










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.3 San José Sep. 2014
Lionfish abundance, size structure and spatial distribution along the Venezuelan coast (Pterois volitans, Pteroinae: Scorpaenidae)
Abundancia, estructura de tamaños y distribución espacial a lo largo de la costa venezolana del pez león (Pterois volitans, Pteroinae: Scorpaenidae)
Abundancia, estructura de tamaños y distribución espacial a lo largo de la costa venezolana del pez león (Pterois volitans, Pteroinae: Scorpaenidae)
Abstract
The recent invasion of lionfish (Pterois volitans) in the Atlantic is considered a new threat to benthic and fish communities in the Caribbean region. This species was first reported in Venezuela in 2009 at various sites. Increasing reports in the past five years suggest lionfish has expanded its range of distribution and habitats. Nevertheless, this information is mostly anecdotal and extensive surveys aimed to determine its abundance, size structure and other ecological aspects encompassing wider spatial scales are necessary to understand the actual role of this species on sub-tidal marine communities in Venezuela. We determined its density and population size structure through visual census along the Venezuelan coast. Visual censuses were made following strip transects at a depth between 5 and 20m and in daylight time, at 19 sites in five localities. Average density ranged between 7 to 55 individuals per hectare among sites. Most individuals were adults and most were found in caves, coexisting with other lionfish or with different species, while others were actively preying. The fish Pterois volitans seems to be well-established along the Venezuelan coast in densities that in some sites appear to be higher than in their Pacific native range but lower than in some invaded localities of the Atlantic.
Key words: Exotic species, visual census, South Caribbean, Pterois volitans.
Resumen
La reciente invasión del pez león (Pterois volitans) en el Atlántico se considera una nueva amenaza para bentónicos y peces de las comunidades en la región del Caribe. Esta especie fue informada por primera vez en varios sitios de Venezuela en 2009. El aumento de informes en los últimos cinco años sugiere que ha ampliado su distribución y hábitats. Sin embargo, esta información es principalmente anecdótica por lo que son necesarios estudios dirigidos a determinar su abundancia, estructura de tallas y otros aspectos ecológicos que abarquen más escalas espaciales para entender el papel real de esta especie en comunidades marinas en Venezuela. Determinamos la densidad y estructura del tamaño poblacional del pez león a través de censos visuales en la costa venezolana. Realizamos censos visuales siguiendo transectos lineales a una profundidad entre 5 y 20m durante el día, en 19 sitios de cinco localidades. La densidad media varia entre 7 y 55 individuos por hectárea entre sitios. La mayoría de los individuos eran adultos que se encontraban en cuevas, coexistiendo con otro peces león o con diferentes especies; mientras que otros estaban forrajeando activamente. El pez Pterois volitans esta bien establecido a lo largo de la costa venezolana y sus densidades en algunos sitios parecen exceder lo informado para su distribución nativa en el Pacífico pero son densidades menores que las informadas en localidades del Atlántico.
Palabras clave: Las especies exóticas, censos visuales, volitans del Sur del Caribe, Pterois
The introduction of exotic species is considered a major threat to biodiversity because it has produced significant phase shifts in marine ecosystems worldwide (Gurevitch & Padilla, 2004; Ruiz, Carlton, Grosholz & Hines, 1997). Among exotic marine species, Pterois volitans, commonly known as lionfish, has recently received special attention. This species is native and widespread in the Indo-pacific but was introduced to the Atlantic probably through numerous releases of ornamental fish since 1985 in Florida (Schofield, 2009). Since then P. volitans has invaded from Florida to North Carolina, most of the Caribbean and the Gulf of Mexico (Schofield, 2010). Lionfish was first reported in Venezuelan in 2009, in Cape Bay in the Central Coast of Venezuela and in Morrocoy National Park. After 2010, reports increased in frequency, and it is presumed that P. volitans moved from west to east along the coast, according to a database created for divers, fishermen and scientists to report sightings the species (http://pezleon.cbm.usb.ve). Currently, P. volitans seems to be well established all along the Venezuelan coast (Lasso-Alcalá & Posada, 2010).
Pterois volitans is a voracious predator feeding from a wide range of fishes and crustaceans (Albins & Hixon, 2008). It has been suggested that the presence of this species in the Atlantic may have negative effects on reef fish communities such as significant reduction in recruitment rates (Albins & Hixon, 2008; 2011). It is also believed that the introduction of lionfish might have indirect effects on the structure of benthic coral communities (Lesser & Slattery, 2011). Therefore it is necessary first to determine its spatial patterns of distribution. This information will be valuable to help identify the processes that might explain these patterns, and more importantly the spatial scales that are relevant to the design of proper experiments.
We present a preliminary survey of abundance and distribution of lionfish encompassing different sites along the western, central and eastern coast of Venezuela aimed at determining the abundance, size structure and patterns of spatial distribution of lionfish. For this, localities separated by hundreds of kilometers and sites within localities separated by hundreds of meters were sampled along the coast of Venezuela.
Materials and Methods
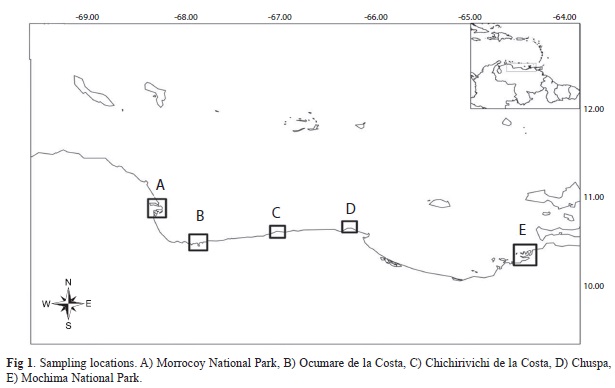
Study sites: The study was conducted at five localities and two to five sites within each locality: (1) Morrocoy National Park (five sites: Caimán, Mero, Boca Seca, Cayo Sombrero and Cayo Norte), (2) Ocumare de la Costa (five sites: Guabinas, Canal de la Ciénaga, Punta de la Ciénaga, Valle Seco and Chuao), (3) Chichirivichi de la Costa (two sites: Punta de Chichirivichi and Bahía de Chichirivichi), (4) Chuspa (two sites: Punta Capino and Quigua), and (5) Mochima National Park (five sites: Punta de Cruces, Tigrillo, Piedra de la Ahogada, Playa Colorada and Burbujas).
Morrocoy national park is located at the western coast of Venezuela (10°51’60”N, 68°14’10”W) it is a marine protected area with major tropical marine communities (seagrass beds, mangrove forest, sandy and rocky shores and coral reefs) occurring in a total area of 320km2 (López-Ordaz, Ortaz & Rodriguez-Quintal, 2009). Ocumare de la Costa (10°29’20”N, 67°49’21”W) Chichiriviche de la Costa (10°33’15”N, 67°14’20.38”W) and Chuspa (10°38’25”N, 66°17’56”W) are located in the Central Coast of Venezuela. This region in characterized by narrow continental margins and mostly rocky shores. Even thought it has some coral reef developments especially in La Cienaga and Guabinas. Mochima is also a National Park located at the eastern coast of Venezuela (10°23’44”N, 64°22’2”W) dominated by scattered coral communities growing on rocky bottoms, these communities being seasonally exposed to coastal upwelling and is considered the most important coral communities in the eastern of Venezuela (Ruiz et al., 1997).
Lionfish surveys: Visual censuses were made to estimate abundance of lionfish. In each site 7 to 12 belt transects (4m wide and 30m long, 120m2) were surveyed, according to reef´s size and at depth varying from 20m to 5m. This was made according to the depth in each reef which was a lot deeper in Ocumare and Chichirichi de la Costa. All visual censuses were made by a single diver with special care taken to thoroughly search every cave, crevice and holes inside each transect. The number of lionfish and the size structure were determined using five size categories (0-10cm, 11-20cm, 21-30cm, 31-40cm, more than 40cm). The criteria for selecting these size classes were based following the AGRRA protocol (Ginsburg, R., Kramer, P., Lang, J., Salle, P., & Steneck, R., 1998). Adults and juveniles were identified through determination of total length, considering fish under 10cm as juvenile. To estimate lionfish biomass we used the allometric relationship W=aLB to convert size to weight, where L=length, a=0.00497 and b=3.201 (Darling, Green, O’Leary & Côté, 2011).
Statistical analysis: A two factor permutation analysis of variance (PERMANOVA), (Anderson, 2005) based on Euclidean distance was used to test the null hypothesis of no differences in lionfish density or biomass between localities (fixed factor with five levels) and across sites (nested factor within localities with 2 to five levels). All analyses were conducted using Primer + Permanova v6.
Results
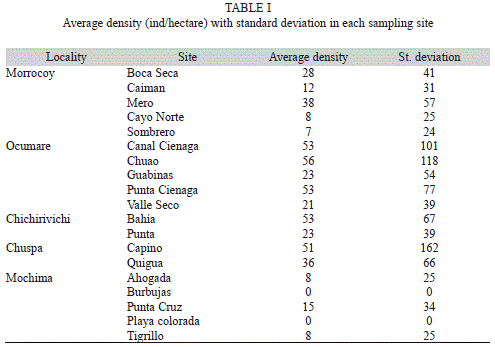
A total of 62 lionfish were found in 200 belt transects, with significant difference in density observed between localities but not across sites within each locality (Fig. 1). Lionfish were found at all sites except in Playa Colorada and Burbujas in Mochima. Mochima was also the locality with the lowest average density (6ind/ha). Average density across sites showed a high variability ranging from 7ind/ha in Cayo Norte (locality Morrocoy), to 55ind/ha in Punta Capino (Chuspa) (Table 1).
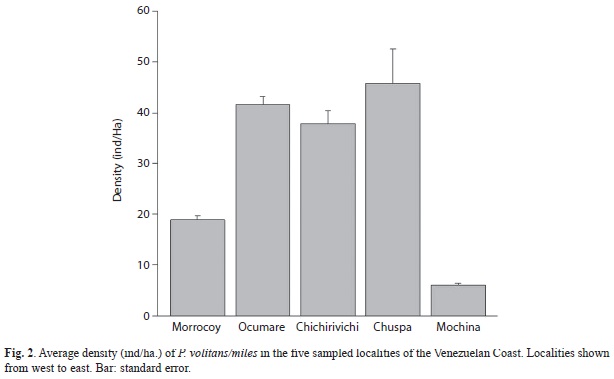
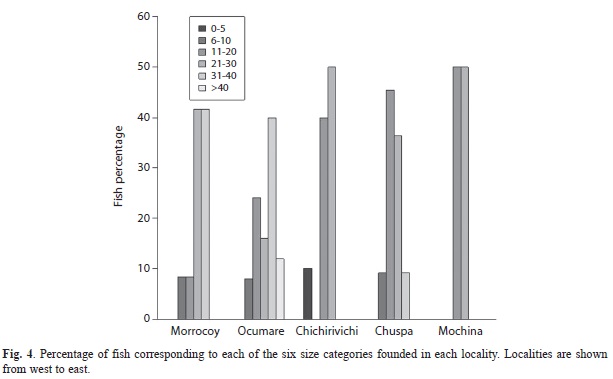
Statistical differences were only found between Ocumare and Mochima (PERMANOVA: p = 0.003) and between Chichirivichi and Mochima (PERMANOVA: p= 0.014). The highest component of variation was found at the level of residual, which indicates a higher variability between replicates from the same site. This high variability between replicates may explain the lack of differences found between most localities. Highest density was found in Chuspa, Ocumare de la Costa and Chichirivichi de la Costa and the lowest in Mochima, where only four individuals in total were observed (Fig. 2). The highest biomass was observed in Ocumare de la Costa (Fig. 3), where most individuals belong to the biggest sizes (Fig. 4).
Regarding their behavior, fifty percent of the lionfish were found hiding in caves and crevices. We also observed lionfish floating over the reef (32%) and individuals actively feeding (10%). Many of the lionfish observed were found in groups composed of up to seven individuals hiding in caves (44%). Almost all of the observed lionfish were adults (94%) ranging in size from 15cm to 45cm. Most individuals belonged to the size category between 30 and 40cm. A few juveniles were found in the sites of Cayo Norte in Morrocoy, Tigrillo in Mochima and Guabinas in Ocumare de la Costa, and in both sites of Chichirivichi de la Costa.
Discussion
This study confirmed that lionfish are well-established and widely spread along the coastline of Venezuela. Densities per site range between 6ind/ha to 55ind/ha and the density for Venezuelan Coast was 25.83±66.51. Average density of lionfish for the Venezuelan Coast seems to be lower than those reported for other invaded localities in the Caribbean, like the Bahamas, but is similar to average densities in North Carolina (Whitfield et al., 2002). Density along the Venezuelan Coast was also similar to some parts of the native range, like the density found off Kenya by Darling et al. (2011) of 25.1ind/ha and also to the average density reported in the Indo Pacific of 26.3ind/ha (Darling et al., 2011; Kulbicki et al., 2012). This study showed that lionfish abundance is extremely variable at every spatial scale, but particularly among replicates making it difficult to compare its abundance at regional level. High variability among replicates might be explained by patterns of patchy distribution displayed by this species which may clump in few caves and crevices while being absent from most of the reef area.
Chichirivichi de la Costa was one of the first localities were P. volitans/miles was reported (January 2010) and the locality with the highest number of reports according to CBM database (http://pezleon.cbm.usb.ve). It was therefore surprising to find only a few lionfish at Punta de Chichiriviche; the majority being of small sizes. This site is a common and popular site for recreational diving where lionfish extraction is practiced. In the second site (Bahia de Chichirivichi), less visited, we found twice the density of individuals, both juveniles and adults. The difference in lionfish abundance between these two sites suggests that regular extraction could maintain low population density of this species and therefore it might be an effective measure of control at a local scale.
Using the CBM database we were able to hypothesize about lionfish dispersion along the Venezuelan coast. We presume it started in the west between November and December of 2009. During 2010 sightings were made in several places of the central coast. We postulate that lionfish did not reach the eastern areas of the Venezuelan coast until the middle of 2011, almost two years after the first report. This may explain why a low average density and so few individuals were found in Mochima National Park during this study. This could also explain the differences in density between Venezuela and other invaded localities like Bahamas, where P. volitans/miles has been present since 2004 (Schofield, 2010). This also means there is the possibility of Venezuela having similar densities in the near future. Nevertheless, it has to be considered that the frequency of the reports from this database depends on the popularity of each diving site and so this conclusion must be considered carefully.
High variability in density of lionfish was found between sites at the same localities. Lionfish density may be influenced by factors acting locally like availability of refuges and structural complexity as it know they spend a lot of time hiding in holes and crevice (Fishelson, 1997). Sites in Ocumare de la Costa are coral reefs with low coral cover (less than 25%, unpublished data) but with a lot of holes and crevices where lionfish was observed specially in sites like La Cienaga, where mostly of the individuals were seeing hiding in caves. In Chichirivichi de la Costa Bay where also found high density. This site was form by group of rocks mixed with artificial structure made of concrete use by fisherman for anchoring their boats. Sites sampled in Mochima were rocky reef mostly flat with few holes and vertical structures.
The low density of lionfish of small sizes, less than 10cm. (only 6%) may be a consequence of the little power of detecting individuals specially of small sizes (Green et al., 2013). It was reported also a high underestimation of P. volitans densities especially in structural complex reef. This suggests that densities reported in this project may by lower than what they actually are, and this could be worse in structural complex reef.
Pterois volitans/miles has been reported as a crepuscular species with nocturnal behavior in its native environment (Fishelson, 1997). In this study, most of the individuals observed were found hiding in caves and crevice. Nevertheless, we observed several individuals actively feeding during the day on juveniles of Stegastes planifrons and adults of Stegastes partitus. We observed a fishing behavior similar to those described by Côté and Maljkovic (2010) wherein lionfish remain motionless and only move to strike and eat prey passing nearby. They are also considered as solitary individuals (Fishelson, 1997), which is contrary to our observation of groups of up to seven individuals of different sizes inhabiting a cave.
In conclusion, this study provides the first baseline of lionfish distribution and abundance based on wide-scale standardized surveys encompassing sites and localities along the coast of Venezuela, southern Caribbean. The results corroborate anecdotal information suggesting that the invasive species had become established invading rocky reef, coral reefs and coral communities on scales that vary from several hundred of meters to hundred of kilometers. Differences in density and biomass at localities suggest that habitat quality (e.g. food resources, environmental setting, etc) for this species might also be variable. The high variability among replicates might suggest a patchy distribution for this species. Future studies must focus in determining the underlying processes that explain these spatial patterns like structural complexity, availability of refuges and abundance of prey.
Acknowledgments
We are thankful to Cesar Herrera and Aldo Croquer for critically reviewing the manuscript and to Luis Miguel Montilla, Jose Cappeletto and Alejandra Hernandez for provide valuable help during dives and visual census. We also thank Juan Jose Cruz-Motta for his contribution in the design of the project. This project was financed by SARCE (South American Research on Coastal Ecosystem).
References
Albins, M. A. & Hixon, M. A. 2011. Worst case scenario: potential long-term effects of invasive predatory lionfish (Pterois volitans) on Atlantic and Caribbean coral-reef communities. Env. Bio. of Fishes. [ Links ]
Albins, M. & Hixon, M. 2008b. Invasive Indo-Pacific lionfish Pterois volitans reduce recruitment of Atlantic coral-reef fishes. Mar. Ecol. Prog Ser. 367: 233–238. [ Links ]
Anderson, M. J. 2005. Permutational multivariate analysis of variance. Department of Statistics, University of Auckland, Auckland. Retrieved from http://www.stat.auckland.ac.nz/~mja/prog/PERMANOVA_UserNotes.pdf. [ Links ]
Côté, I., & Maljkovic, A. 2010. Predation rates of Indo-Pacific lionfish on Bahamian coral reefs. Mar. Ecol. Prog. Ser. 404: 219-225. [ Links ]
Darling, E. S., Green, S. J., O’Leary, J. K., & Côté, I. M. 2011. Indo-Pacific lionfish are larger and more abundant on invaded reefs: a comparison of Kenyan and Bahamian lionfish populations. Biol. Inv. 13(9), 2045-2051. [ Links ]
Fishelson, L. 1997. Experiments and observations on food consumption, growth and starvation in Dendrochirus brachypterus and Pterois volitans (Pteroinae, Scorpaenidae). Env. Bio. of Fishes, 50(4), 397-403. [ Links ]
Ginsburg R., Kramer P., Lang J., Salle P. Steneck R. 1998. Atlantic and Gulf Rapid Reef Assesment (AGRRA). (http://agrra.org). [ Links ]
Green S., N. Tamburello, S. Miller, J. Akins, I. Côté. 2013. Habitat complexity and fish size affect the detection of Indo-Pacific lionfish on invaded reefs. Coral Reefs 32,413-421. [ Links ]
Gurevitch, J., & Padilla, D. 2004. Are invasive species a major cause of extinctions? Trends Ecol. Evolut. 19(9), 470-474. [ Links ]
Hamner, R. M., Freshwater, D. W., & Whitfield, P. E. 2007. Mitochondrial cytochrome b analysis reveals two invasive lionfish species with strong founder effects in the western Atlantic. J. Fish Biol. 71, 214–222. [ Links ]
Kulbicki M, Beets J, Chabanet P, Cure K, Darling E, Floeter SR, Galzin R, Green A, Harmelin, Vivien M, Hixon M, Letourneur Y, Lison de Loma T, McClanahan T, McIlwain J, MouTham G, Myers R, O’Leary JK, Planes S, Vigliola L, Wantiez L. 2012. Distributions of Indo-Pacific lionfishes Pterois spp. in their native ranges: implications for the Atlantic invasion. Mar. Ecol. Prog. Ser. 446, 189-205. [ Links ]
Lasso-Alcalá, O., & Posada, J. 2010. Presence of the invasive red lionfish, Pterois volitans (Linnaeus, 1758), on the coast of Venezuela, southeastern Caribbean Sea. Aquatic Invasions, 5(Supplement 1), S53–S59. [ Links ]
Lesser, M. P., & Slattery, M. 2011. Phase shift to algal dominated communities at mesophotic depths associated with lionfish (Pterois volitans) invasion on a Bahamian coral reef. Biol. Invasions. 13(8), 1855-1868. [ Links ]
López-Ordaz, A., Ortaz, M., & Rodriguez-Quintal, J. G. (2009). Trama trófica de una comunidad de peces en una pradera marina en el Caribe Venezolano. Rev. Biol. Trop. 57(4), 963-975. [ Links ]
Ruiz, G. M., Carlton, J. T., Grosholz, E. D., & Hines, A. H. 1997. Global invasions of marine and estuarine habitats by non-indigenous species: mechanisms, extent, and consequences. Amer. Zoo. 37(6), 621-632. [ Links ]
Schofield, P. 2009. Geographic extent and chronology of the invasion of non-native lionfish (Pterois volitans [Linnaeus 1758] and P. miles [Bennett 1828]) in the Western North Atlantic and Caribbean Sea. Aquat. Inv. 4(3), 473-479.
Schofield, P. 2010. Update on geographic spread of invasive lionfishes (Pterois volitans (Linnaeus, 1758) and P. miles (Bennett, 1828)) in the Western North Atlantic Ocean, Caribbean Sea and Gulf of Mexico. Aquat. Inv. 5(Supplement 1), S117-S122. [ Links ]
Whitfield, P. E., Gardner, T., Vives, S. P., Gilligan, M. R., Courtenay Jr, W. R., Ray, G. C., & Hare, J. A. 2002. Biological invasion of the Indo-Pacific lionfish Pterois volitans along the Atlantic coast of North America. Mar. Ecol. Prog. Ser. 235, 289-297. [ Links ]
Albins, M. & Hixon, M. 2008b. Invasive Indo-Pacific lionfish Pterois volitans reduce recruitment of Atlantic coral-reef fishes. Mar. Ecol. Prog Ser. 367: 233–238. [ Links ]
Anderson, M. J. 2005. Permutational multivariate analysis of variance. Department of Statistics, University of Auckland, Auckland. Retrieved from http://www.stat.auckland.ac.nz/~mja/prog/PERMANOVA_UserNotes.pdf. [ Links ]
Côté, I., & Maljkovic, A. 2010. Predation rates of Indo-Pacific lionfish on Bahamian coral reefs. Mar. Ecol. Prog. Ser. 404: 219-225. [ Links ]
Darling, E. S., Green, S. J., O’Leary, J. K., & Côté, I. M. 2011. Indo-Pacific lionfish are larger and more abundant on invaded reefs: a comparison of Kenyan and Bahamian lionfish populations. Biol. Inv. 13(9), 2045-2051. [ Links ]
Fishelson, L. 1997. Experiments and observations on food consumption, growth and starvation in Dendrochirus brachypterus and Pterois volitans (Pteroinae, Scorpaenidae). Env. Bio. of Fishes, 50(4), 397-403. [ Links ]
Ginsburg R., Kramer P., Lang J., Salle P. Steneck R. 1998. Atlantic and Gulf Rapid Reef Assesment (AGRRA). (http://agrra.org). [ Links ]
Green S., N. Tamburello, S. Miller, J. Akins, I. Côté. 2013. Habitat complexity and fish size affect the detection of Indo-Pacific lionfish on invaded reefs. Coral Reefs 32,413-421. [ Links ]
Gurevitch, J., & Padilla, D. 2004. Are invasive species a major cause of extinctions? Trends Ecol. Evolut. 19(9), 470-474. [ Links ]
Hamner, R. M., Freshwater, D. W., & Whitfield, P. E. 2007. Mitochondrial cytochrome b analysis reveals two invasive lionfish species with strong founder effects in the western Atlantic. J. Fish Biol. 71, 214–222. [ Links ]
Kulbicki M, Beets J, Chabanet P, Cure K, Darling E, Floeter SR, Galzin R, Green A, Harmelin, Vivien M, Hixon M, Letourneur Y, Lison de Loma T, McClanahan T, McIlwain J, MouTham G, Myers R, O’Leary JK, Planes S, Vigliola L, Wantiez L. 2012. Distributions of Indo-Pacific lionfishes Pterois spp. in their native ranges: implications for the Atlantic invasion. Mar. Ecol. Prog. Ser. 446, 189-205. [ Links ]
Lasso-Alcalá, O., & Posada, J. 2010. Presence of the invasive red lionfish, Pterois volitans (Linnaeus, 1758), on the coast of Venezuela, southeastern Caribbean Sea. Aquatic Invasions, 5(Supplement 1), S53–S59. [ Links ]
Lesser, M. P., & Slattery, M. 2011. Phase shift to algal dominated communities at mesophotic depths associated with lionfish (Pterois volitans) invasion on a Bahamian coral reef. Biol. Invasions. 13(8), 1855-1868. [ Links ]
López-Ordaz, A., Ortaz, M., & Rodriguez-Quintal, J. G. (2009). Trama trófica de una comunidad de peces en una pradera marina en el Caribe Venezolano. Rev. Biol. Trop. 57(4), 963-975. [ Links ]
Ruiz, G. M., Carlton, J. T., Grosholz, E. D., & Hines, A. H. 1997. Global invasions of marine and estuarine habitats by non-indigenous species: mechanisms, extent, and consequences. Amer. Zoo. 37(6), 621-632. [ Links ]
Schofield, P. 2009. Geographic extent and chronology of the invasion of non-native lionfish (Pterois volitans [Linnaeus 1758] and P. miles [Bennett 1828]) in the Western North Atlantic and Caribbean Sea. Aquat. Inv. 4(3), 473-479.
Schofield, P. 2010. Update on geographic spread of invasive lionfishes (Pterois volitans (Linnaeus, 1758) and P. miles (Bennett, 1828)) in the Western North Atlantic Ocean, Caribbean Sea and Gulf of Mexico. Aquat. Inv. 5(Supplement 1), S117-S122. [ Links ]
Whitfield, P. E., Gardner, T., Vives, S. P., Gilligan, M. R., Courtenay Jr, W. R., Ray, G. C., & Hare, J. A. 2002. Biological invasion of the Indo-Pacific lionfish Pterois volitans along the Atlantic coast of North America. Mar. Ecol. Prog. Ser. 235, 289-297. [ Links ]
1. Laboratorio de Ecología Experimental, Universidad Simón Bolívar, 1086, Caracas, Venezuela; estebanagudo@yahoo.com
2. Laboratorio de Sensores Remotos, Universidad Simón Bolívar, 1086, Caracas, Venezuela.
Received 01-IX-2013. Corrected 23-II-2014. Accepted 24-III-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}