










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.1 San José Jan./Mar. 2015
Dormancy-breaking requirements of Sophora tomentosa and Erythrina speciosa (Fabaceae) seeds
Requisitos para romper la latencia en semillas de Sophora tomentosa y Erythrina speciosa (Fabaceae)
Requisitos para romper la latencia en semillas de Sophora tomentosa y Erythrina speciosa (Fabaceae)
Carolina Maria Luzia Delgado1*, Alexandre Souza de Paula1, Marisa Santos1 & Maria Terezinha Silveira Paulilo1
Abstract
The physical dormancy of seeds has been poorly studied in species from tropical forests, such as the Atlantic Forest. This study aimed to examine the effect of moderate alternating temperatures on breaking the physical dormancy of seeds, the morphoanatomy and histochemistry of seed coats, and to locate the structure/region responsible for water entrance into the seed, after breaking the physical dormancy of seeds of two woody Fabaceae (subfamily Faboideae) species that occur in the Brazilian Atlantic Forest: Sophora tomentosa and Erythrina speciosa. To assess temperature effect, seeds were incubated in several temperature values that occur in the Atlantic Forest. For morphological and histochemical studies, sections of fixed seeds were subjected to different reagents, and were observed using light or epifluorescence microscopy, to analyze the anatomy and histochemistry of the seed coat. Treated and non-treated seeds were also analyzed using a scanning electron microscope (SEM) to observe the morphology of the seed coat. To localize the specific site of water entrance, the seeds were blocked with glue in different regions and also immersed in ink. In the present work a maximum temperature fluctuation of 15ºC was applied during a period of 20 days and these conditions did not increase the germination of S. tomentosa or E. speciosa. These results may indicate that these seeds require larger fluctuation of temperature than the applied or/and longer period of exposition to the temperature fluctuation. Blocking experiments water inlet combined with SEM analysis of the structures of seed coat for both species showed that besides the lens, the hilum and micropyle are involved in water absorption in seeds scarified with hot water. In seeds of E. speciosa the immersion of scarified seeds into an aniline aqueous solution showed that the solution first entered the seed through the hilum. Both species showed seed morphological and anatomical features for seed coats of the subfamily Faboideae. Lignin and callose were found around all palisade layers and the water impermeability and ecological role of these substances are discussed in the work.
Key words: physical dormancy, seeds, tropical woody Fabaceae, water gaps.
Resumen
La latencia física de las semillas ha sido poco estudiada en las especies de los bosques tropicales, como el bosque atlántico. Este estudio tuvo como objetivo examinar el efecto de las temperaturas moderadas alternantes en romper la latencia física de las semillas, la anatomía y la histoquímica de la cubierta de las semillas, y la localización de la estructura o región responsable de la entrada de agua, después de romper la latencia física de las semillas de Sophora tomentosa y Eythrina speciosa, dos especies leñosas de Fabaceae (subfamilia Faboideae) que presentes en el bosque atlántico de Brasil. Para cumplir con el primer objetivo se incubaron las semillas a varias temperaturas que se dan en el bosque atlántico. Para los estudios morfológicos e histoquímicos se fijaron secciones de semillas sometidos a diferentes reactivos y se observaron usando luz o microscopía de epifluorescencia para analizar la anatomía y la histoquímica de la cubierta de la semilla. Semillas tratadas y no tratadas se analizaron también usando un microscopio electrónico de barrido (MDB) o microscopio estereoscópico (ME) para observar la morfología de la cubierta de la semilla. Para localizar el sitio específico de la entrada de agua, las semillas fueron bloqueadas con pegamento en diferentes regiones y también sumergidas en tinta. En el presente trabajo se aplicó una fluctuación de temperatura máxima de 15°C durante un período de 20 días y estas condiciones no aumentó la germinación de S. tomentosa o E. speciosa. Estos resultados pueden indicar que estas semillas requieren mayor fluctuación de la temperatura que la aplicada y/o un período más largo de exposición a la fluctuación de la temperatura. Experimentos de bloqueo de entrada de agua combinada con el análisis de las estructuras de la cubierta de la semilla para ambas especies (SEM) mostró que, a pesar de la lente, el hilio y micropilo están implicados en la absorción de agua en las semillas escarificadas con agua caliente. En las semillas de E. speciosa la inmersión de semillas escarificadas en una solución acuosa de anilina mostró que la solución entró por primera vez a la semilla a través del hilio. Ambas especies mostraron características morfológicas y anatómicas de semillas con cubierta de la subfamilia Faboideae. La lignina y callosa se encontraron alrededor de todas las capas de empalizada y la impermeabilidad al agua y en el trabajo se discute el papel ecológico de estas sustancias.
Palabras clave: latencia física, semillas, Fabaceae tropicales.
Mature seeds of many plant species, particularly those in the Fabaceae, do not germinate readily under favorable environmental conditions because they are impermeable to water and/or gases (Argel, & Paton, 1999; Bewley, & Black, 1994). This type of impermeability is referred to by Baskin and Baskin (2001) as physical dormancy and is caused by a layer of palisade cells in the seed coat, which have thick, lignified secondary walls (macrosclereids) containing hydrophobic substances (Baskin, & Baskin, 2001).
In physically dormant seeds there are specialized anatomical structures in the coat that develop an opening where water can enter (Gama-Arachchige, Baskin, Geneve, & Baskin, 2013). Under natural conditions, it is also known that temperature is an important environmental factor that influences the breaking of physically dormant seeds (Kondo, & Takahashi, 2004; Jayasuriy, Baskin, Geneve, Baskin, & Chien, 2008; Vázquez-Yanes, & Orozco Segovia, 1982).
The family Fabaceae is the third largest family of angiosperms and the second most economically important family (Judd,Campbel, Kellongg, Steens, & Donogue, 2009). Several species of this group play a vital role in global biogeochemistry because they have nodules with nitrogen-fixing bacteria (Sprent, 2001). In Brazil, this family is represented by about 190 genera and 2 100 species. Many of these taxa play a prominent role in the floras of several vegetation formations, especially those belonging to the Atlantic domain (the Atlantic Forest and other vegetation types along the Atlantic coast), where they are important for their richness and abundance (Lima, 2000). However, there are few studies about the reproductive biology of species of Fabaceae (Brito, Pinheiro, & Sazima, 2010) including those about seed germination (Jayasuriy, Wijetunga, Baskin, & Baskin, 2010). In addition, studies about the physical dormancy of tree species from the Brazilian Atlantic Forest are quite rare, despite the Atlantic Forest being one of the most threatened ecosystems in Latin America (Myers, Mittermeier, Mittermeir, da Fonseca, & Kent, 2000).
This study utilized two species of Fabaceae (subfamily Faboideae), Sophora tomentosa L. and Erythrina speciosa Andr., which have physically dormant seeds. Sophora tomentosa occurs in restinga (Brito et al., 2010), which is a sunny and windy environment near the sea that has water and nutrient shortages (Bresolin, 1979). This species has autochorous and hydrochorous seed dispersal (Bechara, 2003), and produces mature fruits throughout the year but mostly in the summer (Dec-Feb) when there is more rain (Brito et al., 2010; Nogueira, & Arruda, 2006a). Erythrina speciosa is a neotropical tree distributed throughout the Southern and Southeast regions of Brazil (Lorenzi, 2002). It is typically found in fluvial forest and coastal moist plains, as well as flood-prone habitats, and is always in open and secondary formations (Klein, 1969; Medina et al., 2009). This species has autochorous seed dispersal and produces mature fruits at the end of spring and beginning of summer (Lorenzi, 2002) during the period of the year that usually has the largest amount of rainfall.
This work focused on the following aspects: 1) the structure and chemical composition of the seed coats, 2) the structure of the region where water enters the seeds, and 3) the effect of moderate alternating temperatures on breaking the physical dormancy of the seeds.
Materials and Methods
Collection of seeds: Seeds of Sophora tomentosa were collected at Daniela’s beach in July, 2009, and those of Erythrina speciosa in a fragment of Atlantic Forest in November, 2009. Both areas are located in the municipality of Florianópolis, Santa Catarina, Brazil (27º35’36’’ S - 48º35’60’’ W). An approximate quantity of one thousand seeds were collected from numerous plants and used in the study.
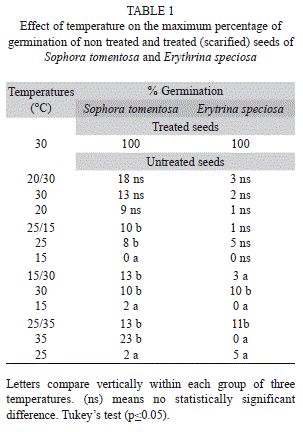
Germination of seeds: Intact seeds were sterilized by immersing them in 5% sodium hypochlorite for five minutes and then washed three times in distilled water. The seeds were placed in transparent plastic boxes on two sheets of filter paper (Whatman No.1, Whatman International Ltd. Maidstone, England) moistened with distilled water. Intact seeds were incubated at 15°C, 20°C, 25°C, 30°C and 35°C and a 12h/12h alternating temperature regime of 35ºC/25ºC, 30ºC/20ºC, 30ºC/15ºC, 25ºC/15ºC with a photoperiod of 12 hours. Hot water scarified seeds were also incubated under the same conditions but only at 30ºC. Four boxes, each with 20 seeds, were utilized both for intact and hot water scarified treatments for each temperature regime. Germinated seeds were counted every two days for 20 days, when the percentage of germination was stabilized. For scarified seeds, to check the efficiency of the period of incubation in hot water, previously weighed seeds in electronic precision balance (Gehaka Model BG200, precision 0.001g) of both species were put in water at 98ºC for 2, 3, 5, 7, 10, 15 and 20min, and weighed (again after these periods, to verify the increase of weight due to the water entrance. The best period was considered to be the one where the weight of seeds started to remain constant. Based on these results, seeds of S. tomentosa and E. speciosa scarified in water at 98°C for 5min and 15min, respectively, were used in the experiments.
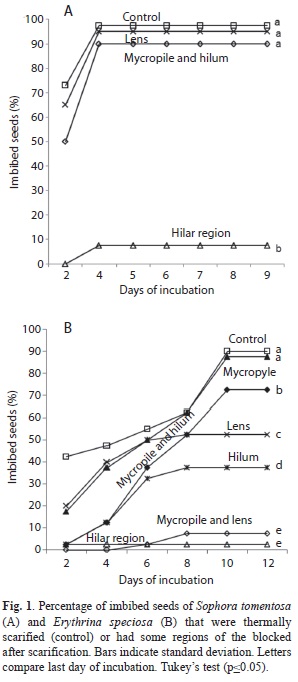
Identification of the site where water enters the seeds: Hot water scarified seeds had parts of the seed coats covered with Super Bonder® glue. Sophora tomentosa seed coats were blocked in the following regions: a) micropyle plus hilum and lens, b) micropyle plus hilum, and c) lens. Erythrina speciosa seed coats were blocked in the a) micropyle plus hilum plus lens, b) hilum, c) lens, d) micropyle, e) micropyle plus hilum, and f) micropyle plus lens. A control group was comprised of non-dormant, non-blocked seeds. Twenty seeds were utilized for each treatment. The seeds were placed in transparent plastic boxes (11x11x3.5cm) on two layers of filter paper with 10mL of distilled water. The boxes were stored at 25ºC with a photoperiod of 12h. Incubated seeds were counted at two-day intervals for 12 days.
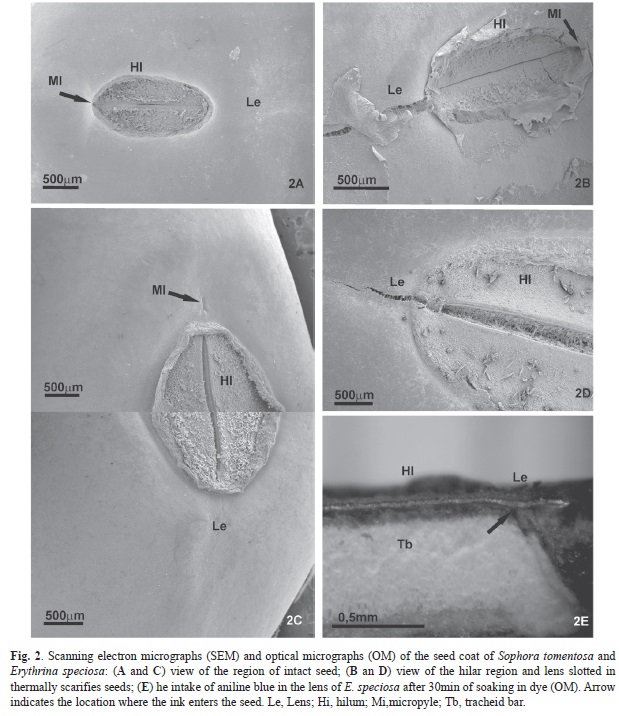
Seed coat features: The hilar region of five intact seeds of both species were cut with a scalpel and fixed in 2.5% glutaraldehyde, in a 0.1M sodium phosphate buffer at pH 7.2, and dehydrated in a graded ethanol series. Sections that were 40µm thick were cut using a sliding microtome. Histochemical tests were then made utilizing Sudan IV (2g/ ethanol 95% 100mL/gliceryne 5mL) for suberin, cutin, oils and waxes; acid phloroglucinol (1%) and iron chloride for lignin (Costa, 1982); and toluidine blue (0.05% toluidine blue en 100mL phosphate buffer 0.1M, pH 6.8) for polychromatic reactions to lignin (blue-green) and cellulose (reddish purple) (O’Brien, Feder, & McCully, 1965). The samples were examined under a Leica DMLS MPS 30 light microscope and images were taken with a Sony digital camera cyber-shot, DSC-W180. For SEM analyses, dehydrated pieces of five intact and five hot water scarified seeds (S. tomentosa and E. speciosa after 5min and 15min, respectively, in water at 98ºC) were dried according Horridge and Tamm (1969) with a CO2 critical point dryer (Leica EM CDP 300). The dried samples were adhered to aluminum stubs, with double-sided carbon tape, and coated with 20nm of gold using a Leica MS SCD500 sputter coater. Samples were observed and documented using a Jeol (model XL30 JSM 6390 LV) scanning electron microscope. To verify the presence of callose in the seeds, sections of non-fixed samples, from the hilar and extra-hilar regions of five intact seeds, were immersed in 0.05% aniline blue with a 0.1M potassium phosphate buffer at pH 8.3 (Ruzin, 1951). As a control, some sections were immersed only in the potassium phosphate buffer. The sections were observed using an Olympus BX41 microscope with a mercury vapor lamp (HBO 100) and a blue epifluorescence filter (UMWU2), at 330–385nm excitement and 420nm emission wavelengths. Images were taken with a Q-imaging digital camera (3.3 mpixel QColor 3C) and the software Q-captures Pro 5.1.
Dye tracking of water movement in the hilar pad: Forty seeds of each species (20 hot water scarified and 20 non-scarified) were immersed in an aqueous solution of 1% aniline blue (modified from Jayasuriya, Baskin, & Baskin, 2007). After 15min, 30min and 60min, five seeds of each treatment of each species were transversely and longitudinally sectioned through the hilum area. The sections (40µm thick) were made with a sliding microtome and observed under an optical microscope (Leica MPS 30 DMLS); images were taken with a Sony digital camera cyber-shot, DSC-W180.
A completely randomized design was used for the germination experiments for data analysis. Arcsine-transformed germination data were analyzed using one-way ANOVA with the software Bioestat (Ayres, Ayres, Ayres, & Santos, 2005). Tukey’s test was performed to compare treatments after ANOVA.
Results
Seed germination: For intact seeds of Sophora tomentosa and Erythrina speciosa incubated at 30ºC, the germination was around 13% and 2%, respectively, while 100% of the scarified seeds of both species germinated. For intact seeds of both species there was no germination at the lowest temperature tested (15°C) with the exception of 1.66% in one experiment using S. tomentosa (Table 1). At the upper limit temperature (35ºC) S. tomentosa seeds germinated but those of E. speciosa did not. The alternating temperatures studied did not promote germination of the species in relation to the constant temperature.
Identification of the site where water enters the seeds: Treatments utilizing glue to block the hilar region of scarified seeds showed, for both species, that the lens, micropyle and hilum are involved in water absorption (Fig. 1). In non-blocked seeds of S. tomentosa (Fig. 1a) treated with hot water, almost 100% absorbed water after 10 days, but when the hilar region was blocked only 10% of the seeds absorbed water. When the hilum-micropyle complex or when the lens was blocked, almost 90% of seeds absorbed water, showing that all of these structures need to be blocked for there to be a large reduction in water intake. In heat-treated and non-blocked seeds of E. speciosa (Fig. 1b) about 90% had imbibed water after 12 days, but when the hilar region was blocked only 10% of the seeds absorbed water. In seeds where the hilum, lens, and micropyle were blocked separately, 35%, 50% and around 90%, respectively, of the seeds imbibed water, which indicates that the hilum and lens were only partly effective in inhibiting water uptake and the micropyle was completely ineffective. However, during the first four days of incubation, blocking the micropyle greatly reduced water uptake compared to seeds without a blocked micropyle.
Seed coat features: In the seed coats of S. tomentosa and E. speciosa the hilum, lens and micropyle are in line, the lens and micropyle are on opposite sides of the hilum, and the middle of the hilum has a groove (Fig. 2a, Fig. 2b, Fig. 2c, Fig. 2d). In S. tomentosa the hilum covers the micropyle. The seed coat structure of the two species is very similar and consists of the extra-hilar region with one layer of columnar palisade cells, one layer of osteosclereids, spongy tissue and crushed cells. The columnar palisade cells are covered by a thick cuticle. In the hilar region there are two layers of palisade cells, the counterpalisade layer and, underneath, the palisade layer. A light line can be seen at the top of the palisade layer in both the hilar and extra-hilar regions. Tracheid bars are visible in the spongy tissue. Histochemical experiments showed that in the hilar region of both species callose is present in the palisade layer and lignin in the counterpalisade layer. In the extra-hilar region callose and lignin are also present in palisade layer, where lignin is mainly in the basal portion of the columnar cells and callose is mainly in the upper portion of these cells. The cuticle reacted positively to Sudan IV. The ultrastructural analysis of the hilar region of S. tomentosa (Fig. 2a) and E. speciosaseeds (Fig. 2c) revealed that in all non-treated seeds the tissues of this region remained intact with no disruptions. However, S. tomentosa seeds thermally scarified with hot water, showed cracks in the lens region (Fig 2b). In scarified seeds of E. speciosa the lens region ruptured and the hilum grove was more extended compared to the control (Fig. 2d). Dye treatments used to visualize the water movement in the hilar region were not conclusive for S. tomentosa; however, in seeds of E. speciosa the dye penetrated the lens and moved toward the vascular bundle (Fig. 2e).
Discussion
Studies of species from tropical forests have shown that alternating temperatures of 15ºC and 20ºC (minimum) and 30ºC and 40ºC (maximum) effectively broke the physical dormancy of seeds (Souza, Voltolini, Santos, & Paulilo, 2012; Vázquez-Yanes, & Orozco-Segovia, 1982). It is known that species exhibiting hard coat dormancy can differ consistently in their responsiveness to temperature fluctuations depending on the amplitude of temperature fluctuation, time of fluctuation and presence or not of cumulative effects of smaller fluctuations for a larger time (Moreno-Casasola, Grime, & Martínez, 1994). In the present work a maximum temperature fluctuation of 15ºC was applied during a period of 20 days and these conditions did not increase the germination of S. tomentosa or E. speciosa. Temperature fluctuation in Brazilian dunes can be over 25ºC (Franco et al., 1984) and the fluctuation used for the species in this work may be less than what is required or these species may require a longer period of fluctuation. Species that require a large fluctuation of temperature or require the cumulative effect of exposure to certain temperatures often only colonize bare soil and open areas and are prevented from becoming established in more stabilized areas covered with vegetation (Moreno-Casasola et al., 1994). This may be the case for the species in this work, but further study is needed on how the physical dormancy of these species is broken in the field, which would give more insight into how factors in natural conditions exercise a controlling influence on seed regeneration of these taxa.
The lens has been reported as being the site of initial water intake in physically dormant seeds of Fabaceae species (Baskin, Baskin, & Li, 2000; Morrison, McClay, Porter and Rish, 1998; Serrato-Valenti, Vries, & Cornara, 1995; Souza et al., 2012). However, in seeds of S. tomentosa, blocking only the lens or the micropyle-hilum complex does not prevent the entry of water into a scarified seed, which indicates that both structures are equally related to the entry of water. This is further supported because there is a significant inhibition of the entry of water into the scarified seed only when these two structures are blocked. The SEM analysis of E. speciosa showed that in scarified seeds the lens ruptured, exposing partially separated macrosclereids, and the opening of the hilar fissure was wider than in non-scarified seeds. Furthermore, the immersion of scarified seeds into an aniline aqueous solution showed that the solution first entered the seed through the hilum. In agreement with these results, (Hu, Wang, Wu, Nan, & Baskin, 2008) observed in Sophora alopecuroides evidence of water intake through the hilum after sulphuric acid scarification of seeds (which showed a wider hilum fissure than non-scarified seeds) and in seeds buried in the field. However, they observed that depending on the time exposed to sulphuric acid, or in the field, the lens and extra-hilar region also become permeable to water. Another structure that has been mentioned as being important to the entrance of water in seeds of Cassia (Fabaceae), which are physically dormant, is the micropyle (Bhattacharya, & Saha, 1990; Paula, Delgado, Paulilo, & Santos, 2012). For S. tomentosa and E. speciosa in the present work, it was interesting to notice that when only the micropyle was blocked, in scarified seeds, the water uptake was inhibited until the second and fourth day (respectively) of seed incubation. This could be explained by the fact that the hilum and the lens are also involved in water absorption, but participate less than the micropyle at the beginning of imbibition, as proposed by Tailor (2005) for physically dormant Fabaceae seeds from Australian pastures. Difference in water uptake between different water gap has been also noted by Hu, Wang, Wu, Nan, and Baskin (2008) and Hu, Wang,Wu, and Baskin (2009). An interesting aspect of water imbibition observed in the present work, and already observed by Hu et al. (2008) and Hut et al. (2009), was the evidence that the water inlet region can be different between initial and subsequent soaking days.
For both species studied, the major features of the seed coat were similar, despite the differences in humidity and sunlight that occur where they grow. In fact, as discussed by Souza and Marcos-Filho (2001), the morphological features of the Fabaceae seed coat are relatively insensitive to environmental conditions and contain distinct taxonomic features. Both species showed seed morphological and anatomical features for seed coats of the subfamily Faboideae, as previously described by Corner (1976), Kelly, Van Staden, and Bell (1992) and Kirkbride, Gunn and Weitzman (2003), such as the lens close to the hilum, the hilum completely visible, and a funicular remnant (rim aril) that is split down the middle lengthwise. In S. tomentosa the micropyle is covered by the hilum as observed by Hu et al. (2008) in S. alopecuroides. Lignin and callose, found around all palisade layer(s), are reported to be impermeable to water and would be responsible for the impediment of water absorption by the physically dormant seeds of Fabaceae (Baskin et al., 2000; Ma,Cholewa, Mohamed , Peterson, & Gijzen, 2004; Varela, & Albornoz 2013) suggest the permeability of Anadenanthera colubrina var. cebil seeds to water is because of the absence of lignin in the palisade layer. Lignin is also ecologically important because it protects the seed against predation (Souza and Marcos Filho, 2001). According to (Dalling Davis, Schutte, & Arnold, 1994), the substances that confer hardness to physical dormancy also promote long-term seed persistence and provide an effective barrier against microbial access, particularly in warm and moist environments where conditions are most conducive to fungal growth. The need for protection against predation occur in seeds of S. tomentosa because the fruits of this species are indehiscent and remain on the plant for several months, which makes them and the seeds within them more susceptible to subsequent predations (Nogueira, & Arruda, 2006b). In the case of E. speciosa, a species found in wet areas with autochorous seed dispersal, the seeds can fall to the wet ground and stay there for a long time. In this case, the presence of lignin is very important because it prevents microbial access. Strong seed coat impermeability also protects against deterioration as seeds age (Brancalion, Novembre, Rodrigues, & Marcos Filho, 2010).
Acknowledgments
This study received financial support from Coordenação de Aperfeiçoamento do Ensino Superior (CAPES), Brazil.
References
Argel, P. J., & Paton, C. J. (1999). Overcoming legume hardseedness. In D. S., Loch, & J. E. Ferguson (Orgs.), Forage seed production: Tropical and sub-tropical species (247-265). Wallingford: CAB International. [ Links ]
Ayres, M., Ayres Junior, M., Ayres, D. L., & Santos, A. S. (2007). BioEstat 5.0: Aplicações estatísticas nas áreas das ciências biomédicas. Belém: Sociedade Civil Mamirauá [ Links ].
Baskin, C. C., & Baskin, J. M. (2001). Seeds: Ecology, biogeography, and evolution of dormancy and germination. San Diego: Academic Press. [ Links ]
Baskin, J. M., Baskin, C. C., & Li, X. (2000). Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biology, 15,139-152. doi: 10.1046/j.1442-1984.2000.00034.x. [ Links ]
Bechara, F. C. (2003). Restauração ecológica de restingas contaminadas por Pinus no Parque Florestal do Rio Vermelho (Dissertação inédita de Mestrado). Universidade Federal de Santa Catarina, Florianópolis, Brasil. [ Links ]
Bewley, J. D., & Black, M. (1994). Seeds: physiology of development and germination. New York: Plenum Publishing. [ Links ]
Bhattacharya, A., & Saha, P. K. (1990). Ultrastructure of seed coat and water uptake pattern of seeds during germination in Cassia sp. Seed Science and Technolgy, 18(1), 97-103. [ Links ]
Brancalion, P. S. H., Novembre, A. D. L. C., Rodrigues, R. R., & Marcos Filho, J. (2010). Dormancy as exaptation to protect mimetic seeds against deterioration before dispersal. Annals of Botany, 105(6), 991-998. [ Links ]
Brito, V. L. G., Pinheiro, M., & Sazima, M. (2010). Sophora tomentosa e Crotalaria vitellina (Fabaceae): biologia reprodutiva e interações com abelhas na restinga de Ubatuba, São Paulo. Biota Neotropica, 10(1), 185-192. [ Links ]
Corner, E. J. H. (1976). The seeds of Dicotyledons. London: University Press. [ Links ]
Costa, A. F. (1982). Farmacognosia. Lisboa: Fundação Calouste Gulbenkian. [ Links ]
Dalling, J. W., Davis, A. S., Schutte, B. J., & Arnold, A. E. (2011). Seed survival in soil: interacting effects of predation, dormancy and the soil microbial community. Journal of Ecology, 99(1) 89-95. [ Links ]
Franco, A. C., Valeriano, D. M., Santos, F. M., Hay, J. D., Henriques, R. P. B., & Medeiros, R. A. (1984). Os microclimas das zonas de vegetação da praia da restinga de Barra do Maricá, Rio de Janeiro (pp. 431-425). In L. D. Lacerda, D. S. D. Araújo, R. Cerqueira, & B. Turcq (Orgs.). Restingas: Origem, estrutura, processos. Niterói: CEUFF. [ Links ]
Gama-Arachchige, N. S., Baskin, J. M., Geneve, R. L., & Baskin, C. C. (2010). Identification and characterization of the water gap in physically dormant seeds of Geraniaceae, with special reference to Geranium carolinianum. Annals of Botany, 105, 997-990. doi: 10.1093/aob/mcq078. [ Links ]
Gama-Arachchige, N. S., Baskin, J. M., Geneve, R. L., & Baskin, C. C. (2013). Identification and characterization of ten new water gaps in seeds and fruits with physical dormancy and classification of water-gap complexes. Annals of Botany, 112, 69-84. doi: 10.1093/aob/mct094. [ Links ]
Horridge, G. A., & Tamm, S. L. (1969). Critical point drying for scanning electron microscopy study of ciliarmotion. Science, 3869, 817-818. [ Links ]
Hu, X. W., Wang, Y. R., Wu, Y. P., Nan, Z. B., & Baskin, C. C. (2008). Role of the lens in physical dormancy in seeds of Sophora alopecuroides L. (Fabaceae) from north-west China. Australian Journal of Agricultural Research, 59, 491-497. doi.org/10.1071/AR07265. [ Links ]
Hu, X. W., Wang, Y. R., Wu, Y. P., & Baskin, C. C. (2009). Role of the lens in controlling water uptake in seeds of two Fabaceae (Papilonoideae) species treated with sulphuric acid and hot water. Seed Science Research, 19, 73-80. doi: 10.1017/S0960258509301099. [ Links ]
Jayasuriya, K. M. G. G., Baskin, C. C., & Baskin, J. M. (2007). Morphology and anatomy of physical dormancy in Ipomoea lacunosa: identification of the water gap in seeds of Convolvulaceae (Solanales). Annals of Botany, 100, 13-21. doi: 10.1093oab/mcm0708. [ Links ]
Jayasuriya, K. M. G. G., Baskin, J. M., Geneve, R. L., Baskin, C. C., & Chien, C. T. (2008). Physical Dormancy in seeds of the holoparasitic angiosperm Cuscuta australis(Convolvulaceae, Cuscuteae):Dormancy-breaking requirements, anatomy of the water gap and sensitivity cycling. Annals of Botany, 102, 39-48. doi: 10.1093/aob/mcn064. [ Links ]
Jayasuriya, K. M. G. G., Wijetunga, A. S. T. B., Baskin, J. M., & Baskin, C. C. (2010). Recalcitrancy and a new kind of epicotyl dormancy in seeds of the understory tropical rainforest tree humboldtia laurifolia (Fabaceae, Ceasalpinioideae). American Journal of Botany, 97(1), 15-26. [ Links ]
Judd, W. S., Campbel, C. S., Kellongg, E. A., Steens P. F., & Donogue, M. J. (2009). Sistemática Vegetal: um enfoque filogenético. Porto Alegre: Artmed. [ Links ]
Kelly, K. M.,Van Staden, J., & Bell, W. E. (1992). Seed coat structure and dormancy. Plant growth Regulation, 11, 201-209. doi:10.1007/BF 00024559. [ Links ]
Kirkbride, J. Jr, Gunn C., & Weitzman A. (2003). Fruits and seeds of genera in subfamily Faboideae (Fabaceae). Technical Bulletin of the United States Department of Agriculture, 1890, 1-1212. [ Links ]
Klein, R. M. (1969). Árvores nativas da Ilha de Santa Catarina. Insula, 3, 3-93. [ Links ]
Kondo, T., & Takahashi, K. (2004). Breaking of physical dormancy and germination ecology for seeds of Thermopsis lupinoides Link. Journal of the Japanese Society of Revegetation Technology, 30(1), 163-168. [ Links ]
Lorenzi, H. (2002). Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas do Brasil. Nova Odessa: Editora Plantarum. [ Links ]
Lima, H. C. (2000). Leguminosas da Mata Atlântica: uma análise da riqueza, padrões de distribuição geográfica e similaridades florísticas em remanescentes florestais do estado do Rio de Janeiro (Tese inédita de Doutorado). Universidade Federal do Rio de Janeiro, Brasil. [ Links ]
Ma, F., Cholewa, W., Mohamed, T., Peterson, C. A., & Gijzen, M. (2004). Cracks in the palisade cuticle of soybean seed coats correlate with their permeability to water. Annals of Botany, 94, 213-228. doi:10.1093/aob/mch133. [ Links ]
Medina, C. L., Sanches, M. C., Tucci, M. L., Sousa, C. A. F., Cuzzuol, G. R. F., & Joly C. A. (2009). Erythrina speciosa (Leguminosae-Papilionoideae) under soil water saturation: morphophysiological and growth responses. Annals of Botany, 104, 671-680. doi: 10.1093/aob/mcp159. [ Links ]
Myers, N., Mittermeier, R. A., Mittermeir, C. G., da Fonseca, G. A., & Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature, 403, 853-858. doi: 10.1038/35002501. [ Links ]
Moreno-Casasola, P., Grime, J. P, & Martínez, L. (1994). A comparative study of the effects of fluctuations in temperature and moisture supply on hard coat dormancy in seeds of coastal tropical legumes in Mexico. Journal of Tropical Ecology, 10, 67-86. doi: 10.1017/S0266467400007720. [ Links ]
Morrison, D. A., McClay, K., Porter, C., & Rish, S. (1998). The role of the lens in controlling heat-induced breakdown of testa-imposed dormancy in native Australian legumes. Annals of Botany, 82, 5-40. doi:10.1006/anbo.1998.0640. [ Links ]
Nogueira, E. M. L., & Arruda, V. L. V. (2006a). Fenologia reprodutiva, polinização e sistema reprodutivo de Sophora tomentosa L. (Leguminosae – Papilionoideae) em restinga da praia da Joaquina, Florianópolis, sul do Brasil. Biotemas, 19(2), 29-36. [ Links ]
Nogueira, E. M. L., & Arruda, V. L. V. (2006b). Frutificação e danos em frutos e sementes de Sophora tomentosa L. (Leguminosae, Papilionoideae) em restinga da praia da Joaquina, Florianópolis, SC. Biotemas, 19(4), 41-48. [ Links ]
O’Brien, T. P., Feder, N., & McCully, M. E. (1965). Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma, 59, 368-373. [ Links ]
Paula, A. S., Delgado, C. M. L., Paulilo, M. T. S., & Santos, M. (2012). Breaking physical dormancy of Cassia leptophylla and Senna macranthera (Fabaceae: Caesalpinioideae) seeds: water absorption and alternating temperatures. Seed Science Research, 22, 259-257. doi:10.1017/S096025851200013X. [ Links ]
Ruzin, S. E. (1951). Plant microtechnique and microscopy. New York: Oxford University Press. [ Links ]
Serrato-Valenti, G., De Vries, M., & Cornara, L. (1995). The hilar region in Leucaena leucocephala Lam. (De Wit) seed: structure, histochemistry and the role of the lens in germination. Annals of Botany, 75(6), 569-574. [ Links ]
Souza, F. H. D., & Marcos Filho, J. (2001). The seed coat as a modulator of seed-environment relationships in Fabaceae. Revista Brasileira de Botanica, 24, 365-375. [ Links ]
Souza, T. V., Voltolini, C. H., Santos, M., & Paulilo, M. T. S. (2012). Water absorption and dormancy-breaking requirements of physically dormant seeds of Schizolobium parahyba (Fabaceae-Caesalpinioideae). Seed Science Research, 22, 169-176. doi:10.1017/S0960258512000013. [ Links ]
Sprent, J. I. (2001). Nodulation in legumes. Kew, Royal Botanic Gardens. [ Links ]
Varela, R.O., & Albornoz, P. L. (2013). Morpho-anatomy, imbibition, viability and germination of the seed of Anadenanthera colubrina var. cebil (Fabaceae). Revista Biologia Tropical, 61(3), 1109-1118. [ Links ]
Vázquez-Yanes, C., & Orozco-Segovia, A. (1982). Seed germination of a tropical rain forest pioneer tree (Heliocarpus donnell-smithii) in response to diurnal fluctuation of temperature. Physiologia plantarum, 56, 295-298. doi.10.1111/j.1399-3054.1982tb00341.x [ Links ]
Ayres, M., Ayres Junior, M., Ayres, D. L., & Santos, A. S. (2007). BioEstat 5.0: Aplicações estatísticas nas áreas das ciências biomédicas. Belém: Sociedade Civil Mamirauá [ Links ].
Baskin, C. C., & Baskin, J. M. (2001). Seeds: Ecology, biogeography, and evolution of dormancy and germination. San Diego: Academic Press. [ Links ]
Baskin, J. M., Baskin, C. C., & Li, X. (2000). Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biology, 15,139-152. doi: 10.1046/j.1442-1984.2000.00034.x. [ Links ]
Bechara, F. C. (2003). Restauração ecológica de restingas contaminadas por Pinus no Parque Florestal do Rio Vermelho (Dissertação inédita de Mestrado). Universidade Federal de Santa Catarina, Florianópolis, Brasil. [ Links ]
Bewley, J. D., & Black, M. (1994). Seeds: physiology of development and germination. New York: Plenum Publishing. [ Links ]
Bhattacharya, A., & Saha, P. K. (1990). Ultrastructure of seed coat and water uptake pattern of seeds during germination in Cassia sp. Seed Science and Technolgy, 18(1), 97-103. [ Links ]
Brancalion, P. S. H., Novembre, A. D. L. C., Rodrigues, R. R., & Marcos Filho, J. (2010). Dormancy as exaptation to protect mimetic seeds against deterioration before dispersal. Annals of Botany, 105(6), 991-998. [ Links ]
Brito, V. L. G., Pinheiro, M., & Sazima, M. (2010). Sophora tomentosa e Crotalaria vitellina (Fabaceae): biologia reprodutiva e interações com abelhas na restinga de Ubatuba, São Paulo. Biota Neotropica, 10(1), 185-192. [ Links ]
Corner, E. J. H. (1976). The seeds of Dicotyledons. London: University Press. [ Links ]
Costa, A. F. (1982). Farmacognosia. Lisboa: Fundação Calouste Gulbenkian. [ Links ]
Dalling, J. W., Davis, A. S., Schutte, B. J., & Arnold, A. E. (2011). Seed survival in soil: interacting effects of predation, dormancy and the soil microbial community. Journal of Ecology, 99(1) 89-95. [ Links ]
Franco, A. C., Valeriano, D. M., Santos, F. M., Hay, J. D., Henriques, R. P. B., & Medeiros, R. A. (1984). Os microclimas das zonas de vegetação da praia da restinga de Barra do Maricá, Rio de Janeiro (pp. 431-425). In L. D. Lacerda, D. S. D. Araújo, R. Cerqueira, & B. Turcq (Orgs.). Restingas: Origem, estrutura, processos. Niterói: CEUFF. [ Links ]
Gama-Arachchige, N. S., Baskin, J. M., Geneve, R. L., & Baskin, C. C. (2010). Identification and characterization of the water gap in physically dormant seeds of Geraniaceae, with special reference to Geranium carolinianum. Annals of Botany, 105, 997-990. doi: 10.1093/aob/mcq078. [ Links ]
Gama-Arachchige, N. S., Baskin, J. M., Geneve, R. L., & Baskin, C. C. (2013). Identification and characterization of ten new water gaps in seeds and fruits with physical dormancy and classification of water-gap complexes. Annals of Botany, 112, 69-84. doi: 10.1093/aob/mct094. [ Links ]
Horridge, G. A., & Tamm, S. L. (1969). Critical point drying for scanning electron microscopy study of ciliarmotion. Science, 3869, 817-818. [ Links ]
Hu, X. W., Wang, Y. R., Wu, Y. P., Nan, Z. B., & Baskin, C. C. (2008). Role of the lens in physical dormancy in seeds of Sophora alopecuroides L. (Fabaceae) from north-west China. Australian Journal of Agricultural Research, 59, 491-497. doi.org/10.1071/AR07265. [ Links ]
Hu, X. W., Wang, Y. R., Wu, Y. P., & Baskin, C. C. (2009). Role of the lens in controlling water uptake in seeds of two Fabaceae (Papilonoideae) species treated with sulphuric acid and hot water. Seed Science Research, 19, 73-80. doi: 10.1017/S0960258509301099. [ Links ]
Jayasuriya, K. M. G. G., Baskin, C. C., & Baskin, J. M. (2007). Morphology and anatomy of physical dormancy in Ipomoea lacunosa: identification of the water gap in seeds of Convolvulaceae (Solanales). Annals of Botany, 100, 13-21. doi: 10.1093oab/mcm0708. [ Links ]
Jayasuriya, K. M. G. G., Baskin, J. M., Geneve, R. L., Baskin, C. C., & Chien, C. T. (2008). Physical Dormancy in seeds of the holoparasitic angiosperm Cuscuta australis(Convolvulaceae, Cuscuteae):Dormancy-breaking requirements, anatomy of the water gap and sensitivity cycling. Annals of Botany, 102, 39-48. doi: 10.1093/aob/mcn064. [ Links ]
Jayasuriya, K. M. G. G., Wijetunga, A. S. T. B., Baskin, J. M., & Baskin, C. C. (2010). Recalcitrancy and a new kind of epicotyl dormancy in seeds of the understory tropical rainforest tree humboldtia laurifolia (Fabaceae, Ceasalpinioideae). American Journal of Botany, 97(1), 15-26. [ Links ]
Judd, W. S., Campbel, C. S., Kellongg, E. A., Steens P. F., & Donogue, M. J. (2009). Sistemática Vegetal: um enfoque filogenético. Porto Alegre: Artmed. [ Links ]
Kelly, K. M.,Van Staden, J., & Bell, W. E. (1992). Seed coat structure and dormancy. Plant growth Regulation, 11, 201-209. doi:10.1007/BF 00024559. [ Links ]
Kirkbride, J. Jr, Gunn C., & Weitzman A. (2003). Fruits and seeds of genera in subfamily Faboideae (Fabaceae). Technical Bulletin of the United States Department of Agriculture, 1890, 1-1212. [ Links ]
Klein, R. M. (1969). Árvores nativas da Ilha de Santa Catarina. Insula, 3, 3-93. [ Links ]
Kondo, T., & Takahashi, K. (2004). Breaking of physical dormancy and germination ecology for seeds of Thermopsis lupinoides Link. Journal of the Japanese Society of Revegetation Technology, 30(1), 163-168. [ Links ]
Lorenzi, H. (2002). Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas do Brasil. Nova Odessa: Editora Plantarum. [ Links ]
Lima, H. C. (2000). Leguminosas da Mata Atlântica: uma análise da riqueza, padrões de distribuição geográfica e similaridades florísticas em remanescentes florestais do estado do Rio de Janeiro (Tese inédita de Doutorado). Universidade Federal do Rio de Janeiro, Brasil. [ Links ]
Ma, F., Cholewa, W., Mohamed, T., Peterson, C. A., & Gijzen, M. (2004). Cracks in the palisade cuticle of soybean seed coats correlate with their permeability to water. Annals of Botany, 94, 213-228. doi:10.1093/aob/mch133. [ Links ]
Medina, C. L., Sanches, M. C., Tucci, M. L., Sousa, C. A. F., Cuzzuol, G. R. F., & Joly C. A. (2009). Erythrina speciosa (Leguminosae-Papilionoideae) under soil water saturation: morphophysiological and growth responses. Annals of Botany, 104, 671-680. doi: 10.1093/aob/mcp159. [ Links ]
Myers, N., Mittermeier, R. A., Mittermeir, C. G., da Fonseca, G. A., & Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature, 403, 853-858. doi: 10.1038/35002501. [ Links ]
Moreno-Casasola, P., Grime, J. P, & Martínez, L. (1994). A comparative study of the effects of fluctuations in temperature and moisture supply on hard coat dormancy in seeds of coastal tropical legumes in Mexico. Journal of Tropical Ecology, 10, 67-86. doi: 10.1017/S0266467400007720. [ Links ]
Morrison, D. A., McClay, K., Porter, C., & Rish, S. (1998). The role of the lens in controlling heat-induced breakdown of testa-imposed dormancy in native Australian legumes. Annals of Botany, 82, 5-40. doi:10.1006/anbo.1998.0640. [ Links ]
Nogueira, E. M. L., & Arruda, V. L. V. (2006a). Fenologia reprodutiva, polinização e sistema reprodutivo de Sophora tomentosa L. (Leguminosae – Papilionoideae) em restinga da praia da Joaquina, Florianópolis, sul do Brasil. Biotemas, 19(2), 29-36. [ Links ]
Nogueira, E. M. L., & Arruda, V. L. V. (2006b). Frutificação e danos em frutos e sementes de Sophora tomentosa L. (Leguminosae, Papilionoideae) em restinga da praia da Joaquina, Florianópolis, SC. Biotemas, 19(4), 41-48. [ Links ]
O’Brien, T. P., Feder, N., & McCully, M. E. (1965). Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma, 59, 368-373. [ Links ]
Paula, A. S., Delgado, C. M. L., Paulilo, M. T. S., & Santos, M. (2012). Breaking physical dormancy of Cassia leptophylla and Senna macranthera (Fabaceae: Caesalpinioideae) seeds: water absorption and alternating temperatures. Seed Science Research, 22, 259-257. doi:10.1017/S096025851200013X. [ Links ]
Ruzin, S. E. (1951). Plant microtechnique and microscopy. New York: Oxford University Press. [ Links ]
Serrato-Valenti, G., De Vries, M., & Cornara, L. (1995). The hilar region in Leucaena leucocephala Lam. (De Wit) seed: structure, histochemistry and the role of the lens in germination. Annals of Botany, 75(6), 569-574. [ Links ]
Souza, F. H. D., & Marcos Filho, J. (2001). The seed coat as a modulator of seed-environment relationships in Fabaceae. Revista Brasileira de Botanica, 24, 365-375. [ Links ]
Souza, T. V., Voltolini, C. H., Santos, M., & Paulilo, M. T. S. (2012). Water absorption and dormancy-breaking requirements of physically dormant seeds of Schizolobium parahyba (Fabaceae-Caesalpinioideae). Seed Science Research, 22, 169-176. doi:10.1017/S0960258512000013. [ Links ]
Sprent, J. I. (2001). Nodulation in legumes. Kew, Royal Botanic Gardens. [ Links ]
Varela, R.O., & Albornoz, P. L. (2013). Morpho-anatomy, imbibition, viability and germination of the seed of Anadenanthera colubrina var. cebil (Fabaceae). Revista Biologia Tropical, 61(3), 1109-1118. [ Links ]
Vázquez-Yanes, C., & Orozco-Segovia, A. (1982). Seed germination of a tropical rain forest pioneer tree (Heliocarpus donnell-smithii) in response to diurnal fluctuation of temperature. Physiologia plantarum, 56, 295-298. doi.10.1111/j.1399-3054.1982tb00341.x [ Links ]
1. Departamento de Botânica, Universidade Federal de Santa Catarina, Florianópolis-SC, Brasil; delgado_carol@yahoo.com.br, alexandredepaula_07@hotmail.com, marisa.santos@ufsc.br, paulilo@ccb.ufsc.br
Received 18-III-2014. Corrected 08-IX-2014. Accepted 10-X-2014.


{kind=link}
{kind=link}
{kind=link}