










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.3 San José Sep. 2014
Population assessment of Acropora palmata (Scleractinia: Acroporidae): relationship between habitat and reef associated species
Evaluación poblacional de Acropora palmata (Scleractinia: Acroporidae): relación entre el hábitat y especie asociados
Evaluación poblacional de Acropora palmata (Scleractinia: Acroporidae): relación entre el hábitat y especie asociados
Abstract
Three decades ago, Acropora palmata was one of the main reef-building coral species throughout the Caribbean, forming an essential component of the structural complexity of shallow coral reef habitats. These colonies still provide microhabitats for settlement, food and shelter to many vertebrates and invertebrates. The recent decline of A. palmata has been followed by a significant loss in spatial heterogeneity and possibly in species diversity. Studies addressing whether dead and living stands of Acropora hold different fish and benthic assemblages are scarce. The status of Acropora colonies and their associated species were assessed in October 2012, at two reef zones of Cayo Sombrero, Venezuela. Visual censuses of fish abundance and the number of macrofaunal individuals were recorded for both live and dead zones. Living Acropora colonies had the lowest abundance (˂31%). In both zoned the fish community was dominated by damselfishes (˂53%) and wrasses (˂36%), the benthic macrofauna by peracarid crustaceans (˂40%) and polychaetes (˂38%). Fish and benthic communities were not correlated with the condition (live or dead) of the Acropora habitats; possibly branching structures provide the necessary shelter and protection no matter if they are dead or alive. More replication is necessary to test this unexpected result.
Key words: Acropora palmata, benthic macrofauna, Caribbean, coral reefs, reef fishes.
Resumen
Hace tres décadas, Acropora palmata fue una de las principales especies de coral constructoras de arrecifes a lo largo del Caribe, formando un componente esencial de la complejidad estructural disponible en los hábitats arrecifales someros. Estas colonias todavía proveen microhábitats para el asentamiento, alimentación y refugio de muchas especies de vertebrados e invertebrados. El declive reciente de A. palmata ha sido precedido por una pérdida significativa de heterogeneidad espacial y se piensa que ésta reducción ha producido una disminución importante en la diversidad de especies. Los estudios señalan que las colonias muertas o vivas de Acropora que sostienen diferentes comunidades de bentos y de peces son escasas. El estado de las colonias de Acropora y sus especies asociadas fueron evaluadas en Octubre de 2012, en dos zonas arrecifales diferentes de Cayo Sombrero, Venezuela. Censos visuales de la abundancia de especies de peces y el número de individuos de la macrofauna fueron también registrados para ambas zonas, tanto vivas como muertas. Las colonias vivas presentaron la menor abundancia (˂31%). En ambos sitios la comunidad de peces fue dominada por los peces damiselas (˂53%) y lábridos (˂36%); la macrofauna bentónica por crustáceos peracáridos (˂40%) y poliquetos (˂38%). Las comunidades de peces y bentos no mostraron correlación con la condición (vivo o muerto) de los hábitats de Acropora; posiblemente las estructuras ramificadas proveen el refugio y protección necesario sin importar si están vivos o muertos. Una mayor replicación es necesaria para probar este resultado inesperado.
Palabras claves: Acropora palmata, arrecifes coralinos, Caribe, macrofauna bentónica, peces arrecifales.
Coral reefs are complex systems that typically support high structural complexities with high spatial heterogeneity, representing one of the most diverse ecosystems of the planet (Glynn, 1991). Unfortunately, multiple factors such as overfishing, sedimentation, nutrient enrichment, global warming and diseases (Rogers, 1990; Aronson & Precht, 2001; Cróquer & Bone, 2003; Gómez et al., 2010) have been directly or indirectly linked to the progressive decline in coral communities, causing important changes on the coral reef structure and increasing the loss of biodiversity and live coral cover over the last few decades (García, Cróquer & Pauls, 2003).
The branching coral Acropora palmata (Lamarck, 1816) has an essential role in coral reefs as it provides the habitat for a myriad of reef-associated species, significant coral cover and adds complex topography to shallow reef environments. Therefore, this species may have an important role in determining the abundance and diversity of many reef fishes and macrofauna populations (Lirman, 1999). The robust and complex morphology of A. palmata provides microhabitats for settlement, food and shelter to many invertebrate reef species (Bonin, 2012), which depend on the presence of Acroporid colonies to survive, probably because of the geological, physical and biological support offered by these corals to numerous shallow reef communities (Baums et al., 2002). Unfortunately, A. palmata has shown an important decline of live tissue cover (˃95%) throughout the Caribbean after a mass mortality caused by the combination of white band disease and hurricanes in the middle of the 80s (Aronson & Precht, 2001). It is believed that this regional demise has reduced habitat complexity and concomitantly the diversity of the associated fauna. Yet, the scarcity of quantitative studies to test this hypothesis is evident.
In Morrocoy National Park (MNP) live stands of A. palmata are rare, despite large patches of dead colonies being common in exposed reefs with strong wave energy. Recently, Martínez and Rodríguez-Quintal (2012) recorded the presence of this branching coral in Cayo Sombrero and they found a predominance of adult and juvenile size colonies with a low impact from partial mortality, bleaching and no signs of diseases. However, factors such as sediment accumulation affected 47.5% of the colonies, followed by algae overgrowth (37.5%), damselfish territories (35%) and anchor damage (22.5%). These findings clearly indicate that this species has managed to survive for decades, despite MNP being subjected to a number of human impacts such as sedimentation, urban development and chronic pollution (Cróquer & Bone, 2003). While different studies have evaluated the structure and distribution of fish communities in MNP (Rodríguez & Villamizar, 2008), no previous comparisons between the organisms associated with live and dead A. palmata zones have ever been conducted. This paper addressed a comparison between fish and benthic assemblages associated with dead and living stands of A. palmata to test if changes in the abundance and species composition of these assemblages were correlated with the condition (dead or alive) of A. palmata.
Materials and Methods
Study site: The Morrocoy National Park (MNP) is a coastal marine park located in the west central coast of Venezuela (10o53’04’’N - 68o12’46’’W), in Golfo Triste region between Tucacas and Chichiriviche populations (Rodríguez & Villamizar, 2008). Surveys to assess the condition of A. palmata stands and the fish and benthic communities were conducted at Cayo Sombrero, a key located at Morrocoy National Park (MNP). Previous surveys indicate the presence of A. palmata to the northwest and the southeast leeward areas of this reef site (Martínez & Rodríguez-Quintal, 2012).
Acropora palmata: We determined the status of A. palmata colonies in October 2012, by counting every colony within four 10m long by 1m wide (10m2) belt transects conducted in two different zones at Cayo Sombrero. One of the zones was dominated by live A. palmata colonies and the other zone was dominated by dead stands of A. palmata. The linear coral cover of all species was recorded at each zone. All Acropora colonies were quantified and classified to determine colony size, using digital images that were analyzed with Image Tool v. 3.0, establishing coral area from each colony. Additionally, the fish and benthic macrofauna communities associated to Acropora zones were assessed in both sites.
Fish community: The coral reef fishes were surveyed in both live and dead Acropora zones using the method of Bortone, Hastings and Oglesby (1986), conducting visual census over A. palmata colonies. This visual census technique was based on the best estimation by eye of fish species abundance that were associated with live and standing dead Acropora areas.
Benthic community: Six sediment samples were collected around live and dead Acropora colonies, using a PVC 7cm diameter cylinder. The samples were preserved in 10% formalin and washed in the lab over a 0.5mm mesh sieve. The material retained on the sieve was fixed with 70% ethanol solution and sorted under a stereo microscope for counting and identification of macrofauna groups associated to A. palmata colonies. Additionally, we identified the main polychaete and peracarid families because of the abundance recorded by these two groups inside the benthic communities.
Statistical analyses: The community structure of live and dead Acropora zones were evaluated by the relative abundance and mean density of A. palmata as well as reef fishes and benthic macrofauna. Changes in the fish and macrofauna assemblages across colony condition were analyzed by Non-Metric Multidimensional scaling (nMDS) calculated from the Bray-Curtis similarity matrix, using square root transformed data. Differences in fish and macrofauna composition between colony conditions were assessed using permutation-based multivariate analysis of variance (PERMANOVA, Anderson, 2001), based on the Bray-Curtis similarity index and using 9 000 randomizations. All these analyses were performed with PRIMER v.6.0 and Statistix v.8.0.
Results
Acropora palmata: The dead Acropora zone was dominated by standing dead A. palmata colonies (75%), followed by a group of zoanthids (20%) and sand substrate (5%), while the live Acropora zone showed that the dominant category was living A. palmata (60%), followed once again by zoanthids (30%) and sand substrate (10%). A total of 62 live and dead A. palmata colonies were found and assessed within 10m2 belt transect (Table 1). Mean densities for live and dead colonies were 0.27±0.15 ind/10m2 (mean±standard deviation) and 0.61±0.52 ind/10m2, respectively. In addition, size structure was only represented by adult A. palmata with sizes varying between 1 900 and 5 500cm2 for dead colonies and between 1 800 and 3 600cm2 for live colonies.
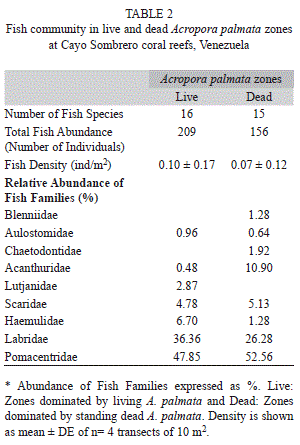
Fish community: A total of 21 species belonging to nine families with similar fish diversity between live and standing dead A. palmata colonies were recorded (Table 2). The total fish abundance consisted of 209 individuals in live A. palmata zones and 156 individuals in standing dead A. palmata. Density in the live zone was 0.10±0.17 ind/m2 and 0.07±0.12 ind/m2 for skeletons of Acropora colonies. The reef fish families in the dead zone were dominated by Pomacentridae with a relative abundance of 52.56%, followed by Labridae with 26.28%. Some families such as Acanthuridae were mainly present in dead Acropora areas. Additionally, the live Acropora zone was dominated by Pomacentridae with relative abundance of 47.85%, followed by Labridae with 36.36%, and some families such as Haemulidae and Lutjanidae recorded their highest abundance over these live colonies (Table 2).
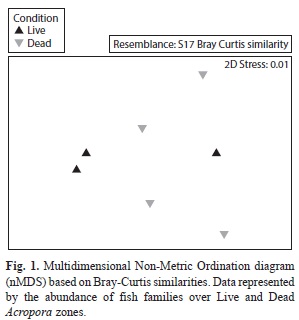
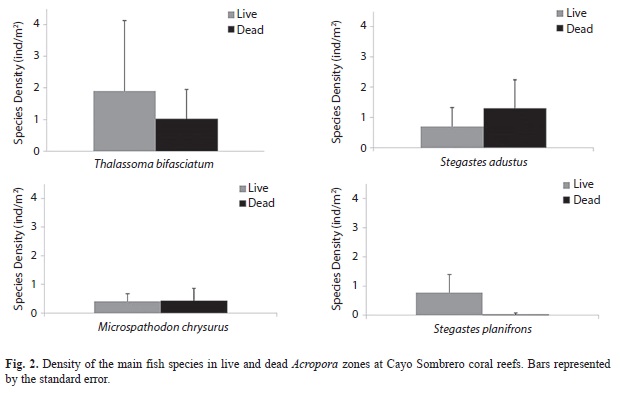
Ordination analyses (nMDS) revealed similar fish communities associated with live and dead A. palmata colonies (Fig. 1). PERMANOVA results supported the nMDS ordination and showed that fish composition was not significantly different between live and standing dead Acropora zones (PERMANOVA, Pseudo-F=0.72, p(PERM)= 0.57). On the other hand, the main fish species observed in live A. palmata were Thalassoma bisfaciatum (˂2 ind/m2), followed by Stegastes planifrons with densities ˂0.80 ind/m2 and S. adustus with ˂0.70 ind/m2 (Fig. 2). Dead Acropora zones were dominated by S. adustus (˂1.50 ind/m2), T. bisfaciatum (˂1 ind/m2) and Microspathodon chrysurus (˂0.50 ind/m2).
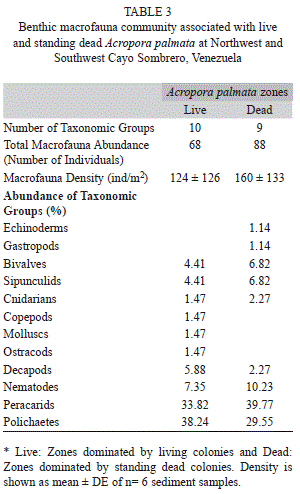
Benthic macrofauna: A total of 12 taxonomic groups were identified in the benthic macrofauna inhabiting the reef sediments, recording a similar diversity between live and dead Acropora zones (Table 3). The total abundance of the overall macrofauna was higher around dead colonies (88 individuals) compared to living colonies (68 individuals), showing the highest mean density of macrofauna in standing dead Acropora zones (160±133 ind/m2) compare to live Acropora (124±126 ind/m2). Also, the identified taxonomic groups were mostly represented by peracarid crustaceans, with relative abundance ˃33%, and polychaetes, with relative abundance ˃29% for both conditions. On the other hand, the remaining benthic macrofauna were relatively rare, comprising <11% of the total in each of the zones (Table 3).
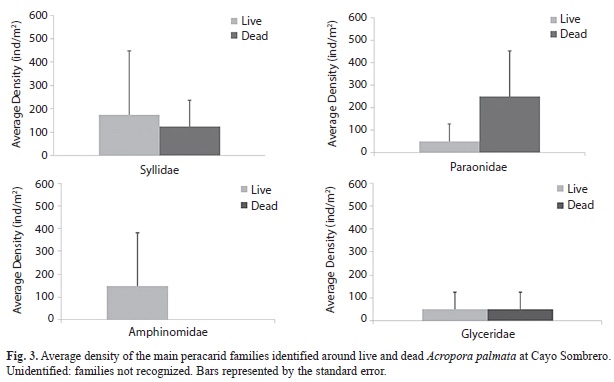
The peracarids were the most abundant group within the benthic macrofauna and they were only represented by three families, from which Isaeidae family was dominant (˃250 ind/m2) in live and dead Acropora zones, followed by Melitidae (˃50 ind/m2) and Sphaeromatidae (˃25 ind/m2) (Fig. 3). Additionally, the “unidentified” group represented those crustaceans that were unable to be classified into families; however, they belonged to the Tanaidacea and Cumacea orders, with densities ˃25 ind/m2 for live and dead zones. The nMDS ordination revealed a similar peracarid community associated with live and standing dead A. palmata condition (Fig. 4). We found no differences between assemblages present in live and dead A. palmata stands (PERMANOVA, Pseudo-F=1.15, p (PERM)= 0.39).
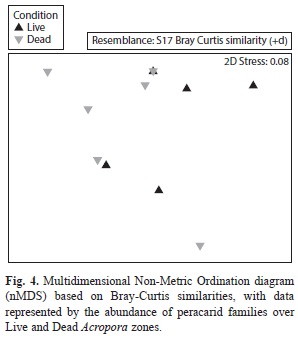
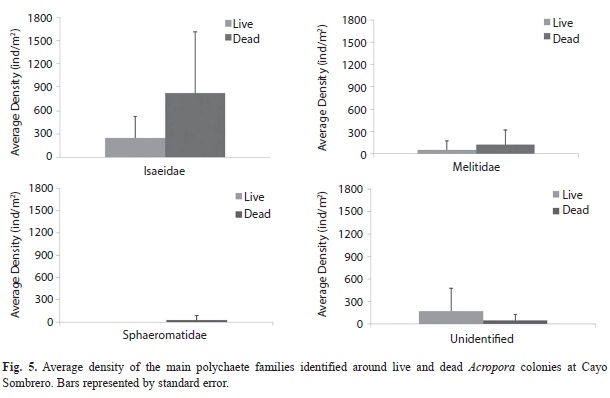
Twelve families of polychaetes were identified in the macrofauna, being dominated by Paraonidae (248±203 ind/m2) around dead Acropora colonies and Syllidae (174±273 ind/m2) around living colonies (Fig. 5). The Amphinomidae family was only present around living colonies (149±231 ind/m2), while Glyceridae reveled the same densities for both colony condition (49±77 ind/m2). As well as peracarids, the nMDS showed a similar polychaete community between live and standing dead A. palmata (Fig. 6), forming two main groups that were defined by the abundance of Syllidae and Paraonidae families. There was not significant differences between the polychaete assemblages for both conditions (PERMANOVA, Pseudo-F=1.88, p (PERM)=0.08).
Discussion
The complex structure provided by the branching species A. palmata has been reported as potential shelter for different reef associated organisms (Lemoine & Valentine, 2012), forming unique habitats with high ecological values at the Caribbean coral reefs. However, over the past few years, many reef areas in Morrocoy have experienced a significant loss of Acroporid corals, leading to the eventual formation of large areas consisting of standing dead A. palmata. Thus, live and dead stands of A. palmata in Sombrero represent two types of habitats that nearly provide the same structural complexity. Our data showed that the density for live A. palmata at Cayo Sombrero was lower than some coral reefs located at Los Roques National Park, such as Cayo Pirata, Crasquí, Carenero, Gran Roque and Cayo de Agua (˃0.69 ind/10 m2) (Zubillaga, Márques, Cróquer & Bastidas, 2008); nevertheless, many other coral reefs such as Boca de Cote, Boca del Medio, Sebastopol and Madrisquí in Los Roques had lower densities (˂0.13 indv/10 m2) than those reported in Cayo Sombrero.
Due to the significant importance of colony condition within a reef environment, we established the hypothesis that those zones dominated by dead A. palmata could be mainly composed of herbivorous species, while those zones with live A. palmata could be composed of other species such as corallivorous and benthivorous species which feed upon living tissues. Although we set all these predictions based on the colony status, the fish and benthic macrofauna community structure did not show significant differences between live and dead A. palmata. Herbivorous fishes were the most abundant condition in both Acropora zones, being dominated by the Pomacentridae family which is typically associated with A. palmata colonies regardless its status. Species in this family are known to defend their territory which includes a shelter hole, grazing area and nest site, showing at the same time some specificity in their relationship with live and dead colonies (Chasqui-Velasco, Alvarado, Acero & Zapata, 2007).
The dominant species recorded at Cayo Sombrero were T. bisfasciatum, S. adustus, M. chrysurus and S. planifrons for both live and dead A. palmata zones. These fishes have also been reported by Rodríguez and Villamizar (2008), who found similar results at the same study site, mentioning that the abundance of T. bisfasciatum and S. adustus could be positively correlated with the substrate type, being mainly associated with dead coral zones covered by algae. Also, these species are very abundant and common in exposed areas with strong wave impact and currents (López-Ordaz & Rodríguez-Quintal, 2010), as was the case in the Acropora shallow reefs at Cayo Sombrero.
Additionally, the benthic macrofauna revealed a similar richness and density compared to other coral reefs such as Madrisquí, Sebastopol and Boca del Medio at Los Roques, where it has been reported between nine and 14 macrofauna groups belonging to reefs sediment around Montastraea faveolata colonies, with densities ˃500 ind/m2 (Bone & López-Ordaz, 2009). Also, the benthic macrofauna did not show significant differences between live and dead Acropora zone, however, we observed that live zones harbored a low density of benthic organisms compared to the dead zone. This low density could be the result of benthivorous fishes which dominate live Acropora reefs, and have a feeding preference for the macrofauna associated with living coral sediments. On the opposite side, the extensions of standing dead A. palmata often harbor herbivorous fishes that only consume the primary producers, leading to the establishment of benthic macrofauna and increasing its abundance around dead colonies.
The most important taxonomic group in terms of abundance was represented by peracarid crustaceans, which are characteristic of the calcareous sediment macrofauna, presenting a spatial distribution that is determinate by several environmental factors such as temperature, chlorophyll concentration and sediment salinity (Escobar-Briones & Jiménez-Guadarrama, 2010). Moreover, the main peracarid families identified around A. palmata colonies were Isaeidae and Melitidae, belonging to the amphipod group, which are often related to coral reef sediments and usually record a high abundance and diversity because of the breeding success and gregarious patterns presented by these families (Winfi & Escobar–Briones, 2007). Besides, the live Acropora area showed the lowest density of peracarid families, possibly because of the presence of benthivorous fishes that include in their diet a large variety of crustaceans (Randall, 1967).
As well as crustaceans, the polychaete worms were also one of the most abundant groups of macrofauna, which typically present both spatial and temporal variations due to the community natural oscillations that occurs through the life cycles of these benthic species (Bone & López-Ordaz, 2009 unpublished). Some polychaete families such as Amphinomidae were only present around living A. palmata colonies, most likely because of the diet preferences. These worms are common in living coral reef zones, being considered as important predators of sessile organisms such as sponges, corals and anemones (Yánez-Rivera, 2009).
A. palmata colonies may function as a physical complex structure, providing shelter and protection for fish and benthic macrofauna communities that are associated with these branching colonies in Cayo Sombrero coral reefs. Although we observed some community variations between live and dead Acropora zones, statistically we did not find significance differences between both conditions, thus, the reef associated species are unlikely related to whether or not live tissue is present, but instead the absent or presence of the physical structure of A. palmata could be the key factor that enables the establishment of different species associated with reef environments.
Acknowledgments
We are grateful to the Marine Community Lab of the Universidad Simón Bolívar for giving the necessary equipments and instruments to support this project. We also appreciate the assistant of I. Ortega for the taxonomic knowledge and the field assistant provided by C. Brett and A. Guerra during this research. Thanks to the CEO (Centro de Estudios Oceanográficos) of the Universidad Simón Bolívar for providing lab facilities during the field trips.
References
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Australian Ecology, 26, 32-46. [ Links ]
Aronson, R. B., & Precht, W. F. (2001). White-band disease and the changing face of Caribbean coral reefs. Hydrobiologia, 460, 25-38. [ Links ]
Baums, I., Bythell, J., Lirman, D., Gladfelter, E., Miller, M., Mueller, E., Ortiz, A., Rueda, P., Vargas, B., Vermeij, M., Vollmer, S., & Weil, E. (2002). Report from the Biology and Ecology Working Group. In Bruckner, A. W. (ed). Proceedings of the Caribbean Acropora workshop: potential application of US endangered species act as a conservation strategy NOAA. Technical Memorandum NMFS-OPR-24, Silver Spring, MD. [ Links ]
Bone, D., & López-Ordaz, A. (2009). Programa de monitoreo ecológico, ecotoxicológico y ambiental de arrecifes coralinos en el Parque Nacional Los Roques comunidades bentónicas asociadas a los sedimentos. Caracas, Venezuela: Unpublished Report. [ Links ]
Bonin, M. C. (2012). Specializing on vulnerable habitat: Acropora selectivity among damselfish recruits and the risk of bleaching-induced habitat loss. Coral Reefs, 31, 287–297. [ Links ]
Bortone, S., Hastings, R. & Oglesby, J. (1986). Quantification of reef fish assemblages: a comparison of several in situ methods. Northeast Gulf Science, 8, 1-22. [ Links ]
Chasqui-Velasco, L., Alvarado, E., Acero, P., & Zapata, F. (2007). Efecto de la herbivoría y coralivoría por peces en la supervivencia de corales trasplantados en el Caribe colombiano. Revista de Biologiá Tropical, 55, 825-837. [ Links ]
Cróquer, A., & Bone, D. (2003). Las enfermedades en corales escleractínidos: ¿Un nuevo problema en el arrecifede Cayo Sombrero, Parque Nacional Morrocoy, Venezuela? Revista de Biologiá Tropical, 4, 167-172. [ Links ]
Escobar-Briones, E., & Jiménez-Guadarrama, L. (2010). Macrocrustáceos (Peracarida, Decapoda) de fondos carbonatados del sector occidental del banco de Campeche en el sur del golfo de México. Revista Mexicana de Biodiversidad, 81, 63-72. [ Links ]
García, A., Cróquer, A., & Pauls, S. (2003). Estado actual de las enfermedades y otros signos de deterioro coralino en siete arrecifes del Parque Nacional Archipiélago de Los Roques, Venezuela. Revista de Biologiá Tropical, 51, 173-180. [ Links ]
Gómez, J. C., Ricaurte, C., Mayo, G., Orejarena, J., Díaz, J. M., & Andrade, C. A. (2010). Cobertura de los fondos y paisajes bentónicos asociados a formaciones diapíricas en los Bancos de Salmedina, Plataforma Continental del Caribe Colombiano. Bologiá Invemar, 39, 117-135. [ Links ]
Glynn, P. (1991). Coral reef bleaching in the 1980s and possible connections with global warming. Trends in Ecology and Evolution, 6, 175-179. [ Links ]
Lemoine, N. P., & Valentine, J. F. (2012). Structurally complex habitats provided by Acropora palmata influence ecosystem processes on a reef in the Florida Keys National Marine Sanctuary. Coral Reefs, 31, 779-786. [ Links ]
Lirman, D. (1999). Reef fish communities associated with Acropora palmata: relationships to benthic attributes. Bulletin of Marine Science, 65, 235-252. [ Links ]
López-Ordaz, A., & Rodríguez-Quintal, J.G. (2010). Ictiofauna asociada a un arrecife somero en el Parque Nacional Morrocoy, Venezuela. Revista de Biologiá Tropical, 58, 163-174. [ Links ]
Martínez, K., & Rodríguez-Quintal, J.G. (2012). Caracterización de las colonias de Acropora palmata (Scleractinia: Acroporidae) en Cayo Sombrero, Parque Nacional Morrocoy, Venezuela. Bological Institute of Oceanography, Venezuela, 51, 67-74. [ Links ]
Randall, J. E. (1967). Food habits of reef fishes of the West Indies. Studies in Tropical Oceanography, 5, 665-847. [ Links ]
Rodríguez, J., & Villamizar, E. 2008. Estructura de la comunidad íctica en dos arrecifes con diferentes grados de afectación en el Parque Nacional Morrocoy, Venezuela. Acta Biologia Venezuela, 28, 61-69. [ Links ]
Rogers, C. S. (1990). Responses of coral reefs and reef organisms to sedimentation. Marine Ecology Progres Series, 62, 185-202. [ Links ]
Winfi, I., & Escobar–Briones, E. (2007). Anfípodos (Crustacea: Gammaridea) del sector norte del Mar Caribe: listado faunístico, registros nuevos y distribución espacial. Revista Mexicana de Biodiversidad, 78, 51- 61. [ Links ]
Yánez-Rivera, B. (2009). Amphinomidae Savigny in Lamarck, 1818. p.p. 77-87. In De León-González, J. A., Bastidas-Zavala, J. R., Carrera-Parra, L. F., García-Garza, M. E., Peña-Rivera, A., Salazar- Vallejo, S. I., & Solís-Weiss, V. (Eds). Poliquetos (Annelida: Polychaeta) de México y América Tropical., México: Universidad Autónoma de Nuevo León, Monterrey. [ Links ]
Zubillaga, A. L., Márques, L. M., Cróquer, A., & Bastidas, C. (2008). Ecological and genetic data indicate recovery of the endangered coral Acropora palmata in Los Roques, Southern Caribbean. Coral Reefs, 27, 63-72. [ Links ]
Aronson, R. B., & Precht, W. F. (2001). White-band disease and the changing face of Caribbean coral reefs. Hydrobiologia, 460, 25-38. [ Links ]
Baums, I., Bythell, J., Lirman, D., Gladfelter, E., Miller, M., Mueller, E., Ortiz, A., Rueda, P., Vargas, B., Vermeij, M., Vollmer, S., & Weil, E. (2002). Report from the Biology and Ecology Working Group. In Bruckner, A. W. (ed). Proceedings of the Caribbean Acropora workshop: potential application of US endangered species act as a conservation strategy NOAA. Technical Memorandum NMFS-OPR-24, Silver Spring, MD. [ Links ]
Bone, D., & López-Ordaz, A. (2009). Programa de monitoreo ecológico, ecotoxicológico y ambiental de arrecifes coralinos en el Parque Nacional Los Roques comunidades bentónicas asociadas a los sedimentos. Caracas, Venezuela: Unpublished Report. [ Links ]
Bonin, M. C. (2012). Specializing on vulnerable habitat: Acropora selectivity among damselfish recruits and the risk of bleaching-induced habitat loss. Coral Reefs, 31, 287–297. [ Links ]
Bortone, S., Hastings, R. & Oglesby, J. (1986). Quantification of reef fish assemblages: a comparison of several in situ methods. Northeast Gulf Science, 8, 1-22. [ Links ]
Chasqui-Velasco, L., Alvarado, E., Acero, P., & Zapata, F. (2007). Efecto de la herbivoría y coralivoría por peces en la supervivencia de corales trasplantados en el Caribe colombiano. Revista de Biologiá Tropical, 55, 825-837. [ Links ]
Cróquer, A., & Bone, D. (2003). Las enfermedades en corales escleractínidos: ¿Un nuevo problema en el arrecifede Cayo Sombrero, Parque Nacional Morrocoy, Venezuela? Revista de Biologiá Tropical, 4, 167-172. [ Links ]
Escobar-Briones, E., & Jiménez-Guadarrama, L. (2010). Macrocrustáceos (Peracarida, Decapoda) de fondos carbonatados del sector occidental del banco de Campeche en el sur del golfo de México. Revista Mexicana de Biodiversidad, 81, 63-72. [ Links ]
García, A., Cróquer, A., & Pauls, S. (2003). Estado actual de las enfermedades y otros signos de deterioro coralino en siete arrecifes del Parque Nacional Archipiélago de Los Roques, Venezuela. Revista de Biologiá Tropical, 51, 173-180. [ Links ]
Gómez, J. C., Ricaurte, C., Mayo, G., Orejarena, J., Díaz, J. M., & Andrade, C. A. (2010). Cobertura de los fondos y paisajes bentónicos asociados a formaciones diapíricas en los Bancos de Salmedina, Plataforma Continental del Caribe Colombiano. Bologiá Invemar, 39, 117-135. [ Links ]
Glynn, P. (1991). Coral reef bleaching in the 1980s and possible connections with global warming. Trends in Ecology and Evolution, 6, 175-179. [ Links ]
Lemoine, N. P., & Valentine, J. F. (2012). Structurally complex habitats provided by Acropora palmata influence ecosystem processes on a reef in the Florida Keys National Marine Sanctuary. Coral Reefs, 31, 779-786. [ Links ]
Lirman, D. (1999). Reef fish communities associated with Acropora palmata: relationships to benthic attributes. Bulletin of Marine Science, 65, 235-252. [ Links ]
López-Ordaz, A., & Rodríguez-Quintal, J.G. (2010). Ictiofauna asociada a un arrecife somero en el Parque Nacional Morrocoy, Venezuela. Revista de Biologiá Tropical, 58, 163-174. [ Links ]
Martínez, K., & Rodríguez-Quintal, J.G. (2012). Caracterización de las colonias de Acropora palmata (Scleractinia: Acroporidae) en Cayo Sombrero, Parque Nacional Morrocoy, Venezuela. Bological Institute of Oceanography, Venezuela, 51, 67-74. [ Links ]
Randall, J. E. (1967). Food habits of reef fishes of the West Indies. Studies in Tropical Oceanography, 5, 665-847. [ Links ]
Rodríguez, J., & Villamizar, E. 2008. Estructura de la comunidad íctica en dos arrecifes con diferentes grados de afectación en el Parque Nacional Morrocoy, Venezuela. Acta Biologia Venezuela, 28, 61-69. [ Links ]
Rogers, C. S. (1990). Responses of coral reefs and reef organisms to sedimentation. Marine Ecology Progres Series, 62, 185-202. [ Links ]
Winfi, I., & Escobar–Briones, E. (2007). Anfípodos (Crustacea: Gammaridea) del sector norte del Mar Caribe: listado faunístico, registros nuevos y distribución espacial. Revista Mexicana de Biodiversidad, 78, 51- 61. [ Links ]
Yánez-Rivera, B. (2009). Amphinomidae Savigny in Lamarck, 1818. p.p. 77-87. In De León-González, J. A., Bastidas-Zavala, J. R., Carrera-Parra, L. F., García-Garza, M. E., Peña-Rivera, A., Salazar- Vallejo, S. I., & Solís-Weiss, V. (Eds). Poliquetos (Annelida: Polychaeta) de México y América Tropical., México: Universidad Autónoma de Nuevo León, Monterrey. [ Links ]
Zubillaga, A. L., Márques, L. M., Cróquer, A., & Bastidas, C. (2008). Ecological and genetic data indicate recovery of the endangered coral Acropora palmata in Los Roques, Southern Caribbean. Coral Reefs, 27, 63-72. [ Links ]
1. Universidad Simón Bolívar, Departamento Biología de Organismos, Postgrado en Ciencias Biológicas, Caracas, Venezuela; kimberlym2004@hotmail.com
2. Universidad Simón Bolívar, Departamento Biología de Organismos, Caracas, Venezuela.
3. Universidad Simón Bolívar, Departamento Estudios Ambientales, Caracas, Venezuela.
4. Universidad Central de Venezuela, Instituto de Zoología Tropical, Postgrado en Ecología, Caracas, Venezuela.
Received 01-IX-2013 Corrected 31-III-2014 Accepted 01-IV-2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}