










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.2 San José Apr./Jun. 2015
Inversión reproductiva de Aratus pisonii (Decapoda: Sesarmidae): diferencias entre hábitats y análisis de rutas
Reproductive investment of Aratus pisonii (Decapoda: Sesarmidae): differences among habitats and path analysis
Reproductive investment of Aratus pisonii (Decapoda: Sesarmidae): differences among habitats and path analysis
Abstract
Aratus pisonii is an abundant Neotropical mangrove crab that forages on fresh leaf tissue in the canopy and lives in a wide spectrum of habitats. It presents great plasticity in some aspects of its life history becoming an excellent subject for comparative studies. The purpose of this study was to investigate possible reproductive variations among populations of this crab in mangroves with different levels of structural development, and explore the direct and indirect effects of environmental variables on reproductive investment. The study was conducted during the rainy season of 2003 and 2004, in North Western Venezuela. Fecundity (number of eggs/female), dry weight of egg mass (PSH) and egg quality (C/Neggs) were evaluated in five populations of this species, in mangroves under different environmental conditions (estuarine, marine, hypersaline) and diverse degrees of structural development (arboreal, arbustive, scrub). Reproductive output (Ro) was obtained by the ratio Ro=PSH/dry body weight (PSC); other estimations of Ro were tested: Ro1=PSH/carapace length (CL), Ro2=fecundity/CL. A path analysis was used to explore causal relationships between environmental and morphological variables and reproductive investment (PSH, fecundity). Fecundity ranged from 6 463±831 eggs (Mean±SE) in the hypersaline arbustive mangrove to a maximum of 24 584±2 393 eggs in estuarine arboreal mangrove, which presents the greatest structural development. The reproductive output of females showed significant differences among mangroves, with the highest values of Ro1, Ro2 in the estuarine mangrove females. The results of the path analysis indicated a multicausal effect of morphological and environmental variables; in particular, the salinity showed the greatest influence on fecundity and PSH. The condition of the estuarine mangrove forests with lower salinity and higher structural development involving higher quality of mangrove leaves and a wider range of potential resources, may explain the higher reproductive output of females observed in this habitat. Rev. Biol. Trop. 63 (2): 385-399. Epub 2015 June 01.
Key words: Crustacea, fecundity, reproductive output, elemental composition of eggs, path analysis, mangrove, Venezuela.
Resumen
Aratus pisonii es un cangrejo abundante de los manglares del Neotrópico, que forrajea sobre el tejido foliar fresco del dosel y vive en una amplia variedad de hábitats. Presenta una gran plasticidad en algunos aspectos de su historia de vida, convirtiéndose en un excelente sujeto para estudios comparativos. El propósito de este estudio fue investigar las posibles variaciones reproductivas entre las poblaciones de este cangrejo en manglares con diferentes niveles de desarrollo estructural, y explorar los efectos directos e indirectos de las variables ambientales sobre la inversión reproductiva. El estudio se realizó durante la temporada de lluvias 2003 y 2004, en la región Noroccidental de Venezuela. Se evaluaron la fecundidad (número de huevos/hembra), el peso seco de la masa de huevos (PSH) y la calidad del huevo (C/Nhuevos) en cinco poblaciones de esta especie, que viven en manglares bajo diferentes condiciones ambientales (estuarinos, marinos, hipersalinos) y diferente grados de desarrollo estructural (arbóreo, arbustivo, achaparrado). El rendimiento reproductivo (Ro) se obtuvo por la relación Ro=PSH/peso seco del cuerpo (PSC); se probaron otras estimaciones de Ro: Ro1=PSH/longitud del caparazón (LC), Ro2=fecundidad/LC. Se utilizó un análisis de rutas para explorar las relaciones causales entre las variables ambientales y morfológicas con la inversión reproductiva (PSH, fecundidad). La fecundidad varió de 6 463±831 huevos (media±ES) en el manglar arbustivo hipersalino, hasta un máximo de 24 584±2 393 huevos en el manglar arbóreo estuarino, que presentó el mayor desarrollo estructural. El rendimiento reproductivo de las hembras mostró diferencias significativas entre manglares, encontrándose los más altos valores de Ro1, Ro2 en las hembras de manglar estuarino. Los resultados del análisis de rutas indicaron un efecto multicausal de variables morfológicas y ambientales; en particular, la salinidad mostró la mayor influencia sobre la fecundidad y el PSH. La condición del manglar estuarino con menor salinidad y mayor desarrollo estructural, que de acuerdo con estudios anteriores supone una mayor calidad de las hojas de mangle y una mayor variedad de recursos potenciales, puede explicar la mayor inversión reproductiva observada en las hembras de este hábitat.
Palabras clave: Crustacea, fecundidad, rendimiento reproductivo, composición elemental de los huevos, análisis de rutas, manglar, Venezuela.
Existe una gran diversidad de patrones de historias de vida en los crustáceos, que se expresan en diferentes estrategias reproductivas y de crecimiento, en los cangrejos Brachyura se encuentran muchos de estos ejemplos; además se asume, generalmente, que estos patrones son adaptativos, moldeados por presiones evolutivas que seleccionan la máxima sobrevivencia de la descendencia a la siguiente generación (Hartnoll, & Gould, 1988). La variabilidad de patrones de historia de vida en crustáceos decápodos ocurre en respuesta a variaciones de las condiciones ambientales (Conde, & Díaz, 1989b; Mashiko, 1990; Hancock, Hughes, & Bunn, 1998) y por efectos estacionales (Díaz, 1980; Ouellet, & Plante, 2004; Bas, Spivak, & Anger, 2007).
En los crustáceos decápodos, los aspectos reproductivos mejor estudiados han sido la fecundidad y el esfuerzo reproductivo (Hines, 1982, 1991; López, Jeri, González, & Rodríguez, 1997; Castiglioni, Santos, Reigada, & Negreiros-Fransozo, 2004; Lima, Soares, & Oshiro, 2006; Bas et al., 2007). La fecundidad, tradicionalmente definida como el número de huevos liberados por una hembra por desove, o durante un periodo determinado durante su ciclo de vida (Hernáez, 2001), constituye una información valiosa la cual permite estimar el potencial reproductivo de una población y realizar estudios comparativos. El esfuerzo reproductivo usualmente definido como la fracción de la energía que un organismo destina a la reproducción (López et al., 1997), puede expresarse como rendimiento reproductivo (Ro) que expresa la relación entre el peso seco de la masa de huevos y del cuerpo de la hembra (Clarke, Hopkins, & Nilssen, 1991). El conocer estos aspectos de la historia de vida de una especie ayuda en la comprensión de su estrategia reproductiva ante las variaciones del ambiente (Sastry, 1970; Stearns, 1976).
Aratus pisonii (Milne-Edwards, 1837) es un cangrejo arborícola que se alimenta principalmente de hojas vivas de mangle, este presenta una amplia distribución en la costa atlántica encontrándose desde Florida hasta Brasil (Hartnoll, 1965; Chance, & Hobss, 1969; Conde, et al., 2000). Se ha evidenciado recientemente, que las poblaciones de la vertiente del Pacífico pertenecen a una especie hermana A. pacificus Thiercelin & Schubart, 2014. A. pisonii habita en una amplia gama de ambientes, desde zonas estuarinas, pasando por ambientes marinos, hasta lagunas hipersalinas, en los cuales se asocia a varias especies de manglar: Rhizophora mangle L., Laguncularia racemosa (L.) Gaertn.f., Avicennia germinans (L.) Stearn (Conde, & Díaz, 1989a, b; Díaz, & Conde, 1989; Conde, 1990; Conde, et al., 2000; Erickson, Saltis, Bell, & Dawes, 2003; Feller, & Chamberlain, 2007; Colpo, Chacur, Guimarães, & Negreiros-Fransozo, 2011).
Esta especie presenta una gran plasticidad en algunos aspectos de su historia de vida y en la dinámica de sus poblaciones (Warner, 1967; Conde, & Díaz, 1989b, 1992a; Conde, Díaz, & Rodríguez, 1989), lo cual la convierte en un interesante sujeto para estudios comparativos. En Venezuela se ha evidenciado que el tamaño corporal al inicio de la madurez sexual varía ampliamente entre diferentes ambientes (Conde, & Díaz, 1992a), al igual que el porcentaje de hembras ovadas, lo cual puede estar asociado a la productividad del hábitat, inferida a partir de la estructura del bosque de manglar (Conde, & Díaz, 1989b; Snedaker, 1989). En particular se ha encontrado que la talla y el porcentaje de hembras ovadas incrementan desde manglares arbustivos y achaparrados, localizados en condiciones hipersalinas, hasta una talla y porcentaje de hembras ovadas máximo observado en manglares estuarinos con mayor altura de los árboles (Conde, & Díaz, 1989b; 1992a, 1992b; Conde et al., 1989). Estudios posteriores han corroborado la existencia de un gradiente de estructura, en un grupo de cinco rodales de manglar localizados en el estado Falcón, los cuales están bajo diferentes condiciones de salinidad, entre otras variables ambientales; dicho gradiente se caracteriza por un progresivo decrecimiento del área basal y altura de los árboles, desde un manglar arbóreo estuarino hasta un manglar achaparrado hipersalino (López, Barreto, & Conde, 2011).
El objetivo de este trabajo fue determinar las diferencias en la inversión reproductiva entre poblaciones de Aratus pisonii, específicamente evaluar si la fecundidad, el rendimiento reproductivo (Ro) y la calidad de los huevos difieren entre manglares con distintos grados de desarrollo estructural, y explorar los efectos directos e indirectos de las variables ambientales sobre la inversión reproductiva.
Materiales y métodos
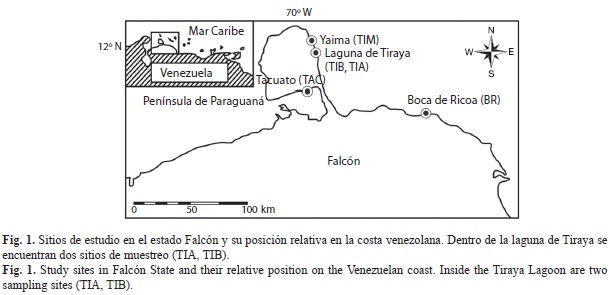
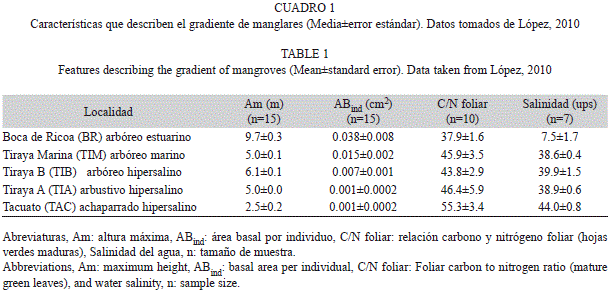
Área de estudio: Se seleccionaron cinco bosques de manglar en la región noroccidental de Venezuela, los cuales conforman un gradiente de complejidad estructural caracterizado por un progresivo decrecimiento del área basal y altura de los árboles, desde un manglar arbóreo estuarino hasta un manglar achaparrado hipersalino (López et al., 2011) (Fig. 1). Para más detalle sobre el gradiente y las características de los hábitats ver López et al. (2011). El manglar con mayor desarrollo estructural se encuentra en la desembocadura del río Ricoa, denominado Boca de Ricoa (BR, 11°30’11” N - 69°12’27” W), cuyas aguas son salobres, el cual presentó las máximas alturas y área basal del grupo de manglares evaluado (Cuadro 1) y donde predominó la especie Laguncularía racemosa. El resto de los manglares se ubican en la península de Paraguaná; estos son un manglar arbóreo marino denominado Tiraya Marina (TIM)(12°03’10” N - 69°51’9” W); otros dos manglares, uno arbóreo y otro arbustivo, TIB (12°01’53” N - 69°51’15” W) y TIA (12°02’05” N - 9°50’48” W) respectivamente, ambos localizados en la laguna hipersalina de Tiraya. La quinta localidad se sitúa al sur de la península de Paraguaná en la laguna hipersalina de Tacuato (TAC, 11°41’21” N - 69°49’58” W), donde se encuentra un manglar achaparrado con la menor altura de todos (Cuadro 1). En los manglares localizados en la península de Paraguaná, Rhizophora mangle fue la especie dominante (López et al., 2011). La precipitación en la desembocadura del río Ricoa presenta dos pulsos al año, uno durante el período de lluvias (octubre-diciembre) y otro menor entre mayo y julio; mientras que en la península de Paraguaná existe una estacionalidad muy marcada, con un periodo de sequía que transcurre entre enero y septiembre, y un periodo lluvioso entre octubre y diciembre (Conde, 1990).
Morfometría, fecundidad y rendimiento reproductivo: En 2003 y 2004, alrededor de 30 hembras ovadas de A. pisonii fueron recolectadas manualmente en cada localidad y año, durante los meses de agosto, septiembre y noviembre, que corresponden a los periodos de transición (agosto) y lluvias (septiembre, noviembre), lo cual aseguraba la presencia de hembras ovadas en todos los hábitats. Cada hembra fue colocada en bolsas separadas para evitar la pérdida de huevos y se preservó en frío (-20°C). Con el fin de minimizar el error relacionado con la pérdida de huevos durante la incubación, del grupo total de hembras por localidad, se seleccionaron aquellas hembras recién ovadas (13-15/localidad) que portaran huevos sin indicios de desarrollo embrionario (Hartnoll, 2006). Se midió el ancho (AC) y largo del caparazón (LC) de cada una, con un vernier de 0.01mm de precisión. Luego se retiró la masa de huevos y se estimó la fecundidad de acuerdo al método volumétrico propuesto por Díaz, Conde, & Bevilacqua (1983), para lo cual, un grupo de 30 huevos fueron inmersos en agua destilada y su diámetro (d) determinado bajo un microscopio estereoscópico; el volumen de cada huevo fue calculado según la fórmula de una esfera V=(4/3) p(d/2)3 (mm3), posteriormente se devolvían los huevos a su masa original y se calculaba el volumen desplazado por la masa de huevos según el método volumétrico (Díaz et al., 1983), con estos datos se obtenía la fecundidad total por hembra.
La masa de huevos y el cuerpo de las hembras fueron secados por separado a 60ºC, hasta obtener un peso constante, luego por medio de una balanza con precisión de 0.1mg se obtuvo el peso seco de la masa de huevos (PSH) y del cuerpo de cada hembra (PSC). El rendimiento reproductivo (Ro) se consiguió mediante la relación señalada por Clarke et al. (1991), donde Ro=PSH/PSC. Se probaron otras estimaciones de Ro empleando el largo del caparazón (LC) como denominador, con el fin de ponderar las variables reproductivas por el tamaño del cangrejo; así se construyeron Ro1=PSH/LC y Ro2=fecundidad/LC.
Materia orgánica y análisis elemental de los huevos y caparazón: Se determinó el porcentaje de materia orgánica de los huevos (%MOhuevos) mediante incineración a 550°C por cuatro horas, sustrayendo el peso de las cenizas del peso seco de los huevos. El contenido de carbono y nitrógeno de los huevos, y del caparazón fue determinado en una submuestra de ±10mg, mediante un analizador elemental Fisons EA 1108-CHNS-O; el tamaño de la muestra fue menor para estos análisis, debido a que la mayor parte del PSH fue empleado para determinar %MOhuevos. Los resultados fueron expresados en porcentaje %C y %N (peso/peso).
Análisis de los resultados: Con el fin de detectar diferencias en las variables biométricas (LC, AC, PSC), reproductivas (PSH, Fecundidad, Vol/huevo, Ro, Ro1, Ro2) y calidad de los huevos (%MOhuevos, C/Nhuevos) entre los diferentes manglares evaluados, se realizaron análisis de varianza de un factor (ANDEVA, factor: localidad). Se comprobaron los supuestos de este análisis (Sokal, & Rohlf, 1995). Cuando no se logró comprobar la homocedasticidad de varianzas, se empleó el test robusto de igualdad de medias Brown-Forsythe (Brown, & Forsythe, 1974). Se emplearon las pruebas a posteriori, Tukey HSD o Games-Howell, ésta última cuando no se cumplía con el supuesto de homosedasticidad. Estos análisis se realizaron utilizando los programas SPSS (1999).
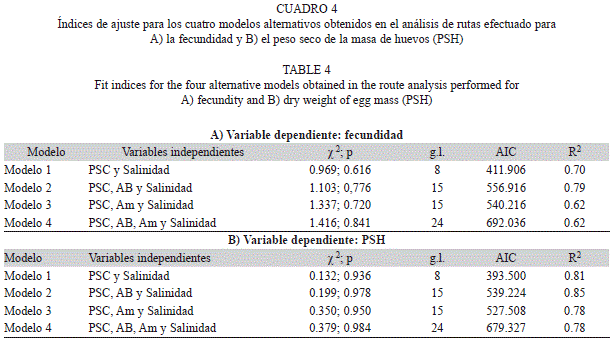
Con el fin de evaluar los efectos de la variabilidad ambiental sobre las características reproductivas de A. pisonii, se aplicó un análisis de rutas (Wright, 1934), englobando las cinco poblaciones de cangrejos examinadas para determinar relaciones multicausales (directas e indirectas), que se establecen entre las variables ambientales y la inversión reproductiva (fecundidad y PSH). En la etapa de especificación de los modelos se incluyeron además de las variables ambientales (área basal, altura máxima, salinidad del agua y C/N foliar), las biométricas (LC, AC, PSC) puesto que son determinantes de la salida reproductiva en cangrejos braquiuros (Hines, 1982); luego se realizaron análisis de regresión múltiple, seleccionando aquellas relaciones con pertinencia biológica (Ortiz-Pulido, 2000). El criterio para seleccionar variables fue comprobar la existencia de significación estadística con las variables respuesta (fecundidad, PSH); adicionalmente las variables colineales fueron excluidas (r>0.9). La identificación de los modelos se realizó de acuerdo a los grados de libertad de éstos, para lo cual se requiere un número positivo de grados de libertad (gl>0), lo que indica que existe suficiente información para su contrastación (Pérez, Medrano, & Sánchez, 2013). Se utilizó la técnica de iteración bootstrap y se estimaron los coeficientes de ruta por máxima verosimilitud, luego de estandarizar todas las variables; se realizó una prueba de bondad de ajuste (χ2) entre la matriz de las relaciones estimadas por el modelo y la matriz de datos observados, esta prueba debe ofrecer un p>0.05 para aceptar el modelo (Ortiz-Pulido, 2000). Finalmente la comparación y escogencia del mejor modelo se obtuvo a partir de la varianza explicada por cada uno (R2) y utilizando el Criterio de Información de Akaike (AIC) (Akaike, 1987). Este criterio plantea que valores inferiores de AIC indican un mejor ajuste respecto a los modelos alternativos (Pérez et al., 2013). Para realizar el análisis de rutas se empleó el programa Ωnyx versión 0.9-729 (von Oertzen, Brandmaier, & Tsang, 2013).
Resultados
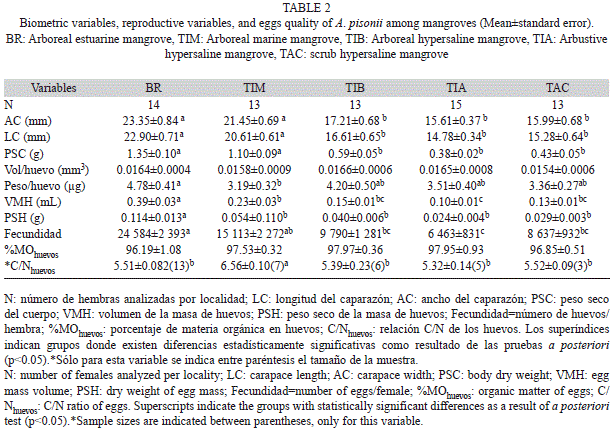
Variables reproductivas y su relación con variables biométricas: En un total de 68 hembras analizadas, la fecundidad promedio fluctuó entre un mínimo de 6 463±832 huevos (media±error estándar) en hembras del manglar arbustivo hipersalino (TIA), y un máximo de 24 584±2 394 huevos en el manglar estuarino (BR) con mayor desarrollo estructural (Cuadro 2). La fecundidad individual fluctuó entre 2 376 huevos/hembra (TIA, AC=17.0mm) hasta un máximo de 36 950 huevos/hembra (BR, AC=26.2mm). El intervalo de tallas evaluado osciló entre 12.0mm de AC en una hembra del manglar achaparrado hipersalino (TAC) y una talla máxima de 27.5mm de AC en el manglar estuarino con mayor desarrollo estructural (BR).
Se comprobó que no existe variación temporal entre años, en la fecundidad y el PSH (t-Student, gl=66, fecundidad: t=0.32, p=0.75; PSH: t=0.43, p=0.67), por lo tanto se agruparon los datos por sector independiente del año para el resto de los análisis. Se encontraron diferencias significativas para todas las variables biométricas entre localidades (ANDEVA, Cuadro 3), observando una disminución del tamaño corporal (talla, peso) desde el manglar arbóreo estuarino de Boca de Ricoa hasta el manglar achaparrado hipersalino de Tacuato (Cuadro 2).
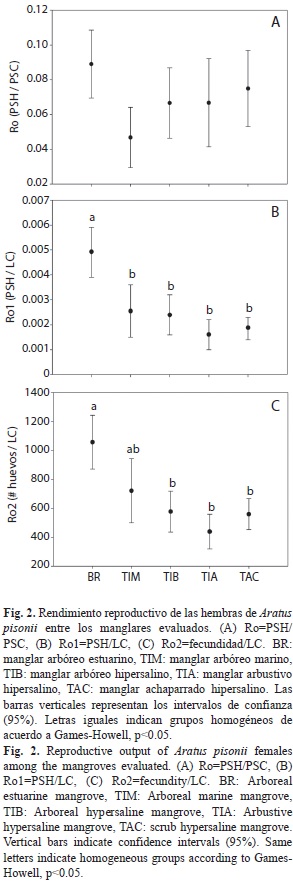
El rendimiento reproductivo estimado de forma clásica (Ro=PSH/PSC) no presentó diferencias significativas entre los manglares examinados (Fig. 2A). Sin embargo, los Ro propuestos en este trabajo (Ro1, Ro2), sí mostraron diferencias significativas (Cuadro 3). Se observó un Ro1 (PSH/LC) significativamente superior en hembras del manglar estuarino con mayor desarrollo estructural (BR), respecto al resto de los manglares en los cuales las hembras presentaron un Ro1 que no se diferenció (Fig. 2B). Mientras que el Ro2 (fecundidad/LC) en hembras del manglar estuarino fue significativamente superior al observado en hembras de los hábitats hipersalinos (TIB, TIA, TAC), pero no se diferenció del observado en aquellas que habitan el manglar marino (TIM) (Fig. 2C).
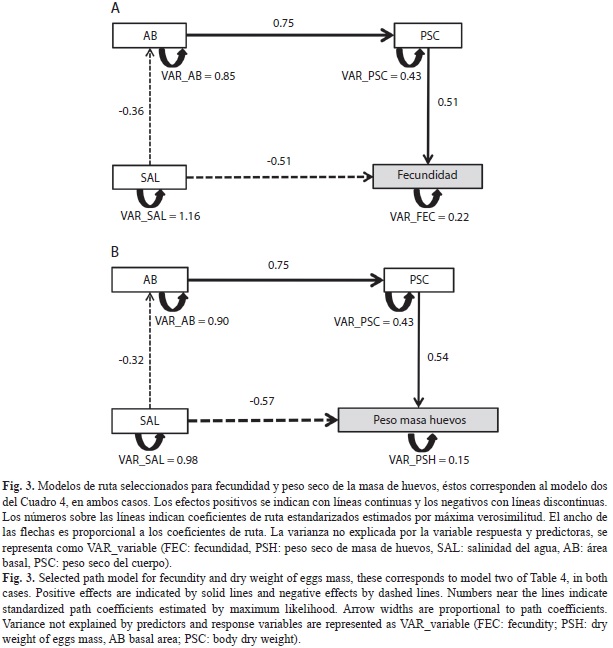
Análisis de rutas: Del total de modelos construidos para evaluar el efecto de las variables morfológicas y ambientales sobre la inversión reproductiva (fecundidad, PSH), se identificaron cuatro de ellos con significación estadística (gl>0; χ2>0.05) y con validez biológica. Se escogió el modelo dos para las dos variables respuesta (Cuadro 4), en los cuales están presentes las variables predictoras: PSC, AB y salinidad (Fig. 3 A y Fig. 3B). Estos modelos explicaron una mayor variación tanto para la fecundidad como para el PSH (R2=0.79 y 0.85 respectivamente) y presentaron un AIC menor en comparación con el modelo más complejo. Aunque los modelos uno y tres son más parsimoniosos, la varianza explicada fue menor a la del modelo seleccionado (Cuadro 4).
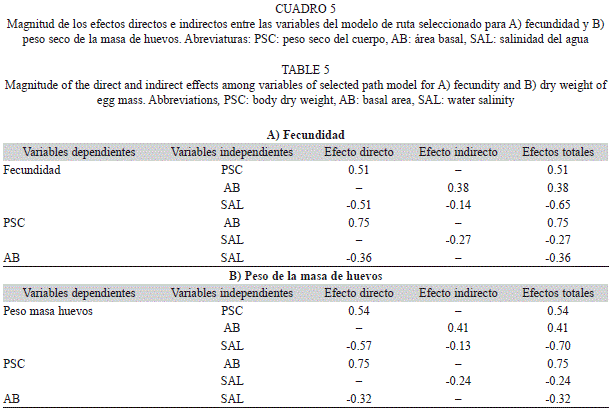
En el modelo seleccionado para fecundidad (Fig. 3A), el mayor efecto total (directo + indirecto) sobre esta variable fue el ejercido por la salinidad (-0.65), otros efectos totales importantes (>0.5) ocurren de mayor a menor entre: el área basal (AB) y el peso seco del cuerpo (PSC), y entre PSC y la fecundidad (Cuadro 5A). El mayor efecto total sobre el peso de los huevos (PSH) también lo ejerce la salinidad (-0.70) (Cuadro 5B); los otros efectos totales más fuertes, mantienen la misma tendencia observada con la fecundidad, sólo que la magnitud varía ligeramente (Cuadro 5, Fig. 3).
Materia orgánica y composición elemental de los huevos: El porcentaje de materia orgánica en los huevos no presentó diferencias entre localidades, mientras que en la relación C/Nhuevos sí las presentó (Cuadro 2, Cuadro 3). El máximo valor de la relación C/Nhuevos se observó en el manglar marino arbóreo (TIM), y en el resto de los manglares el valor C/Nhuevos fue similar. Tanto la materia orgánica de los huevos como la relación C/Nhuevos no se correlacionaron con las variables ambientales examinadas (r, p>0.05).
Discusión
Los resultados de este estudio comprobaron que el rendimiento reproductivo de las hembras de A. pisonii presentó diferencias entre hábitats, encontrando una mayor inversión (Ro1, Ro2) en las hembras del manglar estuarino con mayor desarrollo estructural (BR), respecto al observado en los otros manglares evaluados. Por su parte los resultados del análisis de rutas indicaron un efecto multicausal de las variables morfológicas y ambientales sobre la inversión reproductiva de A. pisonii, en particular la salinidad fue la variable que ejerció la mayor influencia sobre la fecundidad y el peso de la masa de huevos (PSH), con un efecto total (directo + indirecto) negativo de -0.65 para fecundidad y ligeramente superior (-0.70) para PSH. Los análisis de rutas entre otras cosas, permiten visualizar los efectos, tanto directos como indirectos de las variables, ofreciendo una idea más completa de las relaciones que se establecen en el sistema estudiado (Ortiz-Pulido, 2000).
En general, se ha demostrado que la fecundidad incrementa con el peso y la talla del cuerpo en cangrejos braquiuros (Hines, 1982); el mayor determinante para el tamaño de la puesta es el espacio disponible para la acumulación de vitelo en la cavidad corporal (Hines, 1982, 1991). De igual forma en A. pisonii se ha registrado que la talla del cuerpo es la principal determinante de la salida reproductiva, así hembras grandes producirán un mayor número de huevos que hembras de tallas pequeñas (Hines, 1982; Conde, & Díaz, 1989a; Díaz, & Conde, 1989; Leme, & Negreiros-Fransozo, 1998; Conde et al., 2000).
En estudios realizados en Venezuela se han encontrado diferencias en la fecundidad promedio de A. pisonii entre poblaciones de ambientes marino y estuarino, observando 16 379 huevos (7 952-34 057 huevos) en un estuario (Conde, & Díaz, 1989a), y un promedio de 11 577 huevos (3 724-27 134 huevos) en un ambiente marino (Díaz, & Conde, 1989). Nuestros resultados confirman que existen diferencias significativas en la fecundidad y el peso de la masa de huevos (PSH) entre el manglar estuarino y el resto de los manglares. Estas diferencias en el número promedio de huevos entre las distintas poblaciones pueden estar asociadas a la talla promedio de los cangrejos y a la disponibilidad diferencial de recursos ofrecida por cada hábitat; en este trabajo se observan las máximas tallas asociadas a los manglares con mayor área basal y altura máxima, bajo condiciones estuarinas, y las mínimas en manglares de menor altura y área basal, bajo condiciones hipersalinas. Conde & Díaz (1989b) plantearon un modelo conceptual para explicar la amplia plasticidad fenotípica observada en la talla de A. pisonii entre poblaciones asociadas a manglares con distintos niveles de desarrollo estructural planteando que podría ser una respuesta a las condiciones locales de recursos, y proponiendo que las diferencias en la productividad de los manglares podría representar un gradiente de recursos. Negreiros-Fransozo (2002) también encuentra una relación entre la talla de A. pisonii y la estructura del manglar en Brasil, donde los cangrejos más pequeños y los más grandes fueron encontrados en sitios donde el diámetro y densidad de árboles, fueron los mínimos y máximos respectivamente, apoyando la hipótesis de Conde y Díaz (1989b). Castiglioni, Santos, Reigada y Negreiros-Fransozo (2004) reportan las máximas fecundidades para el cangrejo Armases rubripes, en manglares que presentaban el mayor desarrollo estructural, y sugirieron una relación entre la fecundidad y la productividad del manglar producto de un mayor suministro de recursos.
López & Conde (2013) encontraron una asociación directa entre la variedad de rubros consumidos por A. pisonii y la altura del manglar, pero sólo durante el periodo de lluvias, sugiriendo que existe una mayor oferta de recursos en los manglares con mayor desarrollo estructural visible durante esta temporada. Las lluvias generalmente ocasionan un incremento en la productividad de los manglares, apreciable sobre todo en regiones áridas y semiáridas (Day, Conner, Ley-Lou, Day, & Navarro, 1987; Rodríguez, 1987; McKee, 2002), como es el caso de los manglares de la península de Paraguaná de este estudio; esto toma más sentido si consideramos que A. pisonii es una especie catalogada como omnívora oportunista, cuyos individuos deben tomar ventaja estas condiciones para su beneficio (Beever, Simberloff, & King, 1979; Brogim, & Lana, 1997; Díaz, & Conde, 1988; Erickson, Feller, Paul, Kwiatkowski, & Lee, 2008; Riley, Vogel, & Griffen, 2014).
En nuestro sistema de estudio se pudo constatar que todas las poblaciones produjeron huevos de talla (volumen) más o menos similar; sin embargo, las hembras del manglar estuarino -el cual posee mayor desarrollo estructural- presentaron huevos significativamente más pesados que las hembras de las otras poblaciones. Kyomo (2000) consiguió resultados similares a los nuestros cuando comparó entre tres poblaciones de Sesarma intermedia en Japón, observando una fecundidad similar entre poblaciones, y también una talla similar de los huevos, pero encontró diferencias en el peso de los huevos. Los huevos más pesados se observaron en una población donde un temprano incremento en el suministro de alimentos, produjo una anticipada producción de huevos; y sugiere que hubo suficientes nutrientes que se invirtieron en la producción de huevos más pesados. También se ha mencionado que la calidad o cantidad del alimento disponible en cada hábitat, puede causar diferencias en las características reproductivas de poblaciones separadas espacialmente (Kyomo, 2000; Bas, Spivak, & Anger, 2007).
Respecto a la calidad del recurso foliar, se ha encontrado en manglares venezolanos un mayor contenido de polifenoles y menor de nitrógeno, asociado a manglares con menor altura en zonas hipersalinas, donde además se observan que las poblaciones de A. pisonii presentan los menores anchos de caparazón (Conde et al., 1995). López (2010) corrobora el incremento en el contenido de nitrógeno foliar en los manglares con mayor desarrollo estructural y menor salinidad del agua. La literatura menciona que manglares achaparrados son usualmente observados sobre suelos con alta salinidad y pobres en nutrientes, cuyas hojas son más coriáceas y difíciles de manipular por los herbívoros presentando un bajo valor nutricional (Feller, 1995).
En el manglar estuarino de BR se observan altos porcentajes de hembras ovadas durante todo el año, mientras que en las poblaciones de Paraguaná la reproducción presenta un pulso durante los meses de lluvias, y en los manglares hipersalinnos de TIA y TAC es difícil observar hembras ovadas durante los primeros meses del año (Conde 1990; Conde, & Díaz, 1992b; López, 2010). Este comportamiento parece sugerir que el manglar de BR ofrece una mayor calidad del hábitat, ya sea por una mayor disponibilidad de recursos en el año, mayor calidad de las hojas, menor salinidad del agua y un aporte de agua dulce constante, lo cual puede estar manifestándose en el mayor peso de la masa de huevos y fecundidad respecto a los otros manglares evaluados en este trabajo (marino e hipersalinos).
Leme (2006) encuentra valores similares a los observados en este estudio respecto al volumen individual de los huevos de A. pisonii (0.015±0.002mm3, Leme 2006; 0.017±0.002mm3, este estudio); no obstante en relación al peso de cada huevo, nuestros resultados registraron valores inferiores, sobre todo en hembras de los manglares marino e hipersalinos (5.50±0.36µg, Leme 2006; 3.36±0.27µg hipersalino-achaparrado). Las poblaciones que evalúa Leme (2006) se localizan en la desembocadura de un río en Sao Paulo, Brasil, por lo tanto tienen un continuo aporte de agua dulce durante el año, y el peso de los huevos de las hembras de Boca de Ricoa (manglar arbóreo estuarino) en nuestro caso, fue el más similar a los datos obtenidos por Leme (2006). Es decir en los manglares con mayor salinidad, las hembras de A. pisonii presentaron huevos más livianos.
Los resultados del análisis de rutas reflejan el papel relevante de la salinidad sobre la inversión reproductiva de A. pisonii, con efectos tanto indirectos -a través de la estructura de la vegetación- como directos -a través de la influencia sobre el presupuesto energético del cangrejo-. Se ha demostrado que los cangrejos sesármidos comprometen la asignación de energía cuando se encuentran en altas salinidades del agua (48-65ups), de manera que la asignación obtenida a través del consumo de alimento, es usada en la excreción (osmorregulación) y respiración (Gillikin, De Wachter, & Tack, 2004), afectando negativamente la inversión en reproducción o crecimiento; es decir, ocurre una distribución diferencial de recursos entre diversos procesos, estableciéndose una relación costo-beneficio (Stearns, 1976). Por lo tanto, es posible que la energía dirigida a crecimiento o reproducción en los cangrejos de manglares hipersalinos en nuestro estudio, pueda estar comprometida en mantener el equilibrio osmótico y la respiración, restándole energía a otras funciones. Para comprobar esto son necesarias manipulaciones experimentales que evalúen la respuesta fisiológica de A. pisonii bajo diferentes condiciones de salinidad. Sin embargo, otras variables no consideradas, pueden estar influyendo sobre la inversión reproductiva de A. pisonii, por ejemplo aspectos relacionados con la calidad del recurso, diferentes a la relación C/N foliar (no seleccionada por el modelo en el análisis de ruta), como el contenido de fibra cruda y de agua en las hojas (de Lacerda et al., 1986; Faraco, & Lana, 2004) o la disponibilidad de recursos alternativos (algas, frutos, flores, insectos) (Erickson et al., 2003; López, & Conde, 2013). También podría influir la dinámica de las mareas, o las interacciones de competencia, canibalismo y depredación (Wilson, 1989; Feller, & Chamberlain, 2007).
La relación C/Nhuevos en este trabajo, no mostraron la misma tendencia que el C/N foliar, observando en el manglar marino valores más altos, lo cual sugiere que deben existir otras variables no consideradas en este trabajo, que pueden estar influyendo sobre la calidad de los huevos en las hembras de este manglar. En conclusión, la inversión reproductiva de A. pisonii en las poblaciones evaluadas está determinada de forma multicausal, es decir por una combinación de variables (ambientales y morfológicas) que interactúan sobre la fecundidad y el peso de la masa de huevos, donde la salinidad ejerce el efecto más importante. Así se observa la menor fecundidad y PSH en manglares con menor desarrollo estructural asociados a ambientes de elevada salinidad del agua, donde predominan las menores tallas de cangrejos. Se propone que la combinación de variables en el manglar estuarino, menor salinidad, mayor desarrollo estructural, lo cual implica una mayor variedad y disponibilidad de recursos, puede explicar la mayor inversión de energía en reproducción, que en hembras que habitan manglares con menor desarrollo estructural, bajo condiciones ambientales adversas.
Agradecimientos
Este proyecto fue financiado por el Instituto Venezolano de Investigaciones Científicas (IVIC), proyecto N°467. Los autores agradecen a Carlos Méndez, José Montoya y Wilmer Rojas, quienes gentilmente contribuyeron a mejorar el manuscrito. A Eleine Severino por el análisis de muestras en el analizador elemental del Centro de Química, IVIC.
Referencias
Akaike, H. (1987). Factor analysis and AIC. Psychometrika, 52(3), 317-332. [ Links ]
Bas, C. C., Spivak, E. D., & Anger, K. (2007). Seasonal and interpopulational variability in fecundity, egg size, and elemental composition (CHN) of eggs and larvae in a grapsoid crab, Chasmagnathus granulatus. Helgoland Marine Research, 61(4), 225-237. [ Links ]
Beever, J. W., Simberloff, D., & King, L. L. (1979). Herbivory and predation by the mangrove tree crab Aratus pisonii. Oecologia, 43(3), 317-328. [ Links ]
Brogim, R. A., & Lana, P. C. (1997). Espectro alimentar de Aratus pisonii, Chasmagnathus granulata e Sesarma rectum (Decapoda, Grapsidae) em um manguezal na Baía de Paranaguá, Paraná. Iheringia, Série Zoologia, 83, 35-43. [ Links ]
Brown, M., & Forsythe, A. (1974). Robust tests for the equality of variances. Journal of the American Statistical Association, 69(346), 364-367. [ Links ]
Castiglioni, D. S., Santos, S., Reigada, A. L., & Negreiros-Fransozo, M. L. (2004). Reproductive ecology of Armases rubripes (Sesarmidae) from mangroves of southeastern Brazil. Nauplius, 12(2), 109-117. [ Links ]
Chance, F. A., & Hobss, H. H. (1969). The freshwater and terrestrial decapod crustaceans of the West Indies with special reference to Dominica. Bulletin U S National Museum, 292, 1-258. [ Links ]
Clarke, A., Hopkins, C. C., & Nilssen, E. M. (1991). Egg size and reproductive output in the deep-water prawn Pandalus borealis Kroyer, 1838. Functional Ecology, 5(6), 724-730. [ Links ]
Conde, J. E. (1990). Ecología poblacional del cangrejo de mangle Aratus pisonii (H. Milne-Edwards) (Brachyura: Grapsidae) en hábitats extremos. Instituto Venezolano de Investigaciones Científicas. [ Links ]
Conde, J. E., & Díaz, H. (1989a). Productividad del habitat e historias de vida del cangrejo de mangle Aratus pisonii (H. Milne Edwards) (Brachyura, Grapsidae). Boletín Del Instituto Oceanográfico de Venezuela, 28, 113-120. [ Links ]
Conde, J. E., & Díaz, H. (1989b). The mangrove tree crab Aratus pisonii in a tropical estuarine coastal lagoon. Estuarine, Coastal and Shelf Science, 28(6), 639-650. [ Links ]
Conde, J. E., Díaz, H., & Rodríguez, G. (1989). Crecimiento reducido en el cangrejo de mangle Aratus pisonii (H. Milne-Edwards) (Brachyura: Grapsidae). Acta Científica Venezolana, 40, 159-160. [ Links ]
Conde, J. E., & Díaz, H. (1992a). Variations in intraspecific relative size at the onset of maturity (RSOM) in Aratus pisonii (H. Milne Edwards, 1837) (Decapoda, Brachyura, Grapsidae). Crustaceana, 62(2), 214-216. [ Links ]
Conde, J. E., & Díaz, H. (1992b). Extension of the stunting range in ovigerous females of the mangrove crab Aratus pisonii (H. Milne Edwards, 1837) (Decapoda, Brachyura, Grapsidae). Crustaceana, 62(3), 319-322. [ Links ]
Conde, J. E., Flores, S., & Díaz, H. (1995). Nitrogen and tannins in mangrove leaves might explain interpopulation variations in the crab Aratus pisonii. Acta Científica Venezolana, 46, 303-304. [ Links ]
Conde, J. E., Tognella, M. M., Paes, E. T., Soares, M. L., Louro, I. A., & Schaeffer-Novelli, Y. (2000). Population and life history features of the crab Aratus pisonii (Decapoda: Grapsidae) in a subtropical estuary. Interciencia, 25(3), 151-158. [ Links ]
Colpo, K. D., Chacur, M. M., Guimarães, F. J., & Negreiros-Fransozo, M. L. (2011). Subtropical Brazilian mangroves as a refuge of crab (Decapoda: Brachyura) diversity. Biodiversity and Conservation, 20(13), 3239-3250. [ Links ]
Day, J. W., Conner, W. H., Ley-Lou, F., Day, R. H., & Navarro, A. M. (1987). The productivity and composition of mangrove forests, Laguna de Términos, Mexico. Aquatic Botany, 27(3), 267-284. [ Links ]
De Lacerda, L. D., Jose, D. V., de Rezende, C. E., Francisco, M. C., Wasserman, J. C., & Martins, J. C. (1986). Leaf chemical characteristics affecting herbivory in a New World mangrove forest. Biotropica, 18(4), 350-355. [ Links ]
Díaz, H. (1980). The mole crab Emerita talpoida (Say): a case of changing life history pattern. Ecological Monographs, 50(4), 437-456. [ Links ]
Díaz, H., & Conde, J. E. (1988). On the food sources for the mangrove tree crab Aratus pisonii (Brachyura: Grapsidae). Biotropica, 20(4), 348-350. [ Links ]
Díaz, H., & Conde, J. E. (1989). Population dynamics and life history of the mangrove crab Aratus pisonii (Brachyura, Grapsidae) in a marine environment. Bulletin of Marine Science, 45(1), 148-163. [ Links ]
Díaz, H., Conde, J. E., & Bevilacqua, M. (1983). A volumetric method for estimating fecundity in Decapoda. Marine Ecology Progress Series, 10, 203-206. [ Links ]
Erickson, A. A., Feller, I. C., Paul, V. J., Kwiatkowski, L. M., & Lee, W. (2008). Selection of an omnivorous diet by the mangrove tree crab Aratus pisonii in laboratory experiments. Journal of Sea Research, 59(1-2), 59-69. [ Links ]
Erickson, A. A., Saltis, M., Bell, S. S., & Dawes, C. J. (2003). Herbivore feeding preferences as measured by leaf damage and stomatal ingestion: a mangrove crab example. Journal of Experimental Marine Biology and Ecology, 289(1), 123-138. [ Links ]
Faraco, L. F., & Lana, P. (2004). Leaf-consumption levels in subtropical mangroves of Paranaguá Bay (SE Brazil). Wetlands Ecology and Management, 12(2), 115-122. [ Links ]
Feller, I. C. (1995). Effects of Nutrient Enrichment on Growth and Herbivory of Dwarf Red Mangrove (Rhizophora mangle). Ecological Monographs, 65(4), 477-505. [ Links ]
Feller, I. C., & Chamberlain, A. (2007). Herbivore responses to nutrient enrichment and landscape heterogeneity in a mangrove ecosystem. Oecologia, 153(3), 607-616. [ Links ]
Gillikin, D. P., De Wachter, B., & Tack, J. F. (2004). Physiological responses of two ecologically important Kenyan mangrove crabs exposed to altered salinity regimes. Journal of Experimental Marine Biology and Ecology, 301(1), 93-109. [ Links ]
Hancock, M. A., Hughes, J. M., & Bunn, S. E. (1998). Influence of genetic and environmental factors on egg and clutch sizes among populations of Paratya australiensis Kemp (Decapoda: Atyidae) in upland rainforest streams, south-east Queensland. Oecologia, 115(4), 483-491. [ Links ]
Hartnoll, R. G. (1965). Notes on the marine grapsid crabs of Jamaica. Proceedings of the Linnean Society of London, 176(2), 113-147. [ Links ]
Hartnoll, R. G. (2006). Reproductive Investment in Brachyura. Hydrobiologia, 557(1), 31-40. [ Links ]
Hartnoll, R. G., & Gould, P. (1988). Brachyuran life history strategies and the optimization of egg production. Symposia of the Zoological Society of London, 59, 1-9. [ Links ]
Hernáez, P. (2001). Producción y rendimiento reproductivo en Petrolisthes granulosus (Decapoda, Anomura, Porcellanidae) en diferentes localidades del norte de Chile: Una comparación latitudinal. Investigaciones Marinas, 29(1), 73-81. [ Links ]
Hines, A. H. (1982). Allometric constraints and variables of reproductive effort in brachyuran crabs. Marine Biology, 69(3), 309-320. [ Links ]
Hines, A. H. (1991). Fecundity and reproductive output in nine species of Cancer crabs (Crustacea, Brachyura, Cancridae). Canadian Journal of Fisheries and Aquatic Sciences, 48(2), 267-275. [ Links ]
Kyomo, J. (2000). Intraspecific variation of reproductive strategies of the crab Sesarma intermedia: a consequence of habitat variations. Bulletin of Marine Science, 66(1), 157-171. [ Links ]
Leme, M. H. (2006). Investimento reprodutivo e produção de ovos em desovas consecutivas do caranguejo Aratus pisonii (H. Milne Edwards) (Crustacea, Brachyura, Grapsoidea). Revista Brasileira de Zoologia, 23(3), 727-732. [ Links ]
Leme, M. H., & Negreiros-Fransozo, M. L. (1998). Fecundity of Aratus pisonii (Decapoda, Grapsidae) in Ubatuba region, state of Sao Paulo, Brazil. Iheringia, Série Zoologia, 84, 73-77. [ Links ]
Lima, G. V., Soares, M. R., & Oshiro, L. M. (2006). Reproductive biology of the sesarmid crab Armases rubripes (Decapoda, Brachyura) from an estuarine area of the Sahy River, Sepetiba Bay, Rio de Janeiro, Brazil. Iheringia. Série Zoologia, 96(1), 47-52. [ Links ]
López, B. (2010). Ecología trófica y energética del cangrejo de mangle Aratus pisonii (Crustacea: Brachyura: Grapsidae) (H. Milne-Edwards). Instituto Venezolano de Investigaciones Científicas. [ Links ]
López, B., Barreto, M. B., & Conde, J. E. (2011). Caracterización de los manglares de zonas semiáridas en el noroccidente de Venezuela. Interciencia, 36(12), 888-893. [ Links ]
López, B., & Conde, J. E. (2013). Dietary variation in the crab Aratus pisonii (H. Milne Edwards, 1837) (Decapoda, Sesarmidae) in a mangrove gradient in northwestern Venezuela. Crustaceana, 86(9), 1051-1069. [ Links ]
López, L., Jeri, T., González, C., & Rodríguez, S. (1997). Fecundidad y esfuerzo reproductivo de Petrolisthes granulosus (Guérin, 1835) en Iquique, Chile (Decapoda, Anomura, Porcellanidae). Investigaciones Marinas, 25, 159-165. [ Links ]
Mashiko, K. (1990). Diversified egg and clutch sizes among local populations of the fresh-water prawn Macrobrachium nipponense (de Haan). Journal of Crustacean Biology, 10(2), 306-314. [ Links ]
McKee, K. L. (2002). Mangrove ecosystems: definitions, distribution, zonation, forest structure, trophic structure and ecological significance. In I. C. Feller, & M. Sitnik (Eds.), Mangrove ecology workshop manual (pp. 1-6). Washington, DC: Smithsonian Institution Press. [ Links ]
Negreiros-Fransozo, M. L. (2002). Size Variation in the Grapsid Crab Aratus Pisonii (H. Milne-Edwards, 1837) Among Populations of Different Subtropical Mangroves. In E. Escobar-Briones, & F. Alvarez (Eds.), Modern Approaches to the Study of Crustacea (pp. 183-188). Springer US. [ Links ]
Ortiz-Pulido, R. (2000). Análisis de rutas en biología: estadística para sistemas multicausales. Interciencia, 25(7), 329-336. [ Links ]
Ouellet, P., & Plante, F. (2004). An investigation of the sources of variability in American lobster (Homarus americanus) eggs and larvae: female size and reproductive status, and interannual and interpopulation comparisons. Journal of Crustacean Biology, 24(3), 481-495. [ Links ]
Pérez, E., Medrano, L. A., & Sánchez, J. (2013). El Path Analysis: conceptos básicos y ejemplos de aplicación. Revista Argentina de Ciencias Del Comportamiento, 5(1), 52-66. [ Links ]
Riley, M., Vogel, M., & Griffen, B. (2014). Fitness-associated consequences of an omnivorous diet for the mangrove tree crab Aratus pisonii. Aquatic Biology, 20(1), 35-43. [ Links ]
Rodríguez, G. (1987). Structure and production in neotropical mangroves. Trends in Ecology & Evolution, 2(9), 264-267. [ Links ]
Sastry, A. N. (1970). Reproductive physiological variation in latitudinally separated populations of the bay scallop, Aequipecten irradians Lamarck. The Biological Bulletin, 138, 56-65. [ Links ]
Snedaker, S. C. (1989). Overview of ecology of mangroves and information needs for Florida Bay. Bulletin of Marine Science, 44(1), 341-347. [ Links ]
Sokal, R. R., & Rohlf, F. J. (1995). Biometry: the principals and practice of statistics in biological research (3rd ed., p. 887). New York: W. H. Freeman. [ Links ]
SPSS. (1999). SPSS Base 10.0 for Windows user’s guide. Chicago, IL: SPSS Inc. [ Links ]
Stearns, S. C. (1976). Life-History Tactics: A Review of the Ideas. The Quarterly Review of Biology, 51(1), 3-47. [ Links ]
Von Oertzen, T., Brandmaier, A. M., & Tsang, S. (2013). Ωnyx User Guide. [ Links ]
Warner, G. F. (1967). The life history of the mangrove tree crab, Aratus pisonii. Journal of Zoology, 153, 321-335. [ Links ]
Wilson, K. A. (1989). Ecology of mangrove crabs: predation, physical factors and refuges. Bulletin of Marine Science, 44(1), 263-273. [ Links ]
Wright, S. (1934). The Method of Path Coefficients. The Annals of Mathematical Statistics, 5(3), 161-215. [ Links ]
Bas, C. C., Spivak, E. D., & Anger, K. (2007). Seasonal and interpopulational variability in fecundity, egg size, and elemental composition (CHN) of eggs and larvae in a grapsoid crab, Chasmagnathus granulatus. Helgoland Marine Research, 61(4), 225-237. [ Links ]
Beever, J. W., Simberloff, D., & King, L. L. (1979). Herbivory and predation by the mangrove tree crab Aratus pisonii. Oecologia, 43(3), 317-328. [ Links ]
Brogim, R. A., & Lana, P. C. (1997). Espectro alimentar de Aratus pisonii, Chasmagnathus granulata e Sesarma rectum (Decapoda, Grapsidae) em um manguezal na Baía de Paranaguá, Paraná. Iheringia, Série Zoologia, 83, 35-43. [ Links ]
Brown, M., & Forsythe, A. (1974). Robust tests for the equality of variances. Journal of the American Statistical Association, 69(346), 364-367. [ Links ]
Castiglioni, D. S., Santos, S., Reigada, A. L., & Negreiros-Fransozo, M. L. (2004). Reproductive ecology of Armases rubripes (Sesarmidae) from mangroves of southeastern Brazil. Nauplius, 12(2), 109-117. [ Links ]
Chance, F. A., & Hobss, H. H. (1969). The freshwater and terrestrial decapod crustaceans of the West Indies with special reference to Dominica. Bulletin U S National Museum, 292, 1-258. [ Links ]
Clarke, A., Hopkins, C. C., & Nilssen, E. M. (1991). Egg size and reproductive output in the deep-water prawn Pandalus borealis Kroyer, 1838. Functional Ecology, 5(6), 724-730. [ Links ]
Conde, J. E. (1990). Ecología poblacional del cangrejo de mangle Aratus pisonii (H. Milne-Edwards) (Brachyura: Grapsidae) en hábitats extremos. Instituto Venezolano de Investigaciones Científicas. [ Links ]
Conde, J. E., & Díaz, H. (1989a). Productividad del habitat e historias de vida del cangrejo de mangle Aratus pisonii (H. Milne Edwards) (Brachyura, Grapsidae). Boletín Del Instituto Oceanográfico de Venezuela, 28, 113-120. [ Links ]
Conde, J. E., & Díaz, H. (1989b). The mangrove tree crab Aratus pisonii in a tropical estuarine coastal lagoon. Estuarine, Coastal and Shelf Science, 28(6), 639-650. [ Links ]
Conde, J. E., Díaz, H., & Rodríguez, G. (1989). Crecimiento reducido en el cangrejo de mangle Aratus pisonii (H. Milne-Edwards) (Brachyura: Grapsidae). Acta Científica Venezolana, 40, 159-160. [ Links ]
Conde, J. E., & Díaz, H. (1992a). Variations in intraspecific relative size at the onset of maturity (RSOM) in Aratus pisonii (H. Milne Edwards, 1837) (Decapoda, Brachyura, Grapsidae). Crustaceana, 62(2), 214-216. [ Links ]
Conde, J. E., & Díaz, H. (1992b). Extension of the stunting range in ovigerous females of the mangrove crab Aratus pisonii (H. Milne Edwards, 1837) (Decapoda, Brachyura, Grapsidae). Crustaceana, 62(3), 319-322. [ Links ]
Conde, J. E., Flores, S., & Díaz, H. (1995). Nitrogen and tannins in mangrove leaves might explain interpopulation variations in the crab Aratus pisonii. Acta Científica Venezolana, 46, 303-304. [ Links ]
Conde, J. E., Tognella, M. M., Paes, E. T., Soares, M. L., Louro, I. A., & Schaeffer-Novelli, Y. (2000). Population and life history features of the crab Aratus pisonii (Decapoda: Grapsidae) in a subtropical estuary. Interciencia, 25(3), 151-158. [ Links ]
Colpo, K. D., Chacur, M. M., Guimarães, F. J., & Negreiros-Fransozo, M. L. (2011). Subtropical Brazilian mangroves as a refuge of crab (Decapoda: Brachyura) diversity. Biodiversity and Conservation, 20(13), 3239-3250. [ Links ]
Day, J. W., Conner, W. H., Ley-Lou, F., Day, R. H., & Navarro, A. M. (1987). The productivity and composition of mangrove forests, Laguna de Términos, Mexico. Aquatic Botany, 27(3), 267-284. [ Links ]
De Lacerda, L. D., Jose, D. V., de Rezende, C. E., Francisco, M. C., Wasserman, J. C., & Martins, J. C. (1986). Leaf chemical characteristics affecting herbivory in a New World mangrove forest. Biotropica, 18(4), 350-355. [ Links ]
Díaz, H. (1980). The mole crab Emerita talpoida (Say): a case of changing life history pattern. Ecological Monographs, 50(4), 437-456. [ Links ]
Díaz, H., & Conde, J. E. (1988). On the food sources for the mangrove tree crab Aratus pisonii (Brachyura: Grapsidae). Biotropica, 20(4), 348-350. [ Links ]
Díaz, H., & Conde, J. E. (1989). Population dynamics and life history of the mangrove crab Aratus pisonii (Brachyura, Grapsidae) in a marine environment. Bulletin of Marine Science, 45(1), 148-163. [ Links ]
Díaz, H., Conde, J. E., & Bevilacqua, M. (1983). A volumetric method for estimating fecundity in Decapoda. Marine Ecology Progress Series, 10, 203-206. [ Links ]
Erickson, A. A., Feller, I. C., Paul, V. J., Kwiatkowski, L. M., & Lee, W. (2008). Selection of an omnivorous diet by the mangrove tree crab Aratus pisonii in laboratory experiments. Journal of Sea Research, 59(1-2), 59-69. [ Links ]
Erickson, A. A., Saltis, M., Bell, S. S., & Dawes, C. J. (2003). Herbivore feeding preferences as measured by leaf damage and stomatal ingestion: a mangrove crab example. Journal of Experimental Marine Biology and Ecology, 289(1), 123-138. [ Links ]
Faraco, L. F., & Lana, P. (2004). Leaf-consumption levels in subtropical mangroves of Paranaguá Bay (SE Brazil). Wetlands Ecology and Management, 12(2), 115-122. [ Links ]
Feller, I. C. (1995). Effects of Nutrient Enrichment on Growth and Herbivory of Dwarf Red Mangrove (Rhizophora mangle). Ecological Monographs, 65(4), 477-505. [ Links ]
Feller, I. C., & Chamberlain, A. (2007). Herbivore responses to nutrient enrichment and landscape heterogeneity in a mangrove ecosystem. Oecologia, 153(3), 607-616. [ Links ]
Gillikin, D. P., De Wachter, B., & Tack, J. F. (2004). Physiological responses of two ecologically important Kenyan mangrove crabs exposed to altered salinity regimes. Journal of Experimental Marine Biology and Ecology, 301(1), 93-109. [ Links ]
Hancock, M. A., Hughes, J. M., & Bunn, S. E. (1998). Influence of genetic and environmental factors on egg and clutch sizes among populations of Paratya australiensis Kemp (Decapoda: Atyidae) in upland rainforest streams, south-east Queensland. Oecologia, 115(4), 483-491. [ Links ]
Hartnoll, R. G. (1965). Notes on the marine grapsid crabs of Jamaica. Proceedings of the Linnean Society of London, 176(2), 113-147. [ Links ]
Hartnoll, R. G. (2006). Reproductive Investment in Brachyura. Hydrobiologia, 557(1), 31-40. [ Links ]
Hartnoll, R. G., & Gould, P. (1988). Brachyuran life history strategies and the optimization of egg production. Symposia of the Zoological Society of London, 59, 1-9. [ Links ]
Hernáez, P. (2001). Producción y rendimiento reproductivo en Petrolisthes granulosus (Decapoda, Anomura, Porcellanidae) en diferentes localidades del norte de Chile: Una comparación latitudinal. Investigaciones Marinas, 29(1), 73-81. [ Links ]
Hines, A. H. (1982). Allometric constraints and variables of reproductive effort in brachyuran crabs. Marine Biology, 69(3), 309-320. [ Links ]
Hines, A. H. (1991). Fecundity and reproductive output in nine species of Cancer crabs (Crustacea, Brachyura, Cancridae). Canadian Journal of Fisheries and Aquatic Sciences, 48(2), 267-275. [ Links ]
Kyomo, J. (2000). Intraspecific variation of reproductive strategies of the crab Sesarma intermedia: a consequence of habitat variations. Bulletin of Marine Science, 66(1), 157-171. [ Links ]
Leme, M. H. (2006). Investimento reprodutivo e produção de ovos em desovas consecutivas do caranguejo Aratus pisonii (H. Milne Edwards) (Crustacea, Brachyura, Grapsoidea). Revista Brasileira de Zoologia, 23(3), 727-732. [ Links ]
Leme, M. H., & Negreiros-Fransozo, M. L. (1998). Fecundity of Aratus pisonii (Decapoda, Grapsidae) in Ubatuba region, state of Sao Paulo, Brazil. Iheringia, Série Zoologia, 84, 73-77. [ Links ]
Lima, G. V., Soares, M. R., & Oshiro, L. M. (2006). Reproductive biology of the sesarmid crab Armases rubripes (Decapoda, Brachyura) from an estuarine area of the Sahy River, Sepetiba Bay, Rio de Janeiro, Brazil. Iheringia. Série Zoologia, 96(1), 47-52. [ Links ]
López, B. (2010). Ecología trófica y energética del cangrejo de mangle Aratus pisonii (Crustacea: Brachyura: Grapsidae) (H. Milne-Edwards). Instituto Venezolano de Investigaciones Científicas. [ Links ]
López, B., Barreto, M. B., & Conde, J. E. (2011). Caracterización de los manglares de zonas semiáridas en el noroccidente de Venezuela. Interciencia, 36(12), 888-893. [ Links ]
López, B., & Conde, J. E. (2013). Dietary variation in the crab Aratus pisonii (H. Milne Edwards, 1837) (Decapoda, Sesarmidae) in a mangrove gradient in northwestern Venezuela. Crustaceana, 86(9), 1051-1069. [ Links ]
López, L., Jeri, T., González, C., & Rodríguez, S. (1997). Fecundidad y esfuerzo reproductivo de Petrolisthes granulosus (Guérin, 1835) en Iquique, Chile (Decapoda, Anomura, Porcellanidae). Investigaciones Marinas, 25, 159-165. [ Links ]
Mashiko, K. (1990). Diversified egg and clutch sizes among local populations of the fresh-water prawn Macrobrachium nipponense (de Haan). Journal of Crustacean Biology, 10(2), 306-314. [ Links ]
McKee, K. L. (2002). Mangrove ecosystems: definitions, distribution, zonation, forest structure, trophic structure and ecological significance. In I. C. Feller, & M. Sitnik (Eds.), Mangrove ecology workshop manual (pp. 1-6). Washington, DC: Smithsonian Institution Press. [ Links ]
Negreiros-Fransozo, M. L. (2002). Size Variation in the Grapsid Crab Aratus Pisonii (H. Milne-Edwards, 1837) Among Populations of Different Subtropical Mangroves. In E. Escobar-Briones, & F. Alvarez (Eds.), Modern Approaches to the Study of Crustacea (pp. 183-188). Springer US. [ Links ]
Ortiz-Pulido, R. (2000). Análisis de rutas en biología: estadística para sistemas multicausales. Interciencia, 25(7), 329-336. [ Links ]
Ouellet, P., & Plante, F. (2004). An investigation of the sources of variability in American lobster (Homarus americanus) eggs and larvae: female size and reproductive status, and interannual and interpopulation comparisons. Journal of Crustacean Biology, 24(3), 481-495. [ Links ]
Pérez, E., Medrano, L. A., & Sánchez, J. (2013). El Path Analysis: conceptos básicos y ejemplos de aplicación. Revista Argentina de Ciencias Del Comportamiento, 5(1), 52-66. [ Links ]
Riley, M., Vogel, M., & Griffen, B. (2014). Fitness-associated consequences of an omnivorous diet for the mangrove tree crab Aratus pisonii. Aquatic Biology, 20(1), 35-43. [ Links ]
Rodríguez, G. (1987). Structure and production in neotropical mangroves. Trends in Ecology & Evolution, 2(9), 264-267. [ Links ]
Sastry, A. N. (1970). Reproductive physiological variation in latitudinally separated populations of the bay scallop, Aequipecten irradians Lamarck. The Biological Bulletin, 138, 56-65. [ Links ]
Snedaker, S. C. (1989). Overview of ecology of mangroves and information needs for Florida Bay. Bulletin of Marine Science, 44(1), 341-347. [ Links ]
Sokal, R. R., & Rohlf, F. J. (1995). Biometry: the principals and practice of statistics in biological research (3rd ed., p. 887). New York: W. H. Freeman. [ Links ]
SPSS. (1999). SPSS Base 10.0 for Windows user’s guide. Chicago, IL: SPSS Inc. [ Links ]
Stearns, S. C. (1976). Life-History Tactics: A Review of the Ideas. The Quarterly Review of Biology, 51(1), 3-47. [ Links ]
Von Oertzen, T., Brandmaier, A. M., & Tsang, S. (2013). Ωnyx User Guide. [ Links ]
Warner, G. F. (1967). The life history of the mangrove tree crab, Aratus pisonii. Journal of Zoology, 153, 321-335. [ Links ]
Wilson, K. A. (1989). Ecology of mangrove crabs: predation, physical factors and refuges. Bulletin of Marine Science, 44(1), 263-273. [ Links ]
Wright, S. (1934). The Method of Path Coefficients. The Annals of Mathematical Statistics, 5(3), 161-215. [ Links ]
1.Laboratorio de Ecología y Genética de Poblaciones, Centro de Ecología, Instituto Venezolano de Investigaciones Científicas, Apartado Postal 20632, 1020-A Caracas, Venezuela; blopez@ivic.gob.ve, equinter@ivic.gob.ve
Recibido 10-VII-2014. Corregido 18-X-2014. Aceptado 19-XI-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}