










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.2 San José Apr./Jun. 2015
Nest site selection by individual leatherback turtles (Dermochelys coriacea, Testudines: Dermochelyidae) in Tortuguero, Caribbean coast of Costa Rica
Selección del sitio de anidación de individuos de tortuga baula (Dermochelys coriacea, Testudines: Dermochelyidae) en Tortuguero, costa Caribe de Costa Rica
Selección del sitio de anidación de individuos de tortuga baula (Dermochelys coriacea, Testudines: Dermochelyidae) en Tortuguero, costa Caribe de Costa Rica
Abstract
Nest site selection for individual leatherback sea turtles, Dermochelys coriacea, is a matter of dispute. Some authors suggest that a female will tend to randomly scatter her nests to optimize clutch survival at a highly dynamic beach, while others suggest that some site fidelity exists. It is also possible that both strategies exist, depending on the characteristics of each nesting beach, with stable beaches leading to repeating nest site selections and unstable beaches leading to nest scattering. To determine the strategy of the Tortuguero population of D. coriacea, female site preference and repetition were determined by studying whether females repeat their nest zone choices between successive attempts and whether this leads to a correlation in hatching and emergence success of subsequent nests. Nesting data from 1997 to 2008 were used. Perpendicular to the coastline, open sand was preferred in general, regardless of initial choice. This shows a tendency to scatter nests and is consistent with the fact that all vertical zones had a high variability in hatching and emergence success. It is also consistent with nest success not being easily predictable, as shown by the lack of correlation in success of subsequent nesting attempts. Along the coastline, turtles showed a preference for the middle part of the studied section of beach, both at a population level and as a tendency to repeat their initial choice. Interestingly, this zone has the most artificial lights, which leads to slightly lower nest success (though not significantly so) and hatchling disorientation. This finding merits further study for a possibly maladaptive trait and shows the need for increased control of artificial nesting on this beach.
Key words: scatter nest hypothesis, Dermochelys coriacea, nest site repeatability, nesting patterns, nest distribution.
Resumen
La selección de sitio de anidación por parte de individuos de la tortuga baula, Dermochelys coriacea, es tema de discusión. Algunos autores sugieren que una hembra dispersa sus nidos al azar para maximizar su éxito en las inestables playas que elige para anidar, mientras que otros sugieren que existe cierta fidelidad al sitio de anidación. También es posible que ambas estrategias existan, dependiendo de las características de la playa donde anidan, así playas estables llevarían a las hembras a repetir la selección de sitio mientras que las playas inestables las llevarían a dispersar sus nidos. Para determinar la estrategia que usan las hembras de D. coriacea de Tortuguero, se estudió la preferencia de sitio de las hembras así como su repetición en este sitio, desde 1997 hasta 2008. Para la selección de sitio vertical (perpendicular a la costa), las hembras en general prefirieron anidar en arena abierta, independientemente de su escogencia inicial. Esto muestra una tendencia a dispersar los nidos y es consistente con la alta variación en los éxitos de eclosión en todas las zonas, así como la falta de relación entre los éxitos de eclosión de nidos subsecuentes, lo cual muestra que el éxito de los nidos no es predecible. Para la selección horizontal (a lo largo de la costa) se prefirió la parte media de la sección estudiada de la playa, tanto a nivel poblacional como individual (repetición de selección de sitio). Interesantemente, esta zona tiene la mayor cantidad de luces artificiales lo cual lleva a éxitos de eclosión ligeramente más bajos (aunque no significativamente) y a la desorientación de neonatos. Este resultado amerita estudios futuros para determinar si se trata de una tendencia maladaptativa en la población y muestra la importancia de controlar la cantidad de luz artificial en esta playa como medida de conservación.
Palabras clave: hipótesis de dispersión de nidos, Dermochelys coriacea, repetición de sitios de anidación, patrones de anidación, distribución de nidos.
Leatherback females nest every 3-5 years on tropical and subtropical beaches (Sarti Martinez, 2000; Bell, Spotila, Paladino, & Reina, 2003), and can nest up to 13 times in a single season (Reina, Mayor, Spotila, Piedra, & Paladino, 2002). They tend to nest on highly dynamic beaches, some with considerable changes in tide and erosion levels (Eckert, 1987), and may move between different beaches within and between nesting seasons (Pritchard, 1982; Whitmore, & Dutton, 1985; Runemark, 2006).
Adequate nest site selection is a balance between nesting too close to the high tide line, risking nests being washed out (Runemark, 2006), and nesting too close to the vegetation, risking invasion by roots, predation, and possible hatchling disorientation due to artificial lighting on some beaches (Caut, Guirlet, Jouquet, & Girondot, 2006; Katilimis, Urhan, Kaska, & Baskale, 2006; Runemark, 2006).
It has been suggested that sea turtles do not select specific nest sites but rather scatter their nests randomly in order to optimize clutch survival probability (Carr, & Ogren, 1959; Pritchard, 1982; Eckert, 1987; Kamel, & Mrosovsky, 2004). Other authors have suggested that some nest site repetition might exist, with certain females preferring to lay closer to the high tide line and others laying higher up the beach (Kamel, & Mrosovsky, 2004; Mrosovsky, 2006). According to Eckert (1987), females can display either of these strategies depending on the predictability of nest perturbation, with more stable habitats leading to predictable, repeated nesting trends, and unstable habitats leading to scattered nesting.
Nest choice repetition has not been studied in depth and may be used to infer evolutionary potential, with high repetition implying a possibility for a heritable trait (Kamel, & Mrosovsky, 2004; Mrosovsky, 2006). This is a concern because the decreasing population size of leatherbacks (Spotila, Dunham, & Leslie, 1996; Spotila, Reina, Steyermark, Plotkin, & Paladino, 2000; Sarti Martinez, 2000; Troëng, Chacón, & Dick, 2004; NOAA, 2007; Santidrián-Tomillo et al., 2007; Wallace et al., 2011) has prompted the relocation of doomed nests as a conservation strategy (Eckert, & Eckert, 1990; Dutton, Dutton, Chaloupka, & Boulon, 2005; Pike, 2008). Doubts have been raised about whether or not this would select for “bad nesters” (Mrosovsky, 1983; Pike, 2008), females that repeatedly place their nests in low success sites (Kamel, & Mrosovsky, 2004).
The present study aimed to determine whether individual leatherbacks nesting at Tortuguero, Caribbean coast of Costa Rica (a highly dynamic beach), show nest site preferences. This was done by analyzing subsequent nest locations as well as how successful the selected sites were.
Materials and Methods
Study site: This study was carried out at Tortuguero on the Caribbean coast of Costa Rica, in the province of Limón. The 35.6km (22¼mile) long black sand beach is located between the Tortuguero and Parismina river mouths (Fig. 1). For historical reasons, the beach was marked into 1/8mile sections, from North to South (Fowler, 1979). The beach was subject to erosion as well as strong tides and was, therefore, commonly covered by logs and other debris. This may limit nest site availability and influence the movement of sea turtles on the beach (Runemark, 2006; Sarmiento, 2007). Tortuguero village is located mostly (with the exception of a few hotels and the airport) between mile markers 2 5/8 and 3 3/8. Tortuguero National Park starts at mile 3 3/8, and the majority of the beach was within its limits (N. Neeman, pers. obs.).
Data collection: The fieldwork was partially conducted by the first author (NN) as one of several Sea Turtle Conservancy (STC, formerly Caribbean Conservation Corporation) research assistants during the 2007 and 2008 field seasons, as well as the field coordinators. The data were collected during the STC Leatherback Program (which runs annually from March through June), following the current Leatherback Program Monitoring Protocol, with minor modifications since 1997 and detailed in STC’s annual reports (Atkinson, Nolasco, & Harrison, 2011). Data from years prior to 2007 were used with the permission of STC. Overall, the data used herein are from 1997 to 2008, which corresponds to 12 nesting seasons.
For logistical reasons, the fieldwork was concentrated on the northernmost five miles of the beach (Fig. 1), for which monitoring effort and coverage were uniform. Every night of each nesting season, this area was patrolled on foot by at least two research assistants, following variable schedules (normally five hour shifts, between 8pm and 4am). The research assistants identified individual leatherbacks by metal tags placed on their rear flippers; if no tags were found, they attached two new tags and the number or alphanumeric series were recorded. For all leatherbacks that were encountered while the egg chamber was open, research assistants marked the exact nest location by placing three pieces of flagging tape in the vegetation and measuring the distance from each tape to the centre of the egg chamber. The distance from the nest to the most recent, visible high tide line was recorded. The zone of the nest was also noted, using one of three categories: vegetation or full shading (v), border or partial shading (b), and open or no shading (o).
Marked nests were checked each morning during the incubation period to monitor their fate; any evidence that the nest had been washed over or away, predated or poached was recorded. Visible signs of hatching (such as sand caving above the egg chamber or hatchling tracks) were also noted. Two days after hatching was reported, the nest was excavated. For nests where no signs of hatching were observed, excavation took place 75 days after the nest was laid. During excavation eggs were categorized as: empty shells (only counted if more than 50% of the shell was found in one piece), unhatched eggs (divided into three categories depending on the presence and development of embryos), pipped eggs (in which the hatchling broke the shell but didn’t emerge), and predated eggs. Any live or dead hatchlings encountered within the nest chamber were recorded. From this, hatching success was calculated as Ha=E/T, where E is the number of empty shells and T is the total number of eggs; emergence success was calculated as Em=(E-L-D)/T, where L and D are the number of live and dead hatchlings found at excavation, respectively (Atkinson et al., 2011).
Nesting zone definitions: Only females that were observed nesting more than once in the 12 seasons were considered for the analyses. Nest placement was described using horizontal (along the coastline) and vertical (perpendicular to the coastline) distribution on the beach. For the horizontal distribution, the beach was divided into five zones, each one mile in length, assigning zones 0-4 running from North to South: Zone 0 included some nests laid before mile marker zero, and Zone 4 included one nest laid just past the mile five marker (Fig. 1). For the vertical distribution, nests were divided into three zones, taking into account the zone noted during nesting (open sand, vegetation border or vegetation) and the distance to the position of the high tide line at the time when the nest was laid. These zones were defined in order to better reflect the use of the beach by the turtles, i.e. to show where the female chose to nest in relation to the high tide line: Zone I included nests laid below the high tide line or up to 1m above it (due to the difficulty in determining the precise location of the high tide line at night in some occasions); Zone II described nests laid more than 1m above the high tide line but not in the vegetation; Zone III corresponded to nests placed either in the vegetation or on the vegetation border.
Analysis: Since there were limited observations for each female, the data for different seasons were combined, meaning that two or more nests from the same female (either within the same nesting season or between different seasons) were used for the analyses. Subsequent observations were analyzed as pairs, i.e. if a female had more than two nests observed, there were paired as 1&2, 2&3, etc. (for simplicity, all subsequent pairs are referred to as first and second in the text). First, chi-square tests were used to determine whether or not nest placement was independent of zone along the length and width of the beach. Then, in order to analyze the relationship between subsequent nesting events, contingency tables were created for the pairs of nesting events, using the first nest site location as the column and the second location as the row. One such contingency table was created for vertical and a second one for horizontal zone selection. These tables were tested using G tests and chi-square tests of independence (Nordmoe et al., 2004; Zar, 2010). Also, a goodness of fit chi-square was applied when comparing how likely it was to repeat horizontal zone. This test summed all instances in which the second zone is different from the first one (these included four possible cases: since there were five total zones, there were four possible zones that were different from the initial zone) and compared said sum with the instances in which the initial zone was repeated (only one possible case). Initially, a homogeneity test was run to see if summing the zones was correct, and then a chi-square with Yates’s correction for continuity was made with the totals (Zar, 2010).
Both hatching and emergence success between first and second observed nesting events were correlated using a Spearman Rank test (p<0.05) since data were not normally distributed. Finally, Analyses of Variance (ANOVAs) were used to determine if hatching and emergence success differ either between the horizontal zones or between the vertical zones.
Results
The data obtained correspond to 94 pairs of subsequent nesting events of 70 different females. Most females (51) were only observed twice during the study period, 15 females were recorded three times, three were observed on four occasions, and one female was encountered five times (all five observations during the 2001 nesting season).
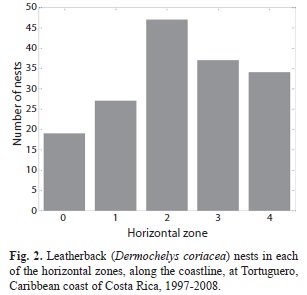
The initial analyses, treating each nest independently, revealed the following results: vertical nest zone selection was non-random, with the majority of nests (137 out of 164) located in Zone II (χ2=186.5, p<0.001, df=2). Horizontal nest site selection was also not random: more nests (47) were laid in Zone 2 and less in Zone 0 (19) than in the other zones (χ2=13.6, p=0.009, df=4, Fig. 2), which showed close to 20% of the nests each, as would be expected if selection was random and nests were distributed uniformly between the five zones.
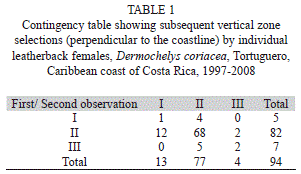
For subsequent nesting attempts, vertical zone selection was independent (G=5.8, p=0.215, df=4, Table 1). However, some patterns were apparent: turtles only moved to adjacent zones for their next nesting attempt (i.e. none moved from I to III or vice versa). The most frequent tendency (88.3% of all observed pairs) was to repeat Zone II. This zone was also chosen after initially choosing Zone I or III (Table 1). Few turtles (14 out of 82) nesting in Zone II moved to Zones I or III. When the analysis was repeated combining Zones I and III into a single category to see whether the initial selection was repeated or not, the result was dependent (χ2 with Yates’s correction for continuity=33.9, p<0.001, df=1, Table 1), which indicated that females that initially selected Zone II tended to repeat it and those that selected other zones tended to move to Zone II.
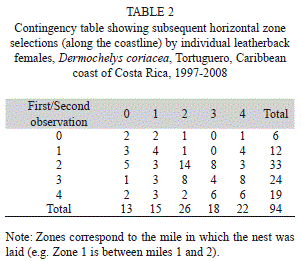
Horizontal nest zone selection was not independent (G=30.5, p=0.016, df=16, Table 2). No clear pattern was visible; however, females often repeated the selection for Zone 2 (14 out of 26 pairs of nesting events had both nests laid in Zone 2). When combining the zones, the results of a heterogeneity chi-square revealed that there were no differences between zones in the likelihood in which they are repeated (χ2=5.79, p=0.215, df=4, Table 2). However, when comparing the total number of females that repeated their initial zone selection (30 in one zone) with those that chose different zones (64 in four zones), it was more likely that the initial choice was repeated (χ 2 with Yates’s correction for continuity =7.61, p=0.006, df=1, Table 2). Most of the subsequent nests (83 out of 94) were encountered within two miles of the first nesting zone.
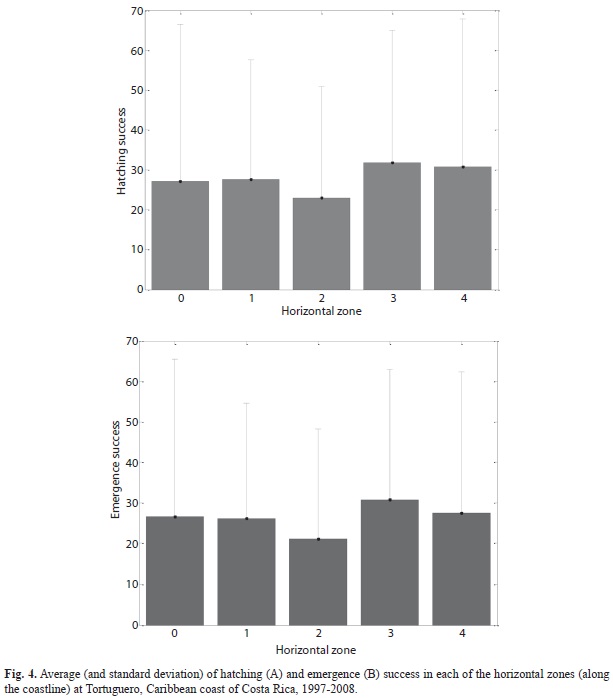
No correlation was found between hatching success of the first nesting event with the second (r=0.21, p=0.090, n=70), nor between these events’ emergence success (r=0.19, p=0.125, n=70). Hatching and emergence success were not different between the vertical zones (hatching: F (2, 137)=2.82, p=0.063; emergence: F (2, 137)=2.64, p=0.075; Fig. 3) or between the horizontal zones (hatching: F (5, 134)=0.59, p=0.701; emergence: F (5, 134)=0.69, p=0.631; Fig. 4).
Discussion
For vertical nest selection, females showed an overall preference for open sand. This was independent of their initial nest choice, thus reflecting a trend at the population level, as opposed to some females specializing in open sand and others in the vegetation or close to the high tide line. The preference is consistent with observations for this species at other locations (Whitmore, & Dutton, 1985; Kamel, & Mrosovsky, 2004; Nordmoe et al., 2004) as well as the idea of balancing the negative consequences of nesting too close to the ocean with those of nesting too close to the vegetation. According to several authors (Caut et al., 2006; Katilimis et al., 2006; Mrosovsky, 2006; Runemark, 2006), this area may be preferred due to its optimal humidity and temperature conditions, though the present study did not find higher hatching or emergence success in this zone. Further studies should compare the physical and chemical characteristics of this part of the beach to determine why it is preferred. Local conditions have been shown to affect nest site selection in other species such as the loggerhead turtle, Caretta caretta (Lamont, & Houser, 2014). For instance, debris might accumulate less in this part of the beach making it more favorable to emergence success.
The low repetition of nest site placement (especially for nests below the high tide line, which all had a hatching success of zero) indicates that the concern for “bad nesters” is unwarranted for this population. The high variability of hatching and emergence success in all zones along with the lack of correlation in hatching and emergence success of subsequent nesting attempts suggests that success is not predictable for females and leads to scattering nests rather than repeating initial choices (Eckert, 1987). This is also consistent with Bjornal & Bolten (2010) who found that green sea turtles (Chelonia mydas) responded more to yearly variation in environmental conditions rather than following individual preferences when selecting nesting sites. However, other species such as the hawksbill turtle (Eretmochelys imbricata) have been shown to strongly repeat individual nest site selection (Kamel, & Mrosovsky, 2005).
For horizontal zone selection, although there was also a trend that mirrors the population preference, there was more evidence of repetition by individuals. This repetition is seen both as repeating the initial zone and as nesting within two miles of their initial location (regardless of whether this was in the same zone or not). A similar tendency was found by Nordmoe et al. (2004) at Playa Grande, Costa Rica, who suggested that the trend to repeat nest choice was not a preference for specific nest sites, but rather a repetition of the initial nest site, regardless of where this first choice was located. This is also apparent in the present study, as females were equally likely to repeat their initial selection in all zones.
Turtles showed a preference for nesting in the part of the beach where the village and, hence, most artificial lighting is located as determined by STC light assessments (Atkinson et al., 2011). Since these assessments were done without notifying the population as well as during nesting hours (Atkinson et al., 2011), they reflect real light use and can affect nesting. This is also reflected in the slightly lower hatching and nesting success in this zone. Additionally, artificial lights cause hatchlings to become disoriented and crawl towards the vegetation rather than towards the ocean (Whitmore, & Dutton, 1985; Witherington, & Martin, 1996; Kamel, & Mrosovsky, 2004). This is not considered in the present study but would further lower the success of these nests. Increased nesting in artificially lit areas contradicts what has been reported for leatherbacks (Nordmoe et al., 2004), though it has been suggested for other species, such as the olive ridley (Lepidochelys olivacea) (F. Bolaños, pers. obs.) and merits further research to understand the underlying reasons for this behavior. This also suggests further action should be taken to reduce artificial lighting on this nesting beach. Similarly to horizontal nest site selection, further studies may help to determine why this part of the beach is selected. It is possible that other factors such as the shape of the beach or the tides lead to favoring this zone (Lamont, & Houser, 2014).
To investigate nest site selection, it is recommended to analyze nesting preferences for individual females observed on different nesting beaches, since it is known that leatherback females may use various beaches (Sarti Martinez, 2000). Such a study could provide additional information, which might broaden our understanding of the nesting activities of individual female leatherbacks and help conservation efforts.
Acknowledgments
Our thanks go to the Sea Turtle Conservancy for providing board and lodging during the stays of the first author as a research assistant and for logistical support, as well as to all the research assistants, field coordinators and volunteer participants without whom the data would not exist. Data collection was possible thanks to a research permit from the Costa Rican Ministry of Energy, Environment and Telecommunications (MINAET). Thanks also to Tiberiu Tesileanu for much help with generating and editing figure 1.
References
Atkinson, C., Nolasco del Aguila, D., & Harrison, E. (2011). Report on the 2010 Leatherback Program at Tortuguero, Costa Rica. Caribbean Conservation Corporation. Retrieved from http://www.conserveturtles.org/pdf/reports/Tortuguero%20Leatherback%20Report%202010.pdf). [ Links ]
Bell, B., Spotila, J. R., Paladino, F. V., & Reina, R. D. (2003). Low reproductive success of leatherback turtles, Dermochelys coriacea, is due to high embryonic mortality. Biological Conservation, 115, 131-138. [ Links ]
Bjorndal, K. A., & Bolten, A. B. (2010). Spatial distribution of green turtle (Chelonia mydas) nests at Tortuguero, Costa Rica. Copeia, 1992, 45-53. [ Links ]
Carr, A., & Ogren, L. (1959). The ecology and migrations of sea turtles. 3, Dermochelys in Costa Rica. American Museum Novitates, 1958, 2-29. [ Links ]
Caut, S., Guirlet, E., Jouquet, P., & Girondot, M. (2006). Influence of nest location and yolkless eggs on the hatching success of leatherback turtle clutches in French Guiana. Canadian Journal of Zoology, 84, 908-915. [ Links ]
Dutton, D. L., Dutton, P. H., Chaloupka, M., & Boulon, R. H. (2005). Increase of a Caribbean leatherback turtle Dermochelys coriacea nesting population linked to long-term nest protection. Biological Conservation, 126, 186-194. [ Links ]
Eckert, K. (1987). Environmental unpredictability and leatherback sea turtle (Dermochelys coriacea) nest loss. Herpetologica, 43, 315-323. [ Links ]
Eckert, K. L., & Eckert, S. A. (1990). Embryo mortality and hatch success in in situ and translocated leatherback sea turtle Dermochelys coriacea eggs. Biological Conservation, 53, 37-46. [ Links ]
Fowler, L. E. (1979). Hatching success and nest predation in the green sea turtle, Chelonia mydas, at Tortuguero, Costa Rica. Ecology, 60, 946-955. [ Links ]
Kamel, S. J., & Mrosovsky, N. (2004). Nest site selection in leatherbacks, Dermochelys coriacea: individual patterns and their consequences. Animal Behaviour, 68, 357-366. [ Links ]
Kamel, S. J., & Mrosovsky, N. (2005). Repeatability of nesting preferences in the hawksbill sea turtle, Eretmochelys imbricata, and their fitness consequences. Animal Behaviour, 70, 819-828. [ Links ]
Katilimis, Y., Urhan, R., Kaska, Y., & Baskale, E. (2006). Invertebrate infestation on eggs and hatchlings of the loggerhead turtle, Caretta caretta, in Dalaman, Turkey. Biodiversity and Conservation, 15, 3721-3730. [ Links ]
Lamont, M. M., & Houser, C. (2014). Spatial distribution of loggerhead turtle (Caretta caretta) emergences along a highly dynamic beach in the northern Gulf of Mexico. Journal of Experimental Marine Biology and Ecology, 453, 98-107. [ Links ]
Mrosovsky, N. (1983). Ecology and nest-site selection of leatherback turtles Dermochelys coriacea. Biological Conservation, 26, 47-56. [ Links ]
Mrosovsky, N. (2006). Distorting gene pools by conservation: Assessing the case of doomed turtle eggs. Environmental Management, 38, 523-31. [ Links ]
NOAA (Turtle Expert Working Group). (2007). An assessment of the leatherback turtle population in the Atlantic Ocean. NOAA Technical Memorandum NMFS-SEFSC-555. [ Links ]
Nordmoe, E. D., Sieg, A. E., Sotherland, P. R., Spotila, J. R., Paladino, F. V., & Reina, R. D. (2004). Nest site fidelity of leatherback turtles at Playa Grande, Costa Rica. Animal Behaviour, 68, 387-394. [ Links ]
Pike, D. (2008). The benefits of nest relocation extend far beyond recruitment: a rejoinder to Mrosovsky. Environmental Management, 41, 461-464. [ Links ]
Pritchard, P. (1982). Nesting of the leatherback turtle, Dermochelys coriacea in Pacific Mexico, with a new estimate of the world population status. Copeia, 1982, 741-747. [ Links ]
Reina, R. D., Mayor, P. A., Spotila, J. R., Piedra, R., & Paladino, F. V. (2002). Nesting ecology of the leatherback turtle, Dermochelys coriacea, at Parque Nacional Marino Las Baulas, Costa Rica, 1988-1989 to 1999-2000. Copeia, 2002, 653-664. [ Links ]
Runemark, A. (2006). Spatial distribution and temperature effects on hatching success of the leatherback turtle Dermochelys coriacea: implications for conservation. Master thesis. Uppsala University, Sweden. [ Links ]
Santidrián-Tomillo, P., Velez, E., Reina, R. D., Piedra, R., Paladino, F. V, Spotila, J. R., & Vélez, E. (2007). Reassessment of the leatherback turtle (Dermochelys coriacea) nesting population at Parque Nacional Marino Las Baulas, Costa Rica: effects of conservation efforts. Chelonian Conservation and Biology, 6, 54-62. [ Links ]
Sarmiento, R. (2007). Caracterización del material maderifero de deriva sobre un sector de la playa de Anidación de Tortugas Marinas, Tortuguero – Costa Rica. STC Internal Document. [ Links ]
Sarti Martinez, A. L. (2000). Dermochelys coriacea. In IUCN 2006. 2006 IUCN Red List of Threatened Species. Retrieved from www.iucnredlist.org
Links ]">
Spotila, J. R., Dunham, A. E., & Leslie, A. J. (1996). Worldwide population decline of Dermochelys coriacea: Are leatherback turtles going extinct? Chelonian Conservation and Biology, 2, 209-222. [ Links ]
Spotila, J. R., Reina, R., Steyermark, A., Plotkin, P. & Paladino, F. (2000). Pacific leatherback turtles face extinction. Nature, 405, 529-530. [ Links ]
Troëng, S., Chacón, D., & Dick, B. (2004). Possible decline in leatherback turtle Dermochelys coriacea nesting along the coast of Caribbean Central America. Oryx, 38, 395-403. [ Links ]
Wallace, B. P., DiMatteo, A. D., Bolten, A. B., Chaloupka, M. Y., Hutchinson, B. J., Abreu-Grobois, F. A., & Mast, R. B. (2011). Global conservation priorities for marine turtles. PloS One, 6, e24510. [ Links ]
Whitmore, C. P., & Dutton, P. H. (1985). Infertility, embryonic mortality and nest-site selection in leatherback and green sea turtles in Suriname. Biological Conservation, 34, 251-272. [ Links ]
Witherington, B., & Martin, R. (1996). Understanding, assessing, and resolving light-pollution problems on sea turtle nesting beaches. Florida Marine Research Institute Technical Report TR-2. [ Links ]
Zar, J. H. (2010). Biostatistical Analysis. USA: Prentice Hall. [ Links ]
Bell, B., Spotila, J. R., Paladino, F. V., & Reina, R. D. (2003). Low reproductive success of leatherback turtles, Dermochelys coriacea, is due to high embryonic mortality. Biological Conservation, 115, 131-138. [ Links ]
Bjorndal, K. A., & Bolten, A. B. (2010). Spatial distribution of green turtle (Chelonia mydas) nests at Tortuguero, Costa Rica. Copeia, 1992, 45-53. [ Links ]
Carr, A., & Ogren, L. (1959). The ecology and migrations of sea turtles. 3, Dermochelys in Costa Rica. American Museum Novitates, 1958, 2-29. [ Links ]
Caut, S., Guirlet, E., Jouquet, P., & Girondot, M. (2006). Influence of nest location and yolkless eggs on the hatching success of leatherback turtle clutches in French Guiana. Canadian Journal of Zoology, 84, 908-915. [ Links ]
Dutton, D. L., Dutton, P. H., Chaloupka, M., & Boulon, R. H. (2005). Increase of a Caribbean leatherback turtle Dermochelys coriacea nesting population linked to long-term nest protection. Biological Conservation, 126, 186-194. [ Links ]
Eckert, K. (1987). Environmental unpredictability and leatherback sea turtle (Dermochelys coriacea) nest loss. Herpetologica, 43, 315-323. [ Links ]
Eckert, K. L., & Eckert, S. A. (1990). Embryo mortality and hatch success in in situ and translocated leatherback sea turtle Dermochelys coriacea eggs. Biological Conservation, 53, 37-46. [ Links ]
Fowler, L. E. (1979). Hatching success and nest predation in the green sea turtle, Chelonia mydas, at Tortuguero, Costa Rica. Ecology, 60, 946-955. [ Links ]
Kamel, S. J., & Mrosovsky, N. (2004). Nest site selection in leatherbacks, Dermochelys coriacea: individual patterns and their consequences. Animal Behaviour, 68, 357-366. [ Links ]
Kamel, S. J., & Mrosovsky, N. (2005). Repeatability of nesting preferences in the hawksbill sea turtle, Eretmochelys imbricata, and their fitness consequences. Animal Behaviour, 70, 819-828. [ Links ]
Katilimis, Y., Urhan, R., Kaska, Y., & Baskale, E. (2006). Invertebrate infestation on eggs and hatchlings of the loggerhead turtle, Caretta caretta, in Dalaman, Turkey. Biodiversity and Conservation, 15, 3721-3730. [ Links ]
Lamont, M. M., & Houser, C. (2014). Spatial distribution of loggerhead turtle (Caretta caretta) emergences along a highly dynamic beach in the northern Gulf of Mexico. Journal of Experimental Marine Biology and Ecology, 453, 98-107. [ Links ]
Mrosovsky, N. (1983). Ecology and nest-site selection of leatherback turtles Dermochelys coriacea. Biological Conservation, 26, 47-56. [ Links ]
Mrosovsky, N. (2006). Distorting gene pools by conservation: Assessing the case of doomed turtle eggs. Environmental Management, 38, 523-31. [ Links ]
NOAA (Turtle Expert Working Group). (2007). An assessment of the leatherback turtle population in the Atlantic Ocean. NOAA Technical Memorandum NMFS-SEFSC-555. [ Links ]
Nordmoe, E. D., Sieg, A. E., Sotherland, P. R., Spotila, J. R., Paladino, F. V., & Reina, R. D. (2004). Nest site fidelity of leatherback turtles at Playa Grande, Costa Rica. Animal Behaviour, 68, 387-394. [ Links ]
Pike, D. (2008). The benefits of nest relocation extend far beyond recruitment: a rejoinder to Mrosovsky. Environmental Management, 41, 461-464. [ Links ]
Pritchard, P. (1982). Nesting of the leatherback turtle, Dermochelys coriacea in Pacific Mexico, with a new estimate of the world population status. Copeia, 1982, 741-747. [ Links ]
Reina, R. D., Mayor, P. A., Spotila, J. R., Piedra, R., & Paladino, F. V. (2002). Nesting ecology of the leatherback turtle, Dermochelys coriacea, at Parque Nacional Marino Las Baulas, Costa Rica, 1988-1989 to 1999-2000. Copeia, 2002, 653-664. [ Links ]
Runemark, A. (2006). Spatial distribution and temperature effects on hatching success of the leatherback turtle Dermochelys coriacea: implications for conservation. Master thesis. Uppsala University, Sweden. [ Links ]
Santidrián-Tomillo, P., Velez, E., Reina, R. D., Piedra, R., Paladino, F. V, Spotila, J. R., & Vélez, E. (2007). Reassessment of the leatherback turtle (Dermochelys coriacea) nesting population at Parque Nacional Marino Las Baulas, Costa Rica: effects of conservation efforts. Chelonian Conservation and Biology, 6, 54-62. [ Links ]
Sarmiento, R. (2007). Caracterización del material maderifero de deriva sobre un sector de la playa de Anidación de Tortugas Marinas, Tortuguero – Costa Rica. STC Internal Document. [ Links ]
Sarti Martinez, A. L. (2000). Dermochelys coriacea. In IUCN 2006. 2006 IUCN Red List of Threatened Species. Retrieved from www.iucnredlist.org
Links ]">
Spotila, J. R., Dunham, A. E., & Leslie, A. J. (1996). Worldwide population decline of Dermochelys coriacea: Are leatherback turtles going extinct? Chelonian Conservation and Biology, 2, 209-222. [ Links ]
Spotila, J. R., Reina, R., Steyermark, A., Plotkin, P. & Paladino, F. (2000). Pacific leatherback turtles face extinction. Nature, 405, 529-530. [ Links ]
Troëng, S., Chacón, D., & Dick, B. (2004). Possible decline in leatherback turtle Dermochelys coriacea nesting along the coast of Caribbean Central America. Oryx, 38, 395-403. [ Links ]
Wallace, B. P., DiMatteo, A. D., Bolten, A. B., Chaloupka, M. Y., Hutchinson, B. J., Abreu-Grobois, F. A., & Mast, R. B. (2011). Global conservation priorities for marine turtles. PloS One, 6, e24510. [ Links ]
Whitmore, C. P., & Dutton, P. H. (1985). Infertility, embryonic mortality and nest-site selection in leatherback and green sea turtles in Suriname. Biological Conservation, 34, 251-272. [ Links ]
Witherington, B., & Martin, R. (1996). Understanding, assessing, and resolving light-pollution problems on sea turtle nesting beaches. Florida Marine Research Institute Technical Report TR-2. [ Links ]
Zar, J. H. (2010). Biostatistical Analysis. USA: Prentice Hall. [ Links ]
1. Escuela de Biología, Universidad de Costa Rica, 11501-2060 San José, Costa Rica; noga.noga@gmail.com, mingowehrtmann@gmx.de, federico.bolanos@ucr.ac.cr
2. Current address: Department of Biology, Drexel University, 3245 Chestnut St, Philadelphia PA 19104, USA.
3. Sea Turtle Conservancy, Apartado Postal 246-2050, San José, Costa Rica; emma@conserveturtles.org
4. Unidad de Investigación Pesquera y Acuicultura (UNIP), Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Universidad de Costa Rica, 11501-2060 San José, Costa Rica.
Received 03-VI-2014. Corrected 10-I-2015. Accepted 05-II-2015.


{kind=link}
{kind=link}
{kind=link}