










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.1 San José Jan./Mar. 2015
Acute toxicity and sublethal effects of the mixture glyphosate (Roundup® Active) and Cosmo-Flux®411F to anuran embryos and tadpoles of four Colombian species
Toxicidad aguda y efectos subletales de la mezcla glifosato (Roundup® Activo) y Cosmo-Flux® 411F en embriones y renacuajos de cuatro especies de anuros colombianos
Toxicidad aguda y efectos subletales de la mezcla glifosato (Roundup® Activo) y Cosmo-Flux® 411F en embriones y renacuajos de cuatro especies de anuros colombianos
Abstract
Glyphosate is the most widely used herbicide in the world with application in agriculture, forestry, industrial weed control, garden and aquatic environments. However, its use is highly controversial for the possible impact on not-target organisms, such as amphibians, which are vanishing at an alarming and rapid rate. Due to the high solubility in water and ionic nature, the glyphosate requires of surfactants to increase activity. In addition, for the control of coca (Erythroxylum coca) and agricultural weeds in Colombia, formulated glyphosate is mixed and sprayed with the adjuvant Cosmo-Flux®411F to increase the penetration and activity of the herbicide. This study evaluates the acute toxic and sublethal effects (embryonic development, tadpole body size, tadpole swimming performance) of the mixture of the formulated glyphosate Roundup® Active and Cosmo-Flux®411F to anuran embryos and tadpoles of four Colombian species under 96h laboratory standard tests and microcosms, which are more similar to field conditions as they include soil, sand and macrophytes. In the laboratory, embryos and tadpoles of Engystomops pustulosus were the most tolerant (LC50=3 904µg a.e./L; LC50=2 799µg a.e./L, respectively), while embryos and tadpoles of Hypsiboas crepitans (LC50=2 203µg a.e./L; LC50=1 424µg a.e./L, respectively) were the most sensitive. R. humboldti and R. marina presented an intermediate toxicity. Embryos were significantly more tolerant to the mixture than tadpoles, which could be likely attributed to the exclusion of chemicals by the embryonic membranes and the lack of organs, such as gills, which are sensitive to surfactants. Sublethal effects were observed for the tadpole body size, but not for the embryonic development and tadpole swimming performance. In microcosms, no toxicity (LC50 could not be estimated), or sublethal responses were observed at concentrations up to fourfold (14.76kg glyphosate a.e./ha) the highest field application rate of 3.69kg glyphosate a.e./ha. Thus, toxicity was less in the microcosms than in laboratory tests, which may be attributed to the presence of sediments and organic matter which rapidly adsorb glyphosate and surfactants such as POEA. It is concluded that the mixture of glyphosate (Roundup® Active) and Cosmo-Flux®411F, as used in the field, has a negligible toxic effect to embryos and tadpoles of the species tested in this study.
Key words: amphibians, ecotoxicology, herbicides, surfactants, survival.
Resumen
El glifosato es el herbicida más usado en el mundo con aplicaciones para la agricultura, control de malezas forestales, industriales, en jardines y ambientes acuáticos. Sin embargo, su uso es altamente controversial por el posible impacto sobre organismos no blanco, como los anfibios, los cuales están desapareciendo a una tasa alarmantemente rápida. Debido a su alta solubilidad en agua y naturaleza iónica, el glifosato requiere de surfactantes para incrementar su actividad. Además, para el control de Erythroxylum coca y de malezas en la agricultura en Colombia, el glifosato formulado es mezclado y rociado con el coadyuvante Cosmo-Flux®411F para incrementar la penetración y actividad del herbicida. Este estudio evalúa los efectos tóxicos agudos y subletales (desarrollo embrionario, tamaño corporal y desempeño natatorio de los renacuajos) de la mezcla del glifosato formulado Roundup® Activo con el Cosmo-Flux®411F en embriones y renacuajos de cuatro especies de anuros colombianos, bajo pruebas de 96h en condiciones estándar de laboratorio y microcosmos, que son más similares a las condiciones de campo al incluir tierra, arena y macrófitas. En laboratorio, los embriones y renacuajos de Engystomops pustulosus fueron los más tolerantes (CL50=3 904µg a.e./L; CL50=2 799µg a.e./L, respectivamente), mientras que los embriones y renacuajos de Hypsiboas crepitans fueron los más sensibles (CL50=2 203µg a.e./L; CL50=1 424µg a.e./L, respectivamente). R. humboldti y R. marina presentaron una toxicidad intermedia. Los embriones fueron más tolerantes a la mezcla que los renacuajos, lo cual podría ser atribuido a la exclusión de los químicos por las membranas embrionarias y a la falta de órganos, como las branquias, que son más sensibles a los surfactantes. Se observaron efectos subletales en el tamaño corporal de los renacuajos, pero no en el desarrollo embrionario ni el desempeño natatorio de los renacuajos. En microcosmos no se observaron efectos tóxicos ni respuestas subletales a concentraciones hasta cuatro veces (14.67kg glifosato a.e./ha) la tasa de aplicación más alta de 3.69kg glifosato a.e./ha. Por lo tanto, la toxicidad fue menor en los microcosmos que en las pruebas de laboratorio, lo que puede ser atribuido a la presencia de sedimentos y materia orgánica que absorbe rápidamente el glifosato y surfactantes como el POEA. Se concluye que la mezcla del glifosato (Roundup® Activo) y Cosmo-Flux®411F, como se aplica en campo, tiene un efecto tóxico bajo en los embriones y renacuajos de las especies estudiadas.
Palabras clave: anfibios, ecotoxicología, herbicidas, supervivencia, surfactantes.
Glyphosate is the most commonly used herbicide in the world with many commercial formulations for application in agriculture (Duke & Powles, 2008; Plötner & Matschke, 2012). However, the use of glyphosate is also highly controversial for its possible impact on not-target organisms. In Colombia, the mixture of formulated glyphosate and the adjuvant Cosmo-Flux®411F is used to eradicate coca, poppy crops (Solomon et al., 2007) and for the control of agricultural weeds (Solomon, Anadón, Cerdeira, Marshall, & Sanin, 2005). Nevertheless, during the aerial or hand spraying, this mix can affect aquatic animals such as amphibians, many of which are threatened with extinction (Mattoon, 2000; Collins & Storfer, 2003; Storfer, 2003). The life cycle and physiological characteristics of amphibians make them particularly susceptible to all environmental stressors (Cowman & Mazanti, 2000). Glyphosate formulations have been considered to induce genotoxic, morphological, biochemical, and physiological effects to anurans (Clements, Ralph, & Petras, 1997; Lajmanovich, Lorenzatti, Maitre, Enrique, & Peltzer, 2003; Cauble & Wagner, 2005; Bernal, Solomon, & Carrasquilla, 2009a; Mann, Hyne, Choung, & Wilson, 2009; Lenkowski, Sanchez-Bravo, & Mclaughlin, 2010; Williams & Semlitsch, 2010; Lajmanovich, Attademo, Peltzer, Junges, & Cabagna, 2011; Relyea, 2012), although low toxicity has been reported under realistic field conditions (Howe et al., 2004; Wojtaszek, Staznik, Chartrand, Stephenson, & Thompson, 2004; Bernal, Solomon, & Carrasquilla, 2009b). Other studies have concluded that the glyphosate and its formulated products represent moderate (Giesy, Dobson, & Solomon, 2000; Sparling, Linder, Bishop, & Krest, 2010) or negligible risk to aquatic organisms (World Health Organization International Program on Chemical Safety, 1994; Giesy et al., 2000; Solomon & Thompson, 2003). These differences could be attributed not only to experimental conditions, but also to types of commercial products tested. Overall, there is a strong debate about the impact of glyphosate on amphibians. Glyphosate is the active ingredient of several herbicide formulations, which contains surfactants such as the ethoxylated tallowamine (POEA) that vary in concentration and may have a greater toxicity than glyphosate itself (Bradberry, Proudfoot, & Vale, 2004). The glyphosate acts as an inhibitor of enzymes involved in the synthesis of aromatic amino acids in growing plants, and must be absorbed through foliage; however, due to its high solubility in water and ionic nature, it does not penetrate easily through the hydrophobic cuticle of leaves. Therefore, surfactants are used in the formulated commercial products. In addition, for the control of coca in Colombia, formulated glyphosate is mixed and sprayed with 2.3% v/v of the adjuvant Cosmo-Flux®411F, which reduce the surface tension of leaves to increase the penetration and activity of the herbicide in coca plants (Solomon et al., 2007). The rate of application of formulated glyphosate is 3.69kg a.e./ha, which is greater than that recommended in agriculture (1.77kg a.e./ha) (Ministry of Environment, Housing and Territorial Development of Colombia, 2007). The toxic effect of Cosmo-Flux®411F has been recently evaluated in Colombian anuran embryos by Henao, Arango and Bernal (2013), where they found that this adjuvant did not exert any lethal consequence at the concentrations sprayed in the field. Another study conducted by Rondón, Ramírez and Eslava (2007) in the fish Piaractus brachypomus, concluded that the Cosmo-Flux®411F produced anatomopathological effects and mortality, but at much larger concentrations (LC50=4 418mg/L) than those applied in the field.
Toxicity studies assessing the effect of glyphosate formulations on amphibians have been mainly performed on larval stages (see review from Plötner & Matschke, 2012), less on juveniles and adults (Mann & Bidwell, 1999; Relyea, 2005; Bernal et al., 2009b), and a few on embryonic stages. For example, Edginton, Sheridan, Stephenson, Thompson and Boermans (2004) evaluated the interactive effect of pH and the glyphosate (Vision®) in embryos and larvae of Rana clamitans, Rana pipiens, Bufo americanus and Xenopus laevis. They reported that larvae were more sensitive to glyphosate than embryos. Triana, Montes and Bernal (2013), on the other hand, evaluated the effect of glyphosate (Roundup® Active) to anuran embryos of four Colombian species, and showed that it was highly toxic in laboratory experiments but less in microcosm conditions. Despite these data, there is no still information about the toxicity of the combined glyphosate and Cosmo-Flux®411F, as sprayed in field, to embryos from Colombian anurans. Bernal et al. (2009a,b) carried out a previous work on the effect of this mix to anuran larvae, juveniles and adults, but they used the formulated glyphosate GLY-41 which is applied to eradicate illicit plantations such as coca (Erythroxylum coca) and opium poppy (Papaver somniferum).
This study was conducted to evaluate the acute toxicity and sublethal effects (embryonic development, tadpole body size, tadpole swimming performance) of the mixture of Roundup® Active and the adjuvant Cosmo-Flux®411F to embryos and tadpoles of four Colombian anuran species. Roundup® Active was selected for this study as this herbicide is widely used in agriculture in Colombia to remove weeds of rice, cotton, soybean and corn fields, which may be used as habitat by anurans.
Materials and Methods
Test organisms: Between two and four freshly egg masses for each study species were collected in the Department of Tolima, Colombia, as follows: Rhinella humboldti (Gallardo, 1965) in Payandé (04°17’51” N - 75°05’48” W), Engystomops pustulosus (Cope, 1864) in Mariquita (04°26’20” N - 75°13’56” W), and Hypsiboas crepitans (Wied-Neuwied, 1824) and Rhinella marina (Linnaeus, 1758) in Potrerillo (04°15’00” N - 74°59’00” W). These species were selected as they occur in areas where the mixture of Roundup® Active and Cosmo-Flux®411F is sprayed, and lay a large number of eggs (Guayara-Barragán & Bernal, 2012). The egg masses were transported to the Laboratory of Herpetology at the University of Tolima, Ibague, Colombia, where they were raised to the experimental Gosner stage ten (embryos) and 25 (tadpoles), in tanks containing tap water (that was dechlorinated by continuous aeration prior to use and at a temperature of 23-25ºC). Organisms were not fed before or during the tests.
Test substances: The commercial formulation of glyphosate, Roundup® Active, which is commonly sprayed by Colombian farmers, was used. It contains 446g/L of potassium salt of N-(phosphonomethyl) glycine, equivalent to 363g/L of glyphosate acid at 20°C. Unfortunately, there is no information about the inactive ingredients in the technical fact sheet of the commercial product, so the presence and proportion of POEA are unknown. The adjuvant Cosmo-Flux®411F contains a mixture of linear and aryl polyethoxylates (17% v/v) and isoparaffins (83% v/v) (Cosmoagro, 2013) and is sold in a concentration of 170g/L at 20°C. We purchased these products from agricultural retailers. Cosmo-Flux®411F was added to the stock test solution at 2.3% v/v to prepare a mixture as used in the field (Solomon et al., 2007; Bernal et al., 2009a). All solutions were prepared with the same dechlorinated tap water used to raise the embryos and tadpoles.
Test procedure-laboratory experiments: A total of 25 embryos and 10 tadpoles per species, with its replicates, were placed separately in each test chambers for a total of 50 embryos and 20 tadpoles per concentration. Biomass loading (defined as the total wet weight of tadpoles per liter of test water) was maintained below 0.6g/L as recommended in ASTM guidelines (1998). Test chambers were 2L glass bowls, containing 1L of test solution, which were indiscriminately positioned in an air-conditioned area of the laboratory, designed to maintain the environmental temperature (24±2ºC) throughout the experiments. After preliminary results of toxicity, the final nominal concentrations were: 0 (negative control: dechlorinated tap water), 325, 750, 1 500, 3 000 and 6 000μg glyphosate a.e./L. Test solutions were renewed daily by transferring the organisms to freshly prepared solutions during the 96h test experiments.
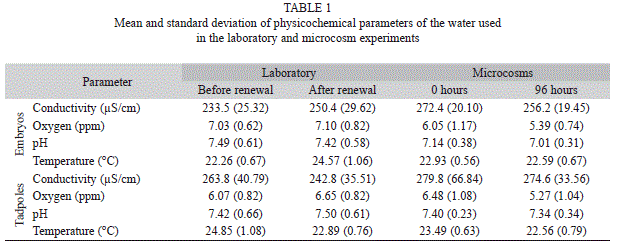
Fluorescent lights (Phillips TLT 20W/54RS) were used for illumination of test bowls. A photoperiod of 12:12h light/dark cycle was controlled by an automatic timer (General Electric PM621). Temperature, dissolved oxygen, conductivity and pH were measured in each test bowl at the beginning, at approximately 24h intervals, and the end of the experiments, including before and after renewals (Table 1). Temperature was measured in test bowls using a liquid-in-glass thermometer. Dissolved oxygen was measured using a portable dissolved oxygen meter (Hanna HI 9146); conductivity and pH were measured using a conductimeter (Hanna HI 8033) and membrane pH meter (HI 8314).
Test procedure-microcosms: A total of 50 embryos and 25 tadpoles per species were placed separately in each of two microcosms for a total of 100 embryos and 50 tadpoles per concentration. Biomass loading was maintained below 0.6g/L as recommended in ASTM guidelines (1998). Microcosms were constructed from polyethylene plastic containers (70cm in diameter, 13cm depth; area: 0.1520m2), which were placed randomly in an indoor but ventilated area of the laboratory at the University of Tolima, with an environmental temperature of 24±2°C and a photoperiod of approximately 12:12h light/dark cycle. At the bottom of the microcosms, 450g of soil, 645g of sand and 10L of dechlorinated tap water to a depth of 12cm above the sediment were added. Then, a fine screen nylon cloth (0.5mm mesh) was placed in the microcosms and pressed into the sediment with four or five small stones (approximately 250g), to facilitate the subsequent collection of embryos and tadpoles, and two leaves (leaf litter) and one macrophyte (Pistia stratiotes) were incorporated. All these materials were obtained from the botanical garden of the University of Tolima, a nonagricultural area. Embryos (Gosner stage ten) and tadpoles (Gosner stage 25) were placed in the water in the middle of the microcosms, which were immediately sprayed with the mix Roundup® Active and Cosmo-Flux®411F, to provide a range of concentration equivalent to 0 (negative control: dechlorinated tap water), 3.69, 7.38, 14.76 and 29.52kg glyphosate a.e./ha.
The amounts of the test substances were measured with a positive displacement pipette and then mixed with 150mL water. This amount was sprayed over the pools, at a height of approximately 20cm, with a small hand-held pump-up garden sprayer of 3L capacity. Spray drift was minimized by spraying just in the center of the pool and with no wind. The physicochemical parameters of the water in the microcosms (Table 1) were similar to those tested in laboratory, and measured at 24, 48, 72 and 96h after the application of the mix. At the 96h of exposure, the nylon mesh was carefully removed from the microcosms and the surviving animals were counted.
Measurements of toxicity and sublethal effects: Mortality was based on the accumulated number of dead and missing animals to 96h of exposure. Toxicity values were estimated through the mean lethal concentration (LC50) and the associated 95% confidence intervals, using the TSK Trimmed Spearman-Karber method (Version 1.5). Sublethal effects at 96h were assessed by measuring the tadpole body size, the stage of embryonic development, and the tadpole swimming performance, as this is expected to relate the ability to evade predators (Fitzpatrick, Reisen, & McCaslin, 2003; Walker et al., 2005). Twenty surviving animals, and ten animals for the laboratory experiments with tadpoles, were tested from the control and each of the tree experimental concentrations lower than the LC50. Photographs of individual tadpoles were taken to measure the body size characters: total length (TL), head width (HW), corporal length (CL) and tail length (tL), using the software ImageJ (http://rsbweb.nih.gov/ij/). This information was analyzed by a MANOVA and the F test (ANOVA) for each variable. Embryos development was compared according to Gosner stages (1960), via ANOVA. Tadpole swimming performance was individually elicited three times in a water-filled rectangular plastic tray (50x1x2cm deep) at 25°C, by the application of a tactile stimulus on the tail. Swimming performance was recorded as the maximum speed (in cm/s) and distance (cm) for each individual. Tadpoles were then photographed and measured (total length) with the software ImageJ (http://rsbweb.nih.gov/ij/). These data were evaluated by an ANCOVA using the total length as covariate.
Results
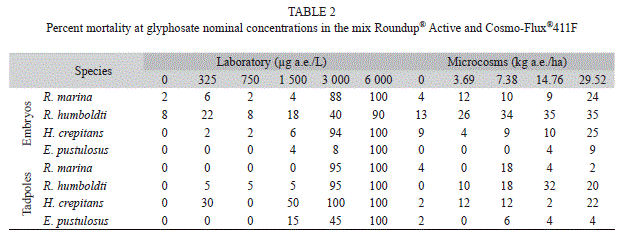
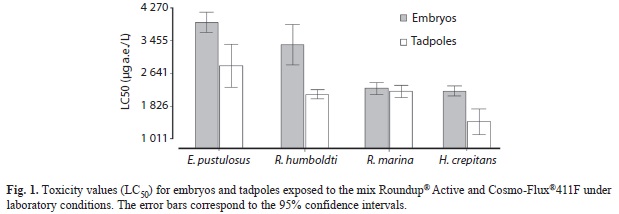
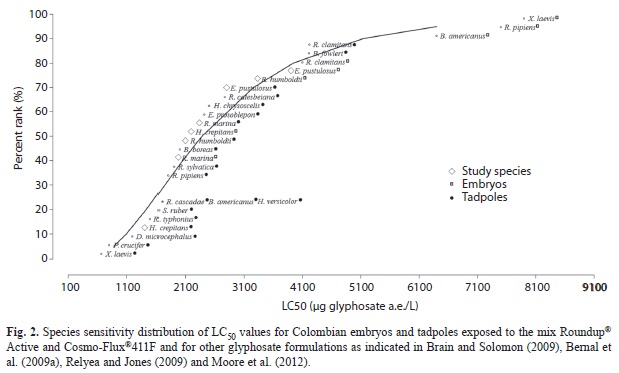
In laboratory experiments, embryos and tadpoles of E. pustulosus were the most tolerant to the mix (embryos: LC50=3 904µg a.e./L; tadpoles: LC50=2 799µg a.e./L), while embryos and tadpoles of H. crepitans (embryos: LC50=2 203µg a.e./L; tadpoles: LC50=1 424µg a.e./L) were the most sensitive (Table 2). R. marina (embryos: LC50=2 270µg a.e./L; tadpoles: LC50=2 170µg a.e./L), and R. humboldti (embryos: LC50=3 336µg a.e./L; tadpoles: LC50=2 121µg a.e./L) presented an intermediate toxicity. Embryos were significantly more tolerant to the mix than tadpoles in three of the four species, but in R. marina there was not a statistical significance at the 95% confidence interval (Fig. 1). The LC50 values for embryos and tadpoles were combined with other data in a sensitivity distribution graph species (SSD) (Fig. 2). The fifth centile of the toxicity distribution was 883μg a.e./L, indicating that 95% of Colombian species are within the toxicity range reported to glyphosate formulations.
Morphometric measurements of larvae (obtained from embryos after 96h exposure) and tadpoles were statistically significant among laboratory treatments (embryos: Hotelling, T2=0.532, p=0.0001; tadpoles: Hotelling, T2=0.46, p<0.0001). For each species, there were significant differences between concentrations (0, 325, 750, 1 500µg a.e./L) for all four morphometric measures (TL, HW, CL, tL) (ANOVA, p<0.05) (Fig. 3), although a decreasing in tadpole body size at the highest sublethal concentration (1 500µg a.e./L) was more evident (Fig. 3). A significant delay in embryonic development was only found between the greatest experimental concentration (organisms in stage 24) and control (organisms in stage 25) (ANOVA, F=4.75, p<0.0043) for R. humboldti. Other species and concentrations did not show any significant change in embryonic development.
In microcosms, mortality was less than 50% for embryos and tadpoles of all the studied species (Table 2), and consequently LC50 values could not be calculated by the TSK method. Additionally, no statistic differences were found among treatments for larval body size, raised from embryos experiments (Hotelling, T2=0.49, p=0.9514), tadpole body size (Hotelling, T2=0.03, p=0.8705), embryonic development (ANOVA, p>0.05), nor tadpole swimming performance (Laboratory: Hotelling, T2=1.4, p=0.1083; Microcosms: Hotelling, T2=2.5, p=0.0827).
Discussion
Amphibians are decreasing at an alarming rate around the world. Several factors have been suggested as possible cause of this declining, among them the herbicides. There is an extensive literature on the toxic effects of the glyphosate on amphibians (Plötner & Matschke, 2012); however, results are contradictory and consequently controversial points of view may be found about its use. Therefore, more information on the impact of ecologically relevant concentrations on survival and sublethal endpoints is essential to determine properly the sensitivity of amphibians (Egea, Relyea, Tejedo, & Torralva, 2012).
In this study, embryos and tadpoles of E. pustulosus were the most resistant to the application of the mix Roundup® Active and Cosmo-Flux®411F, while embryos and tadpoles of H. crepitans showed the greatest sensitivity. These results are in concordance with Bernal et al. (2009a), who found that larvae (stage 25) of E. pustulosus were the most tolerant to the mix GLY-41 and Cosmo-Flux, whereas H. crepitans were the most sensitive. LC50 values could not be calculated for microcosms, this due to the low mortality to exposure concentrations which were up to fourfold the highest field application rate of 3.69kg glyphosate a.e./ha. Toxicity of frog embryos and tadpoles was less in the microcosms than in laboratory tests. The reason for this may be attributed to the presence of sediments and organic matter which rapidly adsorb the glyphosate (Relyea, 2004; Tsui & Chu, 2004; Wojtaszek et al., 2004) and surfactants such as POEA (Wang et al., 2005). In addition, residues of glyphosate decline rapidly within 24h after application (Trumbo, 2005).
Anuran embryos were less sensitive than tadpoles to the mix Roundup® Active and Cosmo-Flux®411F under the same standard laboratory conditions, with the exception of R. marina. This is consistent with Edginton et al. (2004), who found that embryos (Gosner stage eight) of Xenopus laevis., Bufo americanus, Rana clamitans and Rana pipiens exposed to glyphosate formulated Vision® were less sensitive than larvae (Gosner stage 25). The greater sensitivity to pesticides of larval stage as compared with the embryonic stage of amphibians has been also reported by Hall and Swineford (1980), Berrill et al. (1993), Berrill, Bertram, McGillivary, Kolohon and Pauli (1994), Berrill, Coulson, McGillivary and Pauli (1998), Edginton et al. (2004). Surfactants have been demonstrated to cause lysis of gill epithelial cells in the rainbow trout (Partearroyo, Pilling, & Jones, 1991), and affect the ability of the gills to maintain osmotic balance.
In the laboratory experiments, tadpole body size was reduced by the mix at concentrations lower than LC50. Impacts of pesticides on tadpole growth have been reported to be much more prevalent than tadpole survival in some amphibian species (Relyea, 2004) and it may affect populations in the long-term. For instance, slower growth can result in the death of populations if tadpoles are not able to metamorphose before their habitat dries (Wilbur & Collins, 1973). In addition, a smaller body size at metamorphosis has additional long-term fitness effects, such as reduced survival, smaller size at maturity, and lower egg production by females (Semlitsch, Scott, & Pechmann, 1988). Overall, a delay of embryonic development after exposure to the mixture of Roundup® Active and Cosmo-Flux®411F was not observed. Chronic exposures to glyphosate formulations have increased the development time of Bufo americanus, Pseudacris triseriata (Williams & Semlitsch, 2010), and Rana pipiens (Howe et al., 2004), but also accelerated the metamorphosis for Rana cascadae (Cauble & Wagner, 2005). Differences in the biology of the species, the sublethal concentrations and the glyphosate formulations could explain these contradictory results. Tadpole swimming performance was not significant among any of the concentrations tested (lower than LC50), either in laboratory or microcosm experiments. This result was unexpected as other studies have shown that exposure to pesticides caused a significant decrease in the swimming speed for tadpoles of Rana blairi (Bridges, 1997) and Rana berlandieri (Punzo, 2005). It is possible that this commercial formulation Roundup® Active is of low toxicity and does not affect the tadpole swimming performance.
The LC50 values for tadpoles from Colombian frog species showed that sensitivity to glyphosate formulations is similar to that observed in other tropical and temperate species (Maltby, Blake, Brock, & Van den Brink, 2005; Brain & Solomon, 2009; Relyea & Jones, 2009; Moore et al., 2012). This suggests that Roundup formulations have a relatively narrow toxicity range. In addition, that Cosmo-Flux®411F does not significantly increase the toxicity of the glyphosate formulations. At the level of glyphosate considered to be protective of human health (700μg/L a.i., 572μg/L a.e.) (Williams & Semlitsch, 2010), or the amphibian chronic toxicity reference value (TRV) of 740μg/L a.e. (Giesy et al., 2000), which represents the level of chronic exposure expected to result in no deleterious effects to amphibians, the toxicity of the mix Roundup® Active and Cosmo-Flux®411F to Colombian anurans would be low, as the 95% of Colombian embryos and tadpoles would have LC50 greater than these values. On the other hand, neither toxic nor sublethal effects were observed in the microcosms at concentrations up to fourfolds (14.76kg glyphosate a.e./ha) the highest field application rate of 3.69kg glyphosate a.e./ha. We concluded, therefore, that the mix Roundup® Active and Cosmo-Flux®411F, used as recommended in the field, will have negligible effects to anuran embryos and tadpoles of the study species.
Acknowledgment
Funding was provided by Fondo de Investigaciones de la Universidad del Tolima (Project number 490110) and a grant provided by COLCIENCIAS to Marcela Henao as Young Researcher. Collection permit was approved by Corporación Autónoma Regional del Tolima, CORTOLIMA (resolution number 2886 from July 21, 2011). Experiments were authorized by the Bioethics Committee of the Tolima University (October 18, 2011). The authors wish to thank Teófila María Triana and Jorge Luis Turriago for their valuable help in this research, and two anonymous reviewers for their constructive comments to improve the manuscript.
References
ASTM. (1998). Standard guide for conducting the frog embryo teratogenesis Assay-Xenopus (Vol. E 1439-91, pp. 825-836). West Conshohocken, PA: ASTM International. [ Links ]
Bernal, M., Solomon, K., & Carrasquilla, G. (2009a). Toxicity of formulated glyphosate (Glyphos®) and Cosmo-Flux to larval Colombian frogs 1. Laboratory acute toxicity. Journal of Toxicology and Environmental Health, Part A 72, 961-965. [ Links ]
Bernal, M., Solomon, K., & Carrasquilla, G. (2009b). Toxicity of formulated glyphosate (Glyphos®) and Cosmo-Flux to larval and juvenile Colombian frogs 2. Field and laboratory microcosm acute toxicity. Journal of Toxicology and Environmental Health, Part A, 72, 966-973. [ Links ]
Berrill, M., Bertram, S., Wilson, A., Louis, S., Brigham, D., & Stromberg, C. (1993). Lethal and sublethal impacts of pyrethroid insecticides on amphibian embryos and tadpoles. Environmental Toxicology and Chemistry, 12, 522-539. [ Links ]
Berrill, M., Bertram, S., McGillivary, L., Kolohon, M., & Pauli, B. (1994). Effects of low concentrations of forest-use pesticides on frog embryos and tadpoles. Environmental Toxicology and Chemistry, 13, 657-664. [ Links ]
Berrill, M., Coulson, D., McGillivary, L., & Pauli, B. (1998). Toxicity of endosulfan to aquatic stages of anuran amphibians. Environmental Toxicology and Chemistry, 17, 1738-1744. [ Links ]
Bradberry, S. M., Proudfoot, A. T., & Vale, J. A. (2004). Glyphosate poisoning. Toxicological Reviews, 23, 159-167. [ Links ]
Brain, R. A., & Solomon, K. R. (2009). Comparison of the hazards posed to amphibians by the glyphosate spray control program versus the chemical and physical activities of coca production in Colombia. Journal of Toxicology and Environmental Health, Part A: Current Issues, 72, 937-948. [ Links ]
Bridges, C. M. (1997). Tadpole swimming performance and activity affected by acute exposure to sublethal levels of carbaryl. Environmental Toxicology and Chemistry, 16, 1935-1939. [ Links ]
Cauble, K., & Wagner, R. S. (2005). Sublethal effects of the herbicide glyphosate on amphibian metamorphosis and development. Bulletin of Environmental Contamination and Toxicology, 75, 429-435. [ Links ]
Clements, C., Ralph, S., & Petras, M. (1997). Genotoxicity of select herbicides in Rana catesbeiana tadpoles using the alkaline single-cell gel DNA electrophoresis (Comet) Assay. Environmental and Molecular Mutagenesis, 29, 277-288. [ Links ]
Collins, J. P., & Storfer, A. (2003). Global amphibian declines: Sorting the hypotheses. Diversity and Distributions, 9, 89-98. [ Links ]
Cosmoagro. (2013). Colombia. Retrieved from http://www.cosmoagro.com. [ Links ]
Cowman, D. F., & Mazanti, L. E. (2000). Ecotoxicology of ‘‘new generation’’ pesticides to amphibians. In Ecotoxicology of Amphibians and Reptiles (pp. 233-268). Pensacola, FL, USA: SETAC. [ Links ]
Duke, S. O., & Powles, S. B. (2008). Glyphosate: A once-in-a-century herbicide. Pest Management Science, 64, 319-325. [ Links ]
Edginton, A., Sheridan, P., Stephenson, G., Thompson, D., & Boermans, H. (2004). Comparative effects of pH and Vision herbicide on two life stages of four anuran amphibian species. Environmental Toxicology and Chemistry, 23, 815-822. [ Links ]
Egea, A., Relyea, R. A., Tejedo, M., & Torralva, M. (2012). Understanding of the impact of chemicals on amphibians: a meta-analytic review. Ecology and Evolution, 2(7), 1382-1397. [ Links ]
Fitzpatrick, S., Reisen, P., & McCaslin, M. (2003). Pollen-mediated gene flow in alfalfa: A three year summary of field research. Proceedings of the 2003 Central Alfalfa Improvement Conference. National Alfalfa Improvement Conference. Retrieved from http://naaic.org/Meetings/Central2003/Gene_Flow_in_Alfalfa_Abstract_final.doc. [ Links ]
Giesy, J., Dobson, S., & Solomon, K. (2000). Ecotoxicological risk assessment for Roundup® herbicide. Reviews of Environmental Contamination and Toxicology, 167, 35-120. [ Links ]
Glyphosate, World Health Organization International Program on Chemical Safety. (1994). Geneva, vol. 159. Retrieved from http://www.inchem.org/documents/ehc/ehc/ehc159.htm). [ Links ]
Gosner, K. L. (1960). A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica, 16, 183-190. [ Links ]
Guayara-Barragán, M. G., & Bernal, M. H. (2012). Fecundidad y fertilidad en once especies de anuros colombianos con diferentes modos reproductivos. Caldasia, 34, 483-496. [ Links ]
Hall, R. J., & Swineford, D. (1980). Toxic effects of endrin and toxaphene on the southern leopard frog Rana sphenocephala. Environmental Pollution, 23, 53-65. [ Links ]
Henao, L. M., Arango, V. A., & Bernal, M. H. (2013). Toxicidad aguda y efectos subletales del Cosmo-Flux®411F en embriones de cuatro especies de anuros colombianos. Actualidades Biológicas, 35(99), 209-218. [ Links ]
Howe, C. M., Berrill, M., Pauli, B. D., Helbing, C. C., Werry, K., & Veldhoen, N. (2004). Toxicity of glyphosate-based pesticides to four North American frog species. Environmental Toxicology and Chemistry, 23, 1928-1938. [ Links ]
Lajmanovich, R., Lorenzatti, E., Maitre, M., Enrique, S., & Peltzer, P. (2003). Comparative acute toxicity of the commercial herbicides glyphosate to neotropical tadpoles Scinax nasicus (Anura: Hylidae). Fresenius Environmental Bulletin, 12(4), 364-367. [ Links ]
Lajmanovich, R. C., Attademo, A. M., Peltzer, P. M., Junges, C. M., & Cabagna, M. C. (2011). Toxicity of four herbicide formulations with glyphosate on Rhinella arenarum (Anura: Bufonidae) Tadpoles: B-esterases and glutathione S-transferase inhibitors. Archives of Environmental Contamination and Toxicology, 60, 681-689. [ Links ]
Lenkowski, J. R., Sanchez-Bravo, G., & Mclaughlin, K. A. (2010). Low concentrations of atrazine, glyphosate, 2.4-dichlorophenoxyacetic acid, and triadimefon exposures have diverse effects on Xenopus laevis. organ morphogenesis. Journal of Environmental Sciences, 22, 1305-1308. [ Links ]
Maltby, L., Blake, N. N., Brock, T. C. M., & Van den Brink, P. J. (2005). Insecticide species sensitivity distributions: the importance of test species selection and relevance to aquatic ecosystems. Environmental Toxicology and Chemistry, 24, 379-388. [ Links ]
Mann, R. M., & Bidwell, J. R. (1999). The toxicity of glyphosate and several glyphosate formulations to four species of southwestern Australian frogs. Archives of Environmental Contamination and Toxicology, 36, 193-199. [ Links ]
Mann, R. M., Hyne, R. V., Choung, C. B., & Wilson, S. P. (2009). Amphibians and agricultural chemicals: Review of the risk in a complex environment. Environmental Pollution, 157, 2903-2927. [ Links ]
Mattoon, A. (2000). El declive de los anfibios. Worldwatch Institute, 23, 11-21. [ Links ]
Ministry of Environment, Housing and Territorial Development of Colombia, “Resolution Number 1435” (2007). Colombia. Retrieved from http://www.minambiente.gov.co/documentos/res_1435_150807.pdf). [ Links ]
Moore, L. J., Fuentes, L., Rodgers, J. H., Boweman, W. W., Yarrow, G. K., Chao, W. Y., & Bridge, W. C. (2012). Relative toxicity of the components of the original formulation of Roundup® to five North American anurans. Ecotoxicology and Environmental Safety, 78, 128-133. [ Links ]
Partearroyo, M. A., Pilling, S. J., & Jones, M. N. (1991). The lysis of isolated fish (Oncorhynchus mykiss) gill epithelial cells by surfactants. Comparative Biochemistry and Physiology Part C: Comparative Pharmacology, 100, 381-388. [ Links ]
Plötner, J., & Matschke, J. (2012). Acute toxic, sublethal and indirect effects of glyphosate and glyphosate herbicide on amphibians - an overview. Zeitschrift für Feldherpetologie, 19, 1-20. [ Links ]
Punzo, F. (2005). Effects of insecticide (carbaryl) exposure on activity and swimming performance of tadpoles of the Rio Grande Leopard Frog, Rana berlandieri (Anura: Ranidae). Texas Journal of Science, 57(3), 263-272. [ Links ]
Relyea, R. A. (2004). Growth and survival of five amphibian species exposed to combinations of pesticides. Environmental Toxicology and Chemistry, 23(7), 1737-1742. [ Links ]
Relyea, R. A. (2005). The lethal impact of Roundup on aquatic and terrestrial amphibians. Ecological Applications, 15, 1118-1124. [ Links ]
Relyea, R. A., & Jones, D. K. (2009). The toxicity of Roundup Original Max® to 13 species of larval amphibians. Environmental Toxicology and Chemistry, 28(9), 2004-2008. [ Links ]
Relyea, R. A. (2012). New effects of Roundup on amphibians: Predators reduce herbicide mortality; herbicides induce antipredator morphology. Ecological Applications, 22(2), 634-647. [ Links ]
Rondón, I., Ramírez, W., & Eslava, P. (2007). Evaluación de los efectos tóxicos y concentración letal 50 del surfactante Cosmoflux®411F sobre juveniles de cachama blanca (Piaractus brachypomus). Revista Colombiana de Ciencias Pecuarias, 20(4), 431-446. [ Links ]
Semlitsch, R. D., Scott, D. C., & Pechmann, J. H. K. (1988). Time and size at metamorphosis related to adult fitness in Ambystoma talpoideum. Ecology, 69, 184-192. [ Links ]
Solomon, K. R., & Thompson, D. G. (2003). Ecological risk assessment for aquatic organisms from over-water uses of glyphosate. Journal of Toxicology and Environmental Health, Part B, 6, 289-324. [ Links ]
Solomon, K. R., Anadón, A., Cerdeira, A. L., Marshall, J., & Sanin, L. H. (2005). Estudio de los efectos del Programa de Erradicación de Cultivos Ilícitos mediante la aspersión aérea con el herbicida Glifosato (PECIG) y de los cultivos ilícitos en la salud humana y en el medio ambiente. Comisión Interamericana para el Control del Abuso de Drogas (CICAD). [ Links ]
Solomon, K. R., Anadón, A., Carrasquilla, G., Cerdeira, A., Marshall, J., & Sanin, L. H. (2007). Coca and poppy eradication in Colombia: Environmental and human health assessment of aerially applied glyphosate. Reviews of Environmental Contamination and Toxicology, 190, 43-125. [ Links ]
Sparling, D., Linder, G., Bishop, C., & Krest, S. (2010). Ecotoxicology of amphibians and reptiles. Pensacola, Florida: CRC Press. [ Links ]
Storfer, A. (2003). Amphibian declines: future directions. Diversity and Distributions, 9, 151-163. [ Links ]
Triana, T. M., Montes, C. M., & Bernal, M. H. (2013). Efectos letales y subletales del glifosato (Roundup® Activo) en embriones de anuros colombianos. Acta biológica Colombiana, 18(2), 271-278. [ Links ]
Trumbo, J. (2005). An assessment of the hazard of a mixture of the herbicide Rodeo® and the non-ionic surfactant R-11® to aquatic invertebrates and larval amphibians. California fish and game, 91(1), 38-46. [ Links ]
Tsui, M. T., & Chu, L. M. (2004). Comparative toxicity of glyphosate-based herbicides: Aqueous and sediment pore water exposures. Archives of Environmental Contamination and Toxicology, 46, 316-323. [ Links ]
Walker, S. R., Taylor, I. N., Milne, G., Osten, V. A., Hoque, Z., & Farquharson, R. J. (2005). A survey of management and economic impact of weeds in dryland cotton cropping systems of subtropical Australia. Australian Journal of Experimental Agriculture, 45, 79-91. [ Links ]
Wang, N., Besser, J. M., Buckler, D. R., Honegger, J. L., Ingersoll, C. G., Johnson, B. T., & McKee, M. J. (2005). Influence of sediment on the fate and toxicity of a polyethoxylated tallowamine surfactant system (MON 0818) in aquatic microcosms. Chemosphere, 59, 545-551. [ Links ]
Wilbur, H. M., & Collins, J. P. (1973). Ecological aspects of amphibian metamorphosis. Science, 182, 1305-1314. [ Links ]
Williams, B. K., & Semlitsch, R. D. (2010). Larval responses of three midwestern anurans to chronic low-dose exposures of four herbicides. Archives of Environmental Contamination and Toxicology, 58, 819-827. [ Links ]
Wojtaszek, B., Staznik, B., Chartrand, D., Stephenson, G., & Thompson, D. (2004). Effects of Vision® herbicide on mortality, avoidance response, and growth of amphibian larvae in two forest wetlands. Environmental Toxicology and Chemistry, 23(4), 832-842. [ Links ]
Bernal, M., Solomon, K., & Carrasquilla, G. (2009a). Toxicity of formulated glyphosate (Glyphos®) and Cosmo-Flux to larval Colombian frogs 1. Laboratory acute toxicity. Journal of Toxicology and Environmental Health, Part A 72, 961-965. [ Links ]
Bernal, M., Solomon, K., & Carrasquilla, G. (2009b). Toxicity of formulated glyphosate (Glyphos®) and Cosmo-Flux to larval and juvenile Colombian frogs 2. Field and laboratory microcosm acute toxicity. Journal of Toxicology and Environmental Health, Part A, 72, 966-973. [ Links ]
Berrill, M., Bertram, S., Wilson, A., Louis, S., Brigham, D., & Stromberg, C. (1993). Lethal and sublethal impacts of pyrethroid insecticides on amphibian embryos and tadpoles. Environmental Toxicology and Chemistry, 12, 522-539. [ Links ]
Berrill, M., Bertram, S., McGillivary, L., Kolohon, M., & Pauli, B. (1994). Effects of low concentrations of forest-use pesticides on frog embryos and tadpoles. Environmental Toxicology and Chemistry, 13, 657-664. [ Links ]
Berrill, M., Coulson, D., McGillivary, L., & Pauli, B. (1998). Toxicity of endosulfan to aquatic stages of anuran amphibians. Environmental Toxicology and Chemistry, 17, 1738-1744. [ Links ]
Bradberry, S. M., Proudfoot, A. T., & Vale, J. A. (2004). Glyphosate poisoning. Toxicological Reviews, 23, 159-167. [ Links ]
Brain, R. A., & Solomon, K. R. (2009). Comparison of the hazards posed to amphibians by the glyphosate spray control program versus the chemical and physical activities of coca production in Colombia. Journal of Toxicology and Environmental Health, Part A: Current Issues, 72, 937-948. [ Links ]
Bridges, C. M. (1997). Tadpole swimming performance and activity affected by acute exposure to sublethal levels of carbaryl. Environmental Toxicology and Chemistry, 16, 1935-1939. [ Links ]
Cauble, K., & Wagner, R. S. (2005). Sublethal effects of the herbicide glyphosate on amphibian metamorphosis and development. Bulletin of Environmental Contamination and Toxicology, 75, 429-435. [ Links ]
Clements, C., Ralph, S., & Petras, M. (1997). Genotoxicity of select herbicides in Rana catesbeiana tadpoles using the alkaline single-cell gel DNA electrophoresis (Comet) Assay. Environmental and Molecular Mutagenesis, 29, 277-288. [ Links ]
Collins, J. P., & Storfer, A. (2003). Global amphibian declines: Sorting the hypotheses. Diversity and Distributions, 9, 89-98. [ Links ]
Cosmoagro. (2013). Colombia. Retrieved from http://www.cosmoagro.com. [ Links ]
Cowman, D. F., & Mazanti, L. E. (2000). Ecotoxicology of ‘‘new generation’’ pesticides to amphibians. In Ecotoxicology of Amphibians and Reptiles (pp. 233-268). Pensacola, FL, USA: SETAC. [ Links ]
Duke, S. O., & Powles, S. B. (2008). Glyphosate: A once-in-a-century herbicide. Pest Management Science, 64, 319-325. [ Links ]
Edginton, A., Sheridan, P., Stephenson, G., Thompson, D., & Boermans, H. (2004). Comparative effects of pH and Vision herbicide on two life stages of four anuran amphibian species. Environmental Toxicology and Chemistry, 23, 815-822. [ Links ]
Egea, A., Relyea, R. A., Tejedo, M., & Torralva, M. (2012). Understanding of the impact of chemicals on amphibians: a meta-analytic review. Ecology and Evolution, 2(7), 1382-1397. [ Links ]
Fitzpatrick, S., Reisen, P., & McCaslin, M. (2003). Pollen-mediated gene flow in alfalfa: A three year summary of field research. Proceedings of the 2003 Central Alfalfa Improvement Conference. National Alfalfa Improvement Conference. Retrieved from http://naaic.org/Meetings/Central2003/Gene_Flow_in_Alfalfa_Abstract_final.doc. [ Links ]
Giesy, J., Dobson, S., & Solomon, K. (2000). Ecotoxicological risk assessment for Roundup® herbicide. Reviews of Environmental Contamination and Toxicology, 167, 35-120. [ Links ]
Glyphosate, World Health Organization International Program on Chemical Safety. (1994). Geneva, vol. 159. Retrieved from http://www.inchem.org/documents/ehc/ehc/ehc159.htm). [ Links ]
Gosner, K. L. (1960). A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica, 16, 183-190. [ Links ]
Guayara-Barragán, M. G., & Bernal, M. H. (2012). Fecundidad y fertilidad en once especies de anuros colombianos con diferentes modos reproductivos. Caldasia, 34, 483-496. [ Links ]
Hall, R. J., & Swineford, D. (1980). Toxic effects of endrin and toxaphene on the southern leopard frog Rana sphenocephala. Environmental Pollution, 23, 53-65. [ Links ]
Henao, L. M., Arango, V. A., & Bernal, M. H. (2013). Toxicidad aguda y efectos subletales del Cosmo-Flux®411F en embriones de cuatro especies de anuros colombianos. Actualidades Biológicas, 35(99), 209-218. [ Links ]
Howe, C. M., Berrill, M., Pauli, B. D., Helbing, C. C., Werry, K., & Veldhoen, N. (2004). Toxicity of glyphosate-based pesticides to four North American frog species. Environmental Toxicology and Chemistry, 23, 1928-1938. [ Links ]
Lajmanovich, R., Lorenzatti, E., Maitre, M., Enrique, S., & Peltzer, P. (2003). Comparative acute toxicity of the commercial herbicides glyphosate to neotropical tadpoles Scinax nasicus (Anura: Hylidae). Fresenius Environmental Bulletin, 12(4), 364-367. [ Links ]
Lajmanovich, R. C., Attademo, A. M., Peltzer, P. M., Junges, C. M., & Cabagna, M. C. (2011). Toxicity of four herbicide formulations with glyphosate on Rhinella arenarum (Anura: Bufonidae) Tadpoles: B-esterases and glutathione S-transferase inhibitors. Archives of Environmental Contamination and Toxicology, 60, 681-689. [ Links ]
Lenkowski, J. R., Sanchez-Bravo, G., & Mclaughlin, K. A. (2010). Low concentrations of atrazine, glyphosate, 2.4-dichlorophenoxyacetic acid, and triadimefon exposures have diverse effects on Xenopus laevis. organ morphogenesis. Journal of Environmental Sciences, 22, 1305-1308. [ Links ]
Maltby, L., Blake, N. N., Brock, T. C. M., & Van den Brink, P. J. (2005). Insecticide species sensitivity distributions: the importance of test species selection and relevance to aquatic ecosystems. Environmental Toxicology and Chemistry, 24, 379-388. [ Links ]
Mann, R. M., & Bidwell, J. R. (1999). The toxicity of glyphosate and several glyphosate formulations to four species of southwestern Australian frogs. Archives of Environmental Contamination and Toxicology, 36, 193-199. [ Links ]
Mann, R. M., Hyne, R. V., Choung, C. B., & Wilson, S. P. (2009). Amphibians and agricultural chemicals: Review of the risk in a complex environment. Environmental Pollution, 157, 2903-2927. [ Links ]
Mattoon, A. (2000). El declive de los anfibios. Worldwatch Institute, 23, 11-21. [ Links ]
Ministry of Environment, Housing and Territorial Development of Colombia, “Resolution Number 1435” (2007). Colombia. Retrieved from http://www.minambiente.gov.co/documentos/res_1435_150807.pdf). [ Links ]
Moore, L. J., Fuentes, L., Rodgers, J. H., Boweman, W. W., Yarrow, G. K., Chao, W. Y., & Bridge, W. C. (2012). Relative toxicity of the components of the original formulation of Roundup® to five North American anurans. Ecotoxicology and Environmental Safety, 78, 128-133. [ Links ]
Partearroyo, M. A., Pilling, S. J., & Jones, M. N. (1991). The lysis of isolated fish (Oncorhynchus mykiss) gill epithelial cells by surfactants. Comparative Biochemistry and Physiology Part C: Comparative Pharmacology, 100, 381-388. [ Links ]
Plötner, J., & Matschke, J. (2012). Acute toxic, sublethal and indirect effects of glyphosate and glyphosate herbicide on amphibians - an overview. Zeitschrift für Feldherpetologie, 19, 1-20. [ Links ]
Punzo, F. (2005). Effects of insecticide (carbaryl) exposure on activity and swimming performance of tadpoles of the Rio Grande Leopard Frog, Rana berlandieri (Anura: Ranidae). Texas Journal of Science, 57(3), 263-272. [ Links ]
Relyea, R. A. (2004). Growth and survival of five amphibian species exposed to combinations of pesticides. Environmental Toxicology and Chemistry, 23(7), 1737-1742. [ Links ]
Relyea, R. A. (2005). The lethal impact of Roundup on aquatic and terrestrial amphibians. Ecological Applications, 15, 1118-1124. [ Links ]
Relyea, R. A., & Jones, D. K. (2009). The toxicity of Roundup Original Max® to 13 species of larval amphibians. Environmental Toxicology and Chemistry, 28(9), 2004-2008. [ Links ]
Relyea, R. A. (2012). New effects of Roundup on amphibians: Predators reduce herbicide mortality; herbicides induce antipredator morphology. Ecological Applications, 22(2), 634-647. [ Links ]
Rondón, I., Ramírez, W., & Eslava, P. (2007). Evaluación de los efectos tóxicos y concentración letal 50 del surfactante Cosmoflux®411F sobre juveniles de cachama blanca (Piaractus brachypomus). Revista Colombiana de Ciencias Pecuarias, 20(4), 431-446. [ Links ]
Semlitsch, R. D., Scott, D. C., & Pechmann, J. H. K. (1988). Time and size at metamorphosis related to adult fitness in Ambystoma talpoideum. Ecology, 69, 184-192. [ Links ]
Solomon, K. R., & Thompson, D. G. (2003). Ecological risk assessment for aquatic organisms from over-water uses of glyphosate. Journal of Toxicology and Environmental Health, Part B, 6, 289-324. [ Links ]
Solomon, K. R., Anadón, A., Cerdeira, A. L., Marshall, J., & Sanin, L. H. (2005). Estudio de los efectos del Programa de Erradicación de Cultivos Ilícitos mediante la aspersión aérea con el herbicida Glifosato (PECIG) y de los cultivos ilícitos en la salud humana y en el medio ambiente. Comisión Interamericana para el Control del Abuso de Drogas (CICAD). [ Links ]
Solomon, K. R., Anadón, A., Carrasquilla, G., Cerdeira, A., Marshall, J., & Sanin, L. H. (2007). Coca and poppy eradication in Colombia: Environmental and human health assessment of aerially applied glyphosate. Reviews of Environmental Contamination and Toxicology, 190, 43-125. [ Links ]
Sparling, D., Linder, G., Bishop, C., & Krest, S. (2010). Ecotoxicology of amphibians and reptiles. Pensacola, Florida: CRC Press. [ Links ]
Storfer, A. (2003). Amphibian declines: future directions. Diversity and Distributions, 9, 151-163. [ Links ]
Triana, T. M., Montes, C. M., & Bernal, M. H. (2013). Efectos letales y subletales del glifosato (Roundup® Activo) en embriones de anuros colombianos. Acta biológica Colombiana, 18(2), 271-278. [ Links ]
Trumbo, J. (2005). An assessment of the hazard of a mixture of the herbicide Rodeo® and the non-ionic surfactant R-11® to aquatic invertebrates and larval amphibians. California fish and game, 91(1), 38-46. [ Links ]
Tsui, M. T., & Chu, L. M. (2004). Comparative toxicity of glyphosate-based herbicides: Aqueous and sediment pore water exposures. Archives of Environmental Contamination and Toxicology, 46, 316-323. [ Links ]
Walker, S. R., Taylor, I. N., Milne, G., Osten, V. A., Hoque, Z., & Farquharson, R. J. (2005). A survey of management and economic impact of weeds in dryland cotton cropping systems of subtropical Australia. Australian Journal of Experimental Agriculture, 45, 79-91. [ Links ]
Wang, N., Besser, J. M., Buckler, D. R., Honegger, J. L., Ingersoll, C. G., Johnson, B. T., & McKee, M. J. (2005). Influence of sediment on the fate and toxicity of a polyethoxylated tallowamine surfactant system (MON 0818) in aquatic microcosms. Chemosphere, 59, 545-551. [ Links ]
Wilbur, H. M., & Collins, J. P. (1973). Ecological aspects of amphibian metamorphosis. Science, 182, 1305-1314. [ Links ]
Williams, B. K., & Semlitsch, R. D. (2010). Larval responses of three midwestern anurans to chronic low-dose exposures of four herbicides. Archives of Environmental Contamination and Toxicology, 58, 819-827. [ Links ]
Wojtaszek, B., Staznik, B., Chartrand, D., Stephenson, G., & Thompson, D. (2004). Effects of Vision® herbicide on mortality, avoidance response, and growth of amphibian larvae in two forest wetlands. Environmental Toxicology and Chemistry, 23(4), 832-842. [ Links ]
1. Grupo de Herpetología, Eco-Fisiología & Etología, Departamento de Biología, Universidad del Tolima, Calle 42 Barrio Santa Helena Ibagué, Colombia; lmhenaom@ut.edu.co, cmmontesr@ut.edu.co, mhbernal@ut.edu.co
Received 20-XII-2013. Corrected 25-VII-2014. Accepted 26-VIII-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}