










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.1 San José Jan./Mar. 2015
Toxicity induced by dieldrin and chlorpyrifos in the freshwater crayfish Cambarellus montezumae (Cambaridae)
Toxicidad inducida por dieldrín y clorpirifós en Cambarellus montezumae (Cambaridae)
Toxicidad inducida por dieldrín y clorpirifós en Cambarellus montezumae (Cambaridae)
Sandra Díaz-Barriga Arceo1*, Laura Martínez-Tabche2*, Isela Álvarez-González3*, Eugenia López López4* & Eduardo Madrigal-Bujaidar5*
Abstract
The toxicity induced by insecticides in aquatic organisms is of utmost relevance because it may give a clue about the degree of health or damage of the involved ecosystem. In the present report, we determined the effect of dieldrin (DD) and chlorpyrifos (CP) on the freshwater crayfish, Cambarellus montezumae. The organisms (4-6cm in diameter) were collected in the Ignacio Ramírez Reservoir, situated at 50km Northeast of Mexico City, in the Rio Lerma Basin. Initially, we determined the LC50 value with the Probit method, then the DNA damage with single cell gel electrophoresis (comet assay applied at 24, 48, and 72h of exposure) to the brain and hepatopancreas of animals exposed (in reconstituted water) to 0.05 and 0.5µg/L of each insecticide. In the hepatopancreas of the same organisms, we determined the lipid peroxidation by applying the TBARS test. DNA damage and lipid peroxidation were also evaluated with the same methods to organisms exposed in water from the reservoir. In regard to the LC50 at 72h of exposure, we found a value of 5.1µg/L and a value of 5.62µg/L for DD and CP, respectively. The comet assay applied at different exposure times showed significant DNA damage to both organs, with respect to the control level. In the case of DD, statistical significance was observed for the two doses in the whole evaluated schedule. CP was genotoxic in the brain with the high dose at 72h, and in the hepatopancreas with the two tested doses at all evaluated exposure times. Also, a significant lipid peroxidation increase was detected with the two doses of insecticides. In the study with water from the reservoir, a more pronounced DNA damage was detected. Our results showed strong DNA damage induced by both insecticides in the crayfish, as well as a correlation with the lipid peroxidation effect, suggesting that oxidative stress is involved in the genotoxic alteration. Our results also showed the usefulness of the studied organism as well as the applied tests for the evaluation of toxicological effects, and suggested the pertinence of applying the comet assay to other freshwater organisms to evaluate the bioaccumulation of insecticides. Rev. Biol. Trop. 63 (1): 83-96. Epub 2015 March 01.
Key words: genotoxicity, dieldrin, chlorpyrifos, lipoperoxidation, crayfish.
Resumen
La toxicidad inducida por insecticidas en organismos acuáticos es de gran relevancia porque puede orientar sobre el grado de salud o daño del ecosistema involucrado. En el presente estudio determinamos el efecto del dieldrín (DD) y del clorpirifós (CP) en el acocil de agua dulce Cambarellus montezumae. Los organismos (4-6cm de diámetro) se recolectaron en la Represa Ignacio Ramírez, situada a 50km al Noreste de la Ciudad de México, en la cuenca del Río Lerma. Inicialmente determinamos la LC50 con el método de Probit y después el daño al ADN mediante la electroforesis unicelular en gel (ensayo cometa, aplicado a las 24, 48 y 72 h de exposición) en el cerebro y el hepatopáncreas de animales expuestos (en agua reconstituida) a 0.05 y 0.5µg/L de cada insecticida. En el hepatopáncreas de los mismos organismos determinamos la peroxidación lipídica aplicando la prueba de TBARS. El daño al ADN y la peroxidación lipídica también se evaluaron con los mismos métodos en organismos expuestos a los insecticidas en agua de la represa. En relación a la LC50, a las 72h de exposición encontramos un valor de 5.1µg/L y un valor de 5.62µg/L para DD y CP, respectivamente. El ensayo cometa aplicado a diferentes tiempos de exposición mostró un significativo daño al ADN en ambos órganos con respecto al valor del testigo. En el caso del DD se observó significancia estadística para las dos dosis en todo el horario evaluado. CP fue genotóxico en el cerebro con la dosis más alta a las 72 h y en hepatopáncreas con las dos dosis, en todos los tiempos de exposición evaluados. También se detectó un significativo aumento de la peroxidación lipídica con las dos dosis de los insecticidas. En el estudio con el agua de la represa se detectó un daño más pronunciado en el ADN. Nuestros resultados mostraron un fuerte daño al ADN en Cambarellus montezumae por ambos insecticidas, así como una correlación con el efecto de la peroxidación lipídica, lo que sugiere que el estrés oxidativo está involucrado en la alteración genotóxica. Nuestros resultados también mostraron la utilidad del organismo estudiado y de las pruebas aplicadas para evaluar efectos tóxicos, y sugieren la pertinencia de aplicar el ensayo cometa en otros organismos de agua dulce para evaluar la bioacumulación de insecticidas.
Palabras clave: genotoxicidad, dieldrín, clorpirifós, lipoperoxidación, acocil.
Pesticides are widely used in agricultural production to prevent or reduce losses caused by pests, and thus improve yields and quality of the products. These chemicals can be considered as an economic, labor-saving, and efficient tool of pest management (Cooper, & Dobson, 2007). However, their chemical structure, incorrect preparation, application and storage may pose a serious toxicity risk for a number of organisms. In the case of aquatic ecosystems, though agricultural soil is usually the primary recipient of pesticides, adjacent water bodies may become the ultimate recipient for pesticide residues by leaching, runoff, or spray drift (Pereira et al., 2009).
Dieldrin (DD) is a cyclodiene insecticide originally formulated from a waste product of synthetic rubber. It is characterized by a high environmental persistency related with a bioaccumulation because of its resistance to physical degradation and biological metabolism. In invertebrates, the chemical is easily absorbed and transported throughout the body in the blood or in the hemolymph (Jorgenson, 2001). In mammals, its toxicity is mainly manifested in the liver, where it may induce tumors in various experimental animals, although this effect has not been validated in humans (Swaen, de Jong, Slangen, & van Amelsvoort, 2002). The compound may also act as an endocrine disruptor, and may cause reproductive difficulties in terrestrial and aquatic organisms, such as testicular apoptosis and decreased sperm motility (Murray, Lea, Abramovich, Haites, & Fowler, 2001; Jorgenson, 2001; Perobelli et al., 2010).
In the case of its genotoxicity, the chemical was initially considered with weak potency; however, a number of assays have subsequently given positive effects in animals and plants (Cicchetti, Bari, & Argentin, 1999; Pandey, 2008). In the light of its strong toxicity and bio-accumulation in the ecosystems, the use of DD for crop and termite control has been banned in most countries; however, its manufacturing as wood preservative, and as intermediate for other chemical processes has continued; therefore, such compound may be routinely found in the environment (Matsumoto, Kawanaka, Yun, & Oyaizu, 2009).
Chlorpyrifos (CP) is a highly used organophosphorous chemical which has served as insecticide and acaricide. Initially, CP was used in both agricultural and non-agricultural environments to combat a number of insect pests, and was also used as the active component in a wide array of pesticide formulations (Eaton et al., 2008). However, noncrop usage has tended to be restricted due to toxicological and epidemiological information that have suggested a role for CP in neurodevelopmental damage, including its participation in autism, when exposure to the compound is done in early pregnancy (Eaton et al., 2008; Landrigan, 2010).
In the case of its genotoxic potential, a number of assays have shown negative results (Gollapudi, Mendrala, & Linscombe, 1995); however, as previously indicated with respect to DD, other studies have found the opposite in different organisms, including aquatic animals such as Drosophila, mice, Chinese toad, and the fish, Channa punctatus (Rahman, Mahboob, Danadevi, Saleha, & Groover, 2002; Ali et al., 2009; Yin, Zhu, Li, & Liu, 2009; Gupta et al., 2010). Besides, the compound has been detected in a number of aquatic environments, suggesting that it may be involved in ecotoxicological damages (Palma, Palma, Fernandes, Soares, & Barbosa, 2008).
Both insecticides have several common characteristics relevant to the aims of the present study: they are oxidative stress inducers, neurotoxicants in mammals and aquatic organisms, and are environmentally important contaminants because of their level of persistency (Martyniuk et al., 2010; Slotkin, & Seidler, 2010). In genotoxicology, it is highly recommended to apply various tests in order to determine the potentials of a particular chemical. Moreover, these tests are also recommended when contradictory results are reported, as it stands in the present case, because it is well known that each test provides specific information on the type of DNA damage; a knowledge that is relevant to eventually implement antigenotoxic measures. In this context, it is an important fact that no studies with the single cell gel electrophoresis (comet assay) have been made on the presently tested insecticides. The alkaline version of this test is a reliable and relatively rapid assay that mainly expresses single strand breaks and alkaline labile sites induced by the tested agent (Tice et al., 2000). In the present case, its application may contribute to define the genotoxic discrepancy reported on the insecticides, and if considered appropriate because of the present and new studies, it may be included in the field of ecotoxicology, as part of a battery for insecticides toxicological evaluation (Cotelle, & Ferard, 1999).
It is well known that the presently studied compounds may affect a number of molecular and cellular targets, such as mitochondria, or molecules related with apoptosis; however, for the purpose of the present report, it is more pertinent to emphasize their capacity to produce oxidative stress. This has been demonstrated in various models by determining lipid peroxidation, protein carbonils, total glutathione, catalase, superoxide dismutase, glutation peroxidase, and other biomarkers (Verma, & Srivastava, 2003; Hatcher et al., 2007; Narra, 2014). Moreover, it has been reported that DD may potentiate oxidative stress when combined with other agents (Chimeddorj et al., 2013). In this context, malondialdehyde (MDA), in particular, has received attention because it has been considered a toxic messenger that can propagate and amplify oxidative injury (Pizzimenti et al., 2013). Besides, it is known that it can react with DNA forming adducts (deoxyguanosine, deoxyadenosine, and deoxycytidine), which can cause mutagenic and carcinogenic effects and deregulation of cell homeostasis, thus contributing significantly to disease phenotype (Marnett, 2002; Voulgaridou, Anestopoulos, Franco, Panayiotidis, & Pappa, 2011). Moreover, MDA increase has been correlated with the increase in DNA damage (Voulgaridou et al., 2011). The above mentioned information supports our aims for the present report: to determine, initially, the DNA damage induced by the pesticides in crayfishes exposed to reconstituted water, as well as water from the reservoir in which they were obtained. This last assay may help to explore genotoxic differences between exposure to clean and polluted water, with different physicochemical properties; the second aim was to determine the lipoperoxidative capacity of the pesticides (in both types of water), in order to explore a correlation between the DNA damage and oxidative stress. The last purpose of our study was to examine whether the results obtained in Cambarellus montezumae (Cambaridae, Decapoda, Crustacea) may suggest the extension of research on the matter, so as to evaluate the potential of the organism (and the test) to monitor the toxicity elicited by insecticides.
Material and Methods
Chemicals and crayfishes: The following chemicals were obtained from Sigma-Aldrich Chemicals (St Louis, Mo, USA.): DD; CP; phosphate buffer saline (PBS); potassium chloride; tris; sodium carbonate; disodium ethylenediamine-tetra-acetate (EDTA); dimethyl sulfoxide (DMSO); reduced glutathione; hexane/isopropanol; thiobarbituric acid (TBA); trichloroacetic acid (TCA); Coomassie brilliant blue G; ethidium bromide (EB); bovine albumin; and butilated hydroxytoluene (BHT). Low melting point agarose (LMPA) and normal melting point agarose (NMPA) were purchased from GIBCO-BRL (Grand Island, NY, USA.). Nitric acid and dibasic potassium phosphate were obtained from J. T. Baker (Phillisburg, NJ, USA.).
The organisms were collected from the Ignacio Ramírez Reservoir, which is located 50km Northeast of Mexico City (19°27'32'' N - 99°46'34.9'' W), and 2 548m above sea level. A summary of the physicochemical properties of the water body shows a mean temperature of 17.74°C, highly mineralized water by the conductivity and total dissolved solids values (185.5µS/cm, and 0.11g/L, respectively), high turbidity water (111.93NTU), and low values of dissolved oxygen (5.65mg/L), pH close to neutrality (7.43), and nitrogen compounds with dominance of oxidized forms (2.06mg/L, 0.42mg/L, and 0.04mg/L for nitrates, total ammonia, and nitrites, respectively), while the phosphates are lower (0.66mg/L), with hardness of 41mg/L. Crayfishes were captured via a 50x30cm rectangular dip net with a mesh size of 500µm, following the traditional form used on site, and immediately placed in plastic recipients protected from light (60x40x30cm), filled with water from the reservoir, and oxygenated with a pump. Temperature of about 18°C was controlled with reservoir water during the 2h transportation to the laboratory, where they were taxonomically identified as Cambarellus montezumae by M. Sc. Jacinto Elias Sedeño Díaz (member of Macrolatinos group of specialists in aquatic macroinvertebrates). Samples of the organism are in the Aquatic Macroinvertebrate Collection of the National School of Biological Sciences from the National Polytechnic Institute at Mexico City. Then, groups of15 individuals (4-6cm of diameter) were placed in recipients with 6L of reconstituted water (NaHCO3 174mg/L, MgSO4 120mg/L, KCl 6mg/L, and CaSO4 2H2O 120mg/L, at pH 7.8), as well as with gravel and the plant Elodea sp. The aquaria were equipped with systems to maintain the temperature and the aeration. Before the toxicological determinations, the crustaceans were maintained for at least three weeks at 18°C under constant aeration, and were protected from light and fed with Elodea and Artemia salina every third day.
Experimental design: In short term genotoxicity assays it is relevant to avoid or reduce the involvement of confounding effects that may affect the accuracy of the results; in DNA tests in particular, it is useful to discriminate between the true genotoxic effect respect to cytotoxic effects that may also damage the molecule. Therefore, it is recommended to have information on the cytotoxic or toxic effects of the tested compound. In this context, acute toxicity determination is suggested as a preliminary, helpful assay recommended for selecting a proportion of such value for genotoxic evaluation (Vasquez, 2010; Azqueta, Arbillaga, Lopez de Cerain, & Collins, 2013). In our case, we used about 10% of the determined LC50 as the high dose for the comet assay. Then, lipoperoxidation studies were made with the same concentrations, so as to have a more reliable comparison of results. The assay was carried out only in the hepatopancreas because most brain tissue (about 2mm in diameter) was occupied in the comet assay. Besides, hepatopancreas is the organ most actively involved in the metabolic processes. Finally, it is pertinent to mention that the design of the comet experiment was based in accepted recommendations by international agencies which have been followed by our group in previous reports, and included five organisms per group and 100 nucleoids per individual, which have been validated to support the accuracy of the statistical evaluation (Tice et al., 2000; Madrigal-Santillán, Morales-González, Sánchez-Gutiérrez, Reyes-Arellano, & Madrigal-Bujaidar, 2009; García-Medina et al., 2011).
Acute toxicity: EPA Regional and State personnel have generally observed that it is not necessary to conduct a toxicity range-finding test prior to performing a static, acute, toxicity test (Environmental Protection Agency, 1991). However, we followed their suggestion for samples of unknown quality. A down-scaled abbreviated test with groups of five organisms exposed to widely spaced sample dilutions in logarithmic series, from 100% to 0.1%, was carried out. Based on this preliminary data, we applied the definitive test in duplicate. We used animals which were not fed for 24h before the assay. A total of 15 organisms per recipient were then exposed to the tested compounds in the conditions indicated above. In the case of DD, we used 10, 20, 30, 40, and 50µg/L, and in regard to CP, the tested concentrations were 0.04, 0.2, 0.9, 4, 20, and 90µg/L. For the control group, the pesticides' solvent was used (0.1% ethanol). As indicated, the tested range of concentrations was based in preliminary assays. Mortality and behavioural changes were registered at 72h. To obtain the LC50 value, we applied the EPA Probit Analysis Program, version 1.5.
DNA damage determined with the comet assay. Crayfishes exposed to the insecticides in reconstituted water: A total of 15 animals per recipient were exposed to 0.05 and 0.5µg/L of each insecticide, and a group exposed to 0.1% ethanol was used as control. A dissection of the brain and the hepatopancreas was performed in each of five organisms at 24, 48, and 72h. Each organ (15mg of the hepatopancreas, and 5mg of the brain) were placed in 30µL and 20µL of cold PBS, respectively, and homogenized to finally obtain a cell suspension for performing the comet assay. This step is important to maintain the cell isotonicity and high cell viability, as well as to avoid any damage to the tissue that may produce false positive data.
The alkaline comet assay was performed according to the method previously described with slight modifications (García-Medina et al., 2011). Initially, we determined the cellular viability by applying the trypan blue staining. We found a mean of 95.0% and 94.5% of viable cells for the brain and the hepatopancreas, respectively. Then, 20µL of the brain suspension or 10µL of the hepatopancreas was mixed with 75µL of 0.5% LMPA, and placed in a fully frosted, pre-coated microscope slide with 100µL of 1% NMPA. Finally, a third layer of LMPA was added on top. The preparation was then left for 5min at 4°C to allow solidification, and placed at 4°C for 1h in the lisis solution at pH 10 (NaCl 2.5M, EDTA 100mM, trisma base 10mM, 10% DMSO, and 1% triton X-100). Slides were put in a horizontal electrophoresis chamber, where the cells were exposed to an alkaline solution (NaOH 300mM, and EDTA 1mM at pH > 13) to allow DNA unwinding and expression of alkali-labile sites. In the next steps, cells were electrophoresed at 300mA, 25V, and pH>13 for 20min. The process was neutralized by rinsing the slides in trisma base 0.4M at pH 7.4, and finally, the nucleoids of each slide were stained with 50µL of EB (2µg/mL). The analysis was made at 400X magnification by means of a epifluorescent microscope (Axioscope, Carl Zeiss) equipped with a digital camera and a software for the capture, processing and image analysis (Zeiss KS 400 version 3.01). In 100 nucleoids per sample, the length to width index was determined by measuring the image length (T) and dividing the result by nuclear diameter (N), so as to determine the T/N index. Therefore, in this procedure, two distinct measurements per nucleoid were made (Clements, Ralph, & Petras, 1997; Garcia-Medina et al., 2011).
Protein concentration in crayfishes exposed to the insecticides in reconstituted water: Protein quantification was made using the method of Bradford (1976) in five organisms per group. We added 2.5mL of Bradford's reactive to 25µL of the hepatopancreas cell suspension (supernatant) of each of the insecticide exposed and control animals, mixed it, and permitted it to rest for 5min at room temperature before determining the absorbance at 595nm. The readings were interpolated in an albumin type curve made with concentrations from 0.01 to 0.1mg/mL.
Lipid peroxidation in crayfishes exposed to the insecticides in reconstituted water: For this evaluation, in each of the exposed and control animals, we quantified the level of MDA according to the method of Hoving et al. (1992), with slight modifications. The hepatopancreas was homogenized, placed in a tube with 5mL of KCl 0.15M, and centrifuged at 14 000g at -5°C for 15min. Then, 250µL of the supernatant was mixed with 10µL of an aqueous solution of reduced glutation 16.3nmol/L plus 10µL of a methanol solution of BHT 136mmol/L, and 5mL of a mix formed by hexane/isopropanol (3:2v/v). The tubes were agitated for 30min and the solution was centrifuged at 1 000g. Then, to 300µL of each sample, we added 700µL of PBS (pH 7.4) plus the reaction solution constituted by 15% TCA and 15% TBA. The mixture was incubated at 37°C for 45min, centrifuged at 885g for 5min, and the absorbance was registered at 535nm. Results were expressed in mM of MDA/mg protein using a molar extinction coefficient of 1.56 x 105/M.cm.
DNA damage and lipoperoxidation in crayfishes exposed to the insecticides in water from the reservoir: For the comet assay, organisms from the laboratory facilities were placed in recipients with water from the same place where they were collected. Fifteen animals per aquarium, each with 6L of water, were exposed to 0.05, and 0.5µg/L of DD and CP, respectively, as well as to 0.1% ethanol in the control group. Animals were maintained in similar conditions, as described for the previous study, and the determinations were also made in five organisms at 24, 48, and 72h as previously described. The comet assay was performed in both the brain and the hepatopancreas and the lipoperoxidative determination was carried out only for the hepatopancreas samples.
Results
Data registered for the mortality induced by the insecticides in reconstituted water were examined with the Probit test. In this form, at 72h of pesticide exposure, we calculated a LC50 for DD, which was equal to 5.1µg/L (95% CI; 1.7-18.9), and calculated 5.62µg/L (95% CI; 2.9-11.0) for CP.
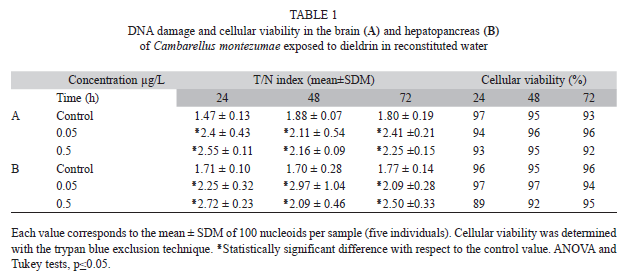
The effect of DD on the DNA of C. montezumae in reconstituted water is shown in table 1. The results obtained in the brain (Table 1A) indicated a similar T/N index mean value of 1.71 in the control animals, along the three selected time points. In comparison with the mean value of the control group, the mean increase observed in the insecticide-treated animals along the studied schedule was 34.5% and 35.6% for the low and high concentration, respectively. Regarding cell viability, the same table shows values higher than 90%.
With respect to hepatopancreas (Table 1B), we observed a similar effect as described above: a mean control level of 1.72 and a slightly higher DNA damage than the observed in the brain. In comparison with the untreated cell level, we found a mean increase of 41.2% for both the low and high concentration along the schedule.
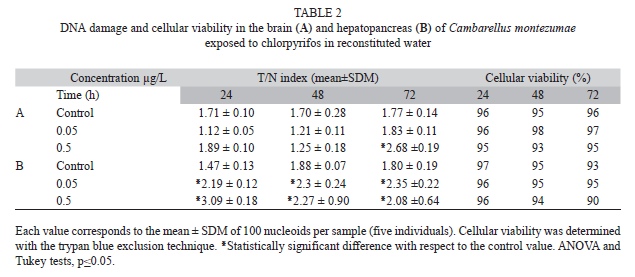
The effect of CP on the DNA of exposed crayfishes in reconstituted water is shown in Table 2. In the brain, we observed no elevation in the value of the T/N index induced by the tested concentrations, except for the effect of 0.5µg/L of CP at 72h, which induced 51.4% more DNA damage than the level of the control animals (Table 2A). Results concerning the hepatopancreas are shown in table 2B. In this organ, the DNA damage induced by CP was more clearly expressed, as shown by a T/N index increase induced with the two tested concentrations of the pesticide at the three evaluated times. The mean increase induced by the low and the high concentration, with respect to the mean control level, was of 33.3 and 45%, respectively. With respect to cellular viability, we found values higher than or equal to 90% in the cells of both brain and hepatopancreas.
Table 3 shows the lipoperoxidative activity induced by the insecticides in reconstituted water. Table 3A shows a constant low level of the activity in the hepatopancreas of control animals along the evaluated schedule, contrary to the significant effect induced by DD with the tested concentrations, as shown by the significant MDA level increase at the three observed time points. Table 3B shows a similar effect induced by CP: a constant low activity in control animals along the assay and a significant elevation of the MDA level with the tested concentrations of the chemical at the three examined times.
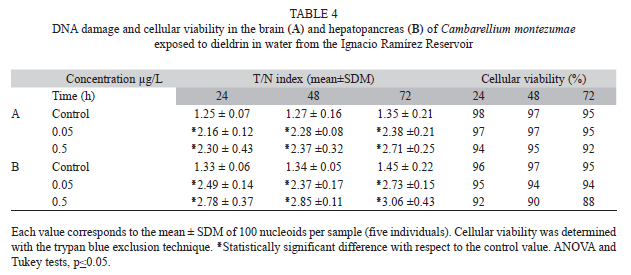
Results concerning the DNA damage in organisms exposed to the insecticides in the water of the reservoir (Tables 4 and 5) showed a stronger effect than the above indicated results (multifactorial ANOVA, p≤0.05). In the brain (Table 4A), data obtained after DD exposure indicated a significant increase in the T/N index induced by the two tested concentrations in the three evaluated times. The mean increase of the low and the high concentration, with respect to the control level, were 75.9 and 90.6%, respectively. In the hepatopancreas (Table 4B), we observed a higher elevation of DNA damage. With respect to the control value, the mean increase was 84.6 and 109% for 0.05 and 0.5µg/L of DD, respectively. In this assay, we also determined a certain decrease in the viability (6.3%) with the high concentration.
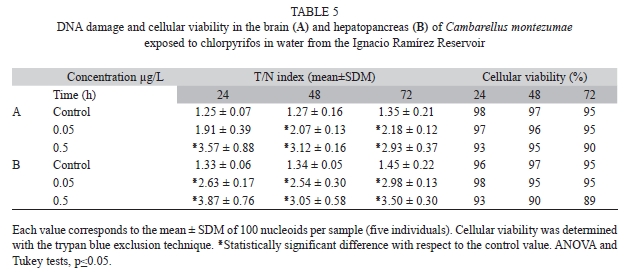
Table 5A shows the effect of CP in the brain of exposed animals. In these cells, we also found an increase in DNA damage by the pesticide. A mean elevation of 58.9 and 97% was found for the low and the high concentrations, respectively; and in the hepatopancreas (Table 5B), the results showed an increase of 101.5 and 126.2% with 0.05 and 0.5µg/L of CP, with respect to the mean control level. The cell viability in the examined organs was high, showing values ranging from 88 to 95%.
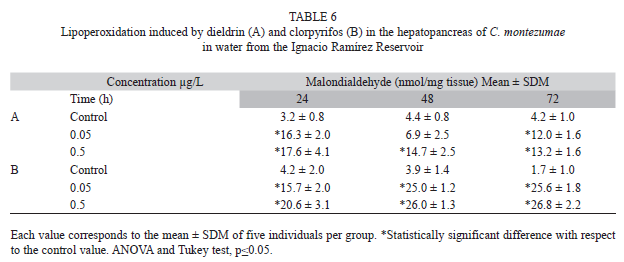
Lipid peroxidation by the insecticides in the water of the reservoir is shown in Table 6. It was observed that the tested concentrations of the two evaluated chemicals increased such activity. The observed lipid damage was somewhat similar to that found in reconstituted water.
Discussion
A number of aquatic crustaceans have been used to evaluate ecotoxicological risks; however, little information about the presently studied insecticides has been reported in these animals. In regard to acute toxicity testing, other pesticides have been studied. Per-methrin, for example, showed a mean static acute 96-h LC50 corresponding to 0.81mg/L in red swamp crayfish Procambarus clarkii; in the same organism, a contrasting LC50 value of 951µg/L was determined for the insecticide chlorantraniliprole, while for the neonicotinoids compounds thiametoxan and dinotefuran, authors reported values of 967µ/L and 2032µg/L, respectively; which are very different from the observed in the pyrethroids clotianidin, lambda-cyhalothrin, and etofen-prox: 59, 0.16 and 0.29µg/L, respectively (Jar-boe, & Romaire, 1991; Barbee, & Stout, 2009; Barbee, McClain, Lanka, & Stout, 2010). In the case of azadirachtin and two neem-based insecticides, their acute toxicity was determined in six aquatic animals, including Procambarus clarkii (Goktepe, Portier, & Ahmedna, 2004). In such study, Daphnia pulex was the only sensitive species toward the assayed pesticides. The expressed data showed high variability of the LC50 values obtained in freshwater crustaceans, which can be related with the species studied, length/weight of the animal, and the chemical structure of the evaluated pesticide, among other variables. These data point to the importance of determining such parameter in each studied organism. In our case, we found a value which was in the same range of the two evaluated insecticides, and that, in the context of the data reported above, suggests that the presently used organism was sensitive towards the tested insecticides, and therefore, appropriate for toxicological studies.
Genotoxic determination of the tested insecticides is relevant because of their known acute and chronic health risk for humans, and for aquatic ecosystems. Moreover, their persistence in the environment may increase such risk. For example, CP, which is one of the most widely used insecticide, has been detected in air, rain, marine sediments, surface waters, drinking water wells, and solid and liquid dietary samples from urban and rural areas (Gebremariam, Beutel, Yonge, Flury, & Harsh, 2012). In fact, the two insecticides have shown bioaccumulation in a number of organisms and organs due to their resistance to physical, biological, chemical and photochemical breakdown. They may remain mainly in lipids of various tissues and participate in long term-and chronic effects (Jorgenson, 2001; Mrema et al., 2013). Experimental studies have confirmed their persistence in the food chain; for example, Vara et al., (2002) detected CP in the small fish Aphanius after the consumption of contaminated Artemia, and suggested a risk for humans. Bioaccumulation has also been reported in various commercial fishes and bivalves (Satyanarayan, Ramakant, & Satyanarayan, 2005; Takabe et al., 2012); moreover, human risk has been documented (Lee et al., 2014). These authors confirmed that dietary exposure to banned organochlorine pesticides represents a higher risk to develop cancer. Therefore, an appropriate regulatory and technical infrastructure is required to monitor insecticides, and to verify their decline in the environment (Tsai, 2010). In these efforts, our results suggest that the comet assay may be useful to periodically monitor the correlation between the amount of insecticide and DNA damage. In this context, it is pertinent to note the strong sensitivity of DNA from the studied crayfish toward the effect of DD and CP. With as low as about 1/100 of their LC50 we observed high DNA breakage with low cellular lethality, suggesting then that the method is suitable to detect geno-toxicity by this type of agents.
Our positive findings are also interesting in the light that few studies have been made in aquatic animals exposed to pesticides by using the comet assay. However, DD had shown alterations in DNA synthesis and repair in mouse liver and brain (Bachowski, Xu, Stevenson, Walborg, & Klauning, 1998; Sava, Velasquez, Song, & Sanchez-Ramos, 2007), and in regard to CP, Galindo Reyes, Leyva, Millan, and Lazcano (2002) found DNA adducts and breaks in shrimp larvae by means of the alkaline unwinding method applied to extracts of DNA. Besides, the specificity of the comet assay has been reported in Astacus leptodactylus, where temperature increase (but no food deprivation or air exposure) had a correlation with an increase in DNA damage (Malev et al., 2010). Therefore, it should be also interesting to extend DNA damage studies to different aquatic insects. These animals spend their life time as larvae and adults in the water bodies and in the surroundings as well, and play an important role in the ecological equilibrium. An example of the potential usefulness of genotoxic determination refers to the contamination found in water bodies and sediments after pesticides are applied to control insects, particularly mosquitoes. For this purpose, ultra-low-volume aerosol applications have been developed to reduce the ecosystem and human damages; however, the hazard cannot be discarded (Pierce, Henry, Blum, & Mueller, 2005; Scheiler, & Peterson, 2010). In this situation, the sensitivity of the comet assay suggests that it can be applied for monitoring purposes, even in non-target organisms such as honeybee, butterfly, Daphnia, and Chironomus, which have been used to evaluate toxicity by pesticide spraying (Park, & Choi, 2009; Hoang, Pryor, Rand, & Frakes, 2011).
The DNA breaking ability of DD and CP may be related with disturbances of redox process that induce oxidative stress. The excessive generation of ROS causes damage to all vital macromolecules, including lipids, proteins and DNA. This indirect action of the chemicals on such molecules has been established by increases in thiobarbituric-acid-reactive species observed in various models, as well as by alterations of antioxidant defences, and elevation of DNA oxidation (Bagchi, Bagchi, Hassoun, & Stohs, 1995; Qiao, Seidler, & Slotkin, 2005; Martyniuk et al., 2010; Slotkin, & Seidler, 2010; Ozkan, Gündüz, Berkóz, Hunt, & Yalin, 2012). Our present data showing a lipid peroxidation increase in crayfish by the insecticides agree with such findings, and suggest a relation of such damage with the observed DNA breakage. However, such correlation should be confirmed with oxidative studies made in the same experimental conditions in proteins, DNA, or antioxidant enzymes. Besides, the testing of other concentrations should be of relevance for determining a concentration-effect relationship that was not observed in the present study.
On the other hand, when C. montezumae was exposed to the pesticides in the water of the reservoir, we observed a higher DNA damage than that determined in reconstituted water. This effect is difficult to explain. A possibility is related with the agricultural activities made in the surroundings of the water body that could contribute with other toxic agents; moreover, examination of water and biota from the dam has shown the presence of metals, as well as organochlorine and organophosphate agents (Favari, López, Martinez, & Diaz, 2002; Galar, Martínez, Sánchez, & López, 2006). Reports on the matter have shown that indeed, polluted water, in six days, may give rise to metal accumulation in the tissue of Procambarus clarkii (Alcorlo, Otero, Crehuet, Baltanás, & Montes, 2006). Moreover, caged freshwater Astacus leptodactylus collected from a reference site and placed for seven days in locations of a river with variable degrees of pollution was able to show a concordant degree of DNA damage, with respect to the level of pollution (Klobucar et al., 2012). However, interaction among water compounds may affect the final result. In some cases, it has been suggested that antagonistic effect may reduce the toxicity (Favari et al., 2002). Therefore, other plausible explanation refers to the physicochemical characteristics of the water of the reservoir, which include high turbidity, probably related with clay, where pollutants can be adsorbed and may interact with the tested insecticides, and the presence of nitrogen compounds, such as ammonia and nitrites, which may increase the detected toxicity, besides of the detected coliforms bacteria in the water body; data which contrast with the controlled laboratory conditions including a 12h dark: light photoperiod, 8.0mg/mL of dissolved oxygen, no turbidity and no contamination. Presently, it is becoming clear that a number of physicochemical properties may act as confounding factors and interfere with the biological effects, thus it has been recommended to include the two aspects for risk assessment studies (Vasquez, & Fatta, 2013).
Freshwater reservoirs can be impacted by a number of hazardous substances mainly related with anthropogenic activities; thus, water quality assessment is relevant for the implementation of the monitoring and remediation programs to minimize the risk of hazardous substances in aquatic ecosystems. Macroinvertebrates play a significant ecological role, since they are an important link between energy inputs and their transfer to other trophic levels through the food web. They participate in the balance between heterotrophs (depending on organic matter from terrestrial origin) and autotrophs that depend on primary production of the periphyton and macrophytes (Hanson, Springer, & Ramírez, 2010). Cambarellus montezumae is an omnivorous crayfish and the food of several fishes and aquatic birds. It is also in contact with contaminants entering the system, particularly with the sediments that accumulate and transfer pollutants to other trophic levels, and thus, able to accumulate contaminants from sediments or the water column. Therefore, they are especially useful as test organisms for identifying the acute and chronic effects of specific pollutants. Our study demonstrated the usefulness of the comet assay applied to this organism. Moreover, its accuracy can be increased when coupled with the examination of other cell/tissue biomarkers, as shown in reports related with the presence of metals, organic pollutants, and environmental disturbances (Antón, Serrano, Angulo, Ferrero, & Rallo, 2000; Holmqvist et al., 2007).
In conclusion, results of the present study established a high efficacy of the comet assay to demonstrate DNA damage induced by the tested insecticides in C. montezumae, as well as a possible relation of such injury with an elevated oxidative stress. Therefore, our approach may be useful for future studies to evaluate bioaccumulation in the same or in other aquatic organisms.
References
Alcorlo, P., Otero, M., Crehuet, M., Baltanás, A., & Montes, C. (2006). The use of the red swamp crayfish (Procambarus clarkii, Girard) as indicator of the bioavailability of heavy metals in environmental monitoring in the River Guadiamar (SW, Spain). The Science of the Total Environment, 366, 380-390. [ Links ]
Ali, D., Nagpure, N. S., Kumar, S., Kumar, R., Kushwaha, B., & Lakra, W. S. (2009). Assesssment of genotoxic and mutagenic effects of chlorpyrifos in freshwater fish Channa punctatus (Bloch) using micronucleus assay and alkaline-single cell gel electrophoresis. Food and Chemical Toxicology, 47, 650-656. [ Links ]
Antón, A., Serrano, T., Angulo, E., Ferrero, G., & Rallo, A. (2000). The use of two species of crayfish as environmental sentinels: the relationship between heavy metal content, cell and tissue biomarkers and physic-chemical characteristics of the environment. The Science of the Total Environment, 247, 239-251. [ Links ]
Azqueta, A., Arbillaga, L., López de Cerain, A., & Collins, A. (2013). Enhancing the sensitivity of the comet assay as a genotoxicity test, by combining it with bacterial repair enzyme FPG. Mutagenesis, 28, 271-277. [ Links ]
Bachowski, S., Xu, Y., Stevenson, D. E., Walborg, E. F. Jr., & Klaunig, J. E. (1998). Role of oxidative stress in the selective toxicity of dieldrin in the mouse liver. Toxicology and Applied Pharmacology, 150, 301-309. [ Links ]
Bagchi, D., Bagchi, M., Hassoun, E. A., & Stohs, S. J. (1995). In vitro and in vivo generation of reactive oxygen species, DNA damage and lactate dehydrogenase leakage by selected pesticides. Toxicology, 704, 129-140. [ Links ]
Barbee, G. C., & Stout, M. J. (2009). Comparative acute toxicity of neonicotinoid and pyrethroid insecticides to non-target crayfish (Procambarus clarkii) associated with rice-crayfish crop rotations. Pest Management Science, 65, 1250-1256. [ Links ]
Barbee, G. C., McClain, W. R., Lanka, S. K., & Stout, M. J. (2010). Acute toxicity of chrorantraniliprole to nontarget crayfish (Procambarus clatkii) associated with rice-crayfish cropping systems. Pest Management Science, 66, 996-1001. [ Links ]
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microorganism quantities of protein utilizing the principle of protein dye binding. Analytical Biochemistry, 72, 248-254. [ Links ]
Chimeddorj, T., Suzuki, T., Murakane, K., Inai, M., Satoh, M., & Oyama, Y. (2013). Synergistic increase in cell lethality by dieldrin and H2O2 in rat thymocytes: effect of dieldrin on the cells exposed to oxidative stress. Chemosphere, 93, 353-358. [ Links ]
Cicchetti, R., Bari, M., & Argentin, G. (1999). Induction of micronuclei in bone marrow by two pesticides and their differentiation with CREST staining: an in vivo study in mice. Mutation Research, 439, 239-248. [ Links ]
Clements, C., Ralph S., & Petras, M. (1997). Genotoxicity of select herbicides in Rana catesbeiana tadpoles using the alkaline single-cell gel DNA electrophoresis (comet) assay. Environmental and Molecular Mutagenesis, 29, 277-288. [ Links ]
Cooper, J., & Dobson, H. (2007). The benefits of pesticides to mankind and the environment. Crop Protection, 26, 1337-1348. [ Links ]
Cotelle, S., & Férard, J. F. (1999). Comet assay in genetic ecotoxicology: A review. Environmental and Molecular Mutagenesis, 34, 246-255. [ Links ]
Eaton, D. L., Daroff, R. B., Autrup, H., Bridges, J., Buffler, P., Costa, L. G., Coyle, J., McKhann, G., Mobley, W. C., Nadel, L., Neubert, D., Schulte-Hermann, R., & Spencer, P. S. (2008). Review of the toxicology of chlorpyrifos with an emphasis on human exposure and neurodevelopment. Critical Reviews in Toxicology, 38, 1-125. [ Links ]
Environmental Protection Agency. (1991). Methods for measuring the acute toxicity of effluents and receiving waters to freshwater and marine organisms. EPA-600/4-90-027. Retrieved from http://nepis.epa.gov/Adobe/PDF/200096OT.PDF [ Links ]
Favari, L., López, E., Martinez-Tabche, L., & Díaz-Pardo, E. (2002). Effect of insecticides on plankton and fish of Ignacio Ramírez reservoir (Mexico): a biochemical and biomagnification study. Ecotoxicology and Environmental Safety, 51, 177-186. [ Links ]
Galar, M. M., Martínez-Tabche, L., Sánchez-Hidalgo, E., & López, E. (2006). Effect of zinc-enriched sediments, in open and isolatyed systems, on three species of benthonic invertebrates. Revista de Biología Tropical, 54, 451-460. [ Links ]
Galindo Reyes, J. G., Leyva, N. R., Millan, O. A., & Laz-cano, G. A. (2002). Effects of pesticides on DNA and protein of shrimp larvae Litopenaeus stylirostris of the California Gulf. Ecotoxicology and Environmental Safety, 53, 191-195. [ Links ]
García-Medina, S., Razo-Estrada, C., Galar-Martinez, M., Cortéz-Barberena, E., Gómez-Oliván, L. M., Alvarez-González, I., & Madrigal-Bujaidar, E. (2011). Geno-toxic and cytotoxic effects induced by aluminum in the lymphocytes of the common carp (Cyprinus carpio). Comparative Biochemistry and Physiology. Part C: Toxicology & Pharmacology, 153, 113-118. [ Links ]
Gebremariam, S. Y., Beutel, M. W., Yonge, D. R., Flury, M., & Harsh, J. B. (2012). Adsortion and desortion of chlorpyrifos to soils and sediments. Review of Environmental Contamination and Toxicology, 215, 123-175. [ Links ]
Goktepe, I., Portier, R., & Ahmedna, M. (2004). Ecological risk assessment of neem-based pesticides. Journal of Environmental Science and Health. Part B, Pesticides, Food contaminants, and Agricultural, 39, 311-320. [ Links ]
Gollapudi, B. B., Mendrala, A. L., & Linscombe, V. A. (1995). Evaluation of the genetic toxicity of the organophosphate insecticide chlorpyrifos. Mutation Research, 342, 25-36. [ Links ]
Gupta, S. C., Mishra, M., Sharma, A., Deepak Balaji, T. G., Kumar, R., Mishra, R. K., & Chowdhuri, D. K. Chlorpyrifos induces apoptosis and DNA damage in Drosophila through generation of reactive oxygen species. Ecotoxicology and Environmental Safety, 73, 1415-1423. [ Links ]
Hanson, P., Springer, M., & Ramírez, A. (2010). Introducción a los grupos de macroinvertebrados acuáticos. Revista de Biología Tropical, 58, 3-37. [ Links ]
Hatcher, J. M., Richardson, J. R., Guillot, T. S., McCor-mack, A. L., Di Monte, D. A., Jones, D. P., Pennell, K. D., & Miller, G. W. (2007). Dieldrin exposure induces oxidative damage in the mouse nigrostriatal dopamine system. Experimental Neurology, 204, 619-630. [ Links ]
Hoang, T. C., Pryor, R. L., Rand, G. M., & Frakes, R. A. Use of butterflies as nontarget insect test species and the acute toxicity and hazard of mosquito control insecticides. Environmental Toxicology and Chemistry, 30, 997-1005. [ Links ]
Holmqvist, N., Stenroth, P., Berglund, O., Nystrom, P., Graneli, W., & Larsson, P. (2007). Persistent organic pollutants (POP) in a benthic omnivore-a comparison between lake and stream crayfish populations. Chemosphere, 66, 1070-1078. [ Links ]
Hoving, E. B., Laing, C., Rutgers, H. M., Teggeler, M., van Doormaal, J. J., & Muskiet, F. A. (1992). Optimized determination of malondialdehyde in plasma lipid extracts using 1,3-diethyl-2-thiobarbituric acid: influence of detection method and relation with lipids and fatty acids in plasma from healthy adults. Clinica Chimica Acta, 208, 63-76. [ Links ]
Jarboe, H. H., & Romaire, R. P. (1991). Acute toxicity of permethrin to four size classes of red swamp crayfish (Procambarus clarkia) and observations of postexposure effects. Archives of Environmental Contamination and Toxicology, 20, 337-342. [ Links ]
Jorgenson, J. L. (2001). Aldrin and dieldrin: a review of research on their production, environmental deposition and fate, bioaccumulation, toxicology, and epidemiology in the United States. Environmental Health Perspectives, 109, 113-139. [ Links ]
Klobucar, G. I., Malev, O., Srut, M., Stambuk, A., Lorenzon, S., Cvetkovic, Z., Ferrero, E. A., & Maguire, I. (2012). Genotoxicity monitoring of freshwater environments using caged crayfish (Astacus leptodactylus). Chemosphere, 87, 62-67. [ Links ]
Landrigan, P. J. (2010). What causes autism? Exploring the environmental contribution. Current Opinion in Pediatrics, 22, 219-225. [ Links ]
Lee, S. H., Ra, J. S., Choi, J. W., Yim, B. J., Jung, M. S., & Kim, S. D. (2014). Human health risks associated with dietary exposure to persistent organic pollutants (POPs) in river water in Korea. The Science of the Total Environment, 470-471, 1362-1369. [ Links ]
Madrigal-Santillán, E., Morales-González, J. A., Sánchez-Gutiérrez, M., Reyes-Arellano, A., & Madrigal-Bujaidar, E. (2009). Investigation on the protective effect of α-mannan against the DNA damage induced by aflatoxin B1 in mouse hepatocytes. International Journal of Molecular Science, 10, 395-406. [ Links ]
Malev, O., Srut, M., Maguire, I., Stambuk, A., Ferrero, E. A., Lorenzon, S., & Klobucar, G. I. (2010). Genotoxic, physiological and immunological effects caused by temperature increase, air exposure or food deprivation in freshwater crayfish Astacus leptodactylus. Comparative Biochemistry and Physiology. Part C: Toxicology & Pharmacology, 152, 433-443. [ Links ]
Marnett, L. J. (2002). Oxy radicals, lipid peroxidation and DNA damage. Toxicology, 181-182, 219-222. [ Links ]
Martyniuk, C. J., Feswick, A., Spade, D. J., Kroll, K. J., Barber, D. S., & Denslow, N. D. (2010). Effects of acute dieldrin exposure on neurotransmitters and global gene transcription in largemouth bass (Micropterus salmoides) hypothalamus. Neurotoxicology, 31, 356-366. [ Links ]
Matsumoto, E., Kawanaka, Y., Yun, S. J., & Oyaizu, H. (2009). Bioremediation of the organochlorine pesticides, dieldrin, and endrin, and their occurrence in the environment. Applied Microbiology and Biotechnology, 84, 205-216. [ Links ]
Mrema, E. J., Rubino, F. M., Brambilla, G., Moretto, A., Tsatsakis, A. M., & Colosio C. (2013). Persistent organochlorinated pesticides and mechanisms of their toxicity. Toxicology, 307, 74-88. [ Links ]
Murray, T. J., Lea, R. G., Abramovich, D. R., Haites, N. E., & Fowler, P. A. (2001). Endocrine disrupting chemicals: effects on human male reproductive health. Early Pregnancy, 5, 80-112. [ Links ]
Narra, M. R. (2014). Tissue-Specific recovery of oxidative and antioxidant effects of chlorpyrifos in the freshwater crab, Batytelphusa guerini. Archives of Environmental Contamination Toxicology, 67, 158-166. [ Links ]
Ozkan, F., Gündüz, S. G., Berkóz, M., Hunt, A. O., & Yalin, S. (2012). The protective role of ascorbic acid (vitamin C) against chlorpyrifos-induced oxidative stress in Oreochromis niloticus. Fish Physiology and Biochemistry, 38, 635-643. [ Links ]
Palma, P., Palma, V. L., Fernandes, R. M., Soares, A. M., & Barbosa, I. R. (2008). Acute toxicity of atrazine, endosulfan sulphate and chrorpyrifos to Vibrio fischeri, Thamnocephalus platyurus, and Daphnia magna, relative to their concentrations in surface waters from the Alentejo region of Portugal. Bulletin of Environmental Contamination and Toxicology, 81, 485-489. [ Links ]
Pandey, R. M. (2008). Cytotoxic effects of pesticides in somatic cells of Vicia faba L. Cytology and Genetics, 42, 373-377. [ Links ]
Park, S. Y., & Choi, J. (2009). Genotoxic effects of non-ylphenol and bisphenol A exposure in aquatic biomonitoring species: freshwater crustacean, Daphnia magna, and aquatic midge, Chironomus riparius. Bulletin of Environmental Contamination and Toxicology, 83, 463-468. [ Links ]
Pereira, J. L., Antunes, S. C., Castro, B. B., Marques, C. R., Gonçalves, A. M., Gonçalves, F., & Pereira, R. (2009). Toxicity evaluation of three pesticides on non-target aquatic and soil organisms: commercial formulation versus active ingredient. Ecotoxicology, 18, 455-463. [ Links ]
Perobelli, J. E., Martinez, M. F., da Silva Franchi, C. A., Fernandez, C. D., de Camargo, J. L., & Kempinas, W. G. (2010). Decreased sperm motility in rats orally exposed to single or mixed pesticides. Journal of Toxicology and Environmental Health, 73, 991-1002. [ Links ]
Pierce, R. H., Henry, M. S., Blum, T. C., & Mueller, E. M. (2005). Aerial and tidal transport of mosquito control pesticides into the Florida Keys National Marine Sanctuary. Revista de Biología Tropical, 53, 117-125. [ Links ]
Pizzimenti, S., Ciamporcero, E., Daga, M., Pettazzoni, P., Arcaro, A., Cetrangolo, G., Minelli, R., Dianzani, C., Lepore, A., Gentile, F., & Barrera, G. (2013). Interaction of aldehydes derived from lipid peroxidation and membrane proteins. Frontiers in Physiology, 4, 242. [ Links ]
Qiao, D., Seidler, F. J., & Slotkin, T. A. (2005). Oxidative mechanisms contributing to the developmental neu-rotoxicity of nicotine and chlorpyrifos. Toxicology and Applied Pharmacoogy, 206, 17-26. [ Links ]
Rahman, M. F., Mahboob, M., Danadevi, K., Saleha, B. B., & Groover, P. (2002). Assessment of genotoxic effects of chorpyriphos and acephate by the comet assay in mice leucocytes. Mutation Research, 516, 139-147. [ Links ]
Satyanarayan, S., Ramakant, R., & Satyanarayan, A. (2005). Bioaccumulation studies of organochlorinated pesticides in tissues of Ciprinus carpio. Journal of Environmental Science and Health. Part B. Pesticides, Food contaminants, and Agricultural Wastes, 40, 397-412. [ Links ]
Sava, V., Velasquez, A., Song, S., & Sanchez-Ramos, J. (2007). Dieldrin elicits a widespread DNA repair and antioxidative response in mouse brain. Journal of Biochemical and Molecular Toxicology, 21, 125-135. [ Links ]
Scheiler, J. J., & Peterson, R. K. (2010). Toxicity and risk of permethrin and naled to non-target insects after adult mosquito management. Ecotoxicology, 19, 1140-1146. [ Links ]
Slotkin, T. A., & Seidler, F. J. (2010). Oxidative stress from diverse developmental neurotoxicants: antioxidants protect against lipid peroxidation without preventing cell loss. Neurotoxicology and Teratology, 32, 124-131. [ Links ]
Swaen, G. M., de Jong, G., Slangen, J. J., & van Amelsvoort, L. G. (2002). Cancer mortality in workers exposed to dieldrin and aldrin: an update. Toxicology and Industrial Health, 18, 63-70. [ Links ]
Takabe, Y., Tsuno, H., Nishimura, F., Tanji, N., Maruno, H., Tsurukawa, M., Suzuki, M., & Matsumura, C. (2012). Bioaccumulation and primary risk assessment of persistent organic pollutants with various bivalves. Water Science and Technology, 66, 2620-2629. [ Links ]
Tice, R. R., Agurell, E., Anderson, D., Burlinson, B., Hartmann, A., Kobayashi, H., Miyamae, Y., Rojas, E., Ryu, J. C., & Sasaki, Y. F. (2000). Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environmental and Molecular Mutagenesis, 35, 206-221. [ Links ]
Tsai, W. T. (2010). Current status and regulatory aspects of pesticides considered to be persistent organic pollutants (POPs) in Taiwan. International Journal of Environmental Research and Public Health, 7, 3615-3627. [ Links ]
Varó, I., Serrano, R., Pitarch, E., Amat, F., López, F. J., & Navarro, J. C. (2002). Bioaccumulation of chlorpyrifos through an experimental food chain: study of protein HSP70 as biomarker of sublethal stress in fish. Archives of Environmental Contamination and Toxicology, 42, 229-235. [ Links ]
Vasquez, M. I., & Fatta-Kassinos, D. (2013). Is the evaluation of "traditional" physicochemical parameters sufficient to explain the potential toxicity of the treated wastewater at sewage treatment plants? Environmental Science and Pollutution Research International, 20, 3516-3528. [ Links ]
Vasquez, M. Z. (2010). Combining the in vivo comet and micronucleus assays: a practical approach to genotoxicity testing and data interpretation. Mutagenesis, 25, 187-199. [ Links ]
Verma, R. S., & Srivastava, N. (2003). Effect of chlorpyrifos on thiobarbituric acid reactive substances, scavenging enzymes and glutathione in rat tissues. Indian Journal of Biochemical and Biophysics, 40, 423-428. [ Links ]
Voulgaridou, G. P., Anestopoulos, I., Franco, R., Panayiotidis, M. I., & Pappa, A. (2011). DNA damage induced by endogenous aldehydes: current state of knowledge. Mutation Research, 711, 13-27. [ Links ]
Yin, X., Zhu, G., Li, X. B., & Liu, S. (2009). Genotoxicity evaluation of chlorpyrifos to amphibian Chinese toad (Amphibian: Anura) by comet assay and micronucleus test. Mutation Research, 680, 2-6. [ Links ]
Ali, D., Nagpure, N. S., Kumar, S., Kumar, R., Kushwaha, B., & Lakra, W. S. (2009). Assesssment of genotoxic and mutagenic effects of chlorpyrifos in freshwater fish Channa punctatus (Bloch) using micronucleus assay and alkaline-single cell gel electrophoresis. Food and Chemical Toxicology, 47, 650-656. [ Links ]
Antón, A., Serrano, T., Angulo, E., Ferrero, G., & Rallo, A. (2000). The use of two species of crayfish as environmental sentinels: the relationship between heavy metal content, cell and tissue biomarkers and physic-chemical characteristics of the environment. The Science of the Total Environment, 247, 239-251. [ Links ]
Azqueta, A., Arbillaga, L., López de Cerain, A., & Collins, A. (2013). Enhancing the sensitivity of the comet assay as a genotoxicity test, by combining it with bacterial repair enzyme FPG. Mutagenesis, 28, 271-277. [ Links ]
Bachowski, S., Xu, Y., Stevenson, D. E., Walborg, E. F. Jr., & Klaunig, J. E. (1998). Role of oxidative stress in the selective toxicity of dieldrin in the mouse liver. Toxicology and Applied Pharmacology, 150, 301-309. [ Links ]
Bagchi, D., Bagchi, M., Hassoun, E. A., & Stohs, S. J. (1995). In vitro and in vivo generation of reactive oxygen species, DNA damage and lactate dehydrogenase leakage by selected pesticides. Toxicology, 704, 129-140. [ Links ]
Barbee, G. C., & Stout, M. J. (2009). Comparative acute toxicity of neonicotinoid and pyrethroid insecticides to non-target crayfish (Procambarus clarkii) associated with rice-crayfish crop rotations. Pest Management Science, 65, 1250-1256. [ Links ]
Barbee, G. C., McClain, W. R., Lanka, S. K., & Stout, M. J. (2010). Acute toxicity of chrorantraniliprole to nontarget crayfish (Procambarus clatkii) associated with rice-crayfish cropping systems. Pest Management Science, 66, 996-1001. [ Links ]
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microorganism quantities of protein utilizing the principle of protein dye binding. Analytical Biochemistry, 72, 248-254. [ Links ]
Chimeddorj, T., Suzuki, T., Murakane, K., Inai, M., Satoh, M., & Oyama, Y. (2013). Synergistic increase in cell lethality by dieldrin and H2O2 in rat thymocytes: effect of dieldrin on the cells exposed to oxidative stress. Chemosphere, 93, 353-358. [ Links ]
Cicchetti, R., Bari, M., & Argentin, G. (1999). Induction of micronuclei in bone marrow by two pesticides and their differentiation with CREST staining: an in vivo study in mice. Mutation Research, 439, 239-248. [ Links ]
Clements, C., Ralph S., & Petras, M. (1997). Genotoxicity of select herbicides in Rana catesbeiana tadpoles using the alkaline single-cell gel DNA electrophoresis (comet) assay. Environmental and Molecular Mutagenesis, 29, 277-288. [ Links ]
Cooper, J., & Dobson, H. (2007). The benefits of pesticides to mankind and the environment. Crop Protection, 26, 1337-1348. [ Links ]
Cotelle, S., & Férard, J. F. (1999). Comet assay in genetic ecotoxicology: A review. Environmental and Molecular Mutagenesis, 34, 246-255. [ Links ]
Eaton, D. L., Daroff, R. B., Autrup, H., Bridges, J., Buffler, P., Costa, L. G., Coyle, J., McKhann, G., Mobley, W. C., Nadel, L., Neubert, D., Schulte-Hermann, R., & Spencer, P. S. (2008). Review of the toxicology of chlorpyrifos with an emphasis on human exposure and neurodevelopment. Critical Reviews in Toxicology, 38, 1-125. [ Links ]
Environmental Protection Agency. (1991). Methods for measuring the acute toxicity of effluents and receiving waters to freshwater and marine organisms. EPA-600/4-90-027. Retrieved from http://nepis.epa.gov/Adobe/PDF/200096OT.PDF [ Links ]
Favari, L., López, E., Martinez-Tabche, L., & Díaz-Pardo, E. (2002). Effect of insecticides on plankton and fish of Ignacio Ramírez reservoir (Mexico): a biochemical and biomagnification study. Ecotoxicology and Environmental Safety, 51, 177-186. [ Links ]
Galar, M. M., Martínez-Tabche, L., Sánchez-Hidalgo, E., & López, E. (2006). Effect of zinc-enriched sediments, in open and isolatyed systems, on three species of benthonic invertebrates. Revista de Biología Tropical, 54, 451-460. [ Links ]
Galindo Reyes, J. G., Leyva, N. R., Millan, O. A., & Laz-cano, G. A. (2002). Effects of pesticides on DNA and protein of shrimp larvae Litopenaeus stylirostris of the California Gulf. Ecotoxicology and Environmental Safety, 53, 191-195. [ Links ]
García-Medina, S., Razo-Estrada, C., Galar-Martinez, M., Cortéz-Barberena, E., Gómez-Oliván, L. M., Alvarez-González, I., & Madrigal-Bujaidar, E. (2011). Geno-toxic and cytotoxic effects induced by aluminum in the lymphocytes of the common carp (Cyprinus carpio). Comparative Biochemistry and Physiology. Part C: Toxicology & Pharmacology, 153, 113-118. [ Links ]
Gebremariam, S. Y., Beutel, M. W., Yonge, D. R., Flury, M., & Harsh, J. B. (2012). Adsortion and desortion of chlorpyrifos to soils and sediments. Review of Environmental Contamination and Toxicology, 215, 123-175. [ Links ]
Goktepe, I., Portier, R., & Ahmedna, M. (2004). Ecological risk assessment of neem-based pesticides. Journal of Environmental Science and Health. Part B, Pesticides, Food contaminants, and Agricultural, 39, 311-320. [ Links ]
Gollapudi, B. B., Mendrala, A. L., & Linscombe, V. A. (1995). Evaluation of the genetic toxicity of the organophosphate insecticide chlorpyrifos. Mutation Research, 342, 25-36. [ Links ]
Gupta, S. C., Mishra, M., Sharma, A., Deepak Balaji, T. G., Kumar, R., Mishra, R. K., & Chowdhuri, D. K. Chlorpyrifos induces apoptosis and DNA damage in Drosophila through generation of reactive oxygen species. Ecotoxicology and Environmental Safety, 73, 1415-1423. [ Links ]
Hanson, P., Springer, M., & Ramírez, A. (2010). Introducción a los grupos de macroinvertebrados acuáticos. Revista de Biología Tropical, 58, 3-37. [ Links ]
Hatcher, J. M., Richardson, J. R., Guillot, T. S., McCor-mack, A. L., Di Monte, D. A., Jones, D. P., Pennell, K. D., & Miller, G. W. (2007). Dieldrin exposure induces oxidative damage in the mouse nigrostriatal dopamine system. Experimental Neurology, 204, 619-630. [ Links ]
Hoang, T. C., Pryor, R. L., Rand, G. M., & Frakes, R. A. Use of butterflies as nontarget insect test species and the acute toxicity and hazard of mosquito control insecticides. Environmental Toxicology and Chemistry, 30, 997-1005. [ Links ]
Holmqvist, N., Stenroth, P., Berglund, O., Nystrom, P., Graneli, W., & Larsson, P. (2007). Persistent organic pollutants (POP) in a benthic omnivore-a comparison between lake and stream crayfish populations. Chemosphere, 66, 1070-1078. [ Links ]
Hoving, E. B., Laing, C., Rutgers, H. M., Teggeler, M., van Doormaal, J. J., & Muskiet, F. A. (1992). Optimized determination of malondialdehyde in plasma lipid extracts using 1,3-diethyl-2-thiobarbituric acid: influence of detection method and relation with lipids and fatty acids in plasma from healthy adults. Clinica Chimica Acta, 208, 63-76. [ Links ]
Jarboe, H. H., & Romaire, R. P. (1991). Acute toxicity of permethrin to four size classes of red swamp crayfish (Procambarus clarkia) and observations of postexposure effects. Archives of Environmental Contamination and Toxicology, 20, 337-342. [ Links ]
Jorgenson, J. L. (2001). Aldrin and dieldrin: a review of research on their production, environmental deposition and fate, bioaccumulation, toxicology, and epidemiology in the United States. Environmental Health Perspectives, 109, 113-139. [ Links ]
Klobucar, G. I., Malev, O., Srut, M., Stambuk, A., Lorenzon, S., Cvetkovic, Z., Ferrero, E. A., & Maguire, I. (2012). Genotoxicity monitoring of freshwater environments using caged crayfish (Astacus leptodactylus). Chemosphere, 87, 62-67. [ Links ]
Landrigan, P. J. (2010). What causes autism? Exploring the environmental contribution. Current Opinion in Pediatrics, 22, 219-225. [ Links ]
Lee, S. H., Ra, J. S., Choi, J. W., Yim, B. J., Jung, M. S., & Kim, S. D. (2014). Human health risks associated with dietary exposure to persistent organic pollutants (POPs) in river water in Korea. The Science of the Total Environment, 470-471, 1362-1369. [ Links ]
Madrigal-Santillán, E., Morales-González, J. A., Sánchez-Gutiérrez, M., Reyes-Arellano, A., & Madrigal-Bujaidar, E. (2009). Investigation on the protective effect of α-mannan against the DNA damage induced by aflatoxin B1 in mouse hepatocytes. International Journal of Molecular Science, 10, 395-406. [ Links ]
Malev, O., Srut, M., Maguire, I., Stambuk, A., Ferrero, E. A., Lorenzon, S., & Klobucar, G. I. (2010). Genotoxic, physiological and immunological effects caused by temperature increase, air exposure or food deprivation in freshwater crayfish Astacus leptodactylus. Comparative Biochemistry and Physiology. Part C: Toxicology & Pharmacology, 152, 433-443. [ Links ]
Marnett, L. J. (2002). Oxy radicals, lipid peroxidation and DNA damage. Toxicology, 181-182, 219-222. [ Links ]
Martyniuk, C. J., Feswick, A., Spade, D. J., Kroll, K. J., Barber, D. S., & Denslow, N. D. (2010). Effects of acute dieldrin exposure on neurotransmitters and global gene transcription in largemouth bass (Micropterus salmoides) hypothalamus. Neurotoxicology, 31, 356-366. [ Links ]
Matsumoto, E., Kawanaka, Y., Yun, S. J., & Oyaizu, H. (2009). Bioremediation of the organochlorine pesticides, dieldrin, and endrin, and their occurrence in the environment. Applied Microbiology and Biotechnology, 84, 205-216. [ Links ]
Mrema, E. J., Rubino, F. M., Brambilla, G., Moretto, A., Tsatsakis, A. M., & Colosio C. (2013). Persistent organochlorinated pesticides and mechanisms of their toxicity. Toxicology, 307, 74-88. [ Links ]
Murray, T. J., Lea, R. G., Abramovich, D. R., Haites, N. E., & Fowler, P. A. (2001). Endocrine disrupting chemicals: effects on human male reproductive health. Early Pregnancy, 5, 80-112. [ Links ]
Narra, M. R. (2014). Tissue-Specific recovery of oxidative and antioxidant effects of chlorpyrifos in the freshwater crab, Batytelphusa guerini. Archives of Environmental Contamination Toxicology, 67, 158-166. [ Links ]
Ozkan, F., Gündüz, S. G., Berkóz, M., Hunt, A. O., & Yalin, S. (2012). The protective role of ascorbic acid (vitamin C) against chlorpyrifos-induced oxidative stress in Oreochromis niloticus. Fish Physiology and Biochemistry, 38, 635-643. [ Links ]
Palma, P., Palma, V. L., Fernandes, R. M., Soares, A. M., & Barbosa, I. R. (2008). Acute toxicity of atrazine, endosulfan sulphate and chrorpyrifos to Vibrio fischeri, Thamnocephalus platyurus, and Daphnia magna, relative to their concentrations in surface waters from the Alentejo region of Portugal. Bulletin of Environmental Contamination and Toxicology, 81, 485-489. [ Links ]
Pandey, R. M. (2008). Cytotoxic effects of pesticides in somatic cells of Vicia faba L. Cytology and Genetics, 42, 373-377. [ Links ]
Park, S. Y., & Choi, J. (2009). Genotoxic effects of non-ylphenol and bisphenol A exposure in aquatic biomonitoring species: freshwater crustacean, Daphnia magna, and aquatic midge, Chironomus riparius. Bulletin of Environmental Contamination and Toxicology, 83, 463-468. [ Links ]
Pereira, J. L., Antunes, S. C., Castro, B. B., Marques, C. R., Gonçalves, A. M., Gonçalves, F., & Pereira, R. (2009). Toxicity evaluation of three pesticides on non-target aquatic and soil organisms: commercial formulation versus active ingredient. Ecotoxicology, 18, 455-463. [ Links ]
Perobelli, J. E., Martinez, M. F., da Silva Franchi, C. A., Fernandez, C. D., de Camargo, J. L., & Kempinas, W. G. (2010). Decreased sperm motility in rats orally exposed to single or mixed pesticides. Journal of Toxicology and Environmental Health, 73, 991-1002. [ Links ]
Pierce, R. H., Henry, M. S., Blum, T. C., & Mueller, E. M. (2005). Aerial and tidal transport of mosquito control pesticides into the Florida Keys National Marine Sanctuary. Revista de Biología Tropical, 53, 117-125. [ Links ]
Pizzimenti, S., Ciamporcero, E., Daga, M., Pettazzoni, P., Arcaro, A., Cetrangolo, G., Minelli, R., Dianzani, C., Lepore, A., Gentile, F., & Barrera, G. (2013). Interaction of aldehydes derived from lipid peroxidation and membrane proteins. Frontiers in Physiology, 4, 242. [ Links ]
Qiao, D., Seidler, F. J., & Slotkin, T. A. (2005). Oxidative mechanisms contributing to the developmental neu-rotoxicity of nicotine and chlorpyrifos. Toxicology and Applied Pharmacoogy, 206, 17-26. [ Links ]
Rahman, M. F., Mahboob, M., Danadevi, K., Saleha, B. B., & Groover, P. (2002). Assessment of genotoxic effects of chorpyriphos and acephate by the comet assay in mice leucocytes. Mutation Research, 516, 139-147. [ Links ]
Satyanarayan, S., Ramakant, R., & Satyanarayan, A. (2005). Bioaccumulation studies of organochlorinated pesticides in tissues of Ciprinus carpio. Journal of Environmental Science and Health. Part B. Pesticides, Food contaminants, and Agricultural Wastes, 40, 397-412. [ Links ]
Sava, V., Velasquez, A., Song, S., & Sanchez-Ramos, J. (2007). Dieldrin elicits a widespread DNA repair and antioxidative response in mouse brain. Journal of Biochemical and Molecular Toxicology, 21, 125-135. [ Links ]
Scheiler, J. J., & Peterson, R. K. (2010). Toxicity and risk of permethrin and naled to non-target insects after adult mosquito management. Ecotoxicology, 19, 1140-1146. [ Links ]
Slotkin, T. A., & Seidler, F. J. (2010). Oxidative stress from diverse developmental neurotoxicants: antioxidants protect against lipid peroxidation without preventing cell loss. Neurotoxicology and Teratology, 32, 124-131. [ Links ]
Swaen, G. M., de Jong, G., Slangen, J. J., & van Amelsvoort, L. G. (2002). Cancer mortality in workers exposed to dieldrin and aldrin: an update. Toxicology and Industrial Health, 18, 63-70. [ Links ]
Takabe, Y., Tsuno, H., Nishimura, F., Tanji, N., Maruno, H., Tsurukawa, M., Suzuki, M., & Matsumura, C. (2012). Bioaccumulation and primary risk assessment of persistent organic pollutants with various bivalves. Water Science and Technology, 66, 2620-2629. [ Links ]
Tice, R. R., Agurell, E., Anderson, D., Burlinson, B., Hartmann, A., Kobayashi, H., Miyamae, Y., Rojas, E., Ryu, J. C., & Sasaki, Y. F. (2000). Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environmental and Molecular Mutagenesis, 35, 206-221. [ Links ]
Tsai, W. T. (2010). Current status and regulatory aspects of pesticides considered to be persistent organic pollutants (POPs) in Taiwan. International Journal of Environmental Research and Public Health, 7, 3615-3627. [ Links ]
Varó, I., Serrano, R., Pitarch, E., Amat, F., López, F. J., & Navarro, J. C. (2002). Bioaccumulation of chlorpyrifos through an experimental food chain: study of protein HSP70 as biomarker of sublethal stress in fish. Archives of Environmental Contamination and Toxicology, 42, 229-235. [ Links ]
Vasquez, M. I., & Fatta-Kassinos, D. (2013). Is the evaluation of "traditional" physicochemical parameters sufficient to explain the potential toxicity of the treated wastewater at sewage treatment plants? Environmental Science and Pollutution Research International, 20, 3516-3528. [ Links ]
Vasquez, M. Z. (2010). Combining the in vivo comet and micronucleus assays: a practical approach to genotoxicity testing and data interpretation. Mutagenesis, 25, 187-199. [ Links ]
Verma, R. S., & Srivastava, N. (2003). Effect of chlorpyrifos on thiobarbituric acid reactive substances, scavenging enzymes and glutathione in rat tissues. Indian Journal of Biochemical and Biophysics, 40, 423-428. [ Links ]
Voulgaridou, G. P., Anestopoulos, I., Franco, R., Panayiotidis, M. I., & Pappa, A. (2011). DNA damage induced by endogenous aldehydes: current state of knowledge. Mutation Research, 711, 13-27. [ Links ]
Yin, X., Zhu, G., Li, X. B., & Liu, S. (2009). Genotoxicity evaluation of chlorpyrifos to amphibian Chinese toad (Amphibian: Anura) by comet assay and micronucleus test. Mutation Research, 680, 2-6. [ Links ]
1 Laboratorio de Genética, Escuela Nacional de Ciencias Biológicas IPN, Av. Wilfredo Massieu, Col. Lindavista. México DF, CP 07738
Laboratorio de Genética, Facultad de Estudios Superiores Cuautitlán UNAM, Av. 1° de Mayo, Col. Santa María las Torres Cuautitlán Izcalli, Estado de México, CP 54740; sadibar@gmail.com
2 Laboratorio de Toxicología Acuática, Escuela Nacional de Ciencias Biológicas IPN, Av. Wilfredo Massieu, Col. Lindavista. México DF, CP 07738; genetica.ipn@gmail.com
3 Laboratorio de Genética, Escuela Nacional de Ciencias Biológicas IPN, Av. Wilfredo Massieu, Col. Lindavista. México DF, CP 07738; isela.alvarez@gmail.com
4 Laboratorio de Bioconservación y Manejo, Escuela Nacional de Ciencias Biológicas IPN, Prol. de Carpio y Plan de Ayala, Col. Santo Tomás, México DF, CP 11340; eulopez@ipn.mx
5 Laboratorio de Genética, Escuela Nacional de Ciencias Biológicas IPN, Av. Wilfredo Massieu, Col. Lindavista. México DF, CP 07738; eduardo.madrigal@lycos.com
Received 03-III-2014. Corrected 01-IX-2014. Accepted 02-X-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}