










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.3 San José Sep. 2014
Fish assemblages on fringing reefs in the southern Caribbean: biodiversity, biomass and feeding types
Ensambles de peces en los arrecifes de franja en el Caribe sur: biodiversidad, biomada y tipos de alimentación
Ensambles de peces en los arrecifes de franja en el Caribe sur: biodiversidad, biomada y tipos de alimentación
Jahson B. Alemu I.1*
Abstract
Reef fish assemblages in the Caribbean are under increasing pressure from human activities. Inadequate enforcement of legislation coupled with unreliable and data-poor landings in Tobago have led to the unregulated exploitation of reef fish for decades. This study addresses the lack of data on major reefs. Visual observations of fish fauna were conducted from November 2011-May 2013 at open access reef sites (Speyside, Charlotteville, Culloden, Arnos Vale, Mt. Irvine, La Guira, Kilgwyn, Plymouth and Black Rock) and one protected area (Buccoo Reef Marine Park). Belt transects surveys were used to determine fish density, species diversity and abundance at the 10-15m depth contour. Fish sizes were converted to biomass using the length-weight relationship of fish W=aLb. Most fish assemblages were dominated by small herbivores (<15cm), in particular Pomacentridae and Scaridae. Few large predators (>40cm) e.g. Serranidae, were noted, which is indicative of fishing pressure. MDS ordination identified three fish assemblages: i) northeastern, ii) southwestern and iii) intermediate. The northwestern cluster (Speyside and Charlotteville) were most representative of reef fish assemblages across the entire island, and exhibited the highest species richness, diversity and biomass. However, the southwestern cluster the highest numerical abundance. The marine protected area contained higher fish biomass, abundance, diversity and richness, but it was only representative of reef fish assemblages on the southwest of the island and not the entire Tobago. Research on the reef fishery, particularly spear fishing, is recommended to determine impact.
Key words: coral reef fish, Tobago, trophism, underwater visual surveys.
Resumen
Los ensamblajes de peces de arrecife en el Caribe están bajo una creciente presión de las actividades humanas. La ejecución inadecuada de la legislación, junto con los desembarques poco fiables y con escasez de datos en Tobago han llevado a una explotación sin reglas durante décadas. Este estudio aborda la falta de datos sobre las comunidades de peces de arrecife en los principales arrecifes. Las observaciones visuales se llevaron a cabo desde noviembre 2011 hasta mayo 2013 en sitios de acceso a los arrecifes abiertos (Speyside, Charlotteville, Culloden, Arnos Vale, el monte Irvine, La Guira, Kilgwyn, Plymouth y Negro Rock) y un área protegida (El parque Buccoo Marine Reef). Se utilizaron encuestas para determinar la densidad de peces, la diversidad y abundancia de especies a 10-15m de profundidad del contorno. El tamaño de los peces se convirtió a biomasa utilizando la relación talla- peso de los peces W=aLb. La mayoría de las comunidades de peces son dominadas por pequeños herbívoros (<15cm), en particular Pomacentridae y Scaridae. Pocos depredadores grandes (>40cm), por ejemplo Serranidae, se observó, lo que es indicativo de la presión de pesca. El análisis de escala multidimensional identificó tres conjuntos distintos de peces indicados como i) del noreste, ii) al suroeste y iii) grupos intermedios. El grupo del noroeste (Speyside y Charlotteville ) es más representativo de comunidades de peces de arrecife en toda la isla, y mostró la mayor riqueza de especies, diversidad y biomasa. Sin embargo, se observó la abundancia más alta de peces en el grupo del suroeste. El área marina protegida contenia mayor biomasa, abundancia, diversidad y riqueza de peces, pero era el único representante de las asociaciones de peces arrecifales en el suroeste de la isla y no de todo Tobago. Se recomienda determinar los impactos de la pesca en los arrecifes, particularmente con el uso de arpón.
Palabras clave: trofismo, Tobago, peces de arrecife de coral, encuestas submarinas visuales.
Coral reef fishes are the most diverse vertebrate communities on Earth (Jones, 1991) and their spatio-temporal distribution is influenced by a combination of complex biological and physical factors. These factors include larval supply (Doherty, 1991), competition (Munday, Jones & Caley, 2001; Holbrook & Schmit, 2002), wave exposure (Fulton, Bellwood & Wainwright, 2005), depth (Srinivasan, 2003) and habitat complexity (Friedlander, Sandin, DeMartini & Sala, 2010). Reef fishes, as predators or herbivores, play an important role in the community dynamics of coral reefs through their interactions with corals, algae, other herbivores and other predators. Disruption in the balance of reef fish assemblages can decrease coral cover and increase algal abundances (Roberts, 1995). Because of this importance, fishes are often a focus of monitoring and management programmes to evaluate the condition of reef communities (Green, Bellwood & Choat, 2009). Historically coral reef fish have been difficult to manage, in part, because different species often have different habitat requirements (Sale, 2002), life history patterns (Appeldoorn, Recksiek, Hill, Pagan & Dennis, 2003) and feeding regimes (Holland, Peterson, Lowe & Wetherbee, 1993).
Reef fish communities around Tobago exist on the margin of coral reefs in the southern Caribbean, and play a critical role in maintaining reef integrity. However, declining reef fish (abundance and biomass) as told in the oral history of Tobago, threatens the mainstay of the local diet and dependent economies. Further, the recent invasion of the lionfish onto Tobago’s reefs poses a real threat to juvenile and small reef fish communities. The purpose of this study is to a) establish some baseline data on the abundance and species richness of reef fishes around Tobago and b) assess any spatial variation in these assemblages.
Materials and Methods
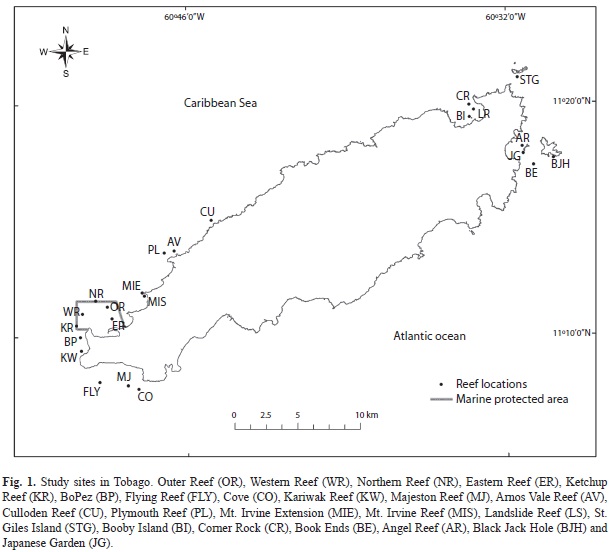
Study site: This study was conducted over an eighteen month period October 2011 to March 2013 on the reefs surrounding Tobago (10°2’ to 11°12’N and 60°30’ to 61°56’W) (Fig. 1), located 7km east off the Paria Peninsula. Study reefs (n=23) were of comparable size(~4ha) and represented the variability of the major reef systems. Tobago’s underwater topography is characterized by two distinct topographic regimes, a shallow limestone gently sloping shelf on the southwest side of the island, and a narrow, rocky, steeply sloping shelf on the northeastern side, upon which a variety of coral have been established (Snake, Rowe, Yule & Wadge, 1986). In this study, the southwest reefs are considered true coral reef (CR) formations consisting of corals colonies built upon a carbonate substrate with distinct reef zonation. Whereas the northeast reefs are considered coral associations (CA) consisting of coral-sponge co-dominated reefs established on rocky non-carbonate based substrata. Reefs along the northeast of Tobago are more exposed to the northeast trade winds and occur in a high energy environment due to the convergence of the Atlantic Ocean and Caribbean Sea. All study reefs were dominated by coral taxa such as Montastraea faveolata, Siderastrea siderea, Diploria strigosa and Colpophyllia natans.
Reef fish census: Reef fish surveys were conducted to estimate abundance, richness and to determine spatial distribution. Surveys were conducted along three 25x5m belttransects along the outer reef slope between 10-15m depth. Along each transect the number of individuals of each species was estimated and the total length of each fish was visually estimated and classified into 10cm classes (<10cm, 10–20cm, >20–30cm, >30–40cm and >40cm). Blennids and gobiids were excluded for this assessment, due to their cryptic nature, difficulty in field identification and field time constraints. Shannon-Weaver diversity indices (H’) [H’= - ∑i pi (log pi)], species richness (S), fish density/100m2 (D) and species abundance (N), were determined for all reefs.
Fish biomass was estimated using the length-weight relationship of fish, W = aLb. The most suitable “a” and “b” constants were acquired from Froese and Pauly (2013), and where no relationship was available for species, that of a closely related species as applied. Trophic guilds were classified according to Froese and Pauly (2013), however in many cases there was no definitive consensus as to the trophism of several species. As a result trophic groups were consolidated and presented as planktivores, primary consumers (herbivores and detritivores), secondary consumer (omnivores and benthivores) and piscivores.
Benthic cover and vertical relief was also measured along benthic transects. Benthic cover was estimated using six 10 x 1m belt transects at each site as described by (Hill & Wilkinson, 2004). Ten non-overlapping 1m2 photos were taken along each transect and analysed using the Coral Point Count with Excel (CPCe) programme (Kohler & Gill, 2006). Sixty random points were overlaid onto each photoquadrat, and the benthos under each point was identified to the lowest taxonomic level, for a total of 3 600 points per site. Only hard coral cover is presented. Vertical relief was measured as the distance between the lowest point and highest point on the substrate along that transect (Luckhurst & Luckhurst, 1978). Low relief was considered to be areas of between 0–1.5m and high relief to be >1.5m.
Bray-Curtis similarity on fourth-root transformed abundance and biomass was calculated, and analysis of similarities (ANOSIM) was conducted to test for differences in similarity between reefs and reef types. The ANOSIM results in a global R statistic that reflects the differences in variability between groups and within groups, and checks for significance of R were performed using permutation tests (Clarke & Warwick, 2001). Two dimensional ordinations were created using non-metric multi-dimensional scaling (MDS), which was used to generate plots in which the distance between points is proportional to their degree of dissimilarity, so closer points are more similar than points farther away (Clarke & Warwick, 2001). MDS plots were generated using entire assemblage level parameters, mean abundances and species biomass. A low stress value is an indicator of low error, similar to a measure of standard deviation (Clarke & Warwick, 2001). The species predominantly responsible for discriminating between assemblages were identified by the similarity percentage (SIMPER).
Results
A total of 15 576 fish, representing 96 species and 23 families were recorded around Tobago, with an average of 677.2±376.3 individuals/site. The most abundant species were Stegastes partitus, Chromis miltilineata and Clepticus parrae which collectively represented approximately 45.0% of the total number of fish observed. The least abundant species were Balistes vetula, Mycteroperca bonaci, Bothus lunatus, Scarus coelestinus, Lutjanus cyanopterus, Lactophyrs quadricornis, M. tigris, Sphyraena barracuda and Abudefdub taurus, with each being only observed once.
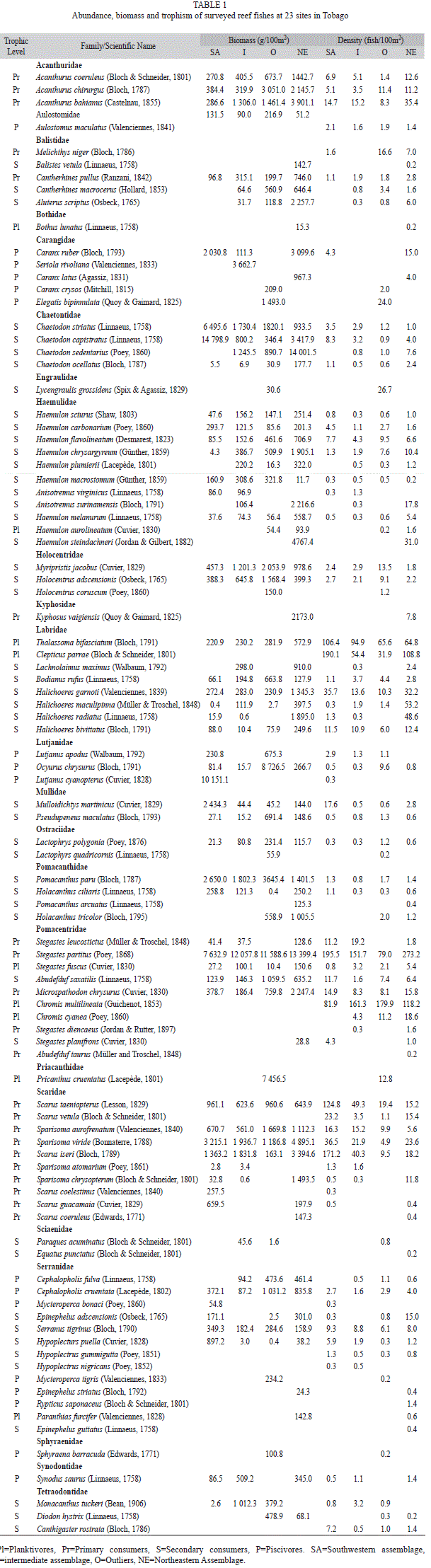
Over 80.0% of the fish observed belonged to the families Pomacentridae (39.2%), Labridae (25.6%) and Scaridae (15.4%) (Table 1). Species from the families Pomacentridae, Labridae, Scaridae, Haemulidae, Acanthuridae and Serranidae were found at all locations, and together with Pomacentridae accounted for 90% of fish observed. Chaetontidae was observed in all but one site. Together, these seven families accounted for ~70.0% of the total biomass, with the highest biomass (21.0%) provided by Pomacentridae (56kg/100m2). Other major contributors to overall biomass included Scaridae (11.4%), Lutjanidae (8.2%), Acanthuridae (6.4%) and Haemulidae (6.2%), with all other noted families contributing <5% each to overall biomass (Table 2).
The most abundant trophic group was the primary consumer (herbivores and detritivores) which accounted for 43.2% of all fish recorded, followed by planktivores (36.5%), secondary consumers (17.8%) and piscivores (2.5%). The primary consumer, Stegastes partitus contributed the most towards total biomass across sites, 19.2% of the total fish biomass. Other major contributors to the overall fish biomass included Chromis multilineata (14.8%), Clepticus parrae (10.6%) and Thalassoma bifasciatum (9.1%). All other species each contributed <7% to overall fish biomass.
Fish assemblages at all locations were dominated by fish ≤10cm in size, probably as a result of the numerical abundance of the inherently small-sized pomacentrids. Fish ≤10cm in size accounted for 71.1% of all fish recorded. The 11-20 and 20-30cm size classes held 22.6% and 4.5% respectively.Very few fish were larger than 30cm. Those recorded larger than 30cm included Sparisoma viridae, Pomacanthus arcuatus, Scarus vetula and Hypoplectrus nigricans.
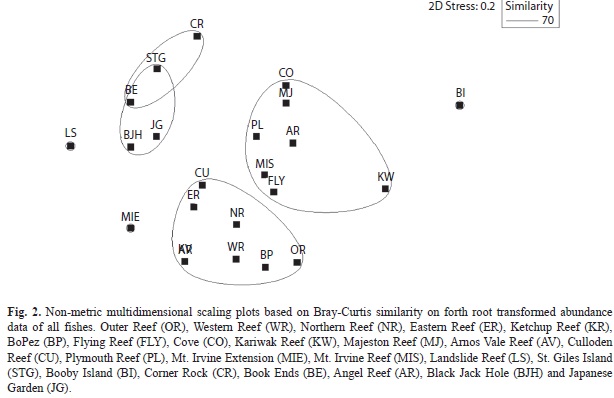
MDS ordination identified three main assemblages of fish based on reef type (ANOSIM, R=0.281, P<0.001), with three outlier groups at LS, BI and MIE based on the numerical abundance of fish (Fig. 2). These assemblages were well separated from the other reefs in ordination space (Stress=0.2). Sites grouped into a northeast Tobago high relief assemblage (CR, STG, JG, BJH and BE),a southwest Tobago low relief assemblage (OR, BP, WR, NR, ER, CU, AV and KR) and an intermediate relief low coral cover assemblage (KW, AR, FLY, MIS, PL, MJ and CO) (Fig 2).Species richness and diversity were generally higher at the northeastern Tobago reefs, particularly at BI (S=38, H’=2.989), STG (S=46, H’=2.95) and BJH (S=45, H’=2.72). The overall highest diversity was occurred at AR (3.003) in the southwest and the lowest H’ occurred at CO (H’=1.234) in the intermediate cluster (Table 2).
Similarly, there was also a significant difference in family composition with reef type (ANOSIM, R=0.263, P<0.002). The northeast assemblage accounted for 55.9% of the total fish biomass and 42.4% of the total fish abundance and the southwest assemblage accounted for 32.5% of total biomass and 36.2% of total abundance. The special interest area, Buccoo Reef Marine Park (BRMP) within the southwestern cluster accounted for 23% of total fish abundance but only 8.8% biomass. This separation in assemblages underlinesthe differences in geomorphologic characteristics on either side of the island, high relief rock-based coral reefs occurring in northeast Tobago and low relief limestone-based coral reefs occurring in southwest Tobago.
With regards to fish biomass, while the general separation of clusters between northeastern and southwestern reefs was maintained in ordination space, there was no significantly relationship with reef type (ANOSIM, R=0.036, P=0.226). In fact the low R values indicate the three groups were not well separated. Non parametric MDS ordination plots based on size class and trophism did not provide distinct fish assemblages, even when reef type was considered.
Overall, the variation in abundance of Acanthuridae, Serranidae, Scaridae, Pomacentridae and Labridae was responsible for most of the dissimilarity among sites and assemblages. In particular, 16 species were responsible for most of the dissimilarity between northeastern, southwestern and intermediate assemblages (SIMPER, average dissimilarity=52.6%). Acanthuridae (particularly Acanthurus bahianus),Serranidae (particularly Serranus tigrinus and Epinephelus adscensionis), Pomacentridae (particularly Chromis multilineata, Stegastes partitus, Microspathodon chrysurus) and Balistidae (particularly Melichthys niger) were mostly found on the northeastern reef cluster, whereas Scaridae (particularlyScarus iseri, S. taeniopteus, Sparisoma viride, S. aurofrenatus) and Labridae (particularly Thalassoma bifasciatum, Clepticus parrae) were dominant in the southern and intermediate reef clusters.
Most top predators/piscivores (primarily Serranidae, Carangidae and Lutjanidae) were noted in the southwesterncohort of fish, which accounted for 45.8% of the total piscivores biomass. It should be noted that ~29.0% of piscivores biomass was detected at a single site in the southwest (BP) which were mainly represented by Ocyurus chrysurus and Lutjanus cyanopterus. The only site where no piscivores were observed was MJ. LS, the northeastern and intermediate assemblages accounted for 24.9%, 19.3% and 5.7% of total piscivores biomass respectively. Sixty five percent of all planktivores were recorded at LS which represented 38.8% of planktivore biomass. These were dominated by small bodied fish such as Chromis multilineata and Thalassoma bifasciatum. Primary consumers were most dominant within the northeast cluster and secondary consumers were most dominant within the southwest cluster (Table 1).
Few fish >40cm were recorded, and where they were noted densities were quite low. The highest densities of large fish occurred at AV (3.5 individuals/100m2), ER (2.7 individuals/100m2) and BP (1.6 individuals/100m2). Overall most fish (93.7%) were <20cm in size. Fish <10cm were similarly distributed among reefs, with the highest densities occurring at LS (7.4 individuals/100m2) in the northeast and KR (7.3 individuals/100m2) in the southwest. The highest densities of fish 11–20cm, 21–30cm and 31–40cm were all noted in the northeastern reefs. Medium sized fish (20-30cm) within southwest cluster and intermediate cluster consisted mainly of Scarus spp, Sparisoma spp, Pomacanthus paru, Mulloidichtys martinicus, Acanthurus spp and Stegastes spp. Whereas, the northeast assemblage displayed a wider range of medium sized such as Thalassoma spp, Halichores spp, Chromis spp, Clepticus sp., Haemulon spp, Mycteroperca tigris, Cephalopholis cruentata, Cephalopholis fulva, Ocyurus chrysurus, Aulostomus maculates and s Scarus spp.
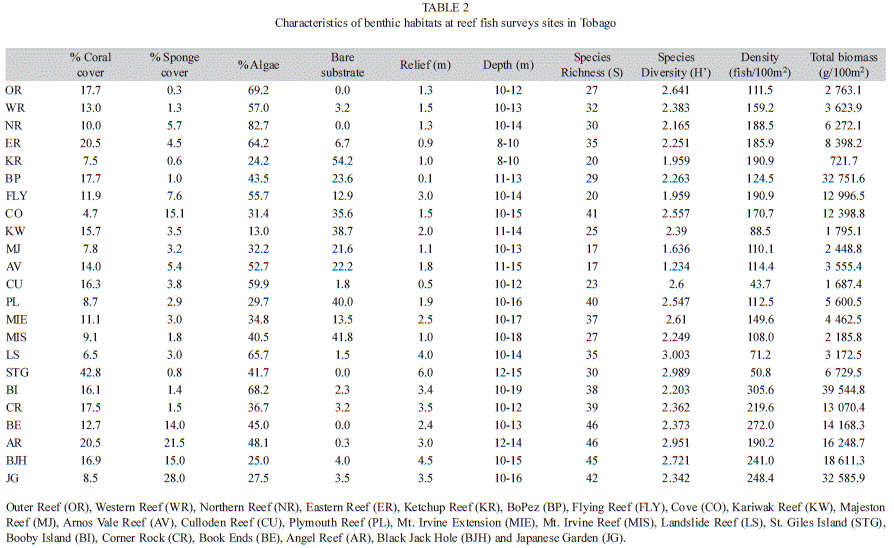
Within the 23 reefs, live coral cover ranged from 4.6% at CO to 42.8% at STG. Macroalgae (including erect coralline algae, turf algae and fleshy macroalgae) was the dominant feature on most reefs, with only four reefs (KW, KR, BJH and JG) having <30% algal cover. Sponge cover ranged from 0.3%28.0%, and together with hard corals was responsible for much of the habitat complexity observed on the reef. The northeastern and southwestern clusters of reefs showed similar coral cover, whereas the lowest coral cover was noted among the intermediate reef cluster. Sponge cover was highest five reefs (CO, BE, AR, BJH and JG), mainly within the northwestern reef cluster, but generally most sites exhibited <6% sponge. Thirteen to eighty three percent (13–82.6%) of reefs were bare substrate (crustose coralline algae, dead coral, rubble and sand) (Table 2). Overall, the northeastern reefs were mostly high relief reefs, with STG exhibiting the highest relief. Mean relief among the southwestern and intermediate reefs was 1.3±1.2m and 1.9±0.7m.
Discussion
This study examined variation in reef fish community structure (diversity, trophism, biomass and richness) across major reef systems in Tobago with the greatest differences occurring between reefs located in the northeastern and southwestern side of the island. The results identify distinct reef fish assemblages, associated with differences in reef type and morphology. Biomass, abundance, diversity and richness of reef fish, varied widely among reefs from northeast to southwest, as a result of shifts in dominance of functional groups and among reefs such as scarids. Overall, fish densities were between 44-306 individuals/100m2 and were dominated by small-bodied fishes of low commercial value (such as pomacentrids and labrids) and few highly valuable commercial species such as serranids and lutjanids. This is similar to other reefs in the Caribbean that have been overfished or highly impacted (Hughes, 1994; Mallela, Roberts, Harrod & Goldspink, 2007; Aguilar-Perera & Appledoorn, 2008). Reef fish biomass in Tobago was low compared to other islands in the region. Newman, Gustavo, Sala & Jackson, (2006) estimated fish biomass in a range of coral reef habitats in the Caribbean and found average biomass estimates to range between 15 and 60g/m2, whereas in Tobago mean biomass 10.7±11.0g/m2.
MDS ordination separated fish populations in three distinct assemblages with three outlier groups. Mean fish density, species richness, and diversity were higher at the northeastern rocky reefs relative to coralline reefs found on the intermediate and southwestern reef cluster. Some taxa exhibited densities several fold higher in northeastern reefs (e.g. pomacentrids and haemulids), than on the intermediate or southwestern reefs and vice versa (e.g. scarids). The differences in densities may be due to the northeastern sites offering greater structural complexity for exploitation by reef fish (Hixon & Beets, 1989; Caley & St. John, 1996; Friedlander, Brown, Jokiel, Smith & Rodgers, 2003; Graham, McClanahan, Letourneur & Galzin, 2007; Benfield, Baxter, Guzman & Mair, 2008; Friedlander et al., 2010). Additionally, the paucity of reef development in Tobago (Spalding, Ravilious & Green, 2001) would force principally reef-associated species exploit less-preferred habitats (e.g. rocky reefs) (Sandin, Vermeij & Hurlbert, 2008) around the island, such as those found on the northeastern reefs.
Structurally complex habitats fragment the area, resulting in numerous microhabitats being formed and encouraging heterogeneous fish assemblages (Gratwicke & Speight, 2005), and this appears to be the case in this study. The gently sloping coralline reefs in the southwest of the island contrast with the steeply sloping rocky reefs and overhangs that characterise the other reefs on the island. The intermediate cluster possesses both coralline reef and rocky reef features, but is also popular recreational fishing areas (spear, fishpot and handline) for coastal communities. We conjecture that the regular removal of fish from these sites and low coral cover are responsible for the paucity of fish. It should be noted, that not all reefs fish assemblages in the southwest or northeast clustered together into an overall southwest assemblage and northeast assemblage, possibly as a result of the differences in the biological and physical character of neighbouring sites (Curley, Kingsford & Gillander, 2002).
The differences in spatial variability of piscivores (Table 1) could be due to spatial differences in habitat availability and food availability. Generally, piscivore density was highest in the northeastern reefs, although most of the piscivore biomass was found at BP in the southwestern cluster. It is plausible that the higher biomass of piscivores such as serranids and lutjanids (Table 1) in the northeastern reefs could be a direct result of the food availability and an indirect result of habitat availability for prey items (Stewart & Jones, 2011). Serranids and lutjanids are roving predators that feed mostly on fish and are associated with coral reefs (Ferreira, Floeter, Gasparini, Ferreira & Joyeux, 2004). Beukers & Jones (1997) however, suggest that habitat availability is a stronger factor than food availability on influencing piscivore abundance on a coral reef. The effect of habitat complexity could either be direct, where it provides fish habitat, or indirect where it influences the distribution of the food source of these species.
Marine protected areas enclosing coral reefs have had a broad range of positive effects such as increased biomass, abundance, average size and diversity of fish and invertebrates (Russ & Alcala, 2011). In this study similar results were observed, supporting previous small scale studies on the impact of MPAs on increasing reef fish populations and biomass (Polunin & Roberts, 1993; Rakitin & Kramer, 1996). It should be noted that connectivity of the MPA to nearby mangrove and seagrass habitats which are suitable for some reef-associated fishes, might contribute to the higher abundance and biomass observed. In fact, the biomass of important herbivores within the MPA such as parrotfish (scarids: ~12.0g/100m2) was almost twice the scarid biomass of nearby reefs (~6.8g/100m2) (Table 1). When compared to other well protected MPA in the region such Los Roques (scarid biomass 60.7g/m2) and Flower Garden Banks (scarid biomass 35.8g/m2) (Posada, Villamizar & Alvarado, 2003), the MPA’s scarid biomass is quite low. On the non-protected fishes reefs of Curacao scarid biomass is approximately 15.2g/m2 and as low as 4.6 on the overfished Montego Bay reefs (J. Jackson et al., unpublished).
This study represents a comprehensive baseline assessment of the reef fish fauna of Tobago and is instructive as to where efforts should be prioritised to conserve reef fish populations from human threats (fishing) and alien invasive threats (Pterois sp.). Additionally, future research on the reef fishery, particularly spears fishing, to determine impacts by these activities on fish populations.
Acknowledgments
I would like to thank Stefan Lue Chin, Sherron Barker, Jonathan Gomez and Addison Titus who assisted in data collection and analysis. Special thanks to Rahanna Juman, Anika Gellineau, Dawn A.T. Phillip and Michelle Cazabon-Mannette for reviewing and commenting. This research was conducted as part of the Project for Ecosystem Services (www.proecoserv.org) while the author was a graduate student at the University of the West Indies. Financial support from the Global Environment Facility (GEF) and UNEP is gratefully acknowledged.
References
Aguilar-Perera, A., & Appeldoorn., R. S. (2008). Spatial distribution of marine fishes along a cross-shelf gradient containing a continuum of mangrove-seagrass-coral reefs off southwestern Puerto Rico. Estuarine, Coastal and Shelf Science, 76, 378-394. [ Links ]
Appeldoorn, R. S., Recksiek C. W., Hill R. L, Pagan F. E., & Dennis, G. D.(1997). Marine protected areas and reef fish movements: the role of habitat in controlling ontogenetic migration. Proceedings of the 8th International Coral Reef Symposium, 2, 1917-1922. [ Links ]
Benfield, S., Baxter, L., Guzman, H. M., & Mair, J. M. (2008). A comparison of coral reef and coral community fish assemblages in pacific Panama and environmental factors governing their structure. Journal of the Marine Biological Association of the United Kingdom, 88, 1331-1341. [ Links ]
Beukers, J. S., & Jones G. P. (1997). Habitat complexity modifies the impact of piscivores on a coral reef fish population. Oecologia, 114, 50-59. [ Links ]
Caley, M. J., & St John, J. (1996). Refuge availability structures assemblages of tropical reef fishes. Journal of Animal Ecology, 65, 414-428. [ Links ]
Clarke, K. R., & Warwick, R. M. (2001). Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd Edition. Plymouth: Primer-E Ltd. [ Links ]
Curley, B. G., Kingsford M. J., & Gillander, B. M. (2002). Spatial and habitat related patterns of temperate reef-fish assemblages: Implications for the design of Marine protected Areas. Marine and Freshwater Research, 53, 1197-1210. [ Links ]
Doherty, J. P. (1991). Spatial and temporal patterns in recruitment. pp. 261-293. In P.F. Sale (ed.). The ecology of fishes on coral reefs. San Diego: Academic Press. [ Links ]
Ferreira, C. E. L., Floeter S. R., Gasparini J. L., Ferreira B. P., & Joyeux J. C. (2004). Trophic structure patterns of Brazilian reef fishes: a latitudinal comparison. Journal of Biogeography, 31, 1093-1106. [ Links ]
Friedlander, A. M., Brown E. K., Jokiel P. L., Smith W. R., & Rodgers K. S. (2003). Effects of habitat, wave exposure, and marine protected area status on coral reef fish assemblages in the Hawaiian archipelago. Coral Reefs, 22, 291-305. [ Links ]
Friedlander, A. M., Sandin S. A., DeMartini E. E., & Sala E. (2010). Spatial patterns of the structure of reef fish assemblages at a pristine atoll in the central Pacific. Marine Ecology Progress Series, 410, 219-231. [ Links ]
Froese, R., & Pauly, D. (eds.). (2013). FishBase.World Wide Web electronic Retrieved from publication.www.fishbase.org, version. [ Links ]
Fulton, C. J., Bellwood, D. R., & Wainwright, P. C. (2005). Wave energy and swimming performance shape coral reef fish assemblages. Proceedings of the Royal Society, 272, 827-832. [ Links ]
Graham, N. A. J., McClanahan T. R., Letourneur, Y., & Galzin, R. (2007). Anthropogenic stressors, inter-specific competition and ENSO effects on a Mauritian coral reef. Environmental Biology of Fishes, 78, 57-69. [ Links ]
Gratwicke, B., & Speight M. R. (2005). Effects of habitat complexityon Caribbean marine fish assemblages. Marine Ecology Progress Series, 292, 301-310. [ Links ]
Green, A. L., Bellwood D. R., & Choat, H. (2009). Monitoring functional groups of herbivorous reef fishes as indicators of coral reef resilience. A practical guide for coral reef managers in the Asia Pacific Region. Gland, Switzerland: IUCN. [ Links ]
Hill, J., & Wilkinson. C. (2004). Methods for Ecological Monitoring of Coral Reefs, Version 1. Townsville, Australia: Australian Institute of Marine Science. [ Links ]
Hixon, M. A., & Beets, J. P. (1989). Predation, prey refuges, and the structure of coral-reef fish assemblages. Ecological Monographs, 63, 77-101. [ Links ]
Holbrook, S. J., & Schmitt, R. J. (2002). Competition for shelter spaces causes density dependent predation mortality in damselfishes. Ecology, 83, 2855-2868. [ Links ]
Holland, K. N., Peterson J. D., Lowe C. G., & Wetherbee B. M. (1993). Movements, distribution and growth rates of the white goatfish Mulloides flavolineatus in a fisheries conservation zone. The Bulletin of Marine Science, 52, 982-992. [ Links ]
Hughes, T. P. (1994). Catastrophes, phase-shifts, and large-scale degradation of a Caribbean coral reef. Science, 265, 1547-1551. [ Links ]
Jones G. P. (1991). Post recruitment process in the ecology of coral reef populations: a multifactorial perspective, pp 294-328. In Sale, P.F. (ed.). The ecology of fishes on coral reefs. San Diego: Academic Press. [ Links ]
Kohler K. E., & Gill S. M. (2006). Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Computers and Geosciences, 32, 1259-1269. [ Links ]
Luckhurst, B. E., & Luckhurst. K. (1978). Analysis of the influence of substrate variables on coral reef fish communities. Marine Biology, 49, 317-323. [ Links ]
Mallela, J., Roberts, C. S, Harrod, C., & Goldspink, C. R. (2007). Distributional patterns and community structure of Caribbean coral reef fishes within a river-impacted bay. Journal of Fish Biology, 70, 523-537. [ Links ]
Munday, P. L., Jones G. P., & Caley, M. J. (2001). Interspecific competition and coexistence in a guild of coral-dwelling fishes. Ecology, 82, 2177-2189. [ Links ]
Newman, M. J. H., Gustavo P. A, Sala, E., & Jackson J. B. C. (2006). Structure of Caribbean coral reef communities across a large gradient of fish biomass. Ecological Letters, 9, 1216-1227. [ Links ]
Polunin, N. V. C., & Roberts C. M. (1993). Greater biomass and value of target coral-reef fishes in two small Caribbean marine reserves. Marine Ecology Progress Series, 100, 167. [ Links ]
Posada, J. M., Villamizar E., & Alvarado D. (2003). Rapid assessment of coral reefs in the Archipiélago Los Roques Nacional Park, Venezuela (Part 2: Fishes). Atoll Research Bulletin, 496, 531-544. [ Links ]
Rakitin, A., &. Kramer D. L. (1996). Effect of a marine reserve on the distribution of coral reef fishes in Barbados. Marine Ecology Progress Series, 131, 97-113. [ Links ]
Roberts, C. M. (1995). Effects of fishing on the ecosystem structure of coral reefs. Conservation Biology, 9, 988-995. [ Links ]
Russ, G. R., & Alcala C. (2011). Enhanced biodiversity beyond marine reserve boundaries: The cup spillith over. Ecological Applications, 21, 241-250. [ Links ]
Sale P. F. (2002). The science we need to develop for more effective management. In Sale, P.F.(ed) Coral reef fishes: dynamics and diversity in a complex ecosystem. London: Academic Press. [ Links ]
Sandin, S. A., Vermeij M. J. A., & Hurlbert A. H. (2008). Island biogeography of Caribbean coral reef fish. Global Ecology and Biogeography, 17, 770-777. [ Links ]
Spalding, M. D., Ravilious C., & Green E. P. (2001). World atlas of coral reefs. University of California, Berkeley: UNEP World Conservation Monitoring Centre. [ Links ]
Snake, W., Rowe D. W., Yule J. D., & Wadge G. (1986). Geologic map of Tobago, West Indies. Retrieved from http://www.gstt.org/Geology/tobago%20map.htm. [ Links ]
Srinivasan, M. (2003). Depth distributions of coral reef fishes: the influence of microhabitat structure, settlement, and post-settlement processes. Oecologia, 137, 76-84. [ Links ]
Stewart, B. D., & Jones G. P. (2011). Association between the abundance of piscivorous fishes and their prey on coral reefs: implications for prey-fish mortality. Marine Biology, 138, 383-397. [ Links ]
Appeldoorn, R. S., Recksiek C. W., Hill R. L, Pagan F. E., & Dennis, G. D.(1997). Marine protected areas and reef fish movements: the role of habitat in controlling ontogenetic migration. Proceedings of the 8th International Coral Reef Symposium, 2, 1917-1922. [ Links ]
Benfield, S., Baxter, L., Guzman, H. M., & Mair, J. M. (2008). A comparison of coral reef and coral community fish assemblages in pacific Panama and environmental factors governing their structure. Journal of the Marine Biological Association of the United Kingdom, 88, 1331-1341. [ Links ]
Beukers, J. S., & Jones G. P. (1997). Habitat complexity modifies the impact of piscivores on a coral reef fish population. Oecologia, 114, 50-59. [ Links ]
Caley, M. J., & St John, J. (1996). Refuge availability structures assemblages of tropical reef fishes. Journal of Animal Ecology, 65, 414-428. [ Links ]
Clarke, K. R., & Warwick, R. M. (2001). Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd Edition. Plymouth: Primer-E Ltd. [ Links ]
Curley, B. G., Kingsford M. J., & Gillander, B. M. (2002). Spatial and habitat related patterns of temperate reef-fish assemblages: Implications for the design of Marine protected Areas. Marine and Freshwater Research, 53, 1197-1210. [ Links ]
Doherty, J. P. (1991). Spatial and temporal patterns in recruitment. pp. 261-293. In P.F. Sale (ed.). The ecology of fishes on coral reefs. San Diego: Academic Press. [ Links ]
Ferreira, C. E. L., Floeter S. R., Gasparini J. L., Ferreira B. P., & Joyeux J. C. (2004). Trophic structure patterns of Brazilian reef fishes: a latitudinal comparison. Journal of Biogeography, 31, 1093-1106. [ Links ]
Friedlander, A. M., Brown E. K., Jokiel P. L., Smith W. R., & Rodgers K. S. (2003). Effects of habitat, wave exposure, and marine protected area status on coral reef fish assemblages in the Hawaiian archipelago. Coral Reefs, 22, 291-305. [ Links ]
Friedlander, A. M., Sandin S. A., DeMartini E. E., & Sala E. (2010). Spatial patterns of the structure of reef fish assemblages at a pristine atoll in the central Pacific. Marine Ecology Progress Series, 410, 219-231. [ Links ]
Froese, R., & Pauly, D. (eds.). (2013). FishBase.World Wide Web electronic Retrieved from publication.www.fishbase.org, version. [ Links ]
Fulton, C. J., Bellwood, D. R., & Wainwright, P. C. (2005). Wave energy and swimming performance shape coral reef fish assemblages. Proceedings of the Royal Society, 272, 827-832. [ Links ]
Graham, N. A. J., McClanahan T. R., Letourneur, Y., & Galzin, R. (2007). Anthropogenic stressors, inter-specific competition and ENSO effects on a Mauritian coral reef. Environmental Biology of Fishes, 78, 57-69. [ Links ]
Gratwicke, B., & Speight M. R. (2005). Effects of habitat complexityon Caribbean marine fish assemblages. Marine Ecology Progress Series, 292, 301-310. [ Links ]
Green, A. L., Bellwood D. R., & Choat, H. (2009). Monitoring functional groups of herbivorous reef fishes as indicators of coral reef resilience. A practical guide for coral reef managers in the Asia Pacific Region. Gland, Switzerland: IUCN. [ Links ]
Hill, J., & Wilkinson. C. (2004). Methods for Ecological Monitoring of Coral Reefs, Version 1. Townsville, Australia: Australian Institute of Marine Science. [ Links ]
Hixon, M. A., & Beets, J. P. (1989). Predation, prey refuges, and the structure of coral-reef fish assemblages. Ecological Monographs, 63, 77-101. [ Links ]
Holbrook, S. J., & Schmitt, R. J. (2002). Competition for shelter spaces causes density dependent predation mortality in damselfishes. Ecology, 83, 2855-2868. [ Links ]
Holland, K. N., Peterson J. D., Lowe C. G., & Wetherbee B. M. (1993). Movements, distribution and growth rates of the white goatfish Mulloides flavolineatus in a fisheries conservation zone. The Bulletin of Marine Science, 52, 982-992. [ Links ]
Hughes, T. P. (1994). Catastrophes, phase-shifts, and large-scale degradation of a Caribbean coral reef. Science, 265, 1547-1551. [ Links ]
Jones G. P. (1991). Post recruitment process in the ecology of coral reef populations: a multifactorial perspective, pp 294-328. In Sale, P.F. (ed.). The ecology of fishes on coral reefs. San Diego: Academic Press. [ Links ]
Kohler K. E., & Gill S. M. (2006). Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Computers and Geosciences, 32, 1259-1269. [ Links ]
Luckhurst, B. E., & Luckhurst. K. (1978). Analysis of the influence of substrate variables on coral reef fish communities. Marine Biology, 49, 317-323. [ Links ]
Mallela, J., Roberts, C. S, Harrod, C., & Goldspink, C. R. (2007). Distributional patterns and community structure of Caribbean coral reef fishes within a river-impacted bay. Journal of Fish Biology, 70, 523-537. [ Links ]
Munday, P. L., Jones G. P., & Caley, M. J. (2001). Interspecific competition and coexistence in a guild of coral-dwelling fishes. Ecology, 82, 2177-2189. [ Links ]
Newman, M. J. H., Gustavo P. A, Sala, E., & Jackson J. B. C. (2006). Structure of Caribbean coral reef communities across a large gradient of fish biomass. Ecological Letters, 9, 1216-1227. [ Links ]
Polunin, N. V. C., & Roberts C. M. (1993). Greater biomass and value of target coral-reef fishes in two small Caribbean marine reserves. Marine Ecology Progress Series, 100, 167. [ Links ]
Posada, J. M., Villamizar E., & Alvarado D. (2003). Rapid assessment of coral reefs in the Archipiélago Los Roques Nacional Park, Venezuela (Part 2: Fishes). Atoll Research Bulletin, 496, 531-544. [ Links ]
Rakitin, A., &. Kramer D. L. (1996). Effect of a marine reserve on the distribution of coral reef fishes in Barbados. Marine Ecology Progress Series, 131, 97-113. [ Links ]
Roberts, C. M. (1995). Effects of fishing on the ecosystem structure of coral reefs. Conservation Biology, 9, 988-995. [ Links ]
Russ, G. R., & Alcala C. (2011). Enhanced biodiversity beyond marine reserve boundaries: The cup spillith over. Ecological Applications, 21, 241-250. [ Links ]
Sale P. F. (2002). The science we need to develop for more effective management. In Sale, P.F.(ed) Coral reef fishes: dynamics and diversity in a complex ecosystem. London: Academic Press. [ Links ]
Sandin, S. A., Vermeij M. J. A., & Hurlbert A. H. (2008). Island biogeography of Caribbean coral reef fish. Global Ecology and Biogeography, 17, 770-777. [ Links ]
Spalding, M. D., Ravilious C., & Green E. P. (2001). World atlas of coral reefs. University of California, Berkeley: UNEP World Conservation Monitoring Centre. [ Links ]
Snake, W., Rowe D. W., Yule J. D., & Wadge G. (1986). Geologic map of Tobago, West Indies. Retrieved from http://www.gstt.org/Geology/tobago%20map.htm. [ Links ]
Srinivasan, M. (2003). Depth distributions of coral reef fishes: the influence of microhabitat structure, settlement, and post-settlement processes. Oecologia, 137, 76-84. [ Links ]
Stewart, B. D., & Jones G. P. (2011). Association between the abundance of piscivorous fishes and their prey on coral reefs: implications for prey-fish mortality. Marine Biology, 138, 383-397. [ Links ]
1. Institute of Marine Affairs, Hilltop Lane, Chaguaramas, Trinidad and Tobago, W.I., Tel: +1 868 678 3718, Fax: +1 868 634-4433; jalemu@ima.gov.tt
Received 22-VIII-2013 Corrected 18-II-2014 Accepted 24-III-2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}