce muy poco sobre el éxito reproductivo de las especies nativas. Canjerana cabralea es un árbol común en el bosque Atlántico Montano y su reproducción es altamente dependiente de los polinizadores. Con el fin de contribuir con el conocimiento particular de esta especie, se recogieron los datos en tres sitios fragmentados y tres de bosques continuos, donde se analizaron los efectos de la fragmentación tanto en relaciones mutualistas (polinización) como interacciones antagónicas (depredación de semillas). Se determinó la producción de frutos y el peso de 25 árboles. También se calculó el número de semillas y el porcentaje de semillas depredadas y abortadas en 7 frutos de 10 árboles por sitio. Frecuencia de visitas de polinizadores a las flores se registraron en dos fragmentos de bosque y en dos sitios de bosque continuo. Nuestros datos muestran que las plantas de C. canjerana produjeron más frutos (Z=-8.24; p<0.0001) y semillas por fruto (Z=-6.58; p=0.002) en el continuo que en los sitios fragmentados. Esto fue probablemente debido a las diferencias en la polinización, porque el número de visitas de polinizadores fue mayor en el bosque continuo que en los fragmentos. Por otro lado, el aborto de semillas (Z=4.08, p<0.001) y la depredación (Z=3.72, p=0.0002) fueron mayores en los sitios fragmentados que en los continuos. Entonces, las interacciones mutualistas y antagónicas se vieron afectadas por la fragmentación, disminuyendo el éxito reproductivo del árbol estudiado. Este estudio fue el primero en mostrar una disminución en el rendimiento reproductivo en los fragmentos de bosque en una especie de árboles forestales atlánticos. Esta disminución puede poner en peligro la estructura de la población y la viabilidad de C. canjerana en fragmentos de bosque.
Palabras clave: Cabralea canjerana, fragmentación del hábitat, interacción planta-polinizador, polinización por polilla, depredación de semillas, producción de semillas, producción de frutos.
Habitat fragmentation may disrupt plant-pollinator interactions by changing the abundance and/or the identity of floral visitors (Lovejoy, 1980; Roubik, 1993; Aizen, & Feinsinger, 1994a; Aizen, & Feinsinger, 1994b; Matheson, Buchamann, O’Toole, Westrich, & Williams, 1996; Murcia, 1996; Prach, Pysek, & Smilauer, 1997). Because floral visitors can differ in their ability to transfer pollen, changes in the pollinator assemblages following fragmentation may affect plant reproductive success (Aizen, & Feinsinger, 1994b). Studies conducted during the past few decades show that fragmentation may affect the reproduction of various plant species in different ways. Most of these studies show decreases in the richness and abundance of pollinators and in the number of pollinator visits in response to fragmentation, leading to a decrease in the reproductive output of plants (Aizen, & Feinsinger, 1994a; Kolb, 2008). In contrast, other studies found no changes in the reproductive output of plants in fragmented habitats (e.g., Aldrich & Hamrick, 1998; Dick, 2001; Lopes, & Buzato, 2007; Aguirre, & Dirzo, 2008). Fragment size may affect plant reproductive output also without any change in pollinator frequency or richness (Ghazoul, 2005; Wagenius, & Lyon, 2010). Plant fertility may be affected by habitat fragmentation indirectly through abiotic changes (Kwak, Velterop, & Andel, 1998), for example, in the light intensity and the availability of water and nutrients in the soil (De Jong, & Klinkhamer, 1989; Saunders, Hobbs, & Margules, 1991).
Abiotic changes caused by habitat fragmentation may also affect the predation rate on plant reproductive structures owing to changes in the local animal community (Cunningham, 2000). Seed predation is an important ecological interaction that may directly impact plant fertility. In some cases, seed predators may cause abortion of the entire fruit (Greig, 1993). Seed predation by insects may change the recruitment rate and the abundance of the host plant (e.g., Jules, 1998; Louda, 1982). Several studies have investigated seed predation in fragmented landscapes and have shown that it may increase with fragmentation (Orrock, Danielson, & Burns, 2003; Fleury, & Galetti, 2006; Galetti, Donatti, & Pires, 2006; Orrock, Levey, & Danielson, 2006). Similarly to plant-pollinator interactions, other studies showed the opposite pattern, where antagonistic interactions may also be disturbed by fragmentation; possibly because the abundance and diversity of antagonistic animals in small populations of plants offer fewer resources to herbivores and seed predators (Colling, & Matthies, 2004; Kéry, Matthies, & Fischer, 2001; Ehlers, & Olesen, 2003).
Effects of habitat fragmentation on plant-pollinator interaction in the Neotropical region have been studied in the Argentinean Chaco (Aizen, & Feinsinger, 1994a; 1994b; Aguilar, & Galetto, 2004), in Mexico (Quesada, Stoner, Rosas-Guerrero, Palacios-Guevara, & Lobo, 2003; Quesada et al., 2004) and in Costa Rica (Cascante, Quesada, Lobo, & Fuchs, 2002). Few studies have been conducted in Brazil (Dick, 2001; Lopes, & Buzato, 2007; Dunley, Freitas, & Galetto, 2009; Elias et al., 2012; Melo, Oliveira, & Franceschinelli, 2014), and only two were performed in the Atlantic forest (Lopes, & Buzato, 2007; Dunley et al., 2009), but none has examine the effect of fragmentation concomitantly on mutualistic (pollination) and antagonistic (seed predation) interactions in this biome. Atlantic forest originally occupied an area of 360 000km2 extending from Northeastern to Southern Brazil. Now, only 7% of this area is covered with natural forest. The remnants of the original forest consist of many small fragments, several medium-sized tracts and a few continuous areas of forest (Tabarelli, Montovani, & Peres, 1999). These remnants have high biological diversity and a high level of endemism (Pinto, Hirota, & Fonseca, 2002), but little is known about the reproductive status of native species.
The present study aims to evaluate pollination, seed predation and reproductive success of plants of Cabralea canjerana subsp. canjerana (Vell.) Mart. in continuous and in isolated forest fragments of the Atlantic forest. This subspecies is a dioecious tree whose reproduction is highly dependent on pollinators. Forest fragmentation affects negatively the population density of C. canjerana (Santos and Ono, unpublished data). However, the effects of forest fragmentation on its reproductive success have not yet been studied. Here, effects of fragmentation on mutualistic (pollination) and antagonistic (seed predation) interactions were analysed to examine how these may influence plant reproductive success. Specifically, we examined pollinator visitation rates and determined the reproductive success in terms of fruit and seed productions as well as in terms of the intensity of pre-dispersal seed predation. We predicted that pollinator visitation rates would be lower in the fragments than in the continuous forest, resulting in lower fruit and seed production. Although seed predation rates may increase or decrease in fragmented habitats, we expected lower plant fertility in fragments than in continuous areas of the Atlantic forest.
Materials and methods
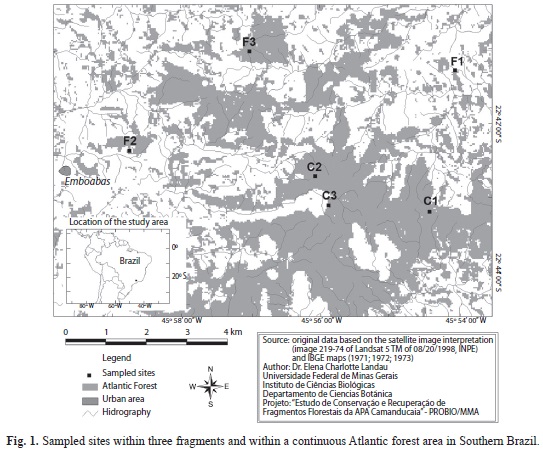
Study site: The study area is located in the Serra da Mantiqueira, Southern Minas Gerais State, in the municipalities of Camanducaia and Gonçalves, Brazil (22° 44’ S, 45° 56’ W, Fig. 1). The predominant vegetation type is wet montane forest. The matrix surrounding the studied fragments is composed of small farms with pasture, potato and carrot fields. Few C. canjerana trees occur in the surrounding matrix.
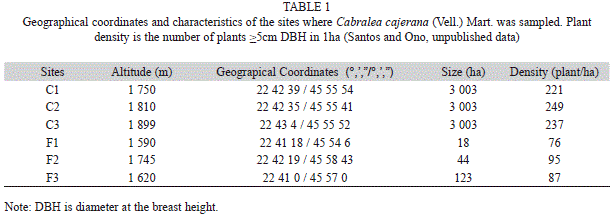
Three fragments (F1, F2 and F3) and three sites in the continuous forest (C1, C2 and C3) were sampled between October of 2001 to October of 2002 (Fig. 1, Table 1). The distances between sampling sites varied from 1.0 to 8.5km. These sites were selected according to the characteristics of the access, size, and mainly to the occurrence of C. canjerana trees. In general, continuous sites are better preserved than the fragments. C1 has experienced selective logging in the past; however, the present owner protects the area from any further disturbance. C2 appears well preserved but shows evidence of earlier selective logging. C3 appears better preserved than C2 and shows no evidence of recent logging. Two isolated fragments (F1 and F2) exhibited strong evidence of selective logging and cattle grazing. F1 was the smallest study fragment (18ha) and F3 was the largest (123ha) (Fig. 1), but F3 has been disturbed by selective logging and grazing. Such as expected, continuous forest sites were better preserved than fragmented ones.
Study species: Cabralea canjerana subsp. canjerana is a very common tree of the montane forests of Southern Minas Gerais State (França, & Stehmann, 2004), where plants can reach 25m tall. It is found in the forests of Central and South America. It has compound pinnate leaves and axillary to cauliflorous inflorescences. This species is dioecious, with white flowers that open at night and are pollinated by moths (Franceschinelli et al., unpublished data). The fruits of this subspecies are red loculicidal capsules with a fleshy pericarp. They contain 1-5 diaspores, which consist of one or two seeds united by the aril. The seeds are ellipsoid and are partially (sometimes completely) covered by the aril. Closed ripe fruits, open fruits and flower buds may occur simultaneously on the same plant due to the long period of fruit development (Franceschinelli et al., unpublished data). The voucher of the species studied is: Herbarium IAC 38690, R.B. Torres, & C.S. Figueiredo 926.
This species was selected because it is abundant in the study area; it occurs primarily in the forest understory and has cauliflory and large red fruits, making it easy to observe its flowers and fruits. The seed aril of C. canjerana is a very important food source for many species of birds and mammals (Pizo, 1997; Carmo, 2005). This species is also valuable for the regeneration of degraded areas (Nóbrega et al., 2008). Besides, its wood was used in carpentry, primarily for roofing and fencing in Southern Brazil (Carvalho, 1994).
Measurements: In each site, all fruits of 25 plants of 7-11m tall were counted when developed but not yet opened. These plants were randomly selected, but trees located in forest edges were avoided. Seven fruits randomly selected from each of these 25 plants were weighted. Seven fruits of 10 plants (selected at random out of the previous 25 trees) were sampled to count the number of seeds per fruit. In those fruits, the number of aborted seeds was counted. Viable seeds may be attacked by dipteran larvae inside the fruit (Carmo, 2005). Thus, seeds attacked by larvae were also counted.
Pollinator visit frequencies (number of visits in 30min) to C. canjerana subsp. canjerana flowers were recorded in two forest fragments (F1 and F2) and in two areas of the continuous forest (C1 and C2). Four randomly selected plants in each area were observed from 20:00 to 22:30h and from 04:00 to 06:30h. Trees located in forest edges were avoided. The observation period lasted 30min per plant and each plant was observed twice or three times per night. Only one inflorescence was observed in each period. A visit was counted as soon as the pollinator visited the flowers to collect nectar. Pollinators visit only few flowers in each visit to an inflorescence of C. canjerana (Carmo, 2005). Each time a pollinator visits an inflorescence, we counted only one visit to this inflorescence, independently of the number of flowers visited in it. The plants observed had similar sizes (8 to 10m tall) and numbers of inflorescences (4 to 6 inflorescences). Each area was sampled for five hours during the flowering peak of C. canjerana (October and November). Some floral visitors were collected and taken to the laboratory to verify the presence of pollen grains on their body.
To test if fragmentation affects C. canjerana reproductive success, we used generalized linear mixed models (GLMMs, Bolker et al. 2009) with reproductive success measures and pollinator visit rates of C. canjerana as response variables and fragmentation status (continuous vs. fragmented landscape) as the predictor variable. In the models for fruit set, each individual plant was a data point, and site identity was used as a random effect to control for the non-independence of the observations made in trees at the same site. In the models for fruit weight, aborted, predated and total seed number, each fruit was a data point and plant identity nested by site identity were used as random effects. In the pollinator visit rate model, each 30min observation period was used as a replicate, with site identity as a random effect. We used a Poisson error structure with log link for the fruit set, aborted, predated and total seed number and number of pollinator visits models and a Gaussian error structure with identity link for the fruit weight model. All analysis were performed on the R environment (R Development Core Team, 2008), using the package lme4 (Bates, Maechler, Bolker, & Walker, 2014).
Results
Cabralea canjerana produced on average 71.7 (±71.0) fruits in continuous forest and 18.5 (±18.3) fruits in fragments, which means 74% less fruits in the fragmented than continuous forest (z-value=-8.24; p<0.0001). The seed number per fruit followed the same pattern, with plants producing on average 6.2 (±1.5) seeds per fruit in fragments and 8.2 (±1.3) seeds per fruit in continuous forest; which means 23% less seeds set in fragments than in continuous forest (z-value= -6.58; p=0.002). However, there was no difference in mean fruit weight between forest types (t-value=-1.641, p=0.104). In continuous forest, fruits weight was on average 5.23g (±1.7) and in fragments 4.6 (±1.7).
The average proportion of aborted seed per fruit was 0.62 (±0.25) in fragments and 0.23 (±0.15) in continuous forest. Therefore, fragments presented 2.1 times more aborted seeds (z-value=4.08, p<0.001) than continuous forest. The average proportion of predated seed per fruit was 0.33 (±0.18) in fragments and 0.07 (±0.08) in continuous forest, resulting in 4.6 times more predated seeds in fragments compared to continuous sites (z-value=3.72, p=0.0002).
Pollen grains were observed on the bodies of the collected moths. However, it was not possible to count them, because of the presence of body scales that easily mixed with the pollen grains, when we tried to remove them. The average number of visits by pollinators was lower in fragments (10.2±4.6) than in continuous forest (5.6±3.0), showing a 45% decrease in pollinator visit rate per inflorescence per tree in the fragments when compared to the continuous forest (z-value=-3.94, p<0.001).
Discussion
Lower fruit and seed set in plants of small habitat fragments have been observed for many species of tropical and temperate vegetation (e.g., Jennersten, 1988; Jennersten, & Nilsson, 1993; Aizen, & Feinsinger, 1994a; Byers, 1995; Oostermeijer, Altenburg, & den Nijs, 1995; Ågren, 1996; Cunningham, 2000; Quesada et al., 2003). Other species either do not show any different response in reproductive output between fragmented and continuous forest or present even higher reproductive output in smaller fragments and open areas than in continuous forest (e.g., Costin, Morgan, & Young, 2001; Dick, 2001; Lopes, & Buzato, 2007; Dunley et al., 2009). Some authors argue that the influence of fragment size and quality depends on the habitat requirements of the plants and the pollinators (Aizen, & Feinsinger, 1994b; Kwak, et al. 1998; Donaldson, Nänni, Zachariades, & Kemper, 2002; Quesada, 2003) and those different groups of species respond differently to habitat fragmentation (Kwak et al., 1998). Cabralea canjerana is an example of a species that shows a reduction in its reproductive success in fragmented habitats. Plants of C. canjerana produce lower fruit, seed per fruit and higher seed predation in fragments than in continuous forest.
Several plant characteristics may contribute to the decrease in reproductive success in fragmented habitats, such as pollinator dependence and self-incompatibility (Rathcke, & Jules, 1993; Aguilar, Ashworth, Galetto, & Aizen, 2006). Sexual system may be another important trait that influences the reproductive output in fragmented habitats. It is probable that the occurrence of male and female flowers in separate plants makes C. canjerana totally dependent on pollinators and vulnerable to their population decreases. The low reproductive success of plants in isolated areas may result from the lower visitation rate of their pollinators to their flowers.
The low visitation rate of pollinators to C. canjerana flowers in fragments may have caused not only the deposition of low numbers of pollen grains on stigmas, but probably lower-quality pollen loads. Moths do not carry large numbers of pollen grains. Although they are efficient at transferring pollen to the stigma, they generally must visit each flower more than once to deposit sufficient pollen grains to fecundate every ovule (Young, 2002). Because pollinators visit only few flowers in each visit to an inflorescence of C. canjerana (Carmo, 2005), probably high visit frequency is important to increase the pollination efficiency and the reproductive success of this species. Variation in visitation rate among fragments of different sizes was also found for other species (Powell, & Powell, 1987; Jennersten, 1988; Quesada et al., 2004).
Fragmentation was negatively related to the density and size of C. canjerana in the areas studied (Santos, & Ono, unpublished data). More intensive selective logging of C. canjerana in the fragments than in continuous forest may have contributed to the decrease in density and plant size in fragments. This may have affected pollination process and reproductive success of C. canjerana in fragmented sites. These populations have also showed higher rates of inbreeding than populations of the continuous forest (Carmo, 2005), which may influence plant fertility as well. Although variations in pollinator visits in different areas are generally related to population size and flower abundance (Murcia, 1990), in C. canjerana, this variation may also be related to differences in the amount and quality of the food resource for the moth larvae. According to Faegri & Pijl (1979), the presence of moths in an area depends more on the occurrence of the plants used by the larvae than on the occurrence of the plants used by the adults. The food resources for the moth larvae may be lower in the fragmented than in the continuous forest.
The reproductive success of C. canjerana was also influenced by seed consumers. In C. canjerana, the number of pre-dispersed seeds damaged by dipteran larvae is higher in fragmented than in continuous forest. Other studies have found similar results (Didham, Ghazoul, Stork, & Davies, 1996; Kruess, & Tscharntke, 1994; Santos, & Telleria, 1994; Simberloff, 1993). High seed predation in fragmented areas has been related to the greater fruit exposure to seed predators and to abiotic variations in factors such as temperature, sunlight and humidity that may influence the populations of seed predators. According to Groom (2001), the high number of predated seeds in small fragments may be related to the impact of fragmentation on the predators of seed consumers. The large amounts of plant diversity generally presented in continuous forest reduce the ability of herbivores to find their host plants (Hämback, & Beckerman, 2003). Cabralea canjerana, which is an understory species, would be better protected from seed preys in the continuous forest, where larger numbers of other plants occur. However, some studies have found that seed predator populations may be disturbed by fragmentation, because plant populations are small and less attractive to seed predators in fragments (Zabel, & Tscharntke, 1998; Farwig, Bleher, von der Gönna, & Böhning-Gaese, 2008; Herrerías-Diego, et al. 2008).
This study is the first to show a decrease in the reproductive output of an Atlantic forest plant species in fragments. Research on the population viability would be important for assessing the future of this species in these forest fragments, especially if population growth rate of C. canjerana is sensitive to changes in fruit and seed production. Moreover, the low reproductive success found in the study fragments may be occurring in other fragments and plant species of the Atlantic forest. This problem may drastically affect the viability of populations in forest fragments, decreasing species diversity in the future and compromising the conservation of this fragmented biome.
Acknowledgments
This work was supported by a grant from PROBIO/MMA (Projeto de Conservação e Utilização Sustentável da Diversidade Biológica Brasileira/Ministério do Meio Ambiente) to the first author. We are indebted to Roderic B. Martines and Antonio Pereira da Silva for their assistance in in the field. We are grateful to Altair Rezende de Souza, Sebastião Loreano, Brás, Antonio Bueno de Souza, Antonio Pereira da Silva and Joaquim Costa for allowing us to work on their properties.
References
Ågren, J. (1996). Population size, pollinator limitation, and seed set in the self-incompatible herb Lythrum salicaria. Ecology, 77, 1779-1790. [ Links ]
Aguilar, R., & Galetto, L. (2004). Effects of forest fragmentation on male and female reproductive success in Cestrum parqui (Solanaceae). Oecologia, 138, 513-520. [ Links ]
Aguilar, R., Ashworth, L., Galetto, L., & Aizen, M. A. (2006). Plant reproductive susceptibility to habitat fragmentation: review and synthesis through a meta-analysis. Ecology Letters, 9, 968-980. [ Links ]
Aguirre, A., & Dirzo, R. (2008). Effects of fragmentation on pollinator abundance and fruit set of an abundant understory palm in a Mexican tropical forest. Biological Conservation, 141, 75-384. [ Links ]
Aizen, M. A., & Feinsinger, P. (1994b). Habitat fragmentation, natives insect pollinators and and feral honey bee in Argentine ‘Chaco Serrano’. Ecological Applications, 4, 378-392. [ Links ]
Aizen, M. A., & Feinsinger, P. (1994a). Forest fragmentation, pollination and plant reproduction in a Chaco dry forest, Argentina. Ecology, 75, 330-351. [ Links ]
Aldrich, P. R., & Hamrick, J. L. (1998). Reproductive dominance of pasture trees in a fragmented tropical forest mosaic. Science, 281, 103-105. [ Links ]
Bates, D., Maechler, M., Bolker, B., & Walker S. (2014). Lme4: Linear mixed-effects models using Eigen and S4. R package version 1.1-7, http://CRAN.R-project.org/package=lme4. [ Links ]
Bolker, B. M., Brooks, M. E. Clark, C. J., Geange, S. W., Poulsen, J. R., Stevens, M. H. H., & White, J. S. (2009). Generalized linear mixed models: a practical guide for ecology and evolution. Trends in Ecology & Evolution, 24, 127-135. [ Links ]
Byers, D. L. (1995). Pollen quantity and quality as explanations for low seed set in small populations exemplified by Eupatorium (Asteraceae). American Journal of Botany, 82, 1000-1006. [ Links ]
Carmo, R. M. (2005). Biologia reprodutiva de Cabralea canjerana subsp. canjerana em Fragmentos de Mata Atlântica do Sul do estado Minas Gerais. (Ph.D. thesis). Universidade Federal de Minas Gerais, Brasil. [ Links ]
Carvalho, P. E. R. (1994). Espécies florestais brasileiras. Recomendações silviculturais, potencialidades e uso da madeira. Brasília DF: Embrapa/CNPF-SPI. [ Links ]
Cascante, A., Quesada, M., Lobo, J., & Fuchs, E. A. (2002). Effects of Dry Tropical Forest Fragmentation on the reproductive success and genetic structure of the tree Samanea saman. Conservation Biology, 16, 1-11. [ Links ]
Colling, G., & Matthies, D. (2004). The effects of plant population size on the interactions between the endangered plant Scorzonera humilis, a specialised herbivore, and a phytopathogenic fungus. Oikos, 105, 71-78. [ Links ]
Costin, B. J., Morgan, J. W., & Young, A. G. (2001). Reproductive success foes not decline in fragmented populations of Leucochrysum albicans subsp. Albicans var. tricolor (Asteraceae). Biological Conservation, 98, 273-284. [ Links ]
Cunningham, S. A. (2000). Depressed pollination in habitat fragments causes low fruit set. Proceedings of the Royal Society of London, B, 267, 1149-1152. [ Links ]
De Jong, T. J., & Klinkhamer, P. G. L. (1989). Limiting factors for seed production in Cynoglossum officinale. Oecologia, 80, 167-171. [ Links ]
Dick, C. W. (2001). Genetic rescue of remnant tropical trees by an alien pollinator. Proceedings of the Royal Society of London B, 268, 2391-2396. [ Links ]
Didham, R. K., Ghazoul, J., Stork, N. E., & Davis, A. J. (1996). Insects in fragmented forests: a functional approach. Trends in Ecology & Evolution, 11, 255-260. [ Links ]
Donaldson, J., Nänni, I., Zachariades, C., & Kemper, J. (2002). Effects of Habitat Fragmentation on Pollinator Diversity and Plant Reproductive Success in Renosterveld Shrublands of South Africa. Conservation Biology, 16, 1267-1276. [ Links ]
Dunley, B., Freitas, L., & Galetto, L. (2009). Reproduction of Byrsonima sericea (Malpighiaceae) in resting fragmented habitats in Southeastern Brazil. Biotropica, 41, 692-699. [ Links ]
Ehlers, B. K., & Olesen, J. M. (2003). Flower and fruit herbivory in a population of Centaurea scabiosa (Asteraceae): importance of population size and isolation. Ecoscience, 10, 45-48. [ Links ]
Elias, M. A. S., Franceschinelli, E. V., Borges, F. J. A., Ferreira, G. M., Juen, L., Carvalho, F. M. V. (2012). Reproductive success of Cardiopetalum calophyllum (Annonaceae) in fragments of Brazilian savanna. Journal of Tropical Ecology, 28, 317-320. [ Links ]
Faegri, K., & van der Pijl, L. (1979). The principles of pollination ecology. New York: Pergamon Press. [ Links ]
Farwig, N., Bleher, B., von der Gönna, S., & Böhning-Gaese, K. (2008). Does forest fragmentation and selective logging affect seed predators and seed predation rates of Prunus africana (Rosaceae)? Biotropica, 40(2), 218-224. [ Links ]
Fleury, M., & Galetti, M. (2006). Forest fragment size and microhabitat effects on palm seed predation. Biological Conservation, 131, 1-13. [ Links ]
França, G. S., & Stehmann, J. R. (2004). Composição florística e estrutura do componente arbóreo de uma floresta altimontana no município de Camanducaia, Minas Gerais, Brasil. Revista Brasileira de Botânica, 27, 19-30. [ Links ]
Galetti, M., Donatti, C., & Pires, A. S. (2006) Seed survival and dispersal of an endemic Atlantic forest palm: the combined effects of defaunation and forest fragmentation. Botanical Journal of Linnean Society, 151, 141-149. [ Links ]
Ghazoul, J. (2005). Pollen and seed dispersal among dispersed plants. Biological Review, 80, 413-443. [ Links ]
Greig, N. (1993). Predispersal seed predation on five Piper species in tropical rainforest. Oecologia, 93, 412-420. [ Links ]
Groom, M. J. (2001). Consequences of subpopulation isolation for pollination, herbivory, and population growth in Clarkia concinna (Onagraceae). Biological Conservation, 100, 55-63. [ Links ]
Hämback, P. A., & Beckerman, A. P. (2003). Herbivory and plant resource competition: a review of two interacting interactions. Oikos, 101, 26-37. [ Links ]
Herrerías-Diego, Y, Quesada, M., Stoner, K. E., Lobo, J. A., Hernández-Flores, Y., & Montoya, G. S. (2008). Effect of forest fragmentation on fruit and seed predation of the tropical dry forest tree Ceiba aesculifolia. Biological Conservation, 141(1), 241-248. [ Links ]
Jennersten, O. (1988). Pollination in Dianthus deltoids (Caryophyllaceae): Effects of habitat fragmentation on visitation and seed set. Conservation Biology, 2, 359-367. [ Links ]
Jennersten, O., & Nilsson, S. G. (1993). Insect flower visitation frequency and seed production in relation to patch size of Viscaria vulgaris (Caryophyllaceae). Oikos, 68, 283-292. [ Links ]
Jules, E. S. (1998). Habitat fragmentation and demographic chances for a common plant: Trillium in old-growth forest. Ecology, 79, 1645-1656. [ Links ]
Kéry, M., Matthies, D., & Fischer, M. (2001). The effect of plant population size on the interactions between the rare plant Gentiana cruciata and its specialized herbivore Maculinea rebeli. Journal of Ecology, 89, 418-427. [ Links ]
Kolb, A. (2008). Habitat fragmentation reduces plant fitness by disturbing pollination and modifying response to herbivory. Biological Conservation, 141, 2540-2549. [ Links ]
Kruess, A., & Tscharntke, T. (1994) Habitat fragmentation, species loss, and biological control. Science, 264, 1581-1584. [ Links ]
Kwak, M. M., Velterop, O., & Andel, J. (1998). Pollen and gene flow in fragmented habitats. Applied Vegetation Science, 1, 37-54. [ Links ]
Lopes, L. E., & Buzato, S. (2007). Variation in pollinator assemblages in a fragmented landscape and its effects on reproductive stages of a self-incompatible treelet Psychotria suterella (Rubiaceae). Oecologia, 154, 305-314. [ Links ]
Louda, S. M. (1982). Distribution ecology: variation in plant recruitment over a gradient in relation to insect seed predation. Ecological Monograph, 52, 25-41. [ Links ]
Lovejoy, T. E. (1980). Discontinuous wilderness: minimum areas for conservation. Parks, 5, 5-13. [ Links ]
Matheson, A., Buchamann, S., O’Toole, C., Westrich, P., & Williams, I. H. (1996). The conservation of bees. London: Academy Press. [ Links ]
Melo, M. S., Oliveira, D. E., & Franceschinelli, E. V., 2014. Low density and fertility of Murici (Byrsonima pachyphylla, Malpighiaceae) in small Cerrado remnants. Acta Botanica Brasilica, 28, 59-64. [ Links ]
Murcia, C. (1990). Effect of floral morphology on pollen receipt and removal in Ipomoea trichocarpa. Ecology, 71, 1098-1109. [ Links ]
Murcia, C. (1996). Forest fragmentation and the pollination on of Neotropical plants. In J. Schelhas, & R. Greenberg (Eds.) Forest Patches in tropical landscapes (pp. 19-37). Washington, DC: Island Press. [ Links ]
Nóbrega, A. M. F., Valeri, S. V., Paula, R. C., and Silva, S. A. (2008). Regeneracao natural em remanescentes florestais e áreas reflorestadas da várzea do rio Mogi-Guacu, Luiz Antonio-SP. Revista Arvore, 32, 909-920. [ Links ]
Oostermeijer, J. G. B., Altenburg, R.G. M., & den Nijs, H. C. M. (1995). Effects of outcrossing distance and selfing on fitness components in the rare Gentiana pneumonanthe (Gentianaceae). Acta Botanica Neerlandica, 44, 257-268. [ Links ]
Orrock, J. L., Danielson, B. J., & Burns, M. J. (2003). Spatial ecology of predator-prey interactions: corridors and patch shape influence seed predation. Ecology, 84, 2589-2599. [ Links ]
Orrock, J. L., Levey, D. J., & Danielson, B. J. (2006). Seed predation, not seed dispersal, explains the landscape-level abundance of an early-successional plant. Journal of Ecology, 94, 838-845. [ Links ]
Pinto, L. P., Hirota, M., & Fonseca, M. (2002). Panorama da Mata Atlântica e dos Campos Sulinos. In: Maury, C. M. (Ed). Biodiversidade brasileira: avaliação e identificação de áreas e ações prioritárias par conservação, utilização sustentável e repartição de benefícios da biodiversidade brasileira (pp. 215-266). Brasília DF: Ministério do Meio Ambiente. [ Links ]
Pizo, M. A. (1997). Seed dispersal and predation in two populations of Cabralea canjerana (Meliaceae) in the Atlantic Forest of southeastern Brazil. Journal of Tropical Ecology, 13, 559-577. [ Links ]
Powell, A. H., & Powell, J. V. N. (1987). Population dynamics of male euglossine bees in Amazonian forest fragments. Biotropica, 19, 176-179. [ Links ]
Prach, K., Pysek, P., & Smilauer, P. (1997). Changes in species traits during succession: A search for pattern. Oikos, 79, 201-205. [ Links ]
Quesada, M., Stoner, K. E., Lobo, J. A., Herrerias-Diego, Y., Palacios-Guevara, C., Munguía-Rosas, M. A., Salazar, K. A. O., & Rosas-Guerrero, V. (2004). Effects of forest fragmentation on pollinator activity and consequences for plant reproductive success and mating patterns in bat-pollinated Bombacaceous trees. Biotropica, 36, 131-138. [ Links ]
Quesada, M., Stoner, K. E., Rosas-Guerrero, V., Palacios-Guevara, C., & Lobo, J. A. (2003). Effects of habitat disruption on the activity of nectarivorous bats (Chiroptera : Phyllostomidae) in a dry tropical forest: implications for the reproductive success of the neotropical tree Ceiba grandiflora. Oecologia, 135, 400-406. [ Links ]
R Development Core Team (2008). An Introduction to R: Notes on R, A Programming Environment for Data Analysis and Graphics (electronic edition): http://cran.r-project.org/doc/manuals/R-intro.html. [ Links ]
Rathcke, B. J., & Jules, E. S. (1993). Habitat fragmentation and plant-pollinator interactions. Current Science, 65, 273-276. [ Links ]
Roubik, D. W. (1993). Ecology and natural history of tropical bees. Cambridge, UK: Cambridge University Press. [ Links ]
Santos, T., & Telleria, J. L. (1994). Influence of forest fragmentation on seed consumption and dispersal of Spanish juniper Juniperus thurifera. Biological Conservation, 70, 129-134. [ Links ]
Saunders, D. A., Hobbs, R. J., & Margules, C. R. (1991). Biological consequences of ecosystem fragmentation: A Review. Conservation Biology, 5, 18-32. [ Links ]
Simberloff, D. (1993). Effects of fragmentation on some Florida ecosystems, and how to redress them. In D. A. Saunders, R. J. Hobbs, & P. R. Ehrlich (Eds.) Nature Conservation 3: Reconstruction of fragmented ecosystems (pp. 174-187). Chipping Norton: Surrey Beatty & Sons. [ Links ]
Tabarelli, M., Montovani, W., & Peres, C. A. (1999). Effects of habitat fragmentation on plant guild structure in the montane Atlantic forest of Southeastern Brazil. Biological Conservation, 91, 119-127. [ Links ]
Wagenius, S., & Lyon, S. P. (2010). Reproduction of Echinacea angustifolia in fragmented prairie is pollen-limited but not pollinator-limited. Ecology, 91, 733-742. [ Links ]
Young, H. (2002). Diurnal and nocturnal pollination of Silene alba (Caryophyllaceae). American Journal of Botany, 89, 433-440. [ Links ]
Zabel, J., & Tscharntke, T. 1998. Does fragmentation of Urtica habitat affect phytophagous and predatory insect differentially? Oecologia, 116, 419-425. [ Links ]
Aguilar, R., & Galetto, L. (2004). Effects of forest fragmentation on male and female reproductive success in Cestrum parqui (Solanaceae). Oecologia, 138, 513-520. [ Links ]
Aguilar, R., Ashworth, L., Galetto, L., & Aizen, M. A. (2006). Plant reproductive susceptibility to habitat fragmentation: review and synthesis through a meta-analysis. Ecology Letters, 9, 968-980. [ Links ]
Aguirre, A., & Dirzo, R. (2008). Effects of fragmentation on pollinator abundance and fruit set of an abundant understory palm in a Mexican tropical forest. Biological Conservation, 141, 75-384. [ Links ]
Aizen, M. A., & Feinsinger, P. (1994b). Habitat fragmentation, natives insect pollinators and and feral honey bee in Argentine ‘Chaco Serrano’. Ecological Applications, 4, 378-392. [ Links ]
Aizen, M. A., & Feinsinger, P. (1994a). Forest fragmentation, pollination and plant reproduction in a Chaco dry forest, Argentina. Ecology, 75, 330-351. [ Links ]
Aldrich, P. R., & Hamrick, J. L. (1998). Reproductive dominance of pasture trees in a fragmented tropical forest mosaic. Science, 281, 103-105. [ Links ]
Bates, D., Maechler, M., Bolker, B., & Walker S. (2014). Lme4: Linear mixed-effects models using Eigen and S4. R package version 1.1-7, http://CRAN.R-project.org/package=lme4. [ Links ]
Bolker, B. M., Brooks, M. E. Clark, C. J., Geange, S. W., Poulsen, J. R., Stevens, M. H. H., & White, J. S. (2009). Generalized linear mixed models: a practical guide for ecology and evolution. Trends in Ecology & Evolution, 24, 127-135. [ Links ]
Byers, D. L. (1995). Pollen quantity and quality as explanations for low seed set in small populations exemplified by Eupatorium (Asteraceae). American Journal of Botany, 82, 1000-1006. [ Links ]
Carmo, R. M. (2005). Biologia reprodutiva de Cabralea canjerana subsp. canjerana em Fragmentos de Mata Atlântica do Sul do estado Minas Gerais. (Ph.D. thesis). Universidade Federal de Minas Gerais, Brasil. [ Links ]
Carvalho, P. E. R. (1994). Espécies florestais brasileiras. Recomendações silviculturais, potencialidades e uso da madeira. Brasília DF: Embrapa/CNPF-SPI. [ Links ]
Cascante, A., Quesada, M., Lobo, J., & Fuchs, E. A. (2002). Effects of Dry Tropical Forest Fragmentation on the reproductive success and genetic structure of the tree Samanea saman. Conservation Biology, 16, 1-11. [ Links ]
Colling, G., & Matthies, D. (2004). The effects of plant population size on the interactions between the endangered plant Scorzonera humilis, a specialised herbivore, and a phytopathogenic fungus. Oikos, 105, 71-78. [ Links ]
Costin, B. J., Morgan, J. W., & Young, A. G. (2001). Reproductive success foes not decline in fragmented populations of Leucochrysum albicans subsp. Albicans var. tricolor (Asteraceae). Biological Conservation, 98, 273-284. [ Links ]
Cunningham, S. A. (2000). Depressed pollination in habitat fragments causes low fruit set. Proceedings of the Royal Society of London, B, 267, 1149-1152. [ Links ]
De Jong, T. J., & Klinkhamer, P. G. L. (1989). Limiting factors for seed production in Cynoglossum officinale. Oecologia, 80, 167-171. [ Links ]
Dick, C. W. (2001). Genetic rescue of remnant tropical trees by an alien pollinator. Proceedings of the Royal Society of London B, 268, 2391-2396. [ Links ]
Didham, R. K., Ghazoul, J., Stork, N. E., & Davis, A. J. (1996). Insects in fragmented forests: a functional approach. Trends in Ecology & Evolution, 11, 255-260. [ Links ]
Donaldson, J., Nänni, I., Zachariades, C., & Kemper, J. (2002). Effects of Habitat Fragmentation on Pollinator Diversity and Plant Reproductive Success in Renosterveld Shrublands of South Africa. Conservation Biology, 16, 1267-1276. [ Links ]
Dunley, B., Freitas, L., & Galetto, L. (2009). Reproduction of Byrsonima sericea (Malpighiaceae) in resting fragmented habitats in Southeastern Brazil. Biotropica, 41, 692-699. [ Links ]
Ehlers, B. K., & Olesen, J. M. (2003). Flower and fruit herbivory in a population of Centaurea scabiosa (Asteraceae): importance of population size and isolation. Ecoscience, 10, 45-48. [ Links ]
Elias, M. A. S., Franceschinelli, E. V., Borges, F. J. A., Ferreira, G. M., Juen, L., Carvalho, F. M. V. (2012). Reproductive success of Cardiopetalum calophyllum (Annonaceae) in fragments of Brazilian savanna. Journal of Tropical Ecology, 28, 317-320. [ Links ]
Faegri, K., & van der Pijl, L. (1979). The principles of pollination ecology. New York: Pergamon Press. [ Links ]
Farwig, N., Bleher, B., von der Gönna, S., & Böhning-Gaese, K. (2008). Does forest fragmentation and selective logging affect seed predators and seed predation rates of Prunus africana (Rosaceae)? Biotropica, 40(2), 218-224. [ Links ]
Fleury, M., & Galetti, M. (2006). Forest fragment size and microhabitat effects on palm seed predation. Biological Conservation, 131, 1-13. [ Links ]
França, G. S., & Stehmann, J. R. (2004). Composição florística e estrutura do componente arbóreo de uma floresta altimontana no município de Camanducaia, Minas Gerais, Brasil. Revista Brasileira de Botânica, 27, 19-30. [ Links ]
Galetti, M., Donatti, C., & Pires, A. S. (2006) Seed survival and dispersal of an endemic Atlantic forest palm: the combined effects of defaunation and forest fragmentation. Botanical Journal of Linnean Society, 151, 141-149. [ Links ]
Ghazoul, J. (2005). Pollen and seed dispersal among dispersed plants. Biological Review, 80, 413-443. [ Links ]
Greig, N. (1993). Predispersal seed predation on five Piper species in tropical rainforest. Oecologia, 93, 412-420. [ Links ]
Groom, M. J. (2001). Consequences of subpopulation isolation for pollination, herbivory, and population growth in Clarkia concinna (Onagraceae). Biological Conservation, 100, 55-63. [ Links ]
Hämback, P. A., & Beckerman, A. P. (2003). Herbivory and plant resource competition: a review of two interacting interactions. Oikos, 101, 26-37. [ Links ]
Herrerías-Diego, Y, Quesada, M., Stoner, K. E., Lobo, J. A., Hernández-Flores, Y., & Montoya, G. S. (2008). Effect of forest fragmentation on fruit and seed predation of the tropical dry forest tree Ceiba aesculifolia. Biological Conservation, 141(1), 241-248. [ Links ]
Jennersten, O. (1988). Pollination in Dianthus deltoids (Caryophyllaceae): Effects of habitat fragmentation on visitation and seed set. Conservation Biology, 2, 359-367. [ Links ]
Jennersten, O., & Nilsson, S. G. (1993). Insect flower visitation frequency and seed production in relation to patch size of Viscaria vulgaris (Caryophyllaceae). Oikos, 68, 283-292. [ Links ]
Jules, E. S. (1998). Habitat fragmentation and demographic chances for a common plant: Trillium in old-growth forest. Ecology, 79, 1645-1656. [ Links ]
Kéry, M., Matthies, D., & Fischer, M. (2001). The effect of plant population size on the interactions between the rare plant Gentiana cruciata and its specialized herbivore Maculinea rebeli. Journal of Ecology, 89, 418-427. [ Links ]
Kolb, A. (2008). Habitat fragmentation reduces plant fitness by disturbing pollination and modifying response to herbivory. Biological Conservation, 141, 2540-2549. [ Links ]
Kruess, A., & Tscharntke, T. (1994) Habitat fragmentation, species loss, and biological control. Science, 264, 1581-1584. [ Links ]
Kwak, M. M., Velterop, O., & Andel, J. (1998). Pollen and gene flow in fragmented habitats. Applied Vegetation Science, 1, 37-54. [ Links ]
Lopes, L. E., & Buzato, S. (2007). Variation in pollinator assemblages in a fragmented landscape and its effects on reproductive stages of a self-incompatible treelet Psychotria suterella (Rubiaceae). Oecologia, 154, 305-314. [ Links ]
Louda, S. M. (1982). Distribution ecology: variation in plant recruitment over a gradient in relation to insect seed predation. Ecological Monograph, 52, 25-41. [ Links ]
Lovejoy, T. E. (1980). Discontinuous wilderness: minimum areas for conservation. Parks, 5, 5-13. [ Links ]
Matheson, A., Buchamann, S., O’Toole, C., Westrich, P., & Williams, I. H. (1996). The conservation of bees. London: Academy Press. [ Links ]
Melo, M. S., Oliveira, D. E., & Franceschinelli, E. V., 2014. Low density and fertility of Murici (Byrsonima pachyphylla, Malpighiaceae) in small Cerrado remnants. Acta Botanica Brasilica, 28, 59-64. [ Links ]
Murcia, C. (1990). Effect of floral morphology on pollen receipt and removal in Ipomoea trichocarpa. Ecology, 71, 1098-1109. [ Links ]
Murcia, C. (1996). Forest fragmentation and the pollination on of Neotropical plants. In J. Schelhas, & R. Greenberg (Eds.) Forest Patches in tropical landscapes (pp. 19-37). Washington, DC: Island Press. [ Links ]
Nóbrega, A. M. F., Valeri, S. V., Paula, R. C., and Silva, S. A. (2008). Regeneracao natural em remanescentes florestais e áreas reflorestadas da várzea do rio Mogi-Guacu, Luiz Antonio-SP. Revista Arvore, 32, 909-920. [ Links ]
Oostermeijer, J. G. B., Altenburg, R.G. M., & den Nijs, H. C. M. (1995). Effects of outcrossing distance and selfing on fitness components in the rare Gentiana pneumonanthe (Gentianaceae). Acta Botanica Neerlandica, 44, 257-268. [ Links ]
Orrock, J. L., Danielson, B. J., & Burns, M. J. (2003). Spatial ecology of predator-prey interactions: corridors and patch shape influence seed predation. Ecology, 84, 2589-2599. [ Links ]
Orrock, J. L., Levey, D. J., & Danielson, B. J. (2006). Seed predation, not seed dispersal, explains the landscape-level abundance of an early-successional plant. Journal of Ecology, 94, 838-845. [ Links ]
Pinto, L. P., Hirota, M., & Fonseca, M. (2002). Panorama da Mata Atlântica e dos Campos Sulinos. In: Maury, C. M. (Ed). Biodiversidade brasileira: avaliação e identificação de áreas e ações prioritárias par conservação, utilização sustentável e repartição de benefícios da biodiversidade brasileira (pp. 215-266). Brasília DF: Ministério do Meio Ambiente. [ Links ]
Pizo, M. A. (1997). Seed dispersal and predation in two populations of Cabralea canjerana (Meliaceae) in the Atlantic Forest of southeastern Brazil. Journal of Tropical Ecology, 13, 559-577. [ Links ]
Powell, A. H., & Powell, J. V. N. (1987). Population dynamics of male euglossine bees in Amazonian forest fragments. Biotropica, 19, 176-179. [ Links ]
Prach, K., Pysek, P., & Smilauer, P. (1997). Changes in species traits during succession: A search for pattern. Oikos, 79, 201-205. [ Links ]
Quesada, M., Stoner, K. E., Lobo, J. A., Herrerias-Diego, Y., Palacios-Guevara, C., Munguía-Rosas, M. A., Salazar, K. A. O., & Rosas-Guerrero, V. (2004). Effects of forest fragmentation on pollinator activity and consequences for plant reproductive success and mating patterns in bat-pollinated Bombacaceous trees. Biotropica, 36, 131-138. [ Links ]
Quesada, M., Stoner, K. E., Rosas-Guerrero, V., Palacios-Guevara, C., & Lobo, J. A. (2003). Effects of habitat disruption on the activity of nectarivorous bats (Chiroptera : Phyllostomidae) in a dry tropical forest: implications for the reproductive success of the neotropical tree Ceiba grandiflora. Oecologia, 135, 400-406. [ Links ]
R Development Core Team (2008). An Introduction to R: Notes on R, A Programming Environment for Data Analysis and Graphics (electronic edition): http://cran.r-project.org/doc/manuals/R-intro.html. [ Links ]
Rathcke, B. J., & Jules, E. S. (1993). Habitat fragmentation and plant-pollinator interactions. Current Science, 65, 273-276. [ Links ]
Roubik, D. W. (1993). Ecology and natural history of tropical bees. Cambridge, UK: Cambridge University Press. [ Links ]
Santos, T., & Telleria, J. L. (1994). Influence of forest fragmentation on seed consumption and dispersal of Spanish juniper Juniperus thurifera. Biological Conservation, 70, 129-134. [ Links ]
Saunders, D. A., Hobbs, R. J., & Margules, C. R. (1991). Biological consequences of ecosystem fragmentation: A Review. Conservation Biology, 5, 18-32. [ Links ]
Simberloff, D. (1993). Effects of fragmentation on some Florida ecosystems, and how to redress them. In D. A. Saunders, R. J. Hobbs, & P. R. Ehrlich (Eds.) Nature Conservation 3: Reconstruction of fragmented ecosystems (pp. 174-187). Chipping Norton: Surrey Beatty & Sons. [ Links ]
Tabarelli, M., Montovani, W., & Peres, C. A. (1999). Effects of habitat fragmentation on plant guild structure in the montane Atlantic forest of Southeastern Brazil. Biological Conservation, 91, 119-127. [ Links ]
Wagenius, S., & Lyon, S. P. (2010). Reproduction of Echinacea angustifolia in fragmented prairie is pollen-limited but not pollinator-limited. Ecology, 91, 733-742. [ Links ]
Young, H. (2002). Diurnal and nocturnal pollination of Silene alba (Caryophyllaceae). American Journal of Botany, 89, 433-440. [ Links ]
Zabel, J., & Tscharntke, T. 1998. Does fragmentation of Urtica habitat affect phytophagous and predatory insect differentially? Oecologia, 116, 419-425. [ Links ]
1. Departamento de Botânica, Instituto de Ciências Biológicas, Universidade Federal de Goiás, Campus II-Samambaia; Goiânia-GO, Brasil; edivanif@gmail.com, goncalves.b.b@gmail.com, carloskoa@gmail.com
2. Departamento de Botânica, ICB, Universidade Federal de Minas Gerais, Av. Antônio Carlos 6627, Pampulha, Belo Horizonte MG, Brazil; roselainicarmo@gmail.com
3. Departamento de Ecologia, Instituto de Ciências Biológicas, Universidade Federal de Goiás, Campus II - Samambaia; Goiânia - GO, Brasil; llbergamini@gmail.com
Received 21-IV-2014. Corrected 07-X-2014. Accepted 19-XI-2014.


{kind=link}
{kind=link}