










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.3 San José Sep. 2014
Bomb-cratered coral reefs in Puerto Rico, the untold story about a novel habitat: from reef destruction to community-based ecological rehabilitation
Los arrecifes de coral con craters-bomba en Puerto Rico, la historia no contada sobre un hábitat inusual: desde la destrucción de arrecifes hasta la rehabilitación ecológica basada en la comunidad
Los arrecifes de coral con craters-bomba en Puerto Rico, la historia no contada sobre un hábitat inusual: desde la destrucción de arrecifes hasta la rehabilitación ecológica basada en la comunidad
Edwin A. Hernández-Delgado1*,2*, Alfredo Montañez-Acuña1,2, Abimarie Otaño-Cruz1,2 & Samuel E. Suleimán-Ramos2
Abstract
Ecological impacts of military bombing activities in Puerto Rico have often been described as minimal, with recurrent allegations of confounding effects by hurricanes, coral diseases and local anthropogenic stressors. Reef craters, though isolated, are associated with major colony fragmentation and framework pulverization, with a net permanent loss of reef bio-construction. In contrast, adjacent non-bombarded reef sections have significantly higher benthic spatial relief and biodiversity. We compared benthic communities on 35-50 year-old bomb-cratered coral reefs at Culebra and Vieques Islands, with adjacent non-impacted sites; 2) coral recruit density and fish community structure within and outside craters; and 3) early effects of a rehabilitation effort using low-tech Staghorn coral Acropora cervicornis farming. Reef craters ranged in size from approximately 50 to 400m2 and were largely dominated by heavily fragmented, flattened benthos, with coral cover usually below 2% and dominance by non-reef building taxa (i.e., filamentous algal turfs, macroalgae). Benthic spatial heterogeneity was lower within craters which also resulted in a lowered functional value as fish nursery ground. Fish species richness, abundance and biomass, and coral recruit density were lower within craters. Low-tech, community-based approaches to culture, harvest and transplant A. cervicornis into formerly bombarded grounds have proved successful in increasing percent coral cover, benthic spatial heterogeneity, and helping rehabilitate nursery ground functions.
Key words: Benthic community structure, bombing impacts, community-based ecological rehabilitation, coral reefs, fish community structure, military activities, novel habitats.
Resumen
Los impactos ecológicos de las actividades militares de bombardeos en Puerto Rico se han descrito a menudo como mínimos, con recurrentes denuncias al confundir efectos por huracanes, enfermedades de corales y estresores antropogénicos locales. Los cráteres de arrecife, aunque aislados, están relacionados con una alta fragmentación de la colonia y pulverización del contorno, con una pérdida neta permanente de arrecife en bio-construcción. En contraste, secciones adyacentes de arrecife no bombardeado tienen mayor biodiversidad y mayor relieve espacial bentónico. Comparamos las comunidades bentónicas en cráteres-bomba de arrecifes de coral con 35-50 años de antigüedad en las islas de Vieques, Puerto Rico, en comparación con los sitios adyacentes no impactados; 2) la densidad de reclutamiento de coral y estructura de la comunidad de peces dentro y fuera de los cráteres; y 3) impactos preliminares de un esfuerzo de rehabilitación basado en la comunidad arrecifal usando tecnología simple con el cultivo del coral Staghorn Acropora cervicornis. Los cráteres de arrecife se distancian en tamaño de aproximadamente 50 a 400m2 y fueron dominados ampliamente por fragmentos de bentos aplanado, con una cubierta de coral generalmente por debajo de 2% y el predominio de taxones no constructores de arrecifes (es decir, tapetes de algas filamentosas, macroalgas). La heterogeneidad espacial bentónica fue significativamente menor dentro de cráteres que también resultaron en un reducido valor funcional como tierra de vivero de peces. La riqueza de especies de peces, abundancia y biomasa y densidad coral recluta fueron significativamente menores dentro de cráteres. Tecnología simple, basada en los enfoques de cultivo de comunidad, la cosecha y transplante de A. cervicornis en terrenos anteriormente bombardeados han demostrado un éxito al aumentar el porcentaje de cobertura de coral, la heterogeneidad espacial bentónica y ayudando a rehabilitar funcionalmente la tierra para vivero.
Palabras clave: estructura de la comunidad bentónica, impactos de bombardeo, rehabilitación ecológica basada en la comunidad, arrecifes de coral, estructura de la comunidad de peces, actividades militares, hábitat inusual
Long-term adverse ecological impacts of military maneuvers on coral reef ecosystems have remained as a concern as there is still limited information in the literature about impacts across multiple spatial and temporal scales. Most studies have often focused on very large spatial scale assessments, which have by default often overlooked some of the acute impacts on bomb-cratered coral reefs at smaller (i.e., fringing reef) spatial scales. Most published accounts were from studies conducted at Vieques Island, Puerto Rico (Raymond, 1978; DON 1979; DON 1980; DON 1986, Raymond & Dodge, 1980; Antonius & Weiner, 1982; GMI, 2003; GMI, 2005, Deslarzes, Nawojchik, Evans, McGarrity & Gehring, 2006; Evans, Nawojchik & Deslarzes, 2006; Hernández-Cruz, Purkis & Riegl, 2006; Kendall & Eschelbach, 2006; McGarrity & Deslarzes, 2006; Riegl, Moyer, Walker, kohler, Gilliam & Dodge, 2008; Bauer, Menza, Foley & Kendall, 2008; Bauer & Kendall, 2010) which were conducted over island wide spatial scales and found minimal destructive ecological impacts of bombing activities at such large scales, concluding that hurricanes and multiple localized human stressors (i.e., sedimentation, fishing) caused significant confounding effects. Even studies which have documented critical acute impacts of bombing and sedimentation across military-impacted coral reefs (IDEA, 1970; Carrera-Rodríguez, 1978; Rogers, Cintrón & Goenaga, 1978; Goenaga, 1986; Goenaga, 1991) did not provide a full quantitative characterization of the localized impacts on bomb-cratered reefs at reef-level spatial scales. None of these studies had either the temporal resolution to address long-term recovery of impacted sites. Therefore, the impacts of habitat fragmentation at across reef spatial scales associated to military activities and its long-term consequences on the recovery ability of local community structure and ecosystem resilience have still been poorly addressed.
Localized bombing impacts on coral reefs still remain controversial, and most of the literature has focused on blast fishing. This is known to cause extensive reef framework destruction across Indo-Pacific (McManus, Reyes & Nañola, 1997; Pet-Soede & Erdmann, 1998) and Red Sea coral reefs (Riegl, 2001), besides its concomitant overexploitation of fishery resources. Blast fishing impacts have caused significant loss of coral cover, an increase in the amount of bare substrate and rubble, and a decline in fish species richness and abundance (Riegl & Luke, 1999). These authors suggested that natural regeneration of impacted reef communities is likely to be very slow, possibly taking several hundred years, and that rehabilitation would be difficult since coral transplants would have to mimic the previously existing community. The frequency and magnitude of military bombing activities in Vieques Island showed a steady significant increase during the cold war years. Rosa-Serrano (1996) documented increasing crater abundance within bombarded areas between 1964 and 1988 using GIS-based analysis, suggesting a long-term increase of physical impacts of bombing. Porter (2000) found unexploded ordnance, leaking toxic 2-4-6-Trinitrotoluene (TNT) on and around reefs, and over 1,000 deteriorating barrels of unknown chemicals on the sunken military vessel USS Killen off southeast Vieques. Porter, Barton and Torres (2011) also found a statistically significant inverse correlation between the coral species richness, colony abundance and species diversity, and the density of military ordnance across reef scales in Vieques. There were also multiple animals across the reef food web polluted with toxic compounds similar to those present in unexploded ordnance. Chromium in sediments, and TNT in both, water and sediments, exponentially increased within areas still littered with unexploded ordnance.
Reef craters present in both, Culebra and Vieques Islands coral reefs are often very small in comparison to the scale of each island, each ranging in size from approximately 50 to 400m2. But these are largely dominated by heavily fragmented flattened benthos, with % coral cover usually below 2% and dominance by non-reef building taxa (i.e., filamentous algal turfs, macroalgae) (Fig. 1a-c). In contrast, adjacent non-bombarded reef zones are still dominated by consolidated benthos, with higher percent living coral cover and larger abundance of reef building species (Fig. 1d-f). Benthic spatial heterogeneity is also significantly lower within crater scales which also results in a lowered functional value as fish nursery ground. The fact that physical disturbance within bombarded grounds was so locally extensive resulted in a mosaic of habitat patches with permanent loss of reef framework and in potentially declining multiple ecosystem functions and services. Therefore, reef craters have become a de facto novel habitat, and as such, there is a need to address the ecological status of benthic and fish communities, as well as their recovery state three to five decades after bombing.
This study was aimed at: 1) documenting the condition of benthic communities within 35-50 year-old reef craters at Culebra and Vieques Islands, Puerto Rico, in comparison to adjacent non-bombarded sites within former military maneuver sites; 2) comparing coral recruit density and fish community structure within and outside reef craters; and 3) addressing the preliminary impacts of a community-based bombarded coral reef rehabilitation effort using low-tech approaches to cultivate threatened staghorn coral, Acropora cervicornis (Lamarck, 1816), and rehabilitate bombarded coral reefs.
Materials and Methods
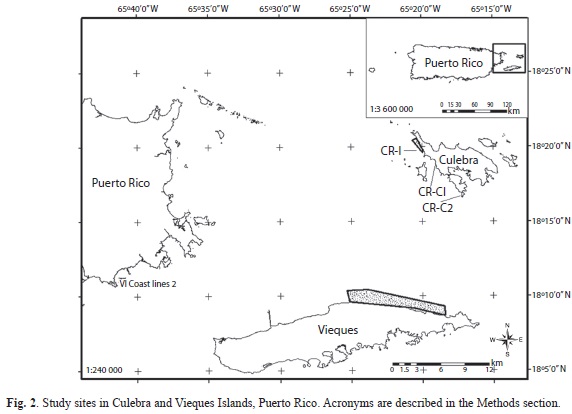
Study sites: Studies were conducted across 15 fringing reef sites, 11 at Culebra Island (located between 18°19.791’N, 65°19.943’W and 18°20.776’N, 65°20.498’W) and 4 at Vieques Island (located between 18°08.784’N, 65°18.482’W and 18°09.698’N, 65°25.073’W), off eastern Puerto Rico (State Plane, NAD83, FIPS PR5200, Fig. 2). Reef craters examined in this study ranged between 50 and 400m2. Sites were selected based on their representativeness of typical reef segments impacted by framework destruction as our aim was documenting what is the status of severely impacted reef sites 35-50 years after bombing impacts. Crater age was estimated from aerial photography and from anecdotal accounts from older fisher folks from both islands, and was a key factor for selecting impacted study sites to have a more accurate estimate of reef recovery trends through time. Control sites were selected on adjacent (usually <250m) sites not directly impacted by bombs. Reefs were subdivided by treatment (impacted-craters [n=7], non-impacted controls [n=8]), and depth (shallow, 1-3m [n=9]; deep, 6-10m [n=6]). In Culebra, sampling was conducted in 6 shallow (3 impacted, 3 controls) and 5 deep (2 impacted, 3 controls) reefs. In Vieques sampling was conducted in 2 shallow (1 impacted, 1 control) and 2 deep (1 impacted, 1 control) reefs. All benthic surveys in Culebra were conducted within the Canal Luis Peña no-take Natural Reserve (CLPNR) where all fishing is prohibited. Fish studies were conducted only in Culebra within the CLPNR to reduce confounding factors with fishing impacts elsewhere. Coral recruitment and reef rehabilitation studies were conducted in Culebra at the bombarded area described above (CR-I1, CR-I2), at Bahía Tamarindo (CR-C1, 18°18.877’N, 65°19.093’W), and at Punta Soldado (CR-C2, 18°16.846’N, 65°17.192’W). Bahía Tamarindo is also located within CLPNR and was used for artillery training activities and amphibious vehicle landing practices between 1920s and 1950s, but was never bombarded. Punta Soldado is located outside CLPNR and was used as a target site during the 1920s but never thereafter. These were used as control sites which underwent different levels of military activities, across different temporal scales, in comparison to direct recent bombing within reef cratered areas at impacted sites until 1970s. Also, these sites are part of a network of coral recruitment monitoring sites.
Benthic community: Benthic habitats were characterized across all sites through 3-6 replicate ten m-long digital video-transects. Number of replicates varied as a function of crater size and covered at least 50-75% of the impacted area within each crater. Transect deployment within each crater was haphazard, often separated by at least 5m. A total of six random, non-overlapping still images/transect were obtained and analyzed with Coral Point Count with excel extensions (v3.6) (Kohler & Gill, 2006) to address percent cover of all benthic components, including coral, algal functional groups (macroalgae, turf, crustose coralline algae [CCA], erect calcareous algae [ECA], Halimeda spp.), cyanobacteria, other components, sand, rubble, and bare substrate. A total of 20 random dots per image were used. Coral species richness, species diversity index (H’n) (Shannon & Weaver, 1948), and evenness (J’n) (Pielou, 1966) were also calculated.
Coral recruits: Coral recruit densities were addressed only in Culebra using triplicate 2.25 x 2m quadrat grids subdivided in 12 replicate 0.75 x 0.50m quadrats/grid from one shallow (CR-IS) and one deep crater (CR-ID), and from two control non-impacted sites at Bahía Tamarindo (CR-C1), and Punta Soldado (CR-C2). High-resolution digital images were collected and all hydrocoral/scleractinian recruits with a diameter below 5cm were counted and identified to the lowest taxon possible.
Fish community structure: Fish communities were characterized only in Culebra using stationary visual censuses within craters (impacted) and adjacent (control) locations following a slight modification from Bohnsack and Bannerot, (1986). Data was collected within a 5 m-radius imaginary cylinder during a period of 15min. All individuals were counted, identified to the lowest taxon possible, and standard fork length was estimated. Size data were used to estimate biomass. Weight-length relationships were calculated following Bohnsack and Harper (1988). Basic information of the fish community structure reported in this study included species richness, abundance, H’n, J’n, total biomass, and piscivore biomass. Reef structural complexity is known to have an important influence on fish community structure (Roberts & Ormond, 1987). A 6-point scale was used to characterize a reef structural heterogeneity index (RSHI) as follows: 0= no vertical relief; 1= low and sparse relief; 2= low but widespread relief; 3= moderately complex; 4= very complex with numerous caves and fissures; 5= exceptionally complex with high coral cover and numerous caves and overhangs (Hawkins et al., 1999).
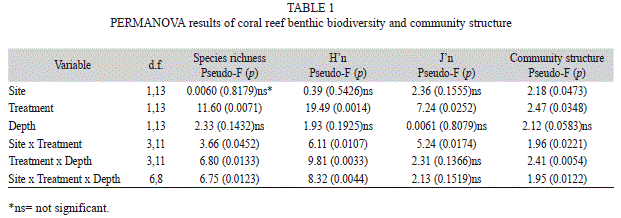
Statistical analyses: A three-way permutational analysis of variance (PERMANOVA) was used to test the null hypothesis of no significant difference in benthic biodiversity parameters and community structure between sites (Culebra, Vieques), treatment level (bombarded areas, non-impacted controls), and depth (1-3m, 6-9m) using PRIMER-e v.6.1.14 (Anderson, Gorley & Clarke, 2008). Principal component ordination (PCO) was used to determine which benthic taxa explained spatial clustering patterns of benthic communities. Proportional data on percent benthic components cover were √-transformed prior to analysis. A one-way PERMANOVA was used to test spatial patterns of coral recruits between bombarded and non-impacted sites in Culebra, followed by PCO. A one-way analysis of similarity (ANOSIM) was used to test spatial patterns of fish communities between bombarded and non-impacted sites in Culebra, followed by a multi-dimensional scaling (MDS) analysis (Clarke & Warwick, 2001). Data were also √-transformed prior to analysis. All tests were based in 10 000 permutations. Fish community data also were correlated (Spearman) with RSHI.
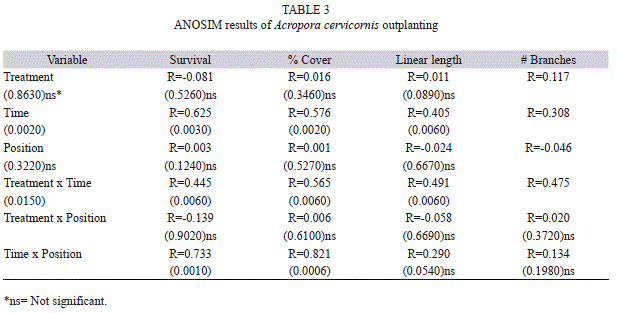
Coral reef rehabilitation: A total of 2 000 corals were harvested from existing low-tech coral farms through the Community-Based Coral Aquaculture and Reef Rehabilitation Project and outplanted to adjacent coral reefs within former military maneuver ranges at two sites in Culebra Island, Bahía Tamarindo and Punta Soldado. Sites selected for outplanting were located within a flat shallow reef (<2.5m) used as artillery maneuver areas at Bahía Tamarindo (impacted site) and at a reef segment at Punta Soldado non-impacted by bombing or artillery maneuvers since 1920s (control site). Corals were attached to masonry nails driven to reef bottom, outplanted in patches of densities ranging from 80 to 120 per 25m2. Survival rates and growth were addressed following two representative patches located at elevated rocky outcrops and two patches adjacent to reef sand pockets at increasing time intervals during a year. A two-way ANOSIM was used to test the null hypotheses of no significant change in coral survival rates, skeletal extension, and branch production through time and between sites.
Results
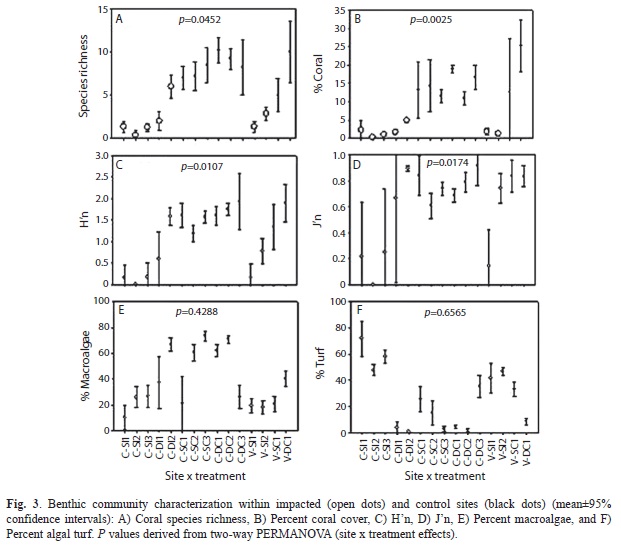
Benthic community: Coral reef benthic communities across bombarded areas showed significantly more physical destruction and an altered coral assemblage in comparison to control non-impacted sites (Table 1). There was a significantly different benthic community structure between sites (p=0.0473) and treatments (p=0.0348). Also, interactions between site and treatment, treatment and depth, and among site, treatment and depth were highly significant. Bombarded sites were characterized by having significantly lower coral species richness (p=0.0452), percent coral cover (p=0.0025), H’n (p=0.0107), and J’n (p=0.0174) (Fig. 3a-d). Mean coral species richness within bombarded bottoms was 2.2/transect, while mean value at adjacent non-impacted control sites was 8.8/transect. Mean living coral cover within bombarded bottoms was 1.9% and 15.7% at control sites. Coral cover was also higher at deeper (13%) than at shallower sites (6.5%). Mean H’n within bombarded bottoms was 0.4912 and 1.6101 at control sites, while mean J’n within bombarded bottoms was 0.4169 and 0.7834 at control sites. Species richness and H’n also had significant treatment x depth, and site x treatment x depth interactions. Macroalgal cover was higher on control sites (47%), in comparison to bombarded areas (29%), while algal turf was higher within bombarded grounds (26%), in comparison to control sites (16%) (Fig. 1e-f). But none of these differences were significant.
The percent relative cover of the most important reef building coral species was significantly lower within bombarded areas (Fig. 4). Montastraea (=Orbicella) annularis (Ellis & Solander, 1786) averaged 0.05% within bombarded areas and 4.1% at control sites, while O. faveolata (Ellis & Solander, 1786), O. franksi (Gregory, 1895), and M. cavernosa Linnaeus, 1767 averaged 1.3, 0.5, and 1.0%, respectively, at control sites. None of these species were present within bombarded areas. Colpophyllia natans (Houttuyn, 1772) averaged 0.01% within bombarded grounds and 0.38% in control sites. Diploria strigosa (Dana, 1846) averaged 0.7% at control sites and was absent within bombarded areas, and Siderastrea siderea (Ellis & Solander, 1786) had a mean 0.14% cover within bombarded areas and 1.8% in control sites.
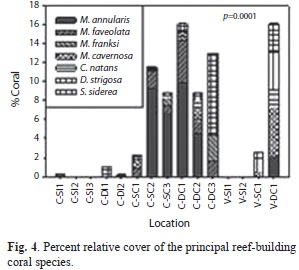
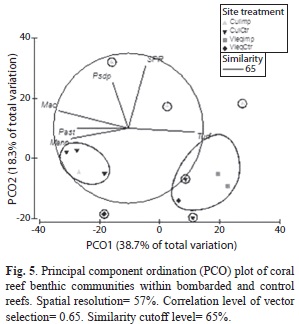
Principal component ordination (PCO) analysis showed two larger clusters of reef communities, and 5 individual sites (Fig. 5). Impacted sites at Culebra were dominated by open substrates composed by a mixture of bare bedrock, rubble and sand pockets (SPR), algal turf, brown macroalgal patches (e.g., Dictyota spp.), and sporadic colonies of octocoral Pseudopterogorgia spp. (Fig. 1a-b). Vieques impacted sites were dominated by algal turfs (Fig. 1c). Culebra control site showed a higher spatial heterogeneity mostly dominated by macroalgae, M. annularis, and P. astreoides (Fig. 1c-d). Control sites at Vieques were also dominated by turf, and in a lesser degree a mixed octocoral community. The proposed PCO solution explained 57% of the observed spatial variation.
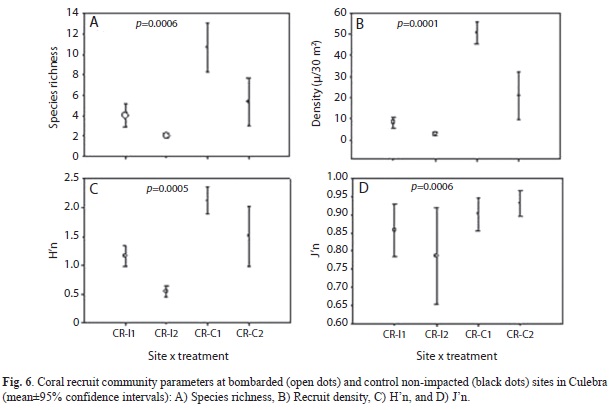
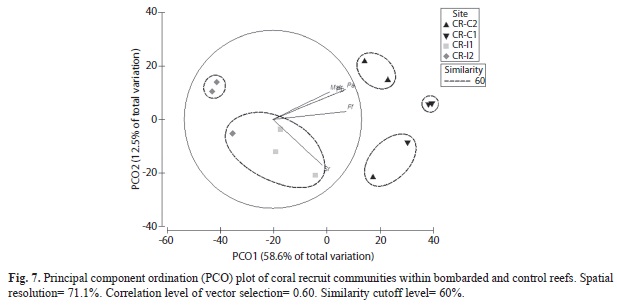
Coral recruits: Coral recruit density was significantly higher (PERMANOVA, Pseudo-F=6.55, p=0.0001) within non-impacted control sites in comparison to bombarded areas. Control site CR-C1 located within CLPNR averaged 51 colonies/30m2, while CR-C2 outside CLPNR averaged 21 colonies/30m2 (Fig. 6). Impacted site CR-I1 averaged 8 colonies/30m2, while CR-I2 averaged less than 3 colonies/30m2. Both impacted sites were also located within CLPNR. ANOSIM analysis showed that coral recruit community structure was significantly different between treatments (R=0.830, p=0.0001). Also, species richness (R=0.736, p=0.0006), and H’n (R=0.747, p=0.0006) were significantly higher at control sites than at bombarded areas. No significant difference in J’n was documented. Brooder species such as Favia fragum (Esper, 1795), Siderastrea radians (Pallas, 1766), and Porites astreoides (Lamarck, 1816) were dominant at control sites, while lower abundances of S. radians and P. astreoides characterized bombarded sites, particularly, at deeper impacted areas. PCO analysis showed five different clusters of reef communities, and five individual sites (Fig. 7). The three clusters composed of control non-impacted sites were explained by P. astreoides, P. porites, F. fragum, and Millepora striata (Lamarck, 1816). Bombarded sites clusters were determined by S. radians. The proposed solution by PCO explained 71.1% of the observed spatial variation.
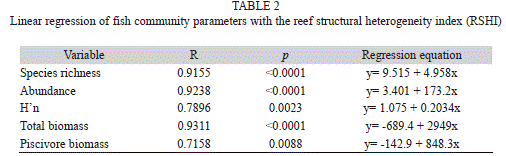
Fish community: Fish community structure also showed significant difference (p<0.0001) between treatment levels that were mostly related to a highly significant decline (p=0.0030) observed in the reef structural heterogeneity index (RSHI) within bombarded sites (Fig. 8a). RSHI had a mean value of 0.69 within bombarded areas and 2.72 within control sites. Fish species richness was significantly higher (23.4 per count) at control sites (p=0.0020) than at bombarded areas (12.6) (Fig. 8b). Fish abundance was also significantly higher (p=0.0002) at control sites (491) versus bombarded sites (108) (Fig. 8c). Also, H’n was significantly higher (p=0.0020) within control areas (1.6744) in comparison to bombarded grounds (1.1716) (Fig. 8d). Total fish biomass was significantly higher (p=0.0001) at control sites (7 697g) than at bombarded areas (999g) (Fig. 8e). Similarly, piscivore biomass was significantly higher (p=0.0002) at control sites (2,406 g) than at bombarded areas (206g) (Fig. 8f). All fish community parameters showed a highly significant linear regression (p<0.0088) with RSHI (Table 2), suggesting the strong permanent negative impacts of bombing activities on the demolition of reef framework and the net decline in fish communities associated to losing spatial benthic heterogeneity. Significant reef functional herbivore guilds such as scrapers, including Scarus iserti Bloch, 1790, S. vetula Schneider, 1801, Sparisomq viride (Bonnaterre, 1788), S. rubiprinne (Valenciennes, 1839), and S. radians (Valenciennes, 1839), and browsers such as Acanthurus coeruleus Schneider, 1801 were largely absent from reef craters, in comparison to adjacent non-bombarded sites. Also, important piscivore guilds such as groupers, including Epinephelus guttatus (Linnaeus, 1758), E. adscensionis (Osbeck, 1765), Cephalopholis fulva (Linnaeus, 1758), and C. cruentata (Lacepède, 1802), and snappers Lutjanus jocu (Schneider, 1801), L. analis (Cuvier, 1828), and L. apodus (Walbaum, 1892) were also absent from reef craters. Fishing impacts was not a factor influencing observed differences in fish community structure within and outside craters as fish data were collected from sites located within the no-take CLPNR.
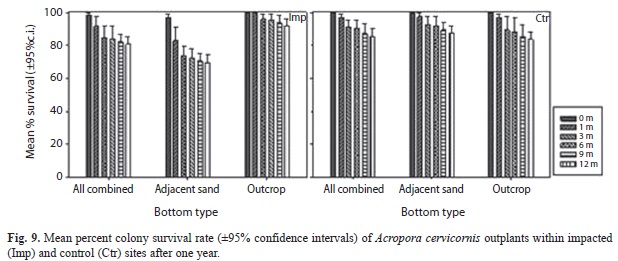
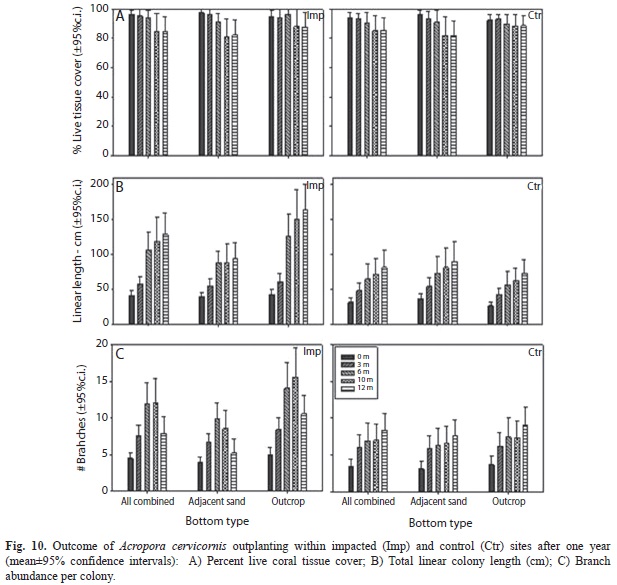
Coral reef rehabilitation: Mean percent colony survival rates of Acropora cervicornis outplants was 81% at impacted sites and 86% at control sites after one year, with a mean survival of 88% at impacted sites and 70% at impacted sites on low-relief reef patches adjacent to sand (Fig. 9). Percent survival at elevated rocky outcrops reached 92% at impacted sites and 84% at control sites. Percent live coral tissue cover on outplanted colonies averaged 85% at both, impacted and control sites after one year (Figure 10a), ranging from 82 to 88% within impacted sites in low-relief patches adjacent to sand and in elevated outcrops, respectively. Mean % live tissue cover ranged from 82 to 89% within impacted sites in low-relief patches adjacent to sand and in elevated outcrops, respectively. Total outplanted colony linear length showed a mean overall increase from 41 to 129cm across impacted sites, and from 32 to 81cm across control sites after one year (Figure 10b). Total outplanted colony branch abundance/colony showed a mean overall increase from 4.5 to 14.4 cm across impacted sites, and from 3.4 to 8.3cm across control sites after one year (Figure 10c). Temporal effects were significant for all variables, but treatment and position effects were not (Table 3).
Discussion
Profound, acute and persistent negative impacts of historical bombing activities were documented in Culebra and Vieques Islands, Puerto Rico, across coral reef craters spatial scales. Severely impacted reef segments were characterized by having significantly lower spatial relief, bedrock exposure and an abundant mixture of unstable turf-covered rubble and bedrock boulders demolished by explosions. These substrates were also characterized by low coral colony abundance, low percent living coral cover, low coral species richness and H’n, when compared to adjacent control sites. Similarly, coral recruit communities were significantly more depauperate within impacted grounds than in control sites, either within or outside the no-take CLPNR, which suggest the persistent inability of coral recruits to survive the natural oceanographic dynamics of unstable substrates within bombarded sites. This is consistent with severe impacts by blast fishing documented elsewhere (Riegl & Luke, 1999; Riegl, 2001). The permanent lack of natural recovery ability of 35-50 years old bomb-cratered coral reef segments, when compared to adjacent non-bombarded control sites dominated by massive reef-building species such as M. annularis species complex, implies that at the local ecosystem scale, bombarded coral reefs have shown a permanent shift in composition and functions, that full recovery of previously existing benthic community structure and spatial heterogeneity may take centuries. Coral recruitment rates of critical reef-building species across the northeast Caribbean region are increasingly low (Rogers, Fitz, III, Gilnack, Beets & Hardin, 1984; Edmunds & Elahi, 2007; Edmunds, Ross & Didden, 2011), suggesting that habitat fragmentation by bombing has resulted in a permanent localized loss of coral reproductive stock and that in combination with natural low recruitment rates of most reef-building species, natural recovery of composition and functions is very unlikely.
Bombarded areas were also characterized by sustaining lower fish species richness, H’n, abundance, and biomass, as a result of the permanent loss and lack of recovery of reef benthic spatial relief. They also had a very low abundance or absence of significant fish functional groups of herbivores and carnivores, including multiple commercially-important species. These findings are consistent with IDEA (1970) which estimated at least 10 times higher fish densities outside cratered reefs in Culebra Island, though no quantitative parameters were provided. Riegl (2001) found that coral cover decreased, bare substratum and rubble increased, and fish communities changed within areas impacted by blast fishing in Egypt. Depauperate fish assemblages within bombarded reef segments were also consistent with declining fish communities documented on reefs that have already shown rapid benthic community decline as a result of climate change impacts (Jones, McCormick, Srinivasan & Eagle, 2004, Graham et al., 2006; Pratchett et al., 2008). Such changes may become more pronounced as coral cover continues to decline and as fishing pressure continues to increase (Pratchett, Hoey & Wilson, 2014). Fishing impacts were not a factor in this study as all fish data collection was conducted within the no-take CLPNR. Status of more diverse and rich fish communities across control sites is consistent with previous accounts across similar spatial scales for the site (Hernández-Delgado, Rosado-Matías & Sabat, 2006). Therefore, differences in fish community structure were presumed to occur at the studied spatial scales as the result of altered benthic community structure and spatial heterogeneity due to bombing activities.
Individual reef craters are often small in size (50-400m2) and isolated in space, which render them as very small spatial units generally disregarded as having low ecological significance as they may represent a small geographical proportion of reef surface area in comparison to island wide spatial scales. Studies of bombing impacts at small spatial scales are still very limited. Dodge (1981) found no significant impacts of military bombing on M. annularis growth rates on individual coral core samples from Vieques, but Macintyre, Raymond & Stuckenrath (1983) found significant destruction by bombing of shallow Acropora palmata (Lamarck, 1816) and Porites porites (Pallas, 1766) frameworks. Porter et al. (2011) also found a statistically significant inverse correlation between the coral species richness, colony abundance and species diversity, and the density of military ordnance across reef scales in Vieques. Nonetheless, at smaller ecological scales (e.g., fringing reef unit), reef craters represent localized mosaics of reef segments that were severely reduced to a flattened, unstable, demolished reef bottom, with depauperate biodiversity, that have shown little or no recovery even after three to five decade temporal scales. Placed within the context of current sea surface warming trends, recurrent massive bleaching events, and documented decline of northeastern Caribbean coral reefs (Miller et al., 2009; Hernández-Pacheco, Hernández-Delgado & Sabat, 2011; Edmunds, 2013), net recovery of ecosystem structure and functions within bombarded grounds is unlikely to occur, rendering them as novel habitats (sensu Graham, Cinner, Norström & Nyström, 2014). This suggests that future trajectories of dramatically changed reef communities constituting novel habitats will be quite different from the past, and embracing novel futures may enable more pragmatic approaches (e.g., rehabilitating ecological functions instead of restoring original diversity) to maintaining or re-building the dominance of massive reef-building corals from the past.
The lack of meaningful natural coral reef recovery within 35-50 year-old reef craters from Culebra is alarming, but surprisingly, still poorly addressed. Our study suggests that coral community recovery is minimal within reef craters and limited to sporadic ephemeral species such as S. radians and P. astreoides. There is increasing evidence that natural coral reef recovery ability from blasting even across small spatial scales can become severely limited with increasing spatial and temporal scale of destruction. Extensively blasted areas for fishing in Indonesia showed no significant recovery within a period of six years despite adequate coral larval supply from adjacent reefs (Fox & Caldwell, 2006). Extensive bombing can result in the formation of unstable coral rubble fields that can move with ocean currents and storm swells, causing extended mortality on adjacent remnant patches of living corals and that can also prevent successful coral larval recruitment over unstable bottoms (Fox, Pet, Dahuri & Caldwell, 2002; 2003; Lindhal, 2003; Raymundo, Maypa, Gomez & Cadiz, 2007). Therefore, reef natural recovery ability within directly bombarded grounds seems poorly probable and will require assisted coral reef rehabilitation methods (Bowden-Kerby, 1997; Raymundo et al., 2007; Hernández-Delgado, Suleimán, Olivo, Fonseca & Lucking, 2011). This can be feasible across small spatial scales similar to those of reef craters. Nonetheless, human intervention is unlikely to be effective on large spatial scales due to prohibitive costs, highlighting the need for a combination of effective management approaches to foster the rehabilitation of reef ecological functions and ecosystem resilience.
A particular concern is that a habitat once dominated by a M. annularis species complex framework has not shown any sign of recovery over the course of several decades through sexual coral larval recruitment. Though coral larval settlement do occur within the crater, coral spat mortality appears to be high largely due to the unstable fragmented nature of the bottom. Considering the significant decline of M. annularis species complex percent living cover across the region (Miller et al., 2009; Hernández-Pacheco et al., 2011, Edmunds, 2013), recovering benthic spatial heterogeneity ecological functions is largely improbable. Therefore, an alternative strategy that can potentially achieve rapid results in rehabilitating shallow reef ecological functions as juvenile fish nursery grounds is the use of community-based, low-tech farming and outplanting of rapid-growing Acropora cervicornis. Low-tech, community-based approaches to culture, harvest and transplant A. cervicornis into formerly bombarded grounds proved highly successful in fomenting increasing benthic spatial heterogeneity, while fostering meaningful community-based participation. Outplanted colonies showed outstanding survival and growth rates. Observed decline occurred as a result of partial coral mortality associated to massive runoff events from deforested steep slopes adjacent to the coastline following heavy rainfall. Higher percent survival rate observed on rocky outcrops at impacted sites (within no-take CLPNR) was the result of lower predation impacts by corallivore gastropod Coralliophila abbreviata Lamarck, 1816 and C. caribaea Abbott, 1958, and by fireworm Hermodice carunculata Pallas, 1766; in comparison to adjacent controls outside the reserve. This could be the result of lack of invertebrate predators across control non-reserve sites, which is consistent with previous accounts of fish community structure from the site (Hernández-Delgado et al., 2006). This suggests that A. cervicornis farming and outplanting is a key successful tool to help rehabilitate shallow reef nursery grounds. But further, it also showed that reef trophic condition is a key element in determining reef rehabilitation success. Therefore, the combination of a no-take marine protected area designation and low-tech coral farming and outplanting are key management tools to foster the rehabilitation of reef ecological functions and ecosystem resilience of impacted sites across reef spatial scales. History has shown that introducing and fostering compliance with coral reef conservation measures in a small island community still traumatized by historical military practices and by past actions by the government perceived by local communities as serious violation of trust has become a paramount challenge. Nonetheless, community-based participatory management approaches have proved to be a highly successful and empowering strategy to rehabilitate impacted coral reefs ecosystems and to educate base communities through hands-on experience on the significance of reef conservation and rehabilitation.
There is also a concern that military impacts on coral reef are ecologically persistent and that they may still represent a risk of toxic pollution further threatening reef recovery. Goenaga (1986, 1991) suggested that the large abundance of unexploded ordnance and the potential leaching of pollutants from bombs in coral reefs may significantly impair their future recreational and fishing value. Porter (2000) found evidence of abundant “unexploded bombs, artillery shells, and shell casings on the coral reef and in adjacent seagrass beds; burial and shading of coral reef organisms by unexploded ordnance and ordnance debris; fracturing of the coral reef framework and the underlying coral bed rock, and the existence of bombs and bomb fragments impregnated into the reef; the existence of parachutes from flares and cluster bomb fragments draped over corals and other coral reef flora and fauna; and the existence of unexploded bombs leaking materials into coral reef environment and creating a limited “dead zone” around the bombs”. Porter et al. (2011) also found a statistically significant inverse correlation between the density of unexploded ordnance and coral species richness, coral colony abundance, and coral species diversity, with reefs with the highest concentrations of bombs and bomb fragments having the lowest health indices and the lowest species diversity. Further, evidence of leaking toxics from unexploded ordnance has also been documented in reef demersal fauna from Vieques (Porter et al., 2011). These factors, in combination with the long-term impacts of uncontrolled, poorly planned land uses in the post-military land development boom in Culebra and Vieques has also resulted in the implementation of a non-sustainable development model with paramount adverse ecological and socio-economic implications to environmental and socio-economic sustainability of the islands (Hernández-Delgado et al., 2012; Ramos-Scharrón, Amador & Hernández-Delgado, 2012). This is an aspect that deserves further research.
Our findings showed that there was still an untold story about bombing impacts across small reef spatial scales, that benthic habitat destruction is ecologically long-lasting (over decadal scales) and that lack of net recovery has resulted in converting impacted reefs in a de facto novel habitat. Natural reef recovery abilities within bombarded reefs need to be continuously monitored. Targeted monitoring efforts will become critical in the context of increasing sea surface temperature and its long-term impacts on coral reefs. Declining reefs across the region due to climate change impacts may aggravate the ability of bombarded reefs to show at least a modest degree of recovery. The lack of natural recovery ability coupled with a declining social-ecological system significantly reduces the probability of ecosystem and socio-economic recovery. A major community-based effort should be launched to foster improved coral reef ecosystem and socio-economic resilience rehabilitation. Integration of local stakeholders should help improve efforts by local natural resource managers and decision-makers to accelerate recovery of ecological functions of degraded reef ecosystems and socio-economic systems, but also to repair communication and trust.
Acknowledgments
This study was possible thanks to the support provided to E.A. Hernández-Delgado by the National Science Foundation HRD #0734826 through the Center for Applied Tropical Ecology and Conservation. We also thank the partial support from the Caribbean Coral Reef Institute of the University of Puerto Rico (NA04NOS4260206, NA05NOS4261159, and NA07NOS4000192) to E.A. Hernández-Delgado, and the partial support provided by NOAA and The Nature Conservancy (MAR-SAM-110110) through Sociedad Ambiente Marino to S. Suleimán-Ramos and E.A. Hernández-Delgado.
References
Anderson, M. J., Gorley R. N., & Clarke K. R. (2008). PERMANOVA + for PRIMER: Guide to Software and Statistical Methods. Plymouth, U.K.: PRIMER-E. [ Links ]
Antonius, A., & Weiner, A. (1982). Coral reefs under fire. Marine Ecology Progress Series, 3, 255-277. [ Links ]
Bauer, L. J., Menza, C., Foley, K. A., & Kendall, M. S. (2008). An ecological characterization of the marine resources of Vieques, Puerto Rico. Part I: Historical data synthesis. Prepared by National Centers for Coastal Ocean Science (NCCOS) Biogeography Branch in cooperation with the Office of Response and Restoration. Silver, Spring, MD.: NOAA Technical Memorandum NOS NCCOS 86. [ Links ]
Bauer, L. J., & Kendall, M. S. (eds.). (2010). An Ecological Characterization of the Marine Resources of Vieques, Puerto Rico Part II: Field Studies of Habitats, Nutrients, Contaminants, Fish, and Benthic Communities. Silver, Spring, MD.: NOAA Tech. Memorandum NOS NCCOS 110. [ Links ]
Bohnsack, J. A., & Bannerot, S. P. (1986). A stationary census technique for quantitatively assessing community structure of coral reef fishes. Silver, Spring, MD.: NOAA Technical Report NMFS 41. [ Links ]
Bohnsack, J. A., & Harper, D. E. (1988). Length-weight relationships of selected marine reef fishes from the Southeastern United States and the Caribbean. Silver, Spring, MD.: NOAA Technical Memorandum NMFS-SEFC-215. [ Links ]
Bowden-Kerby, A. (1997). Coral transplantation in sheltered habitats using unattached fragments and cultured colonies. Proceedings from the 8th International Coral Reef Symposium, 2, 2063-2068. [ Links ]
Carrera-Rodríguez, C. J. (1978). Evaluación sobre las consecuencias de las actividades militares llevadas a cabo en Vieques, P.R. por las Fuerzas Armadas de los EE.UU. de América en los sistemas marinos del este de Vieques, con atención principal a los arrecifes de coral. Report to the Department of Natural Resources, San Juan, Puerto Rico. [ Links ]
Clarke, K. R., & Warwick, R. M. (2001). Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd Ed. Plymouth Marine Laboratory, UK: PRIMER-E, Ltd. [ Links ],
Deslarzes, K. J. P., Nawojchik, R., Evans, D. J., McGarrity, C. J., & Gehring, P. (2006). The condition of fringing reefs off former military training areas at Isla de Culebra and Isla de Vieques, Puerto Rico: preliminary results. Proceedings from the 10th International Coral Reef Symposium, Japan, 1152-1159. [ Links ]
Dodge, R. E. 1981. Growth characteristics of reef-building corals within and external to a naval ordnance range: Vieques, Puerto Rico. Proceedings from the 4th International Coral Reef Symposium, 2, 241-248. [ Links ]
DON (U.S. Department of the Navy). (1979). Draft Environmental Impact Statement for the Continued Use of the Atlantic Fleet Weapons Training Facility Inner Range (Vieques). Prepared by TAMS (Tippetts, Abbett, McCarthy, Stratton) and Ecology and Environment, Inc. [ Links ]
DON (U.S. Department of the Navy). (1980). Final Environmental Impact Statement for the Continued Use of the Atlantic Fleet Weapons Training Facility Inner Range (Vieques). Prepared by TAMS (Tippetts, Abbett, McCarthy, Stratton) and Ecology and Environment, Inc. [ Links ]
DON (U.S. Department of the Navy). (1986). Environmental Assessment of Continued Use of the Atlantic Fleet Weapons Training Facility Inner Range, Vieques, Puerto Rico. Prepared by Ecology and Environment, Inc. [ Links ]
Edmunds, P. J. (2013). Decadal-scale changes in the community structure of coral reefs of St. John, US Virgin Islands. Marine Ecology Progress Series, 489, 107-123. [ Links ]
Edmunds, P. J., & Elahi, R. (2007). The demographics of a 15-year decline in cover of the Caribbean reef coral Montastraea annularis. Ecological Monographs, 77, 3-18. [ Links ]
Edmunds, P. J., Ross, C. L. M., & Didden, C. (2011). High, but localized recruitment of Montastraea annulalris complex in St. John, United States Virgin Islands. Coral Reefs, 30, 123-130
Evans, D. J., Nawojchik, R., & Deslarzes, K. J. P. (2006). The status of coral reef fish populations off former military ranges at the islands of Culebra and Vieques, Puerto Rico: preliminary data. Proceedings from the 10th International Coral Reef Symposium, 1105-1109. [ Links ]
Fox, H. E., Pet, J. S., Dahuri, R., & Caldwell, R. L. (2002). Coral reef restoration after blast fishing in Indonesia. Proceedings from the 9th International Coral Reef Symposium, 2, 969–976. [ Links ]
Fox, H. E., Pet, J. S., Dahuri, R., & Caldwell, R. L. (2003). Recovery in rubble fields: long-term impacts of blast fishing. Marine Pollution Bulletin, 46, 1024-1031. [ Links ]
Fox, H. E., & Caldwell, R. L. (2006). Recovery from blast fishing on coral reefs: A tale of two scales. Ecol. Appl., 16, 1631-1635. [ Links ]
GMI (Geo-Marine, Inc.). (2003). Reef ecosystem baseline assessment survey and monitoring, Vieques Island, Naval Station Roosevelt Roads, Puerto Rico. Prepared for Atlantic Division, Naval Facilities Engineering Command, Norfolk, Virginia. [ Links ]
GMI (Geo-Marine Inc.). (2005). An assessment of the condition of coral reefs off the former Navy bombing ranges at Isla de Culebra and Isla de Vieques, Puerto Rico. Prepared for the Department of Defense, Legacy Resource Management Program, Arlington, Virginia, and US Army Corps of Engineers, Huntsville, Alabama. [ Links ]
Goenaga, C. (1986). Los arrecifes costaneros en Puerto Rico: Estado actual e implicaciones sociales. Science-Ciencia, 13, 78-91. [ Links ]
Goenaga, C. (1991). The state of coral reefs in the wider Caribbean. Interciencia, 15, 12-20. [ Links ]
Graham, N. A. J., Wilson, S. K., Jennings, S., Polunin, N. V. C., Bijoux, J. P., & Robinson, J. (2006). Dynamic fragility of oceanic coral reef ecosystems. Proceedings of the National Academic Sciences. USA, 103, 8425-8429. [ Links ]
Graham, N. A. J., Cinner, J. E., Norström, A.V., & Nyström, M. (2014). Coral reefs as novel ecosystems: embracing new futures. Current Opinion in Environment Sustainability, 7, 9-14. [ Links ]
Hawkins, J. P., Roberts, C. M., Van’t Hof, T., De Meyer, K., Tratalos, J., & Aldam, C. (1999). Effects of recreational scuba diving on Caribbean coral and fish communities. Conservation Biology, 13, 888-897. [ Links ]
Hernández-Cruz, L. R., Purkis, S. J., & Riegl, B. M. (2006). Documenting decadal spatial changes in seagrass and Acropora palmata cover by aerial photography analysis in Vieques, Puerto Rico: 1937-2000. Bulletin of Marine Sciences, 79, 401-414. [ Links ]
Hernández-Delgado, E. A., Rosado-Matías, B.J., & Sabat, A. M. (2006). Management failures and coral decline threatens fish functional groups recovery patterns in the Luis Peña Channel No-Take Natural Reserve, Culebra Island, PR. Proceedings from the Gulf & Caribbean Fisheries Institute, 57, 577-605. [ Links ]
Hernández-Delgado, E. A., Suleimán, S., Olivo, I., Fonseca, J., & Lucking, M. A. (2011). Alternativas de baja tecnología para la rehabilitación de los arrecifes de coral. 178-186. In Seguinot-Barbosa, J (ed.). Islas en Extinción: Impactos Ambientales en las Islas de Puerto Rico. Cataño, PR: Ediciones SM. [ Links ]
Hernández-Delgado, E. A., Ramos-Scharrón, C. E., Guerrero, C., Lucking, M. A., Laureano, R., Méndez-Lázaro, P. A., & Meléndez-Díaz, J. O. (2012).Long-term impacts of tourism and urban development in tropical coastal habitats in a changing climate: Lessons learned from Puerto Rico. 357-398. In Kasimoglu, M. (ed.), Visions from Global Tourism Industry-Creating and Sustaining Competitive Strategies. Puerto Rico: Intech Publications. [ Links ]
Hernández-Pacheco, R., Hernández-Delgado, E. A., & Sabat, A. M. (2011). Demographics of bleaching in the Caribbean reef-building coral Montastraea annularis. Ecosphere 2(1), 1-13. doi:10.1890/ES10-00065.1
Links ]">
IDEA (Instituto de Estudio Ambiental). (1970). Informe sobre Culebra del Instituto de Estudio Ambiental. San Juan, Puerto Rico: Coelgio de Abogados. [ Links ]
Jones, G. P., McCormick, M. I., Srinivasan, M., & Eagle, J. V. (2004). Coral decline threatens fish biodiversity in marine reserves. Proceedings of the National Academic Sciences USA, 101, 8251-8253. [ Links ]
Kendall, M. S., & Eschelbach, K. A. (2006). Spatial analysis of the benthic habitats within the limited-use zones around Vieques. Puerto Rico. Bulletin Marine Science, 79, 389-400. [ Links ]
Kohler, K. E., & Gill, S. M. (2006). Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Computing and Geoscience, 32, 1259-1269. [ Links ]
Lindahl, U. (2003). Coral reef rehabilitation through transplantation of staghorn corals: effects of artificial stabilization and mechanical damages. Coral Reefs, 22, 217-223. [ Links ]
Macintyre, I. G., Raymond, B., & Stuckenrath, R. (1983). Recent history of a fringing reef, Bahia Salina del Sur, Vieques Island, Puerto Rico. Atoll Research Bulletin, 268, 1-9. [ Links ]
McGarrity, C. J., & Deslarzes, K. J. P. (2006). Preliminary data on juvenile corals and coral maladies on reefs off civilian and former military locations at the islands of Culebra and Vieques, Puerto Rico. Proceedings from the 10th International Coral Reef Symposium, 1160-1164. [ Links ]
McManus, J. W., Reyes, Jr R. B., & Nañola, Jr C. L. (1997). Effects of some destructive fishing methods on coral cover and potential rates of recovery. Environmental Management, 21, 69-78. [ Links ]
Miller, J., Muller, E., Rogers, C., Waara, R., Atkinson, A., Whelan, K. R. T., Patterson, M., & Witcher, B. (2009). Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs, 28, 925-937. [ Links ]
Pet-Soede, L., & Erdmann, M. V. (1998). Blast fishing in Southwest Sulawesi, Indonesia. April-June: 4-9.Naga: Center for Living Aquatic Resources Management Quarterly. [ Links ]
Pielou, E. C. (1966). The measurement of diversity in different types of biological collections. Journal of Theoretical Biology, 13, 131-144. [ Links ]
Porter, J. W. (2000). The effects of naval bombardment on the coral reefs of Isla Vieques, Puerto Rico. Atlanta, Georgia: King and Spaulding. [ Links ]
Porter, J. W., Barton, J. V., & Torres, C. (2011). Ecological, radiological, and toxicological effects of naval bombardment on the coral reefs of Isla de Vieques, Puerto Rico. pp 65-122. In Warfare Ecology. Netherlands: Springer. [ Links ]
Pratchett, M. S., Munday, P. L., Wilson, S. K., Graham, N. A. J., Cinner, J. E., Bellwood, D. R., Jones, G. P., Polunin, N. V. C., & McClanahan, T. R. (2008). Effects of climate-induced coral bleaching on coral-reef fishes – Ecological and economic consequences. Oceanography Marine Biology Annual Review, 46, 251-296. [ Links ]
Pratchett, M. S., Hoey, A. S., & Wilson, S. K. (2014). Reef degradation and the loss of critical ecosystem goods and services provided by coral reef fishes. Current Opinion on Environmental Sustainability, 7, 37-43. [ Links ]
Ramos-Scharrón, C. E., Amador, J. M., & Hernández-Delgado, E. A. (2012). An interdisciplinary erosion mitigation approach for coral reef protection – A case study from the eastern Caribbean. 127-160. In Cruzado, A. (Ed.). Marine Ecosystems. InTech Publications. [ Links ]
Raymond, W. (1978). Marine sediment of a Naval bombing range. Fort Lauderdale, FL: Ocean Research and Survey, Inc. [ Links ]
Raymond, W. F., & Dodge, R. E. (1980). 1979 hurricane damage to coral reefs of Vieques. Report to the Commander, Atlantic Division, Naval Facilities Engineering Command, Norfolk, Virginia. [ Links ]
Raymundo, L. J., Maypa, A. P., Gomez, E. D., & Cadiz, P. (2007). Can dynamite-blasted reefs recover? A novel, low-tech approach to stimulating natural recovery in fish and coral populations. Marine Pollution Bulletin, 54, 1009-1019. [ Links ]
Antonius, A., & Weiner, A. (1982). Coral reefs under fire. Marine Ecology Progress Series, 3, 255-277. [ Links ]
Bauer, L. J., Menza, C., Foley, K. A., & Kendall, M. S. (2008). An ecological characterization of the marine resources of Vieques, Puerto Rico. Part I: Historical data synthesis. Prepared by National Centers for Coastal Ocean Science (NCCOS) Biogeography Branch in cooperation with the Office of Response and Restoration. Silver, Spring, MD.: NOAA Technical Memorandum NOS NCCOS 86. [ Links ]
Bauer, L. J., & Kendall, M. S. (eds.). (2010). An Ecological Characterization of the Marine Resources of Vieques, Puerto Rico Part II: Field Studies of Habitats, Nutrients, Contaminants, Fish, and Benthic Communities. Silver, Spring, MD.: NOAA Tech. Memorandum NOS NCCOS 110. [ Links ]
Bohnsack, J. A., & Bannerot, S. P. (1986). A stationary census technique for quantitatively assessing community structure of coral reef fishes. Silver, Spring, MD.: NOAA Technical Report NMFS 41. [ Links ]
Bohnsack, J. A., & Harper, D. E. (1988). Length-weight relationships of selected marine reef fishes from the Southeastern United States and the Caribbean. Silver, Spring, MD.: NOAA Technical Memorandum NMFS-SEFC-215. [ Links ]
Bowden-Kerby, A. (1997). Coral transplantation in sheltered habitats using unattached fragments and cultured colonies. Proceedings from the 8th International Coral Reef Symposium, 2, 2063-2068. [ Links ]
Carrera-Rodríguez, C. J. (1978). Evaluación sobre las consecuencias de las actividades militares llevadas a cabo en Vieques, P.R. por las Fuerzas Armadas de los EE.UU. de América en los sistemas marinos del este de Vieques, con atención principal a los arrecifes de coral. Report to the Department of Natural Resources, San Juan, Puerto Rico. [ Links ]
Clarke, K. R., & Warwick, R. M. (2001). Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd Ed. Plymouth Marine Laboratory, UK: PRIMER-E, Ltd. [ Links ],
Deslarzes, K. J. P., Nawojchik, R., Evans, D. J., McGarrity, C. J., & Gehring, P. (2006). The condition of fringing reefs off former military training areas at Isla de Culebra and Isla de Vieques, Puerto Rico: preliminary results. Proceedings from the 10th International Coral Reef Symposium, Japan, 1152-1159. [ Links ]
Dodge, R. E. 1981. Growth characteristics of reef-building corals within and external to a naval ordnance range: Vieques, Puerto Rico. Proceedings from the 4th International Coral Reef Symposium, 2, 241-248. [ Links ]
DON (U.S. Department of the Navy). (1979). Draft Environmental Impact Statement for the Continued Use of the Atlantic Fleet Weapons Training Facility Inner Range (Vieques). Prepared by TAMS (Tippetts, Abbett, McCarthy, Stratton) and Ecology and Environment, Inc. [ Links ]
DON (U.S. Department of the Navy). (1980). Final Environmental Impact Statement for the Continued Use of the Atlantic Fleet Weapons Training Facility Inner Range (Vieques). Prepared by TAMS (Tippetts, Abbett, McCarthy, Stratton) and Ecology and Environment, Inc. [ Links ]
DON (U.S. Department of the Navy). (1986). Environmental Assessment of Continued Use of the Atlantic Fleet Weapons Training Facility Inner Range, Vieques, Puerto Rico. Prepared by Ecology and Environment, Inc. [ Links ]
Edmunds, P. J. (2013). Decadal-scale changes in the community structure of coral reefs of St. John, US Virgin Islands. Marine Ecology Progress Series, 489, 107-123. [ Links ]
Edmunds, P. J., & Elahi, R. (2007). The demographics of a 15-year decline in cover of the Caribbean reef coral Montastraea annularis. Ecological Monographs, 77, 3-18. [ Links ]
Edmunds, P. J., Ross, C. L. M., & Didden, C. (2011). High, but localized recruitment of Montastraea annulalris complex in St. John, United States Virgin Islands. Coral Reefs, 30, 123-130
Evans, D. J., Nawojchik, R., & Deslarzes, K. J. P. (2006). The status of coral reef fish populations off former military ranges at the islands of Culebra and Vieques, Puerto Rico: preliminary data. Proceedings from the 10th International Coral Reef Symposium, 1105-1109. [ Links ]
Fox, H. E., Pet, J. S., Dahuri, R., & Caldwell, R. L. (2002). Coral reef restoration after blast fishing in Indonesia. Proceedings from the 9th International Coral Reef Symposium, 2, 969–976. [ Links ]
Fox, H. E., Pet, J. S., Dahuri, R., & Caldwell, R. L. (2003). Recovery in rubble fields: long-term impacts of blast fishing. Marine Pollution Bulletin, 46, 1024-1031. [ Links ]
Fox, H. E., & Caldwell, R. L. (2006). Recovery from blast fishing on coral reefs: A tale of two scales. Ecol. Appl., 16, 1631-1635. [ Links ]
GMI (Geo-Marine, Inc.). (2003). Reef ecosystem baseline assessment survey and monitoring, Vieques Island, Naval Station Roosevelt Roads, Puerto Rico. Prepared for Atlantic Division, Naval Facilities Engineering Command, Norfolk, Virginia. [ Links ]
GMI (Geo-Marine Inc.). (2005). An assessment of the condition of coral reefs off the former Navy bombing ranges at Isla de Culebra and Isla de Vieques, Puerto Rico. Prepared for the Department of Defense, Legacy Resource Management Program, Arlington, Virginia, and US Army Corps of Engineers, Huntsville, Alabama. [ Links ]
Goenaga, C. (1986). Los arrecifes costaneros en Puerto Rico: Estado actual e implicaciones sociales. Science-Ciencia, 13, 78-91. [ Links ]
Goenaga, C. (1991). The state of coral reefs in the wider Caribbean. Interciencia, 15, 12-20. [ Links ]
Graham, N. A. J., Wilson, S. K., Jennings, S., Polunin, N. V. C., Bijoux, J. P., & Robinson, J. (2006). Dynamic fragility of oceanic coral reef ecosystems. Proceedings of the National Academic Sciences. USA, 103, 8425-8429. [ Links ]
Graham, N. A. J., Cinner, J. E., Norström, A.V., & Nyström, M. (2014). Coral reefs as novel ecosystems: embracing new futures. Current Opinion in Environment Sustainability, 7, 9-14. [ Links ]
Hawkins, J. P., Roberts, C. M., Van’t Hof, T., De Meyer, K., Tratalos, J., & Aldam, C. (1999). Effects of recreational scuba diving on Caribbean coral and fish communities. Conservation Biology, 13, 888-897. [ Links ]
Hernández-Cruz, L. R., Purkis, S. J., & Riegl, B. M. (2006). Documenting decadal spatial changes in seagrass and Acropora palmata cover by aerial photography analysis in Vieques, Puerto Rico: 1937-2000. Bulletin of Marine Sciences, 79, 401-414. [ Links ]
Hernández-Delgado, E. A., Rosado-Matías, B.J., & Sabat, A. M. (2006). Management failures and coral decline threatens fish functional groups recovery patterns in the Luis Peña Channel No-Take Natural Reserve, Culebra Island, PR. Proceedings from the Gulf & Caribbean Fisheries Institute, 57, 577-605. [ Links ]
Hernández-Delgado, E. A., Suleimán, S., Olivo, I., Fonseca, J., & Lucking, M. A. (2011). Alternativas de baja tecnología para la rehabilitación de los arrecifes de coral. 178-186. In Seguinot-Barbosa, J (ed.). Islas en Extinción: Impactos Ambientales en las Islas de Puerto Rico. Cataño, PR: Ediciones SM. [ Links ]
Hernández-Delgado, E. A., Ramos-Scharrón, C. E., Guerrero, C., Lucking, M. A., Laureano, R., Méndez-Lázaro, P. A., & Meléndez-Díaz, J. O. (2012).Long-term impacts of tourism and urban development in tropical coastal habitats in a changing climate: Lessons learned from Puerto Rico. 357-398. In Kasimoglu, M. (ed.), Visions from Global Tourism Industry-Creating and Sustaining Competitive Strategies. Puerto Rico: Intech Publications. [ Links ]
Hernández-Pacheco, R., Hernández-Delgado, E. A., & Sabat, A. M. (2011). Demographics of bleaching in the Caribbean reef-building coral Montastraea annularis. Ecosphere 2(1), 1-13. doi:10.1890/ES10-00065.1
Links ]">
IDEA (Instituto de Estudio Ambiental). (1970). Informe sobre Culebra del Instituto de Estudio Ambiental. San Juan, Puerto Rico: Coelgio de Abogados. [ Links ]
Jones, G. P., McCormick, M. I., Srinivasan, M., & Eagle, J. V. (2004). Coral decline threatens fish biodiversity in marine reserves. Proceedings of the National Academic Sciences USA, 101, 8251-8253. [ Links ]
Kendall, M. S., & Eschelbach, K. A. (2006). Spatial analysis of the benthic habitats within the limited-use zones around Vieques. Puerto Rico. Bulletin Marine Science, 79, 389-400. [ Links ]
Kohler, K. E., & Gill, S. M. (2006). Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Computing and Geoscience, 32, 1259-1269. [ Links ]
Lindahl, U. (2003). Coral reef rehabilitation through transplantation of staghorn corals: effects of artificial stabilization and mechanical damages. Coral Reefs, 22, 217-223. [ Links ]
Macintyre, I. G., Raymond, B., & Stuckenrath, R. (1983). Recent history of a fringing reef, Bahia Salina del Sur, Vieques Island, Puerto Rico. Atoll Research Bulletin, 268, 1-9. [ Links ]
McGarrity, C. J., & Deslarzes, K. J. P. (2006). Preliminary data on juvenile corals and coral maladies on reefs off civilian and former military locations at the islands of Culebra and Vieques, Puerto Rico. Proceedings from the 10th International Coral Reef Symposium, 1160-1164. [ Links ]
McManus, J. W., Reyes, Jr R. B., & Nañola, Jr C. L. (1997). Effects of some destructive fishing methods on coral cover and potential rates of recovery. Environmental Management, 21, 69-78. [ Links ]
Miller, J., Muller, E., Rogers, C., Waara, R., Atkinson, A., Whelan, K. R. T., Patterson, M., & Witcher, B. (2009). Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs, 28, 925-937. [ Links ]
Pet-Soede, L., & Erdmann, M. V. (1998). Blast fishing in Southwest Sulawesi, Indonesia. April-June: 4-9.Naga: Center for Living Aquatic Resources Management Quarterly. [ Links ]
Pielou, E. C. (1966). The measurement of diversity in different types of biological collections. Journal of Theoretical Biology, 13, 131-144. [ Links ]
Porter, J. W. (2000). The effects of naval bombardment on the coral reefs of Isla Vieques, Puerto Rico. Atlanta, Georgia: King and Spaulding. [ Links ]
Porter, J. W., Barton, J. V., & Torres, C. (2011). Ecological, radiological, and toxicological effects of naval bombardment on the coral reefs of Isla de Vieques, Puerto Rico. pp 65-122. In Warfare Ecology. Netherlands: Springer. [ Links ]
Pratchett, M. S., Munday, P. L., Wilson, S. K., Graham, N. A. J., Cinner, J. E., Bellwood, D. R., Jones, G. P., Polunin, N. V. C., & McClanahan, T. R. (2008). Effects of climate-induced coral bleaching on coral-reef fishes – Ecological and economic consequences. Oceanography Marine Biology Annual Review, 46, 251-296. [ Links ]
Pratchett, M. S., Hoey, A. S., & Wilson, S. K. (2014). Reef degradation and the loss of critical ecosystem goods and services provided by coral reef fishes. Current Opinion on Environmental Sustainability, 7, 37-43. [ Links ]
Ramos-Scharrón, C. E., Amador, J. M., & Hernández-Delgado, E. A. (2012). An interdisciplinary erosion mitigation approach for coral reef protection – A case study from the eastern Caribbean. 127-160. In Cruzado, A. (Ed.). Marine Ecosystems. InTech Publications. [ Links ]
Raymond, W. (1978). Marine sediment of a Naval bombing range. Fort Lauderdale, FL: Ocean Research and Survey, Inc. [ Links ]
Raymond, W. F., & Dodge, R. E. (1980). 1979 hurricane damage to coral reefs of Vieques. Report to the Commander, Atlantic Division, Naval Facilities Engineering Command, Norfolk, Virginia. [ Links ]
Raymundo, L. J., Maypa, A. P., Gomez, E. D., & Cadiz, P. (2007). Can dynamite-blasted reefs recover? A novel, low-tech approach to stimulating natural recovery in fish and coral populations. Marine Pollution Bulletin, 54, 1009-1019. [ Links ]
Riegl, B. (2001). Degradation of reef structure, coral and fish communities in the Red Sea by ship groundings and dynamite fisheries. Bulletin of Marine Science, 69, 595-611. [ Links ]
Riegl, B., & Luke, K. E. (1999). Ecological parameters of dynamited reefs in the northern Red Sea and their relevance to reef rehabilitation. Marine Pollution Bulletin, 37, 488-498. [ Links ]
Riegl, B., Moyer, R. P., Walker, B. K., Kohler, K., Gilliam, D., & Dodge, R. E. (2008). A tale of germs, storms, and bombs: Geomorphology and coral assemblage structure at Vieques (Puerto Rico) compared to St. Croix (U.S. Virgin Islands). Journal of Coastal Research, 24, 1008-1021. [ Links ]
Roberts, C. M., & Ortnond, R. F. G. (1987). Habitat complexity and coral reef fish diversity and abundance on Red Sea fringing reefs. Marine Ecology Progress Series, 41, 1-8. [ Links ]
Rogers, C. S., Cintrón, G., & Goenaga, C. (1978). The impact of military operations on the coral reefs of Vieques and Culebra. Report to the Department of Natural Resources, San Juan, Puerto Rico. [ Links ]
Rogers, C. S., Fitz, III H. C., Gilnack, M., Beets, J., & Hardin, J. (1984). Scleractinian coral recruitment patterns at Salt River Submarine Canyon, St. Croix, U.S. Virgin Islands. Coral Reefs, 3, 69-76. [ Links ]
Rosa-Serrano, N. (1996). Impacto ambiental del bombardeo de la Marina en el area este de la Isla de Vieques. B.Sc. Thesis, Environmental Sciences Program, University of Puerto Rico, San Juan, PR. [ Links ]
Shannon, C. E., & Weaver, W. (1948). The Mathematical Theory of Communication. Urbana, IL.: University of Illinois Press. [ Links ]
Riegl, B., & Luke, K. E. (1999). Ecological parameters of dynamited reefs in the northern Red Sea and their relevance to reef rehabilitation. Marine Pollution Bulletin, 37, 488-498. [ Links ]
Riegl, B., Moyer, R. P., Walker, B. K., Kohler, K., Gilliam, D., & Dodge, R. E. (2008). A tale of germs, storms, and bombs: Geomorphology and coral assemblage structure at Vieques (Puerto Rico) compared to St. Croix (U.S. Virgin Islands). Journal of Coastal Research, 24, 1008-1021. [ Links ]
Roberts, C. M., & Ortnond, R. F. G. (1987). Habitat complexity and coral reef fish diversity and abundance on Red Sea fringing reefs. Marine Ecology Progress Series, 41, 1-8. [ Links ]
Rogers, C. S., Cintrón, G., & Goenaga, C. (1978). The impact of military operations on the coral reefs of Vieques and Culebra. Report to the Department of Natural Resources, San Juan, Puerto Rico. [ Links ]
Rogers, C. S., Fitz, III H. C., Gilnack, M., Beets, J., & Hardin, J. (1984). Scleractinian coral recruitment patterns at Salt River Submarine Canyon, St. Croix, U.S. Virgin Islands. Coral Reefs, 3, 69-76. [ Links ]
Rosa-Serrano, N. (1996). Impacto ambiental del bombardeo de la Marina en el area este de la Isla de Vieques. B.Sc. Thesis, Environmental Sciences Program, University of Puerto Rico, San Juan, PR. [ Links ]
Shannon, C. E., & Weaver, W. (1948). The Mathematical Theory of Communication. Urbana, IL.: University of Illinois Press. [ Links ]
1. University of Puerto Rico, Center for Applied Tropical Ecology and Conservation, Coral Reef Research Group, PO Box 23360, San Juan, PR 00931-3360; fax: 1-787-764-2610; edwin.hernandezdelgado@gmail.com, omsjulio@hotmail.com, alfredo.a.montanez@gmail.com, abimarie07@gmail.com
2. Sociedad Ambiente Marino, PO Box 22158, San Juan, PR 00931-2158; samuelsuleiman@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}