










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.2 San José Apr./Jun. 2015
Non-marine ostracodes from highland lakes in East-central Mexico
Ostrácodos no-marinos de lagos en el altiplano este-centro de México
Ostrácodos no-marinos de lagos en el altiplano este-centro de México
Abstract
The ostracode fauna of the neartic-neotropical transitional zone has remained poorly known until this study. Ten ostracode species inhabit ten highland lakes (five maar lakes (phreatic/phreato-magmatic explosion origin), one volcanic-tectonic lake, three natural dams and one man-made dam) in East-central Mexico. Surface sediments from the deepest part and the littoral zone from all studied lakes were collected. Environmental variables (pH, temperature, dissolved oxygen, conductivity, TDS) were measured in situ and parallel water samples for chemical analysis were collected for habitat description. Widely distributed species in the study area (≥5 lakes) include Cypridopsis vidua, Darwinula stevensoni and Eucandona cf. patzcuaro. Limnocytherina axalapasco is an endemic species and was collected in three maar lakes and in one man-made dam. Rare species included: Chlamydotheca arcuata?, Fabaeformiscandona acuminata?, Ilyocypris gibba?, Limnocythere friabilis?, Potamocypris smaragdina? and Potamocypris unicaudata?. Highest species richness (6 spp.) was found in the large and shallow Lake Metztitlán (2.6km2, 5.5m deep), with the lake water type HCO3->>SO42->Cl--- Ca2+>Na+>Mg2+. The rest of studied lakes (<63m, <27km2) had not more than three species. For instance, only two ostracode species were collected in Lake Alchichica, which is the largest, deepest and most saline studied maar lake. Rev. Biol. Trop. 63 (2): 401-425. Epub 2015 June 01.
Key words: freshwater ostracodes, maar lakes, natural dams, Central Mexico highlands, nearctic-neotropical transitional zone.
Resumen
La fauna de ostrácodos de la zona de transición néartica-neotropical ha sido poco estudiada hasta este estudio. En el este-centro de México, habitan diez especies de ostrácodos en diez lagos: cinco lagos maar (originados por explosiones freáticas/freato-magmáticas), un lago volcánico-tectónico, tres embalses naturales y un embalse artificial. De la parte más profunda y de las zonas litorales de todos los lagos estudiados se recolectaron sedimentos superficiales para el análisis de ostrácodos. También se midieron in situ las variables ambientales: pH, temperatura, oxígeno disuelto, conductividad, SDT, además se recolectaron muestras de agua paralelas para análisis químico con el objetivo de describir el hábitat. Especies con una amplia distribución en el área de estudio (≥5 lagos) incluyen: Cypridopsis vidua, Darwinula stevensoni y Eucandona cf. patzcuaro. Limnocytherina axalapasco es una especie endémica y fue recolectada en tres lagos maar y en el embalse artificial. Especies raras incluyeron: Chlamydotheca arcuata?, Fabaeformiscandona acuminata?, Ilyocypris gibba?, Limnocythere friabilis?, Potamocypris smaragdina? y Potamocypris unicaudata? La riqueza de especies más alta (6 spp.) se encontró en el lago más extenso y somero, lago Metztitlán (2.6km2, 5.5m de profundidad), con el tipo de agua HCO3->>SO42->Cl--- Ca2+>Na+>Mg2+. El resto de lagos estudiados (<63m, <27km2) presentaron no más de tres especies. Por ejemplo, solamente dos especies de ostrácodos fueron recolectadas en el lago Alchichica, el cual es el más extenso, profundo y salino entre los lagos maar estudiados.
Palabras clave: ostrácodos dulceacuícolas, lagos maar, embalses, altiplano del centro México, zona de transición neártica-neotropical.
Microcrustaceans (ostracodes, cladocerans and copepods) are dominant zooplankters in aquatic ecosystems, and are key components of the food web (Dole-Olivier, Galassi, Marmonier, & Creuzé des Chattelliers, 2000; Cohen, 2003). Non-marine ostracodes (Crustacea: Ostracoda) are typically <3mm long and are commonly called “mussel-shrimps”, because their soft parts are protected by two low-Mg calcite shells that form a carapace (Meisch, 2000). Non-marine ostracodes inhabit a variety of aquatic environments such as lakes, lagoons, ponds, rivers and sinkholes (Pérez et al., 2011). Ostracodes are highly sensitive to climate and environmental fluctuations, which makes them great paleobioindicators in paleolimnological studies (Cohen, 2003). Cladocerans and ostracodes are the most important groups of crustaceans in paleolimnological studies because their body segments preserve well in lacustrine sediments. Once their modern ecological requirements are known, ostracodes can be used to quantitatively reconstruct past environmental variables such as conductivity (Mischke et al., 2010), temperature, water chemical composition (Mezquita, Roca, Reed, & Wansard, 2005) and water depth (Pérez et al., 2010a, 2011). This is only possible if species are well identified, and by establishing large local training sets (calibration data), that allows a good knowledge of the species ecological preferences (Viehberg, & Mesquita-Joanes, 2012). The performance of transfer functions based on the previous data depends as well on how reliable and consistent the species identification is. Therefore, a reliable taxonomy is a prerequisite for studies of modern and past environments (Pérez, 2010).
Central Mexico is an interesting area for ostracode studies (species richness, diversity, distribution and ecology) because it is a transitional zone between the Nearctic and the Neotropic. Besides, this region is a sensitive zone between temperate and tropical climatic regimes (Davies, Metcalfe, Caballero, & Juggins, 2002). This study reveals whether ostracode species living in these highland lakes have a Nearctic and/or Neotropical distribution and if endemic species exist. First studies on ostracodes in Central Mexico were carried out by Rio de la Loza and Craveri (1858), de Saussurre (1858), Ehrenberg (1869), in Northern Guatemala by Brehm (1932) and in the Yucatán Peninsula by Furtos (1936). The first studies of ostracodes in the Guatemalan highlands (crater lakes Ayarza, Amatitlán and Atitlán) were carried out by Pérez, Lorenschat, Brenner, Scharf, and Schwalb (2012) and Lorenschat (2009). These authors reported two unknown species of Limnocythere and Ilyocypris, respectively, which could be potential new species, and reported the lack of studies and literature regarding ostracodes from crater lakes in Mexico and Central America.
The extant ostracode fauna in lakes along the Trans-mexican Volcanic Belt (TMVB) and in any freshwater ecosystems in Central Mexico has been poorly studied. There are few paleolimnological studies that have reported fossil ostracodes in late Quaternary sediments (Caballero, Vilaclara, Rodríguez, & Juárez, 2003; Juárez, 2005), however the modern fauna still remains largely unstudied. Caballero et al. (2013) reported the modern ostracode fauna from crater lake Santa María del Oro in Western Mexico, and Hernández, Escobar, and Alcocer (2010) studied the benthic crustacean assemblage of Lake Alchichica. Recently, Cohuo, Pérez, and Karanovic (2014) reported a new endemic ostracode species (Limnocytherina axalapasco) for Mexican crater lakes. In contrast, the microcrustaceans of the Yucatán Peninsula and surrounding areas (Guatemala and Belize) are well known (Lorenschat, 2009; Pérez et al., 2012, 2013). Because of their great importance as proxies of environmental and climatic change, ostracodes in Central Mexico have been used in paleolimnological studies, mostly in the Pátzcuaro Basin (Forester, 1985; Bridgwater, Heaton, & O’Hara, 1999a; Bridgwater, Holmes, & O’Hara, 1999b; Metcalfe et al., 2007), and in the crater lake Santa María del Oro, Western Mexico (Vázquez, Ortega, Rodriguez, Caballero, & Lozano, 2008). However, taxonomic and ecological studies are still missing for these lakes, which could improve their paleolimnological inferences.
The main objective of this study was to provide a short morphological and habitat description, and a compilation of the ecological preferences and distribution for the ostracode fauna that inhabit lakes in East-central Mexico. This is a region where paleolimnological studies have been conducted, and this study will improve environmental and climatic inferences based on ostracodes.
Materials and methods
Study site: This study covers the area from 19°05’ to 20°48’ N and from 97°21’ to 99°18’ W, which includes mainly the Eastern part of the TMVB (Fig. 1). Most studied waterbodies are located in the Oriental basin, while Lake Tecocomulco is part of the basin of Mexico. Lakes Atezca, and Metztitlán are located in the Sierra Madre Oriental, and Lagunas de Zempoala in the Sierra de Chichinautzin. The TMVB crosses the country in a E-W orientation between 19-20° N, revealing a complex subduction geometry (Armienta et al., 2008). Most of the terrain in this region is mountainous (>2 000masl, average temperature is 15°C), as a primarily product of Quaternary volcanism. Abundant aquatic ecosystems of tectonic and volcanic origin are found within the TMVB (Davies et al., 2002; García-Rodríguez, & Tavera, 2002), and lakes dammed by lava flows, or by large scale rock avalanches. Numerous dams (man-made and natural) are found in the region as well (Conserva, & Byrne, 2002; Suter, 2004; de la Lanza-Espino, Gómez-Rodríguez, Islas Islas, Escalante Richards, & Hernández Pulido, 2011). Oriental is a closed hydrological basin located at the Eastern end of the Mexican highlands (mean altitude of 2 300masl) (Caballero, Lozano-García, Vázquez-Selem, & Ortega, 2010). Six maar lakes (phreatic/phreato-magmatic explosion origin) characterize the Eastern TMVB, locally known as the Axalapascos (Armienta et al., 2008). Lakes in the study area display a broad lake maximum depth range, from shallow (Tecuitlapa, 4.5m) to deep lakes (Alchichica, 63m) (Armienta et al., 2008). The trophic state of the lakes range from oligotrophic (Atexcac) to hypereutrophic (Tecuitlapa). The dry climate in the region (425 to 656mm mean annual precipitation) is a product of the orographic barrier of the Sierra Madre Oriental and the Cofre de Perote-Citlatépetl volcanic chain (Caballero et al., 2010). Maar lakes in the Axalapascos region are of special interest for limnologists and biologists (Ramírez-García, & Vázquez-Gutiérrez, 1989; Alcocer, & Bernal-Brooks, 2010), because they have a high level of endemism (Barluenga, Stölting, Salzburger, Muschick, & Meyer, 2006; Alcántara-Rodríguez, Ciros-Pérez, Ortega-Mayagoitia, Serrania-Soto, & Piedra-Ibarra, 2012). These lakes are also important, because of their potential use as high resolution paleoclimatic records (Caballero et al., 2003; Juárez, 2005). The most studied crater lake of Mexico is Lake Alchichica in the Axalapascos (Alcocer, & Lugo, 2003; Alcocer, & Escobar-Briones, 2007; Alcocer, Arce, Zambrano, & Chiappa-Carrara, 2010). Main topics that have been studied include general limnology, endemism, species adaptation, and biogeography (Alcocer, Escobar, Lugo, & Peralta, 1998; Barluenga et al., 2006; Oliva, Lugo, Alcocer, & Cantoral-Uriza, 2006; Alcocer et al., 2010; Kazmierczak et al., 2011; Alcántara-Rodríguez et al., 2012; Cohuo et al., 2014). The Basin of Mexico hosts various relict lacustrine sub-basins such as Xochimilco and Chalco, Texcoco, Zumpango, Xaltocan, Tochac and Tecocomulco (Roy, Caballero, Lozano, Pi, & Morton, 2009). Our study includes Lake Tecocomulco, where already paleolimnological studies (Caballero, Lozano, Ortega, Urrutia, & Macías, 1999; Roy et al., 2009) have been carried out. Lake Tecocomulco is located in the North-Eastern plains, at an altitude of 2 450masl (Roy, Caballero, Lozano, & Smykatz-Kloss, 2008) and with an average annual precipitation of 650mm (Roy et al., 2009).
Sampling and habitat description: Ostracode samples were collected from ten lakes (five maar lakes, one volcanic-tectonic lake, three natural dams, and one man-made dam) during June 2011 (Fig. 1). Surface sediments were retrieved using an Ekman Grab at the lake’s deepest point (Zempoala: 6m, Atezca: 14.4m, Tecocomulco: 0.9m, La Preciosa: 23.5m, Alchichica: 45m, Tecuitlapa: 1.5m, Aljojuca: 40m, Atlagantepec: 1.2m, Quechulac: 32m, Metztitlán: 3m) and at selected littoral zones (0.5m). A total of two samples (deepest point and littoral) were collected at each lake. All samples were immediately preserved in 96% ethanol and stored in 100mL PE-bottles. At each sampling site, water temperature, pH, dissolved oxygen (DO), and electric conductivity were measured in situ with a multiparametric sonde (Hydrolab Quanta), and parallel surface water samples for major ion composition (CO32-, HCO3-, Cl-, SO42-, Na+, K+, Mg2+, Ca2+) were collected, and then refrigerated (4°C) until analyses in the laboratory. Sampling and analyses in the laboratory were made according to Armienta et al. (2008), which are described in American Public Health Association [APHA] (1995, 2005). These analyses were carried out in the Laboratory of Analytical Chemistry, Institute of Geophysics, Universidad Nacional Autónoma de México (UNAM), Mexico.
Ostracode analysis: Ostracodes were extracted from 10mL of all sediment samples using fine brushes under an Olympus SZ stereoscope. Ostracodes with well-preserved soft parts were kept at 5°C in small eppendorf vials filled with 96% ethanol. Hard parts were stored in micropaleontological slides. For an accurate identification, well-preserved specimens were dissected following Meisch (2000). When well-preserved soft parts from adult ostracodes were not collected, we added a question mark after the species name, because identification was only based on their shell morphology and accurate identifications should include observation of both hard and soft parts. Generally it is difficult to collect well-preserved soft parts in samplings based on a limited number of surface sediment samples per lake, which is the case of calibration data sets. In this paper, we present a short description of the ostracode hard parts. Length and height were measured by taking light microscope pictures from valves and then measuring them by using the software Axio Vision Release 4.6.3. Scanning electron microscope (SEM) pictures were taken using a Jeol JSM-5600LV LCM scanning microscope of the Central Microscopy Laboratory, Institute of Physics, UNAM, Mexico. Ostracode specimens are being stored temporarily at the Department of Paleontology, Institute of Geology, UNAM, Mexico.
Results
Highland lakes of East-Central Mexico: Sampled highland lakes include five maar lakes, one volcanic-tectonic lake, three natural dams and one man-made dam (Table 1). The sampled altitudinal gradient ranges from 1 258 to 2 804masl. The shallowest lake is Tecocomulco (0.9m) and the deepest Alchichica (64.6m). The mean lake depth in the region is ~5.2m. Most sampled lakes display small surface areas (≤1.8km2), and the largest lakes are Tecocomulco and Metztitlán with 17.7 and 29.0km2, respectively. Surface water temperatures ranged from 18.7 to 30.0°C, the pH from 7.7 to 10.3, the DO from 4.6 to 7.9mg/L, conductivity from 93 to 3 710µS/cm, except for Lake Alchichica that displays conductivities as much as 12 940µS/cm. The TDS concentration ranges from 0.1 to 9.0g/L.
Studied maar lakes in the Oriental basin, from 2 321-2 371masl, are small (≤1.8km2), and deep (≤64.9m), and are characterized by temperatures from 18.7 to 21.9°C, with the exception of lake Tecuitlapa, which is shallow (2.5m) and displays a much higher surface water temperature (26.2°C). The pH from maar lakes fluctuated from 8.8 to 10.3 and the DO concentration ranged from 4.6 to 6.5mg/L. Electric conductivity and TDS concentration vary (756 to 12 940µS/cm and 0.5 to 9g/L, respectively). Natural dams, from 1 258 to 2 804masl, are shallower than most studied maar lakes (≤16m), and their surface area ranges from 0.1 to 29.0km2. The largest dams is Metztitlán. The only studied man-made dam, lake Atlangatepec, (2 511masl) is small (1.2km2) and shallow (1.2m). Lake Tecocomulco with a volcanic-tectonic origin, is the second highest (2 535masl) and largest studied lake (17.7km2). The natural dams, Lakes Atezca and Metztitlán displayed higher surface temperatures than maar lakes, 26.9°C and 30°C, respectively. The lowest pH (7.7) among all the studied lakes, was determined in the man-made dam Atlangatepec. Electric conductivity (≤505µS/cm) and TDS concentrations (≤0.3g/L) of surface waters from the dams were lower than those from the maar lakes.
The water chemical composition of the lakes is presented in Table 2. The dominant anions in all lakes were carbonates and bicarbonates, except for Lake Alchichica, where chloride dominated. Carbonate and bicarbonate concentrations were higher in lakes Alchichica and Tecuitlapa. Chloride concentrations were relatively high in the maar lakes La Preciosa and Quechulac. Sulfate concentrations were generally low except in Lake Alchichica. The dominant cations were magnesium in La Preciosa and Quechulac, calcium in Metztitlán, Atezca, and Zempoala and sodium-potassium in the rest of the lakes. Among all studied lakes, maar lake Alchichica displayed the highest total ionic concentration (336.25meq/L), and the highest ion concentrations, except for bicarbonate and calcium. Bicarbonate concentration was highest in the maar lake Tecuitlapa and calcium concentration was highest in the dam Metztitlán. Lake Zempoala has the lowest total ionic concentration (2.10meq/L) among studied lakes.
Freshwater ostracode fauna: A total of ten species (Table 3, Fig. 2) were collected in the studied lakes in the highlands of East-central Mexico (Tables 1, 2). Ostracode classification is shown in Table 3. Seven ostracode species belong to the infraorder Cypridocopina, two species to Cytherocopina and one species to Darwinulocopina. The species assemblage of the studied lakes was composed by five cyprids, two candonids, two limnocytherids, and one darwinulid. Table 4 shows the distribution of ostracodes in East-Central Mexico. The most widely distributed species (≤5 lakes) were Cypridopsis vidua, Darwinula stevensoni and Eucandona cf. patzcuaro followed by Limnocytherina axalapasco (four lakes). Rare species (≤4 lakes) included Chlamydotheca arcuata?, Fabaeformiscandona acuminata?, Ilyocypris gibba?, Limnocythere friabilis?, Potamocypris smaragdina? and Potamocypris unicaudata?. Lake Metztitlán displayed the highest species richness (n=6), while the rest of lakes, including the deep Lake Alchichica, presented less than three species. Because of the little information on the extant non-marine ostracode fauna in Mexico, especially in waterbodies of Central Mexico, a short description of the species morphology (RV: right valve; LV left valve) with shell measurements (L: length, H: height) of collected valves (Table 5), as well as their local, regional and worldwide distribution, and ecological preferences are presented below in species alphabetical order:
Chlamydotheca arcuata? (Sars, 1901)
(Fig. 2A, B)
Identification: Furtos (1933), Smith and Delorme (2010).
Size: Adults: L: 1.80-6.00mm, H: 0.99 to 1.80mm. The LV is slightly larger and higher than the RV (Table 5). Juvenile valves collected in this study displayed the following measures: L: 0.60-1.28mm, 0.38-0.75mm (RV, n=3); L: 0.89-1.85mm, H: 0.55-1.09mm (LV, n=2). One collected juvenile valve in this study has the size of an adult ostracode (reported size, see Table 5), however the poor-developed lamellae suggests that it is still an instar.
Morphology: We collected only juvenile single valve. A-2, A-1 instars: Valve surface smooth with delicate setae. Valves large, elongated, oval in lateral view. Anterior and posterior margins round. Ventral margin slightly convexly rounded. Greatest H in front of the middle part of the valve. Posterior margin of RV clearly serrated (ca. 19 short spines). Adults: For a detailed description of adult morphology see Furtos (1933), Smith and Delorme (2010), Díaz and Lopretto (2011) and Rodríguez (2011).
Distribution: Lake Tecocomulco, East-Central Mexico. It has been reported in waterbodies from Nayarit (Caballero et al., 2013), Durango (Rodríguez, 2011; Cohuo, 2012), Quintana Roo (Cohuo, 2012), and Tlaxcala, Mexico (Saldivar-López, 2011). Outside Mexico, it has been collected in Brazil (1901), Paraguay (Daday, 1905), Argentina (Díaz, & Lopretto, 2011), USA (Furtos, 1933; Tressler, 1949; Smith, & Delorme, 2010), and Canada (Delorme, 1970).
Ecological preferences: Nektobenthic. Juvenile specimens were collected at 21.2°C, pH 8.8, DO 5.9mg/L, conductivity 341µS/cm and TDS 0.2g/L. The dominant water ions were Na+, Ca2+, Mg2+, HCO3- and Cl- (Table 2). Juveniles were found in black fine sediment, with high organic matter content and rests of vegetation and aquatic plants. It is a typical tropical species that lives in waters ranging from 24 to 39°C (Smith, & Delorme, 2010; Soria-Caballero, 2010). It has been collected in streams, temporary ponds (Díaz, & Lopretto, 2011), warm springs (Smith, & Delorme, 2010; Soria-Caballero, 2010) and wells (Zara-environmental-LCC, 2010). It shows a preference for slow water currents and it has been collected in waters dominated by Ca2+, Mg2+, K+, Na2+, SO42-, CO32-, displaying a salinity range from 600 to 8 000ppm (Soria-Caballero, 2010).
Cypridopsis vidua (O.F. Müller, 1776)
(Fig. 2C)
Identification: Meisch (2000).
Size: Adults: L: 0.4-0.7mm, H: 0.32-0.43mm. The LV is slightly longer and higher than the RV (Table 5). Valves are usually 0.5 to 0.6mm long (Meisch, 2000).
Morphology: Valves pitted and heart shaped. Ovate to subovate in dorsal view. Dorsal margin arched, and the highest carapace width in the middle. Carapaces usually with four distinct dark green transverse stripes (this study), but it can be found in shades of brown and purple (Smith, & Delorme, 2010). This species is cosmopolitan and therefore presents instraspecific variability in size, shape, surface ornamentation and color (Meisch, 2000).
Distribution: Lakes Zempoala, Atezca, Metztitlán, Tecocomulco, and Aljojuca, East-Central México. It has been reported in Nayarit (Caballero et al., 2013), Michoacán (Bridgwater et al., 1999a), Chihuahua (Palacios-Fest, Carreño, Ortega-Ramírez, & Alvarado-Valdéz, 2002; Chávez, 2011), Sonora (Palacios-Fest, & Dettman, 2001), Mexico. Cohuo (2012) collected this species in waterbodies from Central Mexico to the Yucatán Peninsula. Outside Mexico it has been collected in Guatemala (Pérez et al., 2011; Pérez et al., 2012), USA (Smith, 1993; Smith, & Delorme, 2010), Europe and Japan (Meisch, 2000).
Ecological preferences: Nektobenthic and a plant-dwelling ostracode. Specimens were collected at 19.3-30.0°C, pH 8.8-9.5, DO 4.6-7.9mg/L, conductivity 93-1 152µS/cm, and TDS 0.1-0.7g/L. The dominant water ions were Ca2+, Mg2+, Na+ and HCO3-, SO42-, Cl- (Table 2). This species prefers waters with conductivities ranging from 210 to 1 350mS/cm, waters dominated by Ca2+, Mg2+, Na+ and HCO3-, SO42- (Smith, 1993) and displaying temperatures from 20.2 to 27.6°C (Keyser, 1976; Lorenschat, 2009). Living organisms have been collected at 44m water depth, but it seems to prefer littoral zones (Lorenschat, 2009). It inhabits springs, wetlands, streams and interstitial habitats. This species actively searchs for Chara bed for food on the periphyton and for protection against predators (Karanovic, 2012). Studies have shown that C. vidua is sensitive to herbicides, and it can survive for several hours frozen (Smith, & Delorme, 2010). Preferred sediment is fine to sandy (Lorenschat, 2009).
Darwinula stevensoni
(Brady and Robertson, 1870)
(Fig. 2D)
Identification: Meisch (2000), Pérez, Lorenschat, Brenner, Scharf, and Schwalb (2010b).
Size: Adult females: L: 0.59-0.80mm, H: 0.22-0.30mm. The RV is slightly longer and higher than the LV (Table 5).
Morphology: Valves smooth, whitish, elongated. Posterior margin rounder and broader than anterior. Dorsal margin convex, ventral margin slightly concave and almost straight. RV larger than LV and overlaps LV ventrally. Muscle scars arranged in a rosette and located in front of mid-length in adult carapaces.
Distribution: Lakes Atezca, Metztitlán, Atlagantepec, Quechulac and Tecuitlapa (this study) and Lake La Preciosa (Juárez, 2005), East-central Mexico. It has been reported in Nayarit (Caballero et al., 2013), Michoacán (Bridgwater et al., 1999a), Nuevo León, Tamaulipas (Rodríguez, 2002), Morelos (Almeida Leñero, 1973), Estado de México (Carreño, 1990) and the Yucatán Peninsula, Mexico (Furtos, 1936; Gabriel et al., 2009; Pérez et al., 2011, 2012). Outside Mexico it has been collected in Belize, Guatemala (Pérez et al., 2010b), Nicaragua (Hartmann, 1959), USA (Furtos, 1933; Smith, & Delorme, 2010), among others. It displays a world-wide distribution (Meisch, 2000).
Ecological preferences: Benthic. It was collected at 19.6-30.0°C, pH 7.7-10.3, DO 5.0-7.9mg/L, conductivity 105-3 710µS/cm and TDS 0.1-2.4g/L. The dominant water ions were Ca2+, Mg2+, Na+ and HCO3-, SO42-, Cl- (Table 2). This species displays a preference for muddy and sandy substrates. It prefers low temperatures but it can be found at temperature ranging between 10° and 35°C. It has been collected in waters with a pH as low as 6 (Deckker, 1981; Külköylüoglu, & Vinyard, 2000). Smith (1993) reports a conductivity range in North-central United States for this species from 210 to 925mS/cm. This species shows a preference for water with low salinity, however it can tolerate up to 15‰ (Meisch, 2000; Pérez et al., 2010b), high DO but tolerates waters with a DO between 2 and 14mg/L (Külköylüoglu, & Vinyard, 2000; Smith, & Delorme, 2010). It lives in ponds, lakes, “cenotes”, coastal lagoons, rivers, slow streams, interstitial groundwater (Meisch, 2000; Pérez et al., 2011). The maximum depth where this species has been collected is at 15m in Lakes Petén Itzá and Izabal, Guatemala (Pérez et al. 2012)
Eucandona cf. patzcuaro (Tressler, 1954)
(Figs. 2E, F, G)
Identification: Tressler (1954), Karanovic (2012).
Size: Adult females: L: 0.99-1.34mm, H: 0.55-0.73mm. Adult males: L: 1.12-1.43mm, H: 0.66-0.81mm (Table 5).
Morphology: Valves whitish, smooth, pitted, and sparsely hairy, elongated and kidney-like. Carapace anteriorly round, posteriorly subrounded, bottom concave, greatest height in the posterior half (Fig. 2E, Fig. 2F, Fig. 2G). Valves ornamentated. Five adductor muscles centered in oval (three front, two back). Males display an indentation or notch at the anterodorsal margin and a sharp angle at the anteroventral margin. A small protuberance in the anteroventral margin is characteristic of males. Dorsal margin round, ventral margin posteriorly indented to the anteroventral angle. Females lack the small protuberance on the anteroventral margin. The front of the valve is round and the posterior semiround. A distinct indentation characterizes the back dorsalmargin. The similarity (hard and soft parts) of collected specimens in this study and in Lake Patzcuaro (Tressler, 1954) is high, however, we suspect that it could be a new species restricted to East-central Mexico (see discussion) and therefore decided to report this species as Eucandona cf. patzcuaro. Karanovic (2012) suggests Eucandona patzcuaro (previously Candona patzcuaro).
Distribution: Lakes Metztitlán, Atlagantepec, Alchichica, La Preciosa and Quechulac, East-central Mexico. Eucandona patzcuaro has been reported in Nayarit (Caballero et al., 2013), Puebla (Juárez, 2005; Hernández et al., 2010), Michoacán (Tressler, 1954; Bridgwater et al., 1999b; Garduño-Monroy et al., 2011), Chihuahua (Palacios-Fest et al., 2002), Mexico. Maddocks, Machain-Castillo, and Gío-Argáez (2009) indicate that it is widely distributed in the Gulf coast of Mexico. Outside Mexico it has been collected in USA (Forester, Smith, Palmer, & Curry, 2013).
Ecological preferences: Benthic. Specimens were collected at temperatures 18.7-30°C, pH 7.7- 9.3, DO 5.0-7.0mg/L, conductivity 292-12 940µS/cm, and TDS 0.2-9 g/L. The dominant ions in lake waters were Ca2+, Mg2+, Na+ and HCO3-, SO42-, Cl- (Table 2). Living specimens were collected in a depth of 63m in Lake Alchichica. It was found in different types of substrates: sandy and fine sediment (clay and silts) mixed with small pebbles. Sediments displayed sometimes detritus and small sediment agglomerates. Eucandona patzcuaro shows a preference for alkaline waters (Tressler, 1954), tolerates a wide range of salinity concentrations (200-5 000mg/L), however it prefers low values (Palacios Fest, Cohen, Ruiz, & Blank, 1993; Palacios-Fest, 2010). It has been collected in a wide temperature, 2 to 32°C (Palacios-Fest, 2010). Its presence indicates permanent, slow flow to still waters (Palacios-Fest, 2004).
Fabaeformiscandona acuminata?
(Fischer, 1851)
(Fig. 2H, I)
Identification: Meisch (2000).
Size: Adults: L: 1.12 to 1.60mm. Females: L: 1.12-1.13mm, H: 0.52mm (Table 5). Juveniles: RV (n=2): L: 0.61 to 0.78mm, H: 0.26 to 0.33mm.
Morphology: Adults: see Karanovic (2012). Juveniles: Valve smooth, pitted, with a distinct ornamentation. Valve with a trapezoid shape, anterior margin round, posterior margin elongated. Ventral margin convex. Dorsal margin almost straight.
Distribution: Lake Aljojuca, East-central Mexico. Outside Mexico it has been collected in USA (Forester et al., 2013), Belarus (Nagorskaya, 2002; Nagorskaya, & Keyser, 2005), Europe (Marmonier, & Chatelliers, 1992; Horne, 2007; Szlauer-Lukaszewska, 2012) and in the Arctic (Wetterich, 2008).
Ecological preferences: Benthic. Specimens were found at temperature 21.9°C, pH 9.5, DO 4.6mg/L, conductivity 1 152µS/cm, and TDS 0.7g/L. It was collected in a lake with the following water type HCO3->>Cl->SO42- -- Na2+>Mg2+>>Ca2+. Wetterich (2008) suggests that this species is oligohalophilic. This species has been collected in oxbow lakes, ponds and rivers and shows a preference for sandy and silty substrates (Nagorskaya, & Keyser, 2005). It has been collected among Phragmites australis rushes (Szlauer-Lukaszewska, 2012).
Ilyocypris gibba? (Ramdhor, 1808)
(Fig. 2J, K, L)
Identification: Meisch (2000).
Size: Adults: L: 0.70 to 1.05mm, L: 0.43-0.45mm. Females are longer than males (Table 5). Juveniles: RV (n=2): L: 0.51 to 0.55mm, H: 0.28 to 0.30mm. LV (n=1): L: 0.61mm, H: 0.36mm.
Morphology: Valves subrectangular, covered with small pits, with two transverse sulci. Dorsal margin almost straight and slightly arched over the sulci. Anterior sulcus longer than posterior. Valves display lateral nodes. Nodes in the upper part of the shell are bigger. Anterior and posterior margins of RV and LV present spinules. Front and back part of the shell rounded. Greatest height in the first half of the shell. Only valves were collected, therefore we do not compare between males and females.
Distribution: Lake Metztitlán, East-Central Mexico. It has been reported in Veracruz (Cohuo, 2012), Nuevo León and Tamaulipas (Rodríguez, 2002), Mexico. Outside Mexico it has been collected in Guatemala (Lorenschat, 2009; Pérez et al., 2012), North and South America, Europe, Africa, Middle East, Asia, China (Meisch, 2000; Lorenschat, 2009).
Ecological preferences: Benthic. It was collected at temperature 30°C, pH 8.9, DO 5.8mg/L, conductivity 505µS/cm, and TDS 0.3g/L. The dominant ions in lake waters were Ca2+, Na+, Mg2+, HCO3-, SO42-, and Cl- (Table 2). Klie (1938) collected this species in water not colder than 10°C. However, Smith and Delorme (2010) report that it tolerates waters down to 5°C. Shows a preference for waters between 5 and 15°C. It was collected in waters of Lake Atitlán, Guatemala, at temperature 20.2-21.8°C, salinity 0.1‰, and pH 8.2-8.4. Külköylüoglu (2004) reports environmental variable ranges for this species: pH 6.64-9.80, 7.50-42.0°C, DO 3.0-14.0mg/L, 260-2 800mS/cm (Lorenschat, 2009). It prefers running water but it is also found in lakes and ponds. Preferred substrate is clayey, fine-mudded or sandy (Lorenschat, 2009). It can also be found in temporary pools, springs, slightly salty waters and rice fields and some specimen have been reported from the interstitial habitat (Meisch, 2000; Lorenschat, 2009). It inhabits shallow waters. Lorenschat (2009) collected specimens from the littoral zone (0.5m) down to 44m depth of Lake Atitlán, Guatemala. Ilyocypris gibba requires a minimum of 3mg/L of DO concentration. It has been collected in special biotopes, such as the delta area of rivers in North America (Smith & Delorme, 2010). Ilyocypris gibba has been found abundantly in disturbed aquatic ecosystems (Külköylüoglu, 2004).
Limnocytherina axalapasco
(Cohuo et al., 2014)
(Fig. 2M, N)
Identification: Karanovic (2012), Martens (1996, 2000)
Size: Adult females are smaller than males, L: 0.59-0.71mm, H: 0.27-0.37mm. Adult males: L: 0.62 to 0.75mm, H: 0.30 to 0.49mm (Table 5).
Morphology: Valves with sexual dimorphism. Surface pitted and covered with setae. Four muscle scars arranged vertically. LV overlaps RV. Marginal pore canal unbranched. Males: Anterior part of valves subrectangular. Posterior part rounder than in females. Anterior margin round, with abundant short setae. Posterior margin broad, round and covered with short setae. Ventral margin anteriorly straight, and slightly concave in the middle part of the shell. Posterior ventral margin round and widened. Females: Posterior dorsal margin slightly downward projected, dorsal margin straight. Middle part of the ventral margin slightly concave, a poor-developed flange overlaps it. Male and female can present on the middle part of their valves bumps that sometimes can turn into prominent lateral allae.
Distribution: Lakes Atlagantepec, Alchichica, Quechulac and la Preciosa, East-Central Mexico. It has been reported in Puebla (Juárez, 2005; Hernández et al., 2010; Cohuo et al., 2014). Endemic species, there are no records elsewhere in Mexico and in other countries.
Ecological preferences: Benthic. It was collected at temperatures 19.1-20.3°C, DO 5.0-7.0mg/L, pH 7.7-9.3, conductivity 292-12 950µS/cm, and TDS 0.2-9g/L. The dominant ions in lake waters were Na+, Mg2+, Ca+2, Cl-, HCO32-, and SO42- (Table 2). This species lives in oligotrophic lakes and tolerates high salinities. Ca and Mg seem to be the most important cations for the development of this species and DO seems to be the limiting variable for this species. It is a typical littoral species and its abundance decreases with increasing depth (Hernández et al., 2010). However, Cohuo et al. (2014) reports this species down to 64m. They found this species in alkaline waters dominated mostly by Na, Mg, Cl and HCO3, as well as in waters displaying temperatures 19.1-20.3°C, DO 5.0-6.5mg/L. The preferred substrate seems to be sands with low percentage of silts.
Limnocythere friabilis?
(Benson & MacDonald, 1963)
(Fig. 2O, P)
Identification: Benson and MacDonald (1963), Delorme (1971).
Size: Adults: L: 0.44 to 0.52mm, H: 0.22-0.30mm. Females are smaller than males (Table 5). Valves smaller than other Limnocythere species reported for the region.
Morphology: Valves pitted, finely reticulated. Sexual dimorphism. Greatest carapace width near the middle. Left valve overlaps right valve posteriorly. Anterior and posterior margin bearing small spines. Four muscle scars vertically arranged. Ventral margin concave. Radial pore canals simple and straight. Females: Valves small and subreniform. Greatest height in the anterior half of the valve. Anterior margin broadly rounded, posterior margin narrower than anterior. Dorsal margin slightly convex. Males: Valves subovate-subelliptical, longer, and not as high as female valves. Anterior margin round and posterior margin subrectangular and elongated. Dorsal margin almost straight.
Distribution: Lake Metztitlán, East-Central Mexico. It has been reported in the Gulf of Mexico (Machain-Castillo, & Gío-Argáez, 2004; Maddocks et al., 2009). Outside Mexico it has been collected in the Nearctic climatic zone, including the Gulf of Alaska (Brouwers, 1990) and lakes in the USA (Benson, & MacDonald, 1963; Staplin, 1963; Forester, Colman, Reynolds, & Keigwin, 1994; Forester, & Smith, 1994; Forester et al., 1999; Dennison-Budak, 2010).
Ecological preferences: Benthic. This species was collected at temperature 30°C, pH 8.9, DO 5.8mg/L, conductivity 505µS/cm, and TDS 0.3g/L. The dominant ions in lake waters were Ca2+, Na+, Mg2+, HCO3-, SO42- and Cl- (Table 5). We collected this species at a maximum water depth of 5.5m. This species inhabits freshwater lakes, however it can be found in low-energy river systems (Brouwers, 1990), and it has not been found in wetlands or springs (Smith, & Delorme, 2010). It prefers cold temperatures, and it has been found in the profundal zone of the modern Great Lakes at water depths ranging from 15 to 45m (Curry, & Yansa, 2004). However, Benson and MacDonald (1963) report that this species shows preference for shallower waters. Forester et al. (1994) uses the increase in the abundance of this species as an indicator or shore-line proximity. Curry and Yansa (2004) report that the presence of abundant specimens in relatively shallow zones of large lakes could suggest that the species is thrived in lakes that discharge meltwater or cold groundwater. Light isotopic δ18O values (-18‰) were measured by Dennison-Budak (2010) in ostracodes valvas of L. friabilis from the Glenns Ferry Formation, indicating that this species is associated with groundwater discharge. Dennison-Budak (2010) suggests that the presence of this species indicates oligohaline environments. Machain-Castillo and Gío-Argáez (2004) collected this species in river mouths along the Gulf of Mexico during the rainy season suggesting that this species slightly tolerates slightly saline waters.
Potamocypris smaragdina? Vavra, 1891
(Fig. 2Q)
Identification: Furtos (1933), Meisch (2000).
Size: Adults: L: 0.54 to 0.85mm, H: 0.33 to 0.47mm (Table 5). Meisch (2000) pointed out that the females and males studied from North America are smaller (L: 0.54-0.60mm).
Morphology: Valves smooth, covered with short, strong and backwardly directed hairs. RV shorter than LV. Pore canals found over valves, more concentrated in the ventral half. Carapace seen from the side elongated and subtriangular. RV: Dorsal margin boldly arched. The anterior margin more broadly rounded than the posterior margin. Posterior margin narrow. The greatest height in the middle. LV: Broadly rounded anterior margin and a truncate posterior margin. The posteroventral corner displays a distinct acute angle. Greatest height of LV is in front of the middle. LV encloses the RV anteriorly. Ventral margins of RV and LV slightly sinuated in the central region.
Distribution: Lakes Aljojuca and Metztitlán, East-Central Mexico. It has been reported in Michoacán (Bridgwater et al., 1999a; Garduño-Monroy et al., 2011). Outside Mexico it has been collected in the USA, Canada (Curry, 1999; Meisch, 2000), Argentina (Cusminsky, Pérez, Schwalb, & Whatley, 2005), Russia, Europe (Ferguson, 1958; Meisch, 2000), and China (Li, Liu, Zhang, & Sun, 2010).
Ecological preferences: Nektobenthic. This species was collected at temperature 21.9-30°C, pH 8.9-9.5, DO 4.6-5.8mg/L, conductivity 505-1 152µS/cm, and TDS 0.3-0.7g/L. The dominant ions in lake waters were Ca2+, Na+, Mg2+, CO32-, HCO3-, SO42-, and Cl- (Table 2). This species inhabits environmental conditions in the United States displaying an average mean annual temperature of 10.6°C, mean annual precipitation of 905mm, TDS of 518mg/L, and a HCO3-/Ca2+ of 1.7 (Curry, 1999). Potamocypris smaragdina has a preference for shallow waters, warm and inhabits environments with low effective moisture (Curry, & Baker, 2000), and it displays a strong preference for the presence of aquatic plants (Bridgwater et al., 1999a). It has been reported for littoral zones of lakes, ponds and slow streams. It seems to tolerate slightly salty conditions. It is polythermophilic, oligorheophilic, mesotitanophilic and euryplastic for pH (Meisch, 2000).
Potamocypris unicaudata? Schäfer, 1943
(Fig. 2R)
Identification: Meisch (2000).
Size: Adult: L: 0.68 to 0.87mm (Table 5). Juveniles: LV: L: 0.35mm, H: 0.20mm.
Morphology: Adults: see Meisch (2000). Juveniles: Valves smooth, with only few and sparse setae. Posterior valve margin steeply sloping posteriorly.
Distribution: Lake Tecuitlapa, East-Central Mexico. It has been reported in Puebla (Cohuo, 2012), and Michoacán (Garduño-Monroy et al., 2011). Outside Mexico it has been collected in North America and Europe (Meisch 2000).
Ecological preferences: Nektobenthic. This species was collected at temperature 26.2°C, pH 10.3, DO 5.7mg/L, conductivity 3 710µS/cm, and TDS 2.4g/L. The dominant ions of lake waters were Na+, CO32-, HCO3- and Cl- (Table 2). Cohuo (2012) collected this species in waters with higher DO (8.26mg/L), lower temperature (17.7°C) and conductivity (774µS/cm). It lives in freshwater and slightly brackish habitats. It has been collected in alkaline waters (pH =11.1). The maximum reported water depth of this species is 2m. The preferred substrate seems to be a thin layer of mud overlaying a sandy substrate (Meisch, 2000).
Discussion
Our study reveals that ten ostracode species inhabit ten highland lakes in East-Central Mexico. Species richness of non-marine ostracodes in the studied area is relatively low (≤6 spp. per lake). Similar results have been reported for other areas in Mexico and nearby regions. Caballero et al. (2013) reported six ostracode species in the crater lake Santa María del Oro in Western Mexico. Bridgwater et al. (1999a) reported nine ostracode species in modern and Holocene core samples from Lake Patzcuaro, Michoacán, Mexico. There are other paleoenvironmental studies in Central Mexico reporting the ostracode fauna. However, these reports did not study the extant fauna. Pérez et al. (2011) studied the ostracode fauna of mainly karst aquatic ecosystems from the Yucatán Peninsula and surrounding areas of Belize and Guatemala. Such karst environments are optimal for ostracode development, and therefore species abundances are expected to be high. A total of 29 species were identified and the highest species richness reported per lake was of 10 spp. (Pérez et al., 2011). Nevertheless, valve variability in the Yucatán Peninsula and in Central Mexico is high, which makes an accurate identification difficult. Therefore, such studies should carry out taxonomy-molecular analyses. Higher latitude lakes in North America display similar number of species per lake, as well. For instance, lakes in the Yukon territory, Canada display from three to eight ostracode species (Bunbury, & Gajewski, 2005). In contrast, other regions of the world are habitat a much higher number of ostracode species. The ancient Lake Ohrid, Macedonia and Albania and Lake Titicaca, Peru and Bolivia hold ca. 50 ostracode species each, and approximately half of them are endemic. Lakes Baikal, Russia and Tanganyika, Africa are habitat of ca. 200 species and more than the 90% of which are endemic. The higher number of species is not only attributed to the age of the lakes, but to the interaction of different environmental factors (Martens & Schön, 1999).
The studied aquatic ecosystems include maar lakes, tectonic-volcanic lakes and dams (natural and man-made). Ostracode species richness in maar lakes is low (2-3 spp.) and the ostracode species composition among studied maar lakes shows differences. For instance, the maar lakes La Preciosa, Quechulac and Alchichica share species in common (E. cf. patzcuaro and L. axalapasco) different to those from maar lakes Tecuitlapa (P. unicaudata?) and Aljojuca (C. vidua, F. acuminata?, P. smaragdina?). Lakes Tecuitlapa and Aljojuca do not have species in common, even though they are located close to one another. One reason could be that both lakes display different limnological variables as well as water chemical composition. Conductivity and sodium concentrations in the studied maar lakes were higher (≤12 940µS/cm, ≤115.50meq/L) among other measured cations, which could be a result of high evaporation in the region, and groundwater influence (Armienta et al., 2008). Chloride displayed the highest concentrations among anions (≤110.44meq/L). Crater lake Alchichica is a tropical saline oligotrophic lake (Alcocer, & Filonov, 2007), and the highest concentrations of Na+, Mg2+, K+, Cl-, SO42- and CO32- were encountered in this lake. We found only two ostracode species living in lake Alchichica: Eucandona cf. patzcuaro and Limnocytherina axalapasco. Eucandona patzcuaro, was previously classified as Candona patzcuaro, but Karanovic (2012) suggests that it belongs to the genus Eucandona. Shell variability of this species in the studied lakes seems to be high, and therefore coupled taxonomy and molecular analyses should be carried out in the future to decipher if it is one or more species. The reported salinity tolerance of this species is wide, which is uncommon in freshwater species of this genus, reason why we believe this might be another species restricted to this particular area. Future studies should focus on the revision of E. patzcuaro in Mexican lakes. Limnocytherina axalapasco was only collected in lakes La Preciosa, Alchichica, Atlagantepec, Quechulac and Metztitlán and a recent study (Cohuo et al., 2014) suggests that this species is restricted to East-Central Mexico, and therefore is a new and endemic species for this region. Low species richness and abundances in the study area suggest that the environmental and biological conditions in the lakes are not optimal for ostracodes. However, these are only preliminary results and further studies should be carried out during different seasons to have a better overview of the ostracode fauna and adult and juvenile abundances in highland lakes from East-Central Mexico.
The natural dams Zempoala and Atezca displayed only two species, while Metztitlán displayed the highest species richness (6 spp.) among studied lakes. Lake Metztitlán, a natural dam as well, displayed the highest species richness (6 spp.), and displays a water type HCO3->>SO42->Cl- -- Ca2+>Na+>Mg2+. Ostracode shell is mainly build of calcite and therefore Ca2+ content in lake waters is especially important (Keyser, & Walter, 2004). The highest Ca2+ concentration (≤4.04meq/L) among studied lake waters was determined in Lake Metztitlán, which could partly explain the higher number of ostracode species. Smith (1993) reported that ostracodes of lakes in the USA prefer waters dominated by Ca2+, SO42- and HCO3-, similar to our results. Additionally, Lake Metztitlán is a large and shallow natural dam (29.0km2, 3m deep), with a larger macrophyte cover than in other smaller lakes. This habitat provides shelter, protection against predation and higher food availability.
Chlamydotheca arcuata? was only collected in the volcanic-tectonic lake Tecocomulco. Lake Tecocomulco, a Ramsar site (Ramsar, 2013), is another large and shallow lake, that only displayed two species. The lake displays a higher altitude (2 535masl). Water temperatures in this lake are much colder (21.2°C) than in other lakes, and the lake water type is HCO3->>Cl- -- Na+>>Ca2+-Mg2+. Such environmental conditions seem to be unfavorable for ostracode development. The man-made dam, lake Atlangatepec, shared more species in common with the maar lakes Quechulac, La Preciosa and Alchichica than with other studied lakes.
The relatively low species richness in the studied highland lakes could be explained mainly by the lake water composition of the studied lakes, because it is largely determined by the bedrock geology and climate. Generally, ostracodes are highly sensitive to the change in the water chemical composition (Smith, 1993). The lake waters’ pH, and temperature are high (8.9, 30°C), and there is sufficient DO (5.8mg/L) for an optimal ostracode development, reproduction and colonization. However, our study shows that most highland lakes in East-Central Mexico display as much as three ostracodes species per lake, except for lake Metztitlán. Factors that could influence the presence or absence of ostracodes are the following: competition among species, depredation, food availability, water pollution and human impact, among others.
The Palaearctic zone is the zoographical zone with the highest number of the extant non-marine ostracode species (n=702), followed by the Afrotropical (n=455), Nearctic (n=298) and the Neotropical zones (n=275). One of the reasons why few species are reported for the Neotropical zone is the few number of studies that have been conducted in the region, especially in lakes in Central America. Future studies will shed light information on the actual number of ostracode species inhabiting aquatic ecosystems as well as the number of new and endemic species. Waterbodies in Mexico are located in the Nearctic-Neotropical transitional climatic zone, suggesting the presence of species of Nearctic and/or Neotropical origin, as well as endemic species. Most of the collected ostracode species display a Nearctic distribution. There are as well species with a wide geographic distribution and have been reported for other continents. Limnocythere friabilis is a Nearctic species, while E. patzcuaro is distributed in the Nearctic and Nearctic-Neotropical transitional zone. Chlamydotheca arcuata? has been reported in the Nearctics and Neotropics. Three out of ten ostracode species are widely distributed in East-Central Mexico (five lakes), as well as in other regions of Mexico, and the world. They include: E. cf. patzcuaro, C. vidua and D. stevensoni. Eucandona patzcuaro has been already reported for Mexico (Bridgwater et al., 1999b; Maddocks et al., 2009; Garduño-Monroy et al., 2011; Caballero et al., 2013). Tressler (1954) based the description of Eucandona patzcuaro on two male specimens only. Future studies should provide a description of adult females as well. We collected abundant specimens (adult and juveniles, 90 valves/mL wet sediment). Eucandona patzcuaro is restricted to the region from Central Mexico to South and Western USA. This is supported by Cohuo (2012) and Pérez et al. (2011), who do not report this species for lower latitudes (Southern Mexico, Guatemala and Belize). Lake Alchichica was the lake with the highest salinity (8.5g/L) in our study (Alcocer, & Escobar-Briones, 2007). Eucandona cf. patzcuaro and Limnocytherina axalapasco were the only two species collected in this lake, suggesting that these species can tolerate a broad range of salinity, because they can also be found in fresher waterbodies. Interestingly, adult and juvenile specimens of E. cf. patzcuaro were collected in surface sediments (45m) from the profundal zone. Similar results have been reported by Hernández et al. (2010). Future studies should focus on studying the distribution of this species in the lake (including more sites in the profundal zone), taphonomy, as well as a continuous seasonal sampling. Cypridopsis vidua and D. stevensoni are species with a wider distribution. Both species are highly tolerant and have been reported in all continents (Meisch, 2000). Cypridopsis vidua was most abundant in Lake Aljojuca that displayed lake waters with a high pH (9.5) and dominated by Na+, Mg2+, HCO3- and CO32-. Darwinula stevensoni was reported in five lakes displaying different physical and chemical variables, as well as water chemical composition, confirming the high tolerance of this species reported by Meisch (2000).
A higher number of rare species (6 spp., <five lakes) inhabit the study area. Species that have been reported for other regions in Mexico include: C. arcuata?, I. gibba? and P. smaragdina?. Two unidentified species of the genus Ilyocypris were reported by Cohuo (2012) in Tlacolula, Veracruz. It is possible that the species we collected in East-Central Mexico could be the same as the ones collected by Cohuo (2012) and the species collected in crater lakes in the highlands of Guatemala by Pérez et al. (2012). Nevertheless, further sampling campaigns should be carried out to collect well-preserved adult soft parts for their accurate identification. Meisch (2000) reported that I. gibba is abundant from May to September. Our fieldtrips were between June and October, however, only few single valves were collected and no ostracodes with well-preserved valves. First reports for Central Mexico are F. acuminata? and L. friabilis?. Limnocytherina axalapasco is an endemic species restricted to East-Central Mexico (Cohuo et al., 2014, and this study) while F. acuminata and L. friabilis are distributed in higher latitudes. Limnocytherina axalapasco has been already reported for the region but as Limnocythere itasca (Juárez, 2005), and Limnocythere inopinata (Hernández et al., 2010). Both species are very similar but analysis and comparison of their copulatory organs allows differentiating them.
Our study shows that ostracodes inhabit crater and other highland lakes in East-Central Mexico and some species are abundant and have specific environmental requirements, suggesting their potential as paleoenvironmental indicators. Additional (paleo) bioindicators (cladocerans, chironomids, diatoms, thecamoebians, among others) should be as well taken into account. Few studies have reported their presence and abundances in highland lakes in East-Central Mexico (Caballero et al., 1999; Quiroz Castelán, Díaz Vargas, Trejo Albarrán, & Elizalde Arriaga, 2000; Caballero et al., 2003; Hernández et al., 2010). Maar lakes contain high-resolution climate archives and their little human impact in some cases makes them a unique tool for reconstructing past environment and climate change in Mexico. Future studies will attempt to use ostracodes as indicators of late Quaternary climate change in East-Central Mexico. Intensive sampling and soft part as well as molecular analysis are needed to identify new and endemic species in the area. Future studies will attempt to use ostracodes as paleoenvironmental indicators in maar lakes in East-Central Mexico.
Acknowledgments
We would like to thank all participants in our field trips: Edyta Zawisza (Instituto de Geofísica, UNAM), Alexander Correa-Metrio and Esperanza Torres (Instituto de Geología, UNAM). Special thanks go to María Aurora Armienta and the staff of the Departamento de Química Analítica, Instituto de Geofísica, UNAM for the water chemical analysis. We also thank the agencies and institutions that provided financial support: CONACYT (project number 167621), National Science Foundation (NSF award number 0902864) and the Instituto de Geología, UNAM.
References
Alcántara-Rodríguez, J. A., Ciros-Pérez, J., Ortega-Mayagoitia, E., Serrania-Soto, C. R., & Piedra-Ibarra, E. (2012). Local adaptation in populations of a Brachionus group plicatilis cryptic species inhabiting three deep crater lakes in Central Mexico. Freshwater Biology, 57, 728-740. [ Links ]
Alcocer, J., Escobar, E., Lugo, A., & Peralta, L. (1998). Littoral benthos of the saline crater lakes of the basin of Oriental, Mexico. International Journal of Salt Lake Research, 7, 87-108. [ Links ]
Alcocer, J., & Lugo, A. (2003). Effects of El Niño on the dynamics of Lake Alchichica, central Mexico. Geofísica Internacional, 42, 523-528. [ Links ]
Alcocer, J., & Escobar-Briones, E. (2007). On the ecology of Caecidotea williamsi Escobar-Briones & Alcocer (Crustacea: Isopoda: Asellidae) from Alchichica saline lake, Central Mexico. Hydrobiologia, 576, 103-109. [ Links ]
Alcocer, J., & Filonov, A. (2007). A note on the effects of an individual large rainfall event on saline Lake Alchichica, Mexico. Environmental Geology, 53, 777-783. [ Links ]
Alcocer, J., & Bernal-Brooks, F. W. (2010). Limnology in Mexico. Hydrobiologia, 644, 15-68. [ Links ]
Alcocer, J., Arce, E., Zambrano, L., & Chiappa-Carrara, X. (2010). Poblana alchichica: A threatened silverside species? Verhandlungen Internationale Vereinigung für theoretische und angewandte Limnologie, 30, 1429-1432. [ Links ]
Almeida Leñero, L. (1973). La fauna de ostrácodos en los sedimentos recientes del lago de Tequesquitengo, Morelos (Tesis de licenciatura). Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
APHA (American Public Health Association), AAWWA (American Water Works Association), WPCF (Water Pollution Control Federation). (1995). Standard Methods for the Examination of Water and Wastewater. Washington, D.C., USA: American Health Association. [ Links ]
APHA (American Public Health Association), AAWWA (American Water Works Association), WPCF (Water Pollution Control Federation). (2005). Standard Methods for the Examination of Water and Wastewater. Maryland, USA: American Publich Health Association. [ Links ]
Armienta, M. A., Vilaclara, G., De la Cruz-Reyna, S., Ramos, S., Ceniceros, N., Cruz, O., Aguayo, A., & Arcega-Cabrera, F. (2008). Water chemistry of lakes related to active and inactive Mexican volcanoes. Journal of Volcanology and Geothermal Research, 178, 249-258. [ Links ]
Barluenga, M., Stölting, K. N., Salzburger, W., Muschick, M., & Meyer, A. (2006). Sympatric speciation in Nicaraguan crater lake cichlid fish. Nature, 439, 719-723. [ Links ]
Benson, R. H., & MacDonald, H. C. (1963). Postglacial (holocene) ostracodes from Lake Erie. Palaeontological Contribution University Kansas, 33, 1-26. [ Links ]
Brehm, V. (1932). Notizen zur Süßwasserfauna Guatemalas und Mexikos. Zoologischer Anzeiger, 91, 63-66. [ Links ]
Bridgwater, N. D., Heaton, T. H. E., & O’Hara, S. L. (1999a). A late Holocene paleolimnological record from central Mexico, based on faunal and stable-isotope analysis of ostracod shells. Journal of Paleolimnology, 22, 383-397. [ Links ]
Bridgwater, N. D., Holmes, J. A., & O’Hara, S. L. (1999b). Complex controls on the trace-element chemistry of non-marine ostracods: an example from Lake Pátzcuaro, central Mexico. Palaeogeography, Palaeoclimatology, Palaeoecology, 148, 117-131. [ Links ]
Brouwers, E. (1990). Systematic paleontology of Quaternary ostracode assemblages from the Gulf of Alaska, part 1: Families Cytherellidae, Bairdiidae, Cytheridae, Leptocytheridae, Limnocytheridae, Eucytheridae, Krithidae, Cushmanideidae. Washington, USA: United States Government printing office. [ Links ]
Bunbury, J., & Gajewski, K. (2005). Quantitative analysis of freshwater ostracode assemblages in southwestern Yukon Territory, Canada. Hydrobiologia, 545, 117-128. [ Links ]
Caballero, M., Lozano, S., Ortega, B., Urrutia, J., & Macias, J. L. (1999). Environmental characteristics of Lake Tecocomulco, northern basin of Mexico, for the last 50,000 years. Journal of Paleolimnology, 22, 399-411. [ Links ]
Caballero, M., Vilaclara, G., Rodríguez, A., & Juárez, D. (2003). Short-term climatic change in lake sediments from lake Alchichica, Oriental, Mexico. Geofísica Internacional, 42, 529-537. [ Links ]
Caballero, M., Lozano-García, S., Vázquez-Selem, L., & Ortega, B. (2010). Evidencias de cambio climático y ambiental en registros glaciales y en cuencas lacustres del centro de México durante el último máximo glacial. Boletín de la Sociedad Geológica Mexicana, 62, 359-377. [ Links ]
Caballero, M., Rodríguez, A., Vilaclara, G., Ortega, B., Roy, P., & Lozano, S. (2013). Hydrochemistry, ostracods and diatoms in a deep, tropical crater lake in western Mexico. Journal of Limnology, 72, 512-523. [ Links ]
Carreño, A. L. (1990). Ostrácodos lacustres del paleolago de Texcoco. Revista de la Sociedad Mexicana de Paleontología, 3, 117-129. [ Links ]
Chávez, C. M. (2011). Paleoecología de ostrácodos en los últimos 45 cal ka A.P., procedentes de los sedimentos del paleolago Babícora, desierto de Chihuahua (Tesis de licenciatura). Facultad de Ingeniería, Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
Cohen, A. (2003). Paleolimnology. New York, USA: Oxford University Press. [ Links ]
Cohuo, S. (2012). Revisión taxonómica (morfológica y molecular) de los ostrácodos dulceacuícolas del centro-sureste de México (Tesis de maestría). Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
Cohuo, S., Pérez, L., & Karanovic, I. (2014). On Limnocytherina axalapasco, a new freshwater ostracod (Ostracoda: Limnocytheridae) from Mexican crater lakes. Revista de Biología Tropical, 62, 15-32. [ Links ]
Conserva, M. E., & Byrne, R. (2002). Late Holocene Vegetation Change in the Sierra Madre Oriental of Central Mexico. Quaternary Research, 58, 122-129. [ Links ]
Curry, B. B. (1999). An environmental tolerance index for ostracodes as indicators of physical and chemical factors in aquatic habitats. Palaeogeography, Palaeoclimatology, Palaeoecology, 148, 51-63. [ Links ]
Curry, B. B., & Baker, R. G. (2000). Palaeohydrology, vegetation, and climate since the late Illinois Episode (~130 ka) in south-central Illinois. Palaeogeography, Palaeoclimatology, Palaeoecology, 155, 59-81. [ Links ]
Curry, B. B., & Yansa, C. H. (2004). Evidence for stagnation of the Harvard Sublove (Lake Michigan Lobe) in Northeastern Illinois, U.S.A., from 24000 to 17600 BP and subsequent Tundra-like ice-marginal paleoenvironments from 17600 to 15700 BP. Geógraphie physique et Quaternaire, 58, 305-321. [ Links ]
Cusminsky, G. C., Pérez, P. A., Schwalb, A., & Whatley, R. (2005). Recent Lacustrine Ostracods from Patagonia, Argentina. Revista Española de Micropaleontología, 37, 431-450. [ Links ]
Daday, E. (1905). Untersuchungen über die Süsswasser-Mikrofauna Paraguays. Zoologica, 18, 1-374. [ Links ]
Davies, S. J., Metcalfe, S. E., Caballero, M. E., & Juggins, S. (2002). Developing diatom-based transfer functions for Central Mexican lakes. Hydrobiologia, 467, 199-213. [ Links ]
de la Lanza-Espino, G., Gómez-Rodríguez, G., Islas Islas, A., Escalante Richards, V., & Hernández Pulido, S. (2011). Analysis of the effect of El Niño and La Niña on Tecocomulco Lake, central basin, Mexico. Hidrobiológica, 21, 249-259. [ Links ]
de Saussure, H. (1858). Mèmoire sur divers crustacés nouveaux du Mexique et des Antilles Mémoirs pour servir a l’histoire naturelle du Mexique, des Antilles et des États-Unis -- 1er mémoire. Ginebra, Suiza: Imprimerie Jules G. Fick. [ Links ]
Deckker, P. (1981). Ostracods of athalassic saline lakes. Hydrobiologia, 81-82, 131-144. [ Links ]
Delorme, L. D. (1970). Freshwater ostracodes of Canada. Part I. Subfamily Cypridinae. Canadian Journal of Zoology, 48, 153-168. [ Links ]
Delorme, D. (1971). Freshwater ostracodes of Canada. Part V: Families Limnocytheridae, Loxoconchidae. Canadian Journal of Zoology, 49, 43-64. [ Links ]
Dennison-Budak, C. W. (2010). Ostracodes as indicators of the paleoenvironment in the Pliocene Glenns Ferry Formation, Glenns Ferry Lake, Idaho (Master’s thesis). Kent State University, Ohio, USA. [ Links ]
Díaz, A. R., & Lopretto, E. C. (2011). The genus Chlamydotheca Saussure (Crustacea: Ostracoda) in northeastern Argentina. Nauplius, 19, 97-107. [ Links ]
Dole-Olivier, M. J., Galassi, D. M. P., Marmonier, P., & Creuzé des Chatelliers, M. (2000). The biology and ecology of lotic microcrustaceans. Freshwater Biology, 44, 63-91. [ Links ]
Ehrenberg, C. G. (1869). Über mächtige Gebirgsschichten vorherrschend aus mikroskopischen Bacillarien unter und bei der Stadt Mexiko. Berlin, Germany: Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin, Physikalische Klasse 1869. [ Links ]
Ferguson, E., Jr. (1958). Freshwater Ostracods From South Carolina. American Midland Naturalist, 59, 111-119. [ Links ]
Forester, R. M. (1985). Limnocythere bradburyi n.sp.: A modern ostracode from central Mexico and possible Quaternary paleoclimatic indicator. Journal of Paleontology, 59, 8-20. [ Links ]
Forester, R. M., & Smith, A. J. (1994). Late glacial climate estimates for southern Nevada, the ostracode fossil record. High Level Radioactive Waste Management, 1, 2553-2561. [ Links ]
Forester, R. M., Colman, S. M., Reynolds, R. L., & Keigwin, L. D. (1994). Lake Michigan’s Late Quaternary Limnological and Climate History From Ostracode, Oxygen Isotope, and Magnetic Susceptibility. Journal of Great Lakes Research, 20, 93-107. [ Links ]
Forester, R., Bradbury, J. P., Carter, C., Lundstrom, S., Mahan, S., Marshall, B., Paces, J., Whelan, J., Neymark, L., Elvidge-Tuma, A., Hemphill, M., Sharpe, S., & Wigand, P. (1999). The climatic and hydrologic history of southern Nevada during the late Quaternary. Denver, Colorado, USA: U.S. Geological Survey. [ Links ]
Forester, R., Smith, A. J., Palmer, D. F., & Curry, B. (2013). NANODe Version 1: North American Non-Marine Ostracode Database. USA: U.S. Geological Survey, Kent State University,National Science Foundation Programs: Biological Sciences, Earth Systems History, and Hydrologic Sciences. [ Links ]
Furtos, N. (1933). The Ostracoda of Ohio. In Ohio Biological Survey (Vol. V (6), pp. 413-524). Columbus, USA: The Ohio State University Press. [ Links ]
Furtos, N. (1936). On the Ostracoda from the cenotes of Yucatán and vicinity. In The cenotes of Yucatan, a zoological and hydrographic survey (pp. 89-115). Washington, USA: Carnegie Intitution of Washington. [ Links ]
Gabriel, J., Reinhardt, E., Peros, M., Davidson, D., Hengstum, P., & Beddows, P. (2009). Palaeoenvironmental evolution of Cenote Aktun Ha (Carwash) on the Yucatan Peninsula, Mexico and its response to Holocene sea-level rise. Journal of Paleolimnology, 42, 199-213. [ Links ]
García-Rodríguez, J., & Tavera, R. (2002). Phytoplankton composition and biomass in a shallow monomictic tropical lake. Hydrobiologia, 467, 91-98. [ Links ]
Garduño-Monroy, V. H., Soria-Caballero, D. C., Israde-Alcántara, I., Hernández Madrigal, M., Rodríguez-Ramírez, A., Ostroumov, M., Rodríguez-Pascua, M. A., Chacón-Torres, A., & Mora- [ Links ]
Chaparro, J. C. (2011). Evidence of tsunami events in the Paleolimnological record of Lake Pátzcuaro, Michoacán, Mexico. Geofísica Internacional, 50, 147-161. [ Links ]
Hartmann, G. (1959). Beitrag zur Kenntnis des Nicaragua-Sees unter besonderer Berücksichtigung seiner Ostracoden. Zoologischer Anzeiger, 162, 269-294. [ Links ]
Hernández, M., Escobar, E., & Alcocer, J. (2010). Benthic crustaceans assemblage in a tropical, saline lake. Revista Mexicana de Biodiversidad, 81, 133-140. [ Links ]
Horne, D. J. (2007). A mutual temperature range method for Quaternary palaeoclimatic analysis using European nonmarine Ostracoda. Quaternary Science Reviews, 26, 1398-1415. [ Links ]
Juárez, D. (2005). Registro de cambios paleoambientales en sedimentos del lago “La Preciosa”, Puebla, con base en el estudio de ostrácodos (Tesis de maestría). Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
Karanovic, I. (2012). Recent freshwater ostracods of the world. Berlin Heidelberg, Germany: Springer. [ Links ]
Kazmierczak, J., Kempe, S., Kremer, B., López-García, P., Moreira, D., & Tavera, R. (2011). Hydrochemistry and microbialites of the alkaline crater lake Alchichica, Mexico. Facies, 57, 543-570. [ Links ]
Keyser, D. (1976). Zur Kenntnis der brackigen mangrovebewachsenen Weichböden Südwest-Floridas unter besonderer Berücksitchtigung ihrer Ostracodenfauna. Doktorarbeit. Hamburg, Germany: Fachbereich Biologie, Universität Hamburg. [ Links ]
Keyser, D., & Walter, R. (2004). Calcification in ostracodes. Revista Española de Micropaleontología, 36, 1-11. [ Links ]
Külköylüoglu, O., & Vinyard, G. L. (2000). Distribution and ecology of freshwater Ostracoda (Crustacea) collected from springs of Nevada, Idaho, and Oregon: a preliminary study. Western North American Naturalist, 60, 291-303. [ Links ]
Külköylüoglu, O. (2004). On the usage of ostracods (Crustacea) as bioindicator species in different aquatic habitats in the Bolu region, Turkey. Ecological Indicators, 4, 139-147. [ Links ]
Li, X., Liu, W., Zhang, L., & Sun, Z. (2010). Distribution of Recent ostracod species in the Lake Qinghai area in northwestern China and its ecological significance. Ecological Indicators, 10, 880-890. [ Links ]
Lorenschat, J. (2009). Ostracoden aus Hoch- und Tieflandseen in südlichen Guatemala als Bioindikatoren. Diplomarbeit. Braunschweig, Germany: Institut für Umweltgeologie, Technische Universität Braunschweig. [ Links ]
Machain-Castillo, M. L., & Gío-Argáez, R. (2004). Ostrácodos bentónicos del sur del Golfo de México. In M. Caso, I. Pisanty, & R. Ezcurra (Eds.), Diagnóstico ambiental del Golfo de México (pp. 161-171). México: Instituto Nacional de Ecología (INE-SEMARNAT). [ Links ]
Maddocks, R. F., Machain-Castillo, M. L., & Gío-Argáez, R. (2009). Podocopan Ostracoda (Crustacea) of the Gulf of Mexico. In D. L. Felder & D. K. Camp, (Eds.), Gulf of Mexico-Origins, Water and Biota. Biodiversity (pp. 877-894). Texas, USA: Texas A & M Press, College. [ Links ]
Marmonier, P., & Chatelliers, M. C. D. (1992). Biogeography of the Benthic and Interstitial Living Ostracods (Crustacea) of the Rhone River (France). Journal of Biogeography, 19, 693-704. [ Links ]
Martens, K. (1996). On Korannocythere gen. nov. (Crustacea, Ostracoda), a new genus of temporary pool limnocytherids from southern Africa, with the description of three new species and a generic reassessment of the Limnocytherinae. Royal Sciences Naturelles de Belgique Biology, 66. [ Links ]
Martens, K., & Schön, I. (1999). Crustacean Biodiversity in Ancient Lakes: A Review. Crustaceana, 72, 899-910. [ Links ]
Martens, K. (2000). Factors affecting the divergence of mate recognition systems in the Limnocytherinae (Crustacea, Ostracoda). Hydrobiologia, 419, 83-101. [ Links ]
Meisch, C. (2000). Freshwater Ostracoda of western and central Europe. Heidelberg, Berlin, Germany: Spektrum Akademischer Verlag GmbH. [ Links ]
Metcalfe, S. E., Davies, S. J., Braisby, J. D., Leng, M. J., Newton, A. J., Terrett, N. L., & O’Hara, S. L. (2007). Long and short-term change in the Pátzcuaro Basin, central Mexico. Palaeogeography, Palaeoclimatology, Palaeoecology, 247, 272-295. [ Links ]
Mezquita, F., Roca, J. R., Reed, J. M., & Wansard, G. (2005). Quantifying species-environment relationships in non-marine Ostracoda for ecological and palaeoecological studies: examples using Iberian data. Palaeogeography, Palaeoclimatology, Palaeoecology, 225, 93-117. [ Links ]
Mischke, S., Almogi-Labin, A., Ortal, R., Rosenfeld, A., Schwab, M. J., & Boomer, I. (2010). Quantitative reconstruction of lake conductivity in the Quaternary of the Near East (Israel) using ostracods. Journal of Paleolimnology, 43, 667-688. [ Links ]
Nagorskaya, L. (2002). Living freshwater Ostracoda (Crustacea) of a floodplain relict oak forest (Polesye, Belarus). Revista Española de Micropaleontología, 36, 135-145. [ Links ]
Nagorskaya, L., & Keyser, D. (2005). Habitat diversity and ostracod distribution patterns in Belarus. Hydrobiologia, 538, 167-178. [ Links ]
Oliva, M. G., Lugo, A., Alcocer, J., & Cantoral-Uriza, E. A. (2006). Cyclotella alchichicana sp. nov. from a saline mexican lake. Diatom Research, 21, 81-89. [ Links ]
Palacios Fest, M., Cohen, A., Ruiz, J., & Blank, B. (1993). Comparative paleoclimatic interpretations from nonmarine ostracodes using faunal assemblages, trace elements, shell chemistry and stable isotope data. Geophysical Monograph, 78, 179-189. [ Links ]
Palacios-Fest, M. R., & Dettman, D. L. (2001). Temperature controls monthly variation in ostracode valve Mg/Ca: Cypridopsys vidua from a small lake in Sonora, Mexico. Geochimica et Cosmochimica Acta, 65, 2499-2507. [ Links ]
Palacios-Fest, M., Carreño, A., Ortega-Ramírez, J., & Alvarado-Valdéz, G. (2002). A paleoenvironmental reconstruction of Laguna Babícora, Chihuahua, Mexico based on ostracode paleoecology and trace element shell chemistry. Journal of Paleolimnology, 27, 185-206. [ Links ]
Palacios-Fest, M. (2004). Ostracode paleoecology of Red Mountain Site, Mesa, Arizona. In Life on the Lehi Terrace: the Archaeology of the Red Mountain Freeway between Country Club Drive and Gilbert Road. USA: Arizona Department of Transportation Environmental Planning Group, ADOT Project RAM 600-8-326, TRACS 202L-MA20-H3872-01D, 737-750. [ Links ]
Palacios-Fest, M. (2010). Late Holocene paleoenvironmental history of the upper west Amarillo Creek Valley at Archaeological Site 41PT185/C, Texas, USA. Boletín de la Sociedad Geológica Mexicana, 62, 399-436. [ Links ]
Pérez, L. (2010). Non-marine ostracodes from the Yucatán Peninsula as late Quaternary paleoenvironmental indicators (Doctoral thesis). Architektur, Bauingenieurwesen und Umweltwissenschaften, Technische Universität Braunschweig, Braunschweig, Germany. [ Links ]
Pérez, L., Bugja, R., Massaferro, J., Steeb, P., van Geldern, R., Frenzel, P., Brenner, M., Scharf, B., & Schwalb, A. (2010a). Post-Columbian environmental history of Lago Petén Itzá, Guatemala. Revista Mexicana de Ciencias Geológicas, 27, 490-507. [ Links ]
Pérez, L., Lorenschat, J., Brenner, M., Scharf, B., & Schwalb, A. (2010b). Extant freshwater ostracodes (Crustacea: Ostracoda) from Lago Petén Itzá, Guatemala. Revista de Biología Tropical, 58, 871-895. [ Links ]
Pérez, L., Frenzel, P., Brenner, M., Escobar, E., Hoelzmann, P., Scharf, B., & Schwalb, A. (2011). Late Quaternary (24-10 ka BP) environmental history of the Neotropical lowlands inferred from ostracodes in sediments of Lago Petén Itzá, Guatemala. Journal of Paleolimnology, 46, 59-74. [ Links ]
Pérez, L., Lorenschat, J., Brenner, M., Scharf, B., & Schwalb, A. (2012). Non-marine ostracodes (Crustacea) of Guatemala. In E. Cano & J. Shuster (Eds.), Biodiversidad de Guatemala II (pp. 121-131). Guatemala: Universidad del Valle de Guatemala. [ Links ]
Pérez, L., Lorenschat, J., Massaferro, J., Pailles, C., Sylvestre, F., Hollwedel, W., Brandorff, G.-O. , Brenner, M., Islebe, G., Lozano, S., Scharf, B., & Schwalb, A. (2013). Bioindicators of climate and trophic state in lowland and highland aquatic ecosystems of the Northern Neotropics. Revista de Biología Tropical, 61, 603-644. [ Links ]
Quiroz Castelán, H., Díaz Vargas, M., Trejo Albarrán, R., & Elizalde Arriaga, E. (2000). Aspectos sobre la abundancia y distribución de los principales grupos de la fauna bentónica en el lago “Zempoala”, Morelos, México. Ciencia y mar, 4, 39-50. [ Links ]
Ramírez-García, P., & Vázquez-Gutiérrez, F. (1989). Contribuciones al estudio limnobotánico de la zona litoral de seis lagos cráter del Estado de Puebla. Anales del Instituto de Ciencias del Mar y Limnología, 323, 1-16. [ Links ]
Ramsar. (2013). Humedales de México. Retrieved from http://ramsar.conanp.gob.mx/lsr.php [ Links ]
Rio de la Loza, L., & Craveri, C. (1858). Opúsculo sobre los Pozos Artesianos y las Aguas Naturales de más uso en la Ciudad de México, con algunas noticias relativas al Corte Geológico del Valle, y una lista de las Plantas que vegetan en las inmediaciones del Desierto Viejo. Boletín de la Sociedad Mexicana de Geografía y Estadística, 6, 9-28. [ Links ]
Rodríguez, G. (2002). Biodiversidad de los crustáceos dulceacuícolas del centro de Nuevo León y noroeste de Tamaulipas (Reporte final de proyecto S104, CONABIO). Departamento de Zoología de Invertebrados, Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, Nuevo León, México. Retrived from http://www.conabio.gob.mx/institucion/cgi-bin/datos.cgi?Letras=S & Numero=104 [ Links ]
Rodríguez, E. (2011). Estudio morfológico de ostrácodos dulceacuícolas de Durango y Veracruz, México (Tesis de licenciatura). Facultad de Ciencias, Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
Roy, P. D., Caballero, M., Lozano, R., & Smykatz-Kloss, W. (2008). Geochemistry of late quaternary sediments from Tecocomulco lake, central Mexico: Implication to chemical weathering and provenance. Chemie der Erde - Geochemistry, 68, 383-393. [ Links ]
Roy, P. D., Caballero, M., Lozano, R., Pi, T., & Morton, O. (2009). Late Pleistocene-Holocene geochemical history inferred from Lake Tecocomulco sediments, Bain of Mexico, Mexico. Geochemical Journal, 43, 49-64. [ Links ]
Saldivar-López, D. B. (2011). Platelmintos y crustáceos presentes en la vegetación acuática, sus condiciones de hábitat y sus relaciones interespecíficas, en la Presa de Apizaquito (Tesis de licenciatura). División de Ciencias Biológicas y de la Salud, Departamento El Hombre y su ambiente, Universidad Autónoma Metropolitana, Unidad Xochimilco, México. [ Links ]
Sars, G. (1901). Contributions to the knowledge of the fresh-water Entomostraca of South America as shown by artificial hatching from dried material. Part II. Copepoda-Ostracoda. Archiv fur Mathematik og Naturvidenskab, 24, 1-52. [ Links ]
Smith, A. J. (1993). Lacustrine ostracodes as hydrochemical indicators in lakes of the north-central United States. Journal of Paleolimnology, 8, 121-134. [ Links ]
Smith, A. J., & Delorme, L. D. (2010). Ostracoda. In J. Thorp & A. Covich (Eds.), Ecology and Classification of North American Freshwater Invertebrates (pp. 725-771). United Kingdom:Academic Press, Elsevier. [ Links ]
Soria Caballero, D. C. (2010). Estudio bioestratigráfico y mineralógico de la ciénega de Zacapu, Michoacán (Tesis de maestría). Instituto de Investigaciones Metalúrgicas, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, México. [ Links ]
Staplin, F. L. (1963). Subfamilies Cyclocyprinae, Cypridopsinae, Ilyocyprinae, families Darwinulidae and Cytheridae. Stratigraphic ranges and assemblage patters. Journal of Paleontology, 37, 1164-1203. [ Links ]
Suter, M. (2004). A neotectonic-geomorphologic investigation of the prehistoric rock avalanche damming Laguna de Metztitlán (Hidalgo State, East-central Mexico). Revista Mexicana de Ciencias Geológicas, 21, 397-411. [ Links ]
Szlauer-Lukaszewska, A. (2012). Ostracod assemblages in relation to littoral plant communities of a shallow lake (Lake Swidwie, Poland). International Review of Hydrobiology, 97, 262-275. [ Links ]
Tressler, W. L. (1949). Fresh-water Ostracoda from Brazil. Proceeding of the United States National Museum, 100, 61-83. [ Links ]
Tressler, W. L. (1954). Fresh-water Ostracoda from Texas and Mexico. Journal of Washington Academy Science, 44, 138-149. [ Links ]
Vázquez, G. V., Ortega, B., Rodríguez, A., Caballero, M., & Lozano, S. (2008). Mineralogía magnética como indicador de sequía en los sedimentos lacustres de los últimos ca. 2,600 años de Santa María del Oro, occidente de México. Revista Mexicana de Ciencias Geológicas, 25, 21-38. [ Links ]
Viehberg, F. A., & Mesquita-Joanes, F. (2012). Chapter 4 - Quantitative Transfer Function Approaches in Palaeoclimatic Reconstruction Using Quaternary Ostracods. In D. J. Horne, J. A. Holmes, J. Rodriguez-Lazaro, & F. Viehberg (Eds.), Developments in Quaternary Sciences (pp. 47-64). Amsterdam, Netherlands: Elsevier. [ Links ]
Wetterich, S. (2008). Freshwater ostracods as bioindicators in Arctic periglacial regions. Thesis. Potsdam, Germany: Mathematisch-Naturwissenschaftlichen Fakultät Universität Potsdam. [ Links ]
Zara-environmental-LCC. (2010). Final report for deep aquifer biota study of the Edwards Aquifer. Texas, USA: Zara-environmental-LCC. [ Links ]
Alcocer, J., Escobar, E., Lugo, A., & Peralta, L. (1998). Littoral benthos of the saline crater lakes of the basin of Oriental, Mexico. International Journal of Salt Lake Research, 7, 87-108. [ Links ]
Alcocer, J., & Lugo, A. (2003). Effects of El Niño on the dynamics of Lake Alchichica, central Mexico. Geofísica Internacional, 42, 523-528. [ Links ]
Alcocer, J., & Escobar-Briones, E. (2007). On the ecology of Caecidotea williamsi Escobar-Briones & Alcocer (Crustacea: Isopoda: Asellidae) from Alchichica saline lake, Central Mexico. Hydrobiologia, 576, 103-109. [ Links ]
Alcocer, J., & Filonov, A. (2007). A note on the effects of an individual large rainfall event on saline Lake Alchichica, Mexico. Environmental Geology, 53, 777-783. [ Links ]
Alcocer, J., & Bernal-Brooks, F. W. (2010). Limnology in Mexico. Hydrobiologia, 644, 15-68. [ Links ]
Alcocer, J., Arce, E., Zambrano, L., & Chiappa-Carrara, X. (2010). Poblana alchichica: A threatened silverside species? Verhandlungen Internationale Vereinigung für theoretische und angewandte Limnologie, 30, 1429-1432. [ Links ]
Almeida Leñero, L. (1973). La fauna de ostrácodos en los sedimentos recientes del lago de Tequesquitengo, Morelos (Tesis de licenciatura). Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
APHA (American Public Health Association), AAWWA (American Water Works Association), WPCF (Water Pollution Control Federation). (1995). Standard Methods for the Examination of Water and Wastewater. Washington, D.C., USA: American Health Association. [ Links ]
APHA (American Public Health Association), AAWWA (American Water Works Association), WPCF (Water Pollution Control Federation). (2005). Standard Methods for the Examination of Water and Wastewater. Maryland, USA: American Publich Health Association. [ Links ]
Armienta, M. A., Vilaclara, G., De la Cruz-Reyna, S., Ramos, S., Ceniceros, N., Cruz, O., Aguayo, A., & Arcega-Cabrera, F. (2008). Water chemistry of lakes related to active and inactive Mexican volcanoes. Journal of Volcanology and Geothermal Research, 178, 249-258. [ Links ]
Barluenga, M., Stölting, K. N., Salzburger, W., Muschick, M., & Meyer, A. (2006). Sympatric speciation in Nicaraguan crater lake cichlid fish. Nature, 439, 719-723. [ Links ]
Benson, R. H., & MacDonald, H. C. (1963). Postglacial (holocene) ostracodes from Lake Erie. Palaeontological Contribution University Kansas, 33, 1-26. [ Links ]
Brehm, V. (1932). Notizen zur Süßwasserfauna Guatemalas und Mexikos. Zoologischer Anzeiger, 91, 63-66. [ Links ]
Bridgwater, N. D., Heaton, T. H. E., & O’Hara, S. L. (1999a). A late Holocene paleolimnological record from central Mexico, based on faunal and stable-isotope analysis of ostracod shells. Journal of Paleolimnology, 22, 383-397. [ Links ]
Bridgwater, N. D., Holmes, J. A., & O’Hara, S. L. (1999b). Complex controls on the trace-element chemistry of non-marine ostracods: an example from Lake Pátzcuaro, central Mexico. Palaeogeography, Palaeoclimatology, Palaeoecology, 148, 117-131. [ Links ]
Brouwers, E. (1990). Systematic paleontology of Quaternary ostracode assemblages from the Gulf of Alaska, part 1: Families Cytherellidae, Bairdiidae, Cytheridae, Leptocytheridae, Limnocytheridae, Eucytheridae, Krithidae, Cushmanideidae. Washington, USA: United States Government printing office. [ Links ]
Bunbury, J., & Gajewski, K. (2005). Quantitative analysis of freshwater ostracode assemblages in southwestern Yukon Territory, Canada. Hydrobiologia, 545, 117-128. [ Links ]
Caballero, M., Lozano, S., Ortega, B., Urrutia, J., & Macias, J. L. (1999). Environmental characteristics of Lake Tecocomulco, northern basin of Mexico, for the last 50,000 years. Journal of Paleolimnology, 22, 399-411. [ Links ]
Caballero, M., Vilaclara, G., Rodríguez, A., & Juárez, D. (2003). Short-term climatic change in lake sediments from lake Alchichica, Oriental, Mexico. Geofísica Internacional, 42, 529-537. [ Links ]
Caballero, M., Lozano-García, S., Vázquez-Selem, L., & Ortega, B. (2010). Evidencias de cambio climático y ambiental en registros glaciales y en cuencas lacustres del centro de México durante el último máximo glacial. Boletín de la Sociedad Geológica Mexicana, 62, 359-377. [ Links ]
Caballero, M., Rodríguez, A., Vilaclara, G., Ortega, B., Roy, P., & Lozano, S. (2013). Hydrochemistry, ostracods and diatoms in a deep, tropical crater lake in western Mexico. Journal of Limnology, 72, 512-523. [ Links ]
Carreño, A. L. (1990). Ostrácodos lacustres del paleolago de Texcoco. Revista de la Sociedad Mexicana de Paleontología, 3, 117-129. [ Links ]
Chávez, C. M. (2011). Paleoecología de ostrácodos en los últimos 45 cal ka A.P., procedentes de los sedimentos del paleolago Babícora, desierto de Chihuahua (Tesis de licenciatura). Facultad de Ingeniería, Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
Cohen, A. (2003). Paleolimnology. New York, USA: Oxford University Press. [ Links ]
Cohuo, S. (2012). Revisión taxonómica (morfológica y molecular) de los ostrácodos dulceacuícolas del centro-sureste de México (Tesis de maestría). Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
Cohuo, S., Pérez, L., & Karanovic, I. (2014). On Limnocytherina axalapasco, a new freshwater ostracod (Ostracoda: Limnocytheridae) from Mexican crater lakes. Revista de Biología Tropical, 62, 15-32. [ Links ]
Conserva, M. E., & Byrne, R. (2002). Late Holocene Vegetation Change in the Sierra Madre Oriental of Central Mexico. Quaternary Research, 58, 122-129. [ Links ]
Curry, B. B. (1999). An environmental tolerance index for ostracodes as indicators of physical and chemical factors in aquatic habitats. Palaeogeography, Palaeoclimatology, Palaeoecology, 148, 51-63. [ Links ]
Curry, B. B., & Baker, R. G. (2000). Palaeohydrology, vegetation, and climate since the late Illinois Episode (~130 ka) in south-central Illinois. Palaeogeography, Palaeoclimatology, Palaeoecology, 155, 59-81. [ Links ]
Curry, B. B., & Yansa, C. H. (2004). Evidence for stagnation of the Harvard Sublove (Lake Michigan Lobe) in Northeastern Illinois, U.S.A., from 24000 to 17600 BP and subsequent Tundra-like ice-marginal paleoenvironments from 17600 to 15700 BP. Geógraphie physique et Quaternaire, 58, 305-321. [ Links ]
Cusminsky, G. C., Pérez, P. A., Schwalb, A., & Whatley, R. (2005). Recent Lacustrine Ostracods from Patagonia, Argentina. Revista Española de Micropaleontología, 37, 431-450. [ Links ]
Daday, E. (1905). Untersuchungen über die Süsswasser-Mikrofauna Paraguays. Zoologica, 18, 1-374. [ Links ]
Davies, S. J., Metcalfe, S. E., Caballero, M. E., & Juggins, S. (2002). Developing diatom-based transfer functions for Central Mexican lakes. Hydrobiologia, 467, 199-213. [ Links ]
de la Lanza-Espino, G., Gómez-Rodríguez, G., Islas Islas, A., Escalante Richards, V., & Hernández Pulido, S. (2011). Analysis of the effect of El Niño and La Niña on Tecocomulco Lake, central basin, Mexico. Hidrobiológica, 21, 249-259. [ Links ]
de Saussure, H. (1858). Mèmoire sur divers crustacés nouveaux du Mexique et des Antilles Mémoirs pour servir a l’histoire naturelle du Mexique, des Antilles et des États-Unis -- 1er mémoire. Ginebra, Suiza: Imprimerie Jules G. Fick. [ Links ]
Deckker, P. (1981). Ostracods of athalassic saline lakes. Hydrobiologia, 81-82, 131-144. [ Links ]
Delorme, L. D. (1970). Freshwater ostracodes of Canada. Part I. Subfamily Cypridinae. Canadian Journal of Zoology, 48, 153-168. [ Links ]
Delorme, D. (1971). Freshwater ostracodes of Canada. Part V: Families Limnocytheridae, Loxoconchidae. Canadian Journal of Zoology, 49, 43-64. [ Links ]
Dennison-Budak, C. W. (2010). Ostracodes as indicators of the paleoenvironment in the Pliocene Glenns Ferry Formation, Glenns Ferry Lake, Idaho (Master’s thesis). Kent State University, Ohio, USA. [ Links ]
Díaz, A. R., & Lopretto, E. C. (2011). The genus Chlamydotheca Saussure (Crustacea: Ostracoda) in northeastern Argentina. Nauplius, 19, 97-107. [ Links ]
Dole-Olivier, M. J., Galassi, D. M. P., Marmonier, P., & Creuzé des Chatelliers, M. (2000). The biology and ecology of lotic microcrustaceans. Freshwater Biology, 44, 63-91. [ Links ]
Ehrenberg, C. G. (1869). Über mächtige Gebirgsschichten vorherrschend aus mikroskopischen Bacillarien unter und bei der Stadt Mexiko. Berlin, Germany: Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin, Physikalische Klasse 1869. [ Links ]
Ferguson, E., Jr. (1958). Freshwater Ostracods From South Carolina. American Midland Naturalist, 59, 111-119. [ Links ]
Forester, R. M. (1985). Limnocythere bradburyi n.sp.: A modern ostracode from central Mexico and possible Quaternary paleoclimatic indicator. Journal of Paleontology, 59, 8-20. [ Links ]
Forester, R. M., & Smith, A. J. (1994). Late glacial climate estimates for southern Nevada, the ostracode fossil record. High Level Radioactive Waste Management, 1, 2553-2561. [ Links ]
Forester, R. M., Colman, S. M., Reynolds, R. L., & Keigwin, L. D. (1994). Lake Michigan’s Late Quaternary Limnological and Climate History From Ostracode, Oxygen Isotope, and Magnetic Susceptibility. Journal of Great Lakes Research, 20, 93-107. [ Links ]
Forester, R., Bradbury, J. P., Carter, C., Lundstrom, S., Mahan, S., Marshall, B., Paces, J., Whelan, J., Neymark, L., Elvidge-Tuma, A., Hemphill, M., Sharpe, S., & Wigand, P. (1999). The climatic and hydrologic history of southern Nevada during the late Quaternary. Denver, Colorado, USA: U.S. Geological Survey. [ Links ]
Forester, R., Smith, A. J., Palmer, D. F., & Curry, B. (2013). NANODe Version 1: North American Non-Marine Ostracode Database. USA: U.S. Geological Survey, Kent State University,National Science Foundation Programs: Biological Sciences, Earth Systems History, and Hydrologic Sciences. [ Links ]
Furtos, N. (1933). The Ostracoda of Ohio. In Ohio Biological Survey (Vol. V (6), pp. 413-524). Columbus, USA: The Ohio State University Press. [ Links ]
Furtos, N. (1936). On the Ostracoda from the cenotes of Yucatán and vicinity. In The cenotes of Yucatan, a zoological and hydrographic survey (pp. 89-115). Washington, USA: Carnegie Intitution of Washington. [ Links ]
Gabriel, J., Reinhardt, E., Peros, M., Davidson, D., Hengstum, P., & Beddows, P. (2009). Palaeoenvironmental evolution of Cenote Aktun Ha (Carwash) on the Yucatan Peninsula, Mexico and its response to Holocene sea-level rise. Journal of Paleolimnology, 42, 199-213. [ Links ]
García-Rodríguez, J., & Tavera, R. (2002). Phytoplankton composition and biomass in a shallow monomictic tropical lake. Hydrobiologia, 467, 91-98. [ Links ]
Garduño-Monroy, V. H., Soria-Caballero, D. C., Israde-Alcántara, I., Hernández Madrigal, M., Rodríguez-Ramírez, A., Ostroumov, M., Rodríguez-Pascua, M. A., Chacón-Torres, A., & Mora- [ Links ]
Chaparro, J. C. (2011). Evidence of tsunami events in the Paleolimnological record of Lake Pátzcuaro, Michoacán, Mexico. Geofísica Internacional, 50, 147-161. [ Links ]
Hartmann, G. (1959). Beitrag zur Kenntnis des Nicaragua-Sees unter besonderer Berücksichtigung seiner Ostracoden. Zoologischer Anzeiger, 162, 269-294. [ Links ]
Hernández, M., Escobar, E., & Alcocer, J. (2010). Benthic crustaceans assemblage in a tropical, saline lake. Revista Mexicana de Biodiversidad, 81, 133-140. [ Links ]
Horne, D. J. (2007). A mutual temperature range method for Quaternary palaeoclimatic analysis using European nonmarine Ostracoda. Quaternary Science Reviews, 26, 1398-1415. [ Links ]
Juárez, D. (2005). Registro de cambios paleoambientales en sedimentos del lago “La Preciosa”, Puebla, con base en el estudio de ostrácodos (Tesis de maestría). Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
Karanovic, I. (2012). Recent freshwater ostracods of the world. Berlin Heidelberg, Germany: Springer. [ Links ]
Kazmierczak, J., Kempe, S., Kremer, B., López-García, P., Moreira, D., & Tavera, R. (2011). Hydrochemistry and microbialites of the alkaline crater lake Alchichica, Mexico. Facies, 57, 543-570. [ Links ]
Keyser, D. (1976). Zur Kenntnis der brackigen mangrovebewachsenen Weichböden Südwest-Floridas unter besonderer Berücksitchtigung ihrer Ostracodenfauna. Doktorarbeit. Hamburg, Germany: Fachbereich Biologie, Universität Hamburg. [ Links ]
Keyser, D., & Walter, R. (2004). Calcification in ostracodes. Revista Española de Micropaleontología, 36, 1-11. [ Links ]
Külköylüoglu, O., & Vinyard, G. L. (2000). Distribution and ecology of freshwater Ostracoda (Crustacea) collected from springs of Nevada, Idaho, and Oregon: a preliminary study. Western North American Naturalist, 60, 291-303. [ Links ]
Külköylüoglu, O. (2004). On the usage of ostracods (Crustacea) as bioindicator species in different aquatic habitats in the Bolu region, Turkey. Ecological Indicators, 4, 139-147. [ Links ]
Li, X., Liu, W., Zhang, L., & Sun, Z. (2010). Distribution of Recent ostracod species in the Lake Qinghai area in northwestern China and its ecological significance. Ecological Indicators, 10, 880-890. [ Links ]
Lorenschat, J. (2009). Ostracoden aus Hoch- und Tieflandseen in südlichen Guatemala als Bioindikatoren. Diplomarbeit. Braunschweig, Germany: Institut für Umweltgeologie, Technische Universität Braunschweig. [ Links ]
Machain-Castillo, M. L., & Gío-Argáez, R. (2004). Ostrácodos bentónicos del sur del Golfo de México. In M. Caso, I. Pisanty, & R. Ezcurra (Eds.), Diagnóstico ambiental del Golfo de México (pp. 161-171). México: Instituto Nacional de Ecología (INE-SEMARNAT). [ Links ]
Maddocks, R. F., Machain-Castillo, M. L., & Gío-Argáez, R. (2009). Podocopan Ostracoda (Crustacea) of the Gulf of Mexico. In D. L. Felder & D. K. Camp, (Eds.), Gulf of Mexico-Origins, Water and Biota. Biodiversity (pp. 877-894). Texas, USA: Texas A & M Press, College. [ Links ]
Marmonier, P., & Chatelliers, M. C. D. (1992). Biogeography of the Benthic and Interstitial Living Ostracods (Crustacea) of the Rhone River (France). Journal of Biogeography, 19, 693-704. [ Links ]
Martens, K. (1996). On Korannocythere gen. nov. (Crustacea, Ostracoda), a new genus of temporary pool limnocytherids from southern Africa, with the description of three new species and a generic reassessment of the Limnocytherinae. Royal Sciences Naturelles de Belgique Biology, 66. [ Links ]
Martens, K., & Schön, I. (1999). Crustacean Biodiversity in Ancient Lakes: A Review. Crustaceana, 72, 899-910. [ Links ]
Martens, K. (2000). Factors affecting the divergence of mate recognition systems in the Limnocytherinae (Crustacea, Ostracoda). Hydrobiologia, 419, 83-101. [ Links ]
Meisch, C. (2000). Freshwater Ostracoda of western and central Europe. Heidelberg, Berlin, Germany: Spektrum Akademischer Verlag GmbH. [ Links ]
Metcalfe, S. E., Davies, S. J., Braisby, J. D., Leng, M. J., Newton, A. J., Terrett, N. L., & O’Hara, S. L. (2007). Long and short-term change in the Pátzcuaro Basin, central Mexico. Palaeogeography, Palaeoclimatology, Palaeoecology, 247, 272-295. [ Links ]
Mezquita, F., Roca, J. R., Reed, J. M., & Wansard, G. (2005). Quantifying species-environment relationships in non-marine Ostracoda for ecological and palaeoecological studies: examples using Iberian data. Palaeogeography, Palaeoclimatology, Palaeoecology, 225, 93-117. [ Links ]
Mischke, S., Almogi-Labin, A., Ortal, R., Rosenfeld, A., Schwab, M. J., & Boomer, I. (2010). Quantitative reconstruction of lake conductivity in the Quaternary of the Near East (Israel) using ostracods. Journal of Paleolimnology, 43, 667-688. [ Links ]
Nagorskaya, L. (2002). Living freshwater Ostracoda (Crustacea) of a floodplain relict oak forest (Polesye, Belarus). Revista Española de Micropaleontología, 36, 135-145. [ Links ]
Nagorskaya, L., & Keyser, D. (2005). Habitat diversity and ostracod distribution patterns in Belarus. Hydrobiologia, 538, 167-178. [ Links ]
Oliva, M. G., Lugo, A., Alcocer, J., & Cantoral-Uriza, E. A. (2006). Cyclotella alchichicana sp. nov. from a saline mexican lake. Diatom Research, 21, 81-89. [ Links ]
Palacios Fest, M., Cohen, A., Ruiz, J., & Blank, B. (1993). Comparative paleoclimatic interpretations from nonmarine ostracodes using faunal assemblages, trace elements, shell chemistry and stable isotope data. Geophysical Monograph, 78, 179-189. [ Links ]
Palacios-Fest, M. R., & Dettman, D. L. (2001). Temperature controls monthly variation in ostracode valve Mg/Ca: Cypridopsys vidua from a small lake in Sonora, Mexico. Geochimica et Cosmochimica Acta, 65, 2499-2507. [ Links ]
Palacios-Fest, M., Carreño, A., Ortega-Ramírez, J., & Alvarado-Valdéz, G. (2002). A paleoenvironmental reconstruction of Laguna Babícora, Chihuahua, Mexico based on ostracode paleoecology and trace element shell chemistry. Journal of Paleolimnology, 27, 185-206. [ Links ]
Palacios-Fest, M. (2004). Ostracode paleoecology of Red Mountain Site, Mesa, Arizona. In Life on the Lehi Terrace: the Archaeology of the Red Mountain Freeway between Country Club Drive and Gilbert Road. USA: Arizona Department of Transportation Environmental Planning Group, ADOT Project RAM 600-8-326, TRACS 202L-MA20-H3872-01D, 737-750. [ Links ]
Palacios-Fest, M. (2010). Late Holocene paleoenvironmental history of the upper west Amarillo Creek Valley at Archaeological Site 41PT185/C, Texas, USA. Boletín de la Sociedad Geológica Mexicana, 62, 399-436. [ Links ]
Pérez, L. (2010). Non-marine ostracodes from the Yucatán Peninsula as late Quaternary paleoenvironmental indicators (Doctoral thesis). Architektur, Bauingenieurwesen und Umweltwissenschaften, Technische Universität Braunschweig, Braunschweig, Germany. [ Links ]
Pérez, L., Bugja, R., Massaferro, J., Steeb, P., van Geldern, R., Frenzel, P., Brenner, M., Scharf, B., & Schwalb, A. (2010a). Post-Columbian environmental history of Lago Petén Itzá, Guatemala. Revista Mexicana de Ciencias Geológicas, 27, 490-507. [ Links ]
Pérez, L., Lorenschat, J., Brenner, M., Scharf, B., & Schwalb, A. (2010b). Extant freshwater ostracodes (Crustacea: Ostracoda) from Lago Petén Itzá, Guatemala. Revista de Biología Tropical, 58, 871-895. [ Links ]
Pérez, L., Frenzel, P., Brenner, M., Escobar, E., Hoelzmann, P., Scharf, B., & Schwalb, A. (2011). Late Quaternary (24-10 ka BP) environmental history of the Neotropical lowlands inferred from ostracodes in sediments of Lago Petén Itzá, Guatemala. Journal of Paleolimnology, 46, 59-74. [ Links ]
Pérez, L., Lorenschat, J., Brenner, M., Scharf, B., & Schwalb, A. (2012). Non-marine ostracodes (Crustacea) of Guatemala. In E. Cano & J. Shuster (Eds.), Biodiversidad de Guatemala II (pp. 121-131). Guatemala: Universidad del Valle de Guatemala. [ Links ]
Pérez, L., Lorenschat, J., Massaferro, J., Pailles, C., Sylvestre, F., Hollwedel, W., Brandorff, G.-O. , Brenner, M., Islebe, G., Lozano, S., Scharf, B., & Schwalb, A. (2013). Bioindicators of climate and trophic state in lowland and highland aquatic ecosystems of the Northern Neotropics. Revista de Biología Tropical, 61, 603-644. [ Links ]
Quiroz Castelán, H., Díaz Vargas, M., Trejo Albarrán, R., & Elizalde Arriaga, E. (2000). Aspectos sobre la abundancia y distribución de los principales grupos de la fauna bentónica en el lago “Zempoala”, Morelos, México. Ciencia y mar, 4, 39-50. [ Links ]
Ramírez-García, P., & Vázquez-Gutiérrez, F. (1989). Contribuciones al estudio limnobotánico de la zona litoral de seis lagos cráter del Estado de Puebla. Anales del Instituto de Ciencias del Mar y Limnología, 323, 1-16. [ Links ]
Ramsar. (2013). Humedales de México. Retrieved from http://ramsar.conanp.gob.mx/lsr.php [ Links ]
Rio de la Loza, L., & Craveri, C. (1858). Opúsculo sobre los Pozos Artesianos y las Aguas Naturales de más uso en la Ciudad de México, con algunas noticias relativas al Corte Geológico del Valle, y una lista de las Plantas que vegetan en las inmediaciones del Desierto Viejo. Boletín de la Sociedad Mexicana de Geografía y Estadística, 6, 9-28. [ Links ]
Rodríguez, G. (2002). Biodiversidad de los crustáceos dulceacuícolas del centro de Nuevo León y noroeste de Tamaulipas (Reporte final de proyecto S104, CONABIO). Departamento de Zoología de Invertebrados, Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, Nuevo León, México. Retrived from http://www.conabio.gob.mx/institucion/cgi-bin/datos.cgi?Letras=S & Numero=104 [ Links ]
Rodríguez, E. (2011). Estudio morfológico de ostrácodos dulceacuícolas de Durango y Veracruz, México (Tesis de licenciatura). Facultad de Ciencias, Universidad Nacional Autónoma de México, Distrito Federal, México. [ Links ]
Roy, P. D., Caballero, M., Lozano, R., & Smykatz-Kloss, W. (2008). Geochemistry of late quaternary sediments from Tecocomulco lake, central Mexico: Implication to chemical weathering and provenance. Chemie der Erde - Geochemistry, 68, 383-393. [ Links ]
Roy, P. D., Caballero, M., Lozano, R., Pi, T., & Morton, O. (2009). Late Pleistocene-Holocene geochemical history inferred from Lake Tecocomulco sediments, Bain of Mexico, Mexico. Geochemical Journal, 43, 49-64. [ Links ]
Saldivar-López, D. B. (2011). Platelmintos y crustáceos presentes en la vegetación acuática, sus condiciones de hábitat y sus relaciones interespecíficas, en la Presa de Apizaquito (Tesis de licenciatura). División de Ciencias Biológicas y de la Salud, Departamento El Hombre y su ambiente, Universidad Autónoma Metropolitana, Unidad Xochimilco, México. [ Links ]
Sars, G. (1901). Contributions to the knowledge of the fresh-water Entomostraca of South America as shown by artificial hatching from dried material. Part II. Copepoda-Ostracoda. Archiv fur Mathematik og Naturvidenskab, 24, 1-52. [ Links ]
Smith, A. J. (1993). Lacustrine ostracodes as hydrochemical indicators in lakes of the north-central United States. Journal of Paleolimnology, 8, 121-134. [ Links ]
Smith, A. J., & Delorme, L. D. (2010). Ostracoda. In J. Thorp & A. Covich (Eds.), Ecology and Classification of North American Freshwater Invertebrates (pp. 725-771). United Kingdom:Academic Press, Elsevier. [ Links ]
Soria Caballero, D. C. (2010). Estudio bioestratigráfico y mineralógico de la ciénega de Zacapu, Michoacán (Tesis de maestría). Instituto de Investigaciones Metalúrgicas, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, México. [ Links ]
Staplin, F. L. (1963). Subfamilies Cyclocyprinae, Cypridopsinae, Ilyocyprinae, families Darwinulidae and Cytheridae. Stratigraphic ranges and assemblage patters. Journal of Paleontology, 37, 1164-1203. [ Links ]
Suter, M. (2004). A neotectonic-geomorphologic investigation of the prehistoric rock avalanche damming Laguna de Metztitlán (Hidalgo State, East-central Mexico). Revista Mexicana de Ciencias Geológicas, 21, 397-411. [ Links ]
Szlauer-Lukaszewska, A. (2012). Ostracod assemblages in relation to littoral plant communities of a shallow lake (Lake Swidwie, Poland). International Review of Hydrobiology, 97, 262-275. [ Links ]
Tressler, W. L. (1949). Fresh-water Ostracoda from Brazil. Proceeding of the United States National Museum, 100, 61-83. [ Links ]
Tressler, W. L. (1954). Fresh-water Ostracoda from Texas and Mexico. Journal of Washington Academy Science, 44, 138-149. [ Links ]
Vázquez, G. V., Ortega, B., Rodríguez, A., Caballero, M., & Lozano, S. (2008). Mineralogía magnética como indicador de sequía en los sedimentos lacustres de los últimos ca. 2,600 años de Santa María del Oro, occidente de México. Revista Mexicana de Ciencias Geológicas, 25, 21-38. [ Links ]
Viehberg, F. A., & Mesquita-Joanes, F. (2012). Chapter 4 - Quantitative Transfer Function Approaches in Palaeoclimatic Reconstruction Using Quaternary Ostracods. In D. J. Horne, J. A. Holmes, J. Rodriguez-Lazaro, & F. Viehberg (Eds.), Developments in Quaternary Sciences (pp. 47-64). Amsterdam, Netherlands: Elsevier. [ Links ]
Wetterich, S. (2008). Freshwater ostracods as bioindicators in Arctic periglacial regions. Thesis. Potsdam, Germany: Mathematisch-Naturwissenschaftlichen Fakultät Universität Potsdam. [ Links ]
Zara-environmental-LCC. (2010). Final report for deep aquifer biota study of the Edwards Aquifer. Texas, USA: Zara-environmental-LCC. [ Links ]
1. Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510, Distrito Federal, México; lcpereza@geologia.unam.mx, mslozano@unam.mx
2. Instituto de Geofísica, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510, Distrito Federal, México; maga@geofisica.unam.mx
Received 21-VII-2014. Corrected 10-X-2014. Accepted 14-XI-2014.


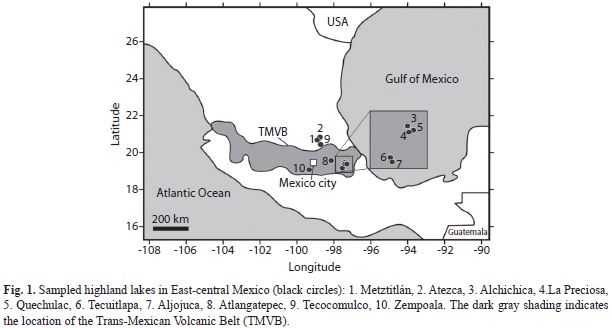{kind=link}
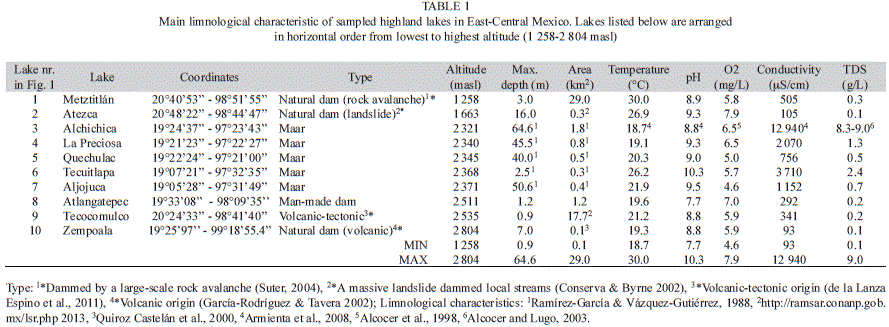{kind=link}
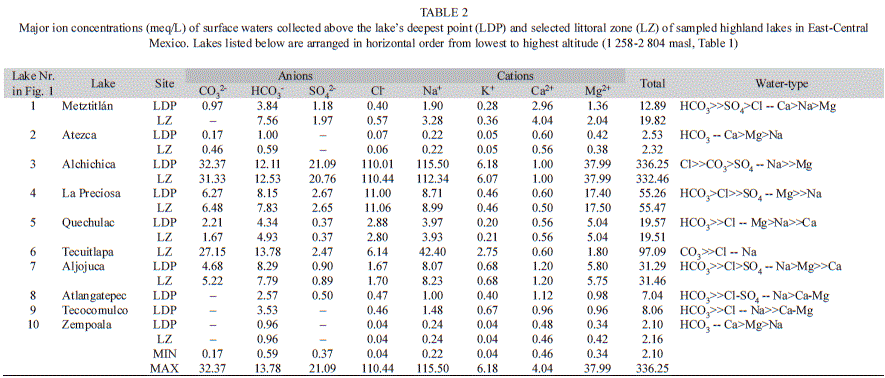{kind=link}
{kind=link}
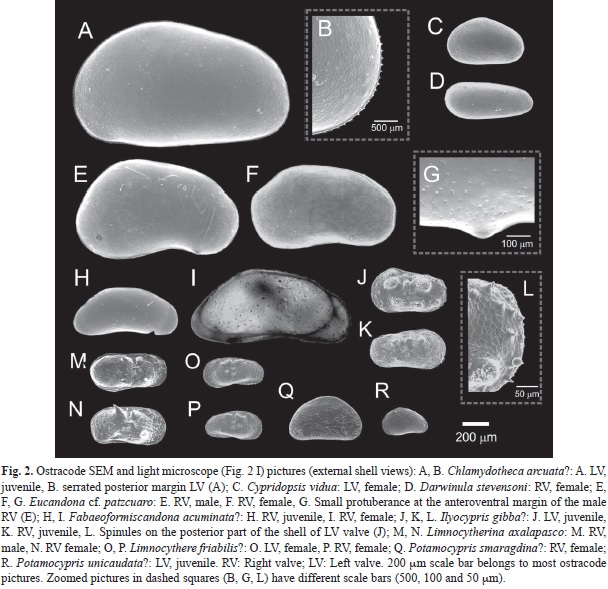{kind=link}
{kind=link}
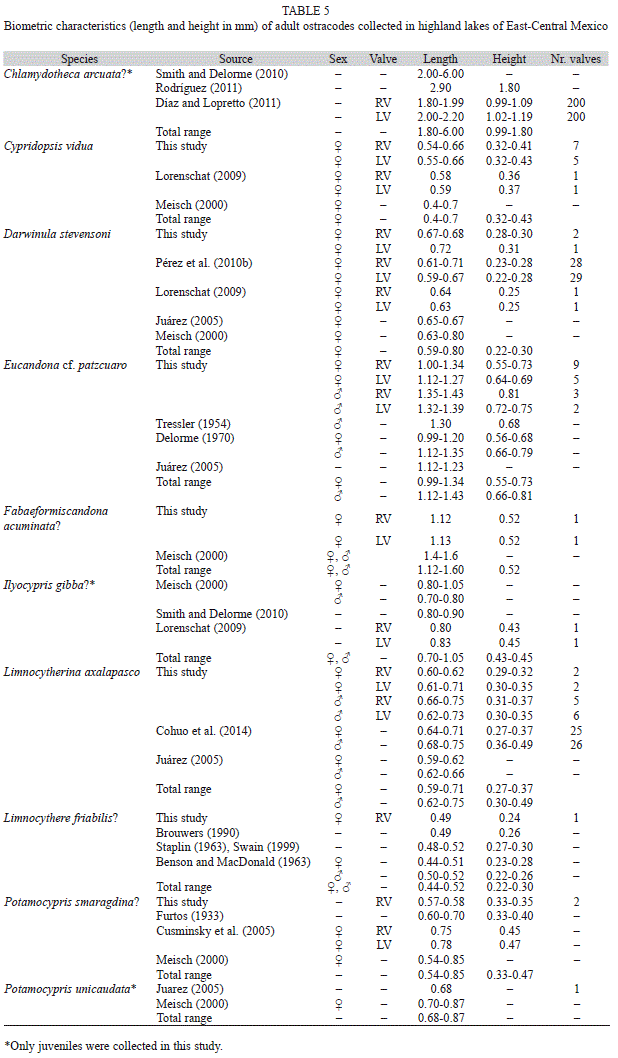{kind=link}