










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.3 San José Sep. 2014
The status of coral reefs and associated fishes and invertebrates of commercial importance in Pedro Bank, Jamaica
Estado de los arrecifes de coral y comunidades de peces e invertebrados con interés comercial asociadas al Banco de Pedro, Jamaica
Estado de los arrecifes de coral y comunidades de peces e invertebrados con interés comercial asociadas al Banco de Pedro, Jamaica
Abstract
The coral reefs located off the north coast of the Jamaican mainland are some of the best and most studied reefs in the world. Coral reefs of Pedro Bank, Jamaica were assessed in March, 2012 as part of the KSLOF Global Reef Expedition using a modified Atlantic and Gulf Rapid Reef Assessment (AGRRA) protocol. The main objectives were to: 1) characterize the distribution, structure and health of coral reefs; and 2) evaluate the population status of commercially important reef fishes and invertebrates. This work was conducted to assist in characterizing coral reef habitats within and outside a proposed fishery reserve, and identify other possible conservation zones. Within 20 reefs, live coral cover ranged from 4.9% to 19.2%. Coral communities were dominated by small corals (esp. Agaricia, Porites and Siderastrea) although many sites had high abundances of large colonies of Montastraea annularis and M. faveolata, and these were generally in good condition. A single area, within the proposed fishery reserve, had extensive Acropora cervicornis thickets, and several shallow locations had small, but recovering A. palmata stands. Macroalgal cover at all sites was relatively low, with only three sites having greater than 30% cover; crustose coralline algae (CCA) was high, with eight sites exceeding 20% cover. Fish biomass at all sites near the Cays was low, with a dominance of herbivores (parrotfish and surgeonfish) and a near absence of groupers, snappers and other commercially important species. While parrotfish were the most abundant fish, these were all extremely small (mean size= 12cm; <4% over 29cm), and they were dominated by red band parrotfish (Sparisoma aurofrenatum) followed by striped parrotfish (Scarus iseri). While coral communities remain in better condition than most coastal reefs in Jamaica, intense fishing pressure using fish traps (main target species: surgeonfish) and hookah/spear fishing (main target: parrotfish) is of grave concern to the future persistence of these reefs. The proposed fishery reserve encompasses some of the best coral reef habitat near the Cays, but this MPA should be expanded to encompass other habitats and MPAs should be considered for bank reefs at the northwestern end, as well as Banner Reef and Blowers Rock.
Key words: Pedro Bank, Jamaica, coral reef health, fish community structure, marine protected areas.
Resumen
Los arrecifes de coral ubicados en la costa norte de Jamaica son los mejores y mas estudiados corales del mundo. Los corales de arrecife del Banco de Pedro, Jamaica, fueron evaluados en marzo 2012 como parte de la expedición global KSLOF utilizando una modificación del protocolo de estimación rápida de arrecifes para el Atlántico y el Caribe (AGRRA). Los objetivos principales fueron: 1) caracterizar la distribución, estructura y salud de los arrecifes de coral; y 2) evaluar la condición de las poblaciones de peces arrecifales e invertebrados de interés comercial. Este trabajo fue realizado con el propósito de colaborar en la caracterización de hábitats arrecifales, tanto dentro como fuera de un área sugerida como reserva marina, e identificar otras áreas de conservación. Para los 20 arrecifes, la cobertura de coral vivo varió entre 4.9% y 19.2%. Las comunidades de corales son dominadas por corales pequeños (Agaricia, Porites y Siderastrea), aunque en varias localidades se presentó una mayor abundancia de colonias grandes de M. annularis y M. faveolata, y estos se encontraban por lo general en buenas condiciones. Dentro de la reserva pesquera propuesta, una sola área presentó fragmentos de A. cervicornis, y varias localidades someras presentaron pequeños fragmentos de A. palmata, pero en proceso de recuperación. La cobertura de macroalgas en todas las áreas fue relativamente baja, solo tres lugares presentaron más de un 30%; la cobertura de algas coralinas calcárea (CCA) fue alta, ocho lugares excedieron el 20%. La biomasa de peces fue baja en todos los sitios cercanos a los cayos, y hubo dominancia de herbívoros (pez loro y cirujanos), y en general ausencia de chernas, pargos y otras especies de peces de interés comercial. Aunque los peces loro fueron los más abundantes, estos presentaron tamaños extremadamente pequeños (tamaño promedio = 12cm; <4% por encima de 29cm), y estuvieron dominados por el pez loro de banda roja (Sparisoma aurofrenatum), y por el pez loro de rayas (Scarus iseri). Mientras que las comunidades de coral permanecen en mejor condición que muchos de los arrecifes costeros de Jamaica, existe una intensa presión pesquera usando trampas de pesca (principal objetivo: cirujanos) y de compresores de aire (hookah)/ pesca con arpón (principal objetivo: pez loro) es preocupante, y compromete la persistencia y el futuro de estos arrecifes. La reserva marina (MPA) que se sugiere incluye algunos de los mejores hábitats de arrecife coralino cerca de los cayos, pero es recomendable que se amplíe para incluir otros hábitats y que se consideren áreas marinas protegidas en los bancos de arrecife del extremo noroeste, así como el arrecife Banner y Blowers Rock.
Palabras clave: Banco de Pedro, Jamaica, salud del arrecife de coral, estructura comunitaria de peces, áreas marinas protegidas.
The coral reefs located off the north coast of the Jamaican mainland are some of the best and most studied reefs in the world (Hughes, 1994). In contrast, very few studies have been conducted on the banks located off the south coast of Jamaica. The first and only comprehensive coral reef assessment was conducted on a portion of Pedro Bank in 2005 (Kramer, 2006). Other studies have focused on fisheries, socioeconomics and terrestrial assessments (Nicholson & Hartsuijter, 1982; Munro, 1983; Koslow, Hanley & Wicklund, 1988; Espuet, 2006; Hay, 2006; Kramer, 2006). These studies have documented the progressive expansion of human populations on the Cays during the 1990s, and their effects on the reefs and associated resources. The importance of the Pedro Bank as a base for fishing has steadily increased as Jamaica’s nearshore fisheries continue to decline. The influx of fishermen, together with the adoption of more efficient and destructive fishing techniques such as Antillean Z-traps and hookah spearfishing is leading to the overexploitation of lobster, conch, and finfish throughout Pedro Bank (Aiken & Kong, 2000; Koslow et al., 1988; Munro, 1983; Nicholson & Hartsuijker, 1982). Further, the increase in permanent inhabitants on Pedro Cays has resulted in destruction of coastal vegetation, excessive trash and marine debris, discharge of untreated sewage into surrounding waters, and diminished habitat for nesting turtles and seabirds (Espeut, 2006; Hay, 2006).
Since the mid-2000s, conservation agencies in Jamaica have been working to implement the Pedro Bank Coral Reef Management Project. This project has included biological and socioeconomic assessments, feasibility studies, and efforts to control and minimize overfishing and degradation of coral reefs and coral cays due to unsustainable human settlement. A primary goal has involved the development of a zoning strategy, including the adoption of a fishery reserve off southwest Cay. To help inform constituents of the value of these resources and the benefits that would ensue through establishment of networks of marine protected areas (MPAs), KSLOF and partners conducted an extensive survey of coral reef habitats within Pedro Bank. The current research used the Atlantic and Gulf Rapid Reef Assessment (AGRRA) protocol, which is the same methodology applied in 2005 (Kramer, 2006). The areas assessed in 2005 were reevaluated during this study, and we expanded the surveys to new locations. Here we present data on the status of coral reefs and changes that have occurred since 2005, and provide recommendations on potential sites that would benefit from protection as MPAs.
Materials and Methods
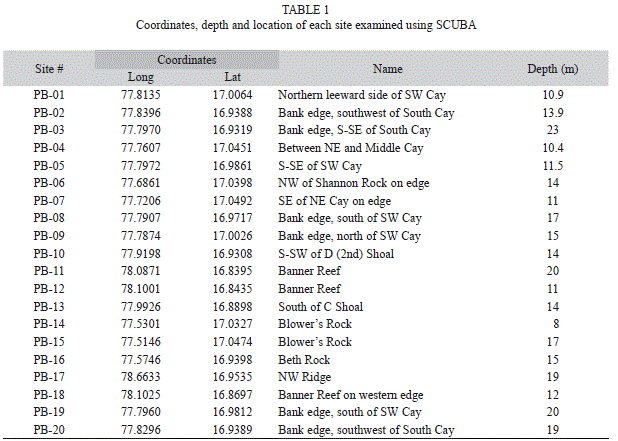
Study sites: Pedro Bank is a submerged bank rising abruptly from about 500m depth. It is located about 58km off Jamaica at its closest point (Portland Point), and roughly 98km from Kingston, Jamaica. It extends over an area of 8 040km2 and has a circumference of roughly 590km. Much of the bank is relatively shallow (about 10-24m depth) with extensive seagrass beds, rubble and sandy habitats, and scattered patch reefs. The bank gradually deepens in a NW direction. The south and southeast sides of the bank face into the Caribbean current and have the best developed reefs. The coral reef community structure was assessed at 20 sites along the S/SE sides, within 18 fore reef locations and two patch reefs (Table 1). At each site, data were collected on benthic community structure, corals, fishes and motile invertebrates using the Atlantic and Gulf Rapid Reef Assessment (AGRRA) protocol.
Benthic cover: Cover of benthic organisms was estimated using a minimum of six 10m long transects per site (158 transects total) and one to three 10m x 1m photo-transects. The organism and substrate type were recorded every 10cm for a total of 100 points per transect. Substrates were categorized as hard-ground, rubble, sand/silt, live coral, and dead coral. All corals were identified to species and recorded as live, bleached, recently dead or long dead. Other invertebrates were identified to the lowest taxonomic level possible. Algae were divided into five functional groups (fleshy macroalgae, erect coralline algae, crustose coralline algae, turf algae, cyanobacteria). Additional measurements of algal height were recorded for macroalgae.
Coral: Coral species diversity, abundance, size structure and health were assessed with 74 belt transects, each 1 x 10m in length. Each coral within the 10m2 area, 4cm or larger in diameter, was identified, measured (length, width and height) and assessed. Visual estimates of tissue loss were recorded for each colony using a 1m bar marked in 1cm increments for scale. If the coral exhibited tissue loss, estimates of the amount of remaining tissue and percent that died were made based on the entire colony surface. Tissue loss was categorized as recent mortality (white skeleton, occurring within the last 1-5 days), transitional mortality (exposed skeleton with filamentous green algae and diatom colonization, 6-30 days) and old mortality (>30 days). For each coral with partial or whole colony mortality, the cause of mortality was identified if possible. The diagnosis included an assessment of the type of disease, extent of bleaching, predation, competition, overgrowth or other causes of mortality. Each coral was first carefully examined to identify cryptic predators. Lesions were diagnosed into four categories: recent tissue loss, skeletal damage, color change, and unusual growth patterns; an individual colony could have multiple characteristics (e.g. color change and recent tissue loss). The location (apical, basal, medial) and pattern of tissue loss (linear, annular, focal, multifocal, and coalescing) were recorded, and a field name (e.g. black band disease, yellow band disease, dark spots disease) was assigned.
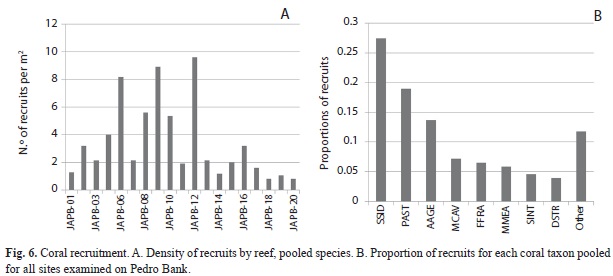
The abundance of recruits (corals smaller than 4cm) was assessed using a minimum of five 0.25m2 quadrats per transect, with each quadrat located at fixed, predetermined intervals (2, 4, 6, 8, 10m, alternating between right and left side of the transect line). Recruits were identified in both benthic and coral transects. Recruits were divided into two categories: <2cm diameter and 2-3.9cm diameter.
Motile invertebrates: The abundance of large motile invertebrates (crustaceans, molluscs and echinoderms) was quantified using random circular plots, each 10m diameter (total area = 314m2), with 1-4 plots assessed per reef. One diver would extend a 10m line from a central point and then swim slowly in a circle while a second diver would record the numbers of each species of invertebrate seen within the plot. Additional roving surveys were undertaken to assess presence of invertebrates in cryptic areas, including ledges and caves. Invertebrates were also counted and recorded within coral belt transects.
Reef fish: The reef fish assemblage (approximately 70 species) was quantified within a minimum of six 30 x 2m belt transects per site (187 transects total). All species were identified and their size was estimated to the nearest 5cm using a T-bar marked in 5cm increments for scale. The assessment focused on species that are ecologically relevant to the health of reefs and also important for commercial or recreational fisheries, with emphasis on herbivores, invertivores and larger piscivores. Roving surveys were also undertaken to characterize species diversity.
Data analysis: All data were initially entered into Microsoft Excel spreadsheets with PRIMER software used for graphical and comparative analysis. Species diversity, richness and evenness was calculated using the Shannon – Weiner index (H`): [H` = - Σi pi (log pi)]; Margalef’s species richness (d): d = (S - 1)/ log (N), where S= number of species; N = number of individuals; and Pielou’s evenness (J`): J` = H`/ log (S) to determine how evenly individuals were distributed among different species.
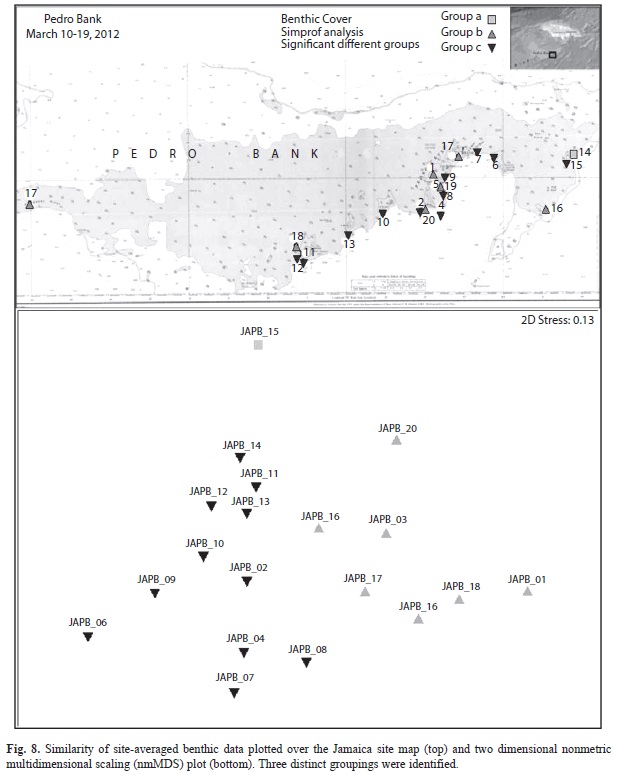
Clustering of benthic data, coral composition and fish biomass/abundance by site was examined using multi-dimensional scaling (MDS) followed by similarity profiles (SIMPROF) analysis to determine the factors that contribute most to a particular grouping. Individual contribution (e.g. fish taxa, fish functional groups, coral species) to the similarity of resulting groups was estimated using the similarity percentage (SIMPER) analysis of untransformed datasets. SIMPER analysis results were visually inspected and biotopes were determined based on similarities, dissimilarities, taxa distribution and relative abundance (semi-quantitative data). Analysis of Similarity (ANOSIM) testing was also employed to evaluate relationships between benthic and fish attributes. These tests compare sites based upon ranked, species similarity measures. Coral species abundances were log transformed to create a Bray-Curtis dissimilarity matrix (d). ANOSIM results were graphically interpreted using non-metric, multi-dimensional scaling (MDS) (Clarke & Warwick, 2001; Clarke & Gorley, 2006).
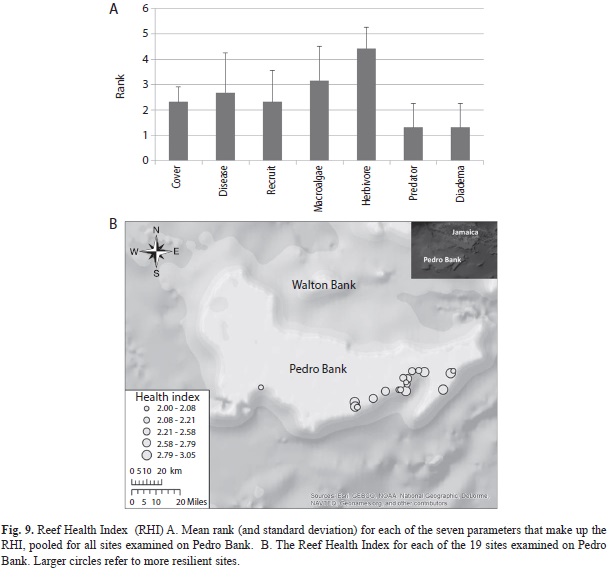
A Reef Health Index (RHI) was calculated for each dive site using seven specific biological indicators assessed during the field surveys. The grades were calculated by converting the mean for each indicator into a rank of 1 (critical) to 5 (very good). Seven parameters, grouped into two categories, were used: a Coral Index, comprised of coral cover, coral disease prevalence and coral recruitment; and a Reef Biota Index, comprised of a macroalgal index, herbivorous fish abundance (parrotfish and surgeon fish only), commercial fish abundance (grouper and snapper only), and Diadema abundance. The ranked scores of the three Coral measures and the four Reef Biota measures and these two sub-indices were then averaged to calculate an integrated reef health index. Threshold values for each rank were based on data ranges presented in the Healthy Reef Initiative report (HRI, 2008).
Results
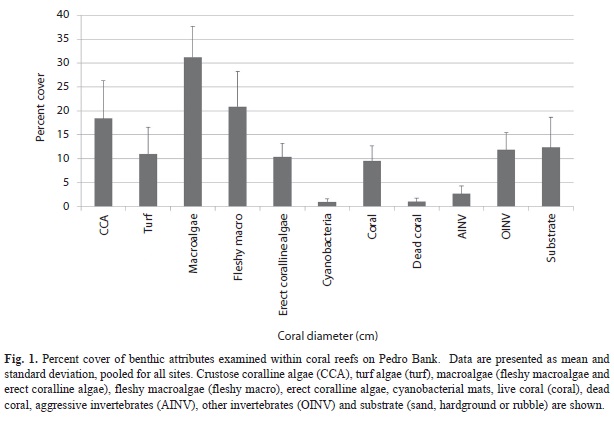
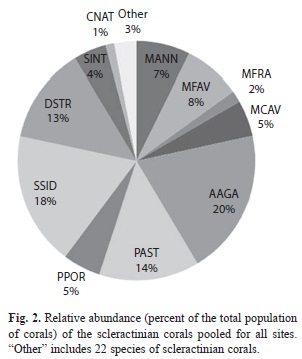
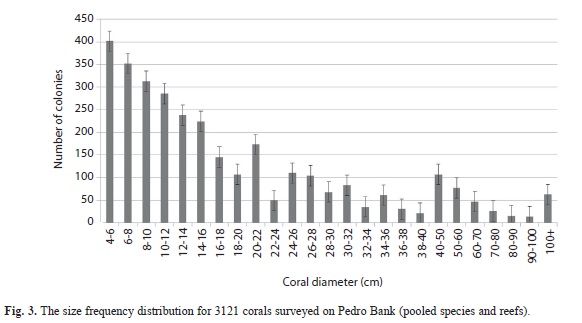
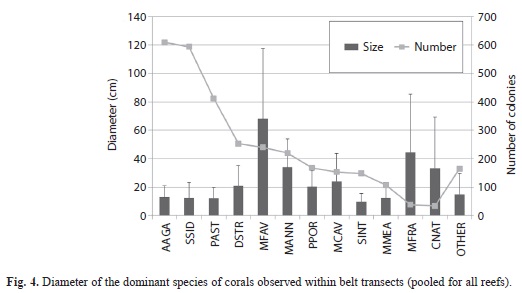
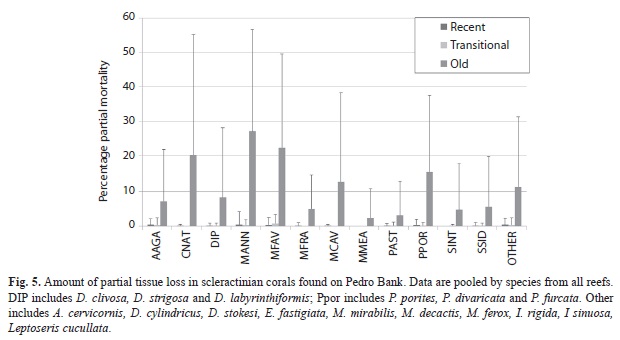
Benthic community structure: Within coral reef habitats, over 60% of the bottom was covered by algae, most of which was fleshy and erect coralline macroalgae (31%) and crustose coralline algae (18%). Live coral cover ranged from 4.9% (PB-01) to 19.2% (PB-20). Mean cover of other invertebrates was 14.5%, while 12.3% of the bottom consisted of uncolonized substrate (sand, rubble or hardground); 1% was dead coral (Fig. 1). Sites contained a total of 33 species of scleractinian corals. The most abundant corals were Agaricia, Siderastrea, Porites and the Montastraea annularis complex, respectively (Fig. 2). Most corals were small (mean diameter= 20cm), with 30% of all colonies 21cm or larger and 2% of the corals 100cm or larger (Fig. 3). The only corals with a mean diameter that exceeded 20cm were the M. annularis complex, M. cavernosa, Colpophyllia natans, Dendrogyra cylindricus, Acropora palmata and A. cervicornis (Fig. 4). A single taxon, M. faveolata, was dominated by colonies that exceeded 50cm. All corals exhibited a very low percent partial mortality (mean= 10%) and virtually no recent or transitional mortality (<1%). The greatest amount of partial tissue loss was observed in the largest corals, including C. natans (20%), M. annularis (22%) and M. faveolata (27%) (Fig. 5). Coral recruits were dominated by S. siderea (27%), P. astreoides M. cavernosa (7%), Favia fragum (6.5%) and Meandrina meandrites (5.8%), with all species (pooled) occurring at a density of 3.3/m2 (Fig. 6).
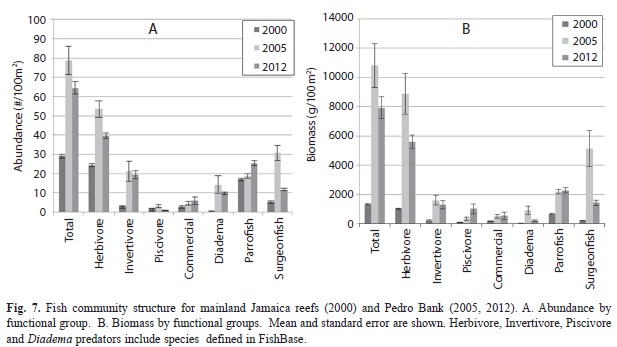
Reef fish communities: Reef fish populations exhibited a fairly low diversity (116 species), abundance (65 fish/100m2), size (mean=13cm) and biomass (9 430g/100m2). Populations were dominated by herbivores, with parrotfish occurring at the highest density (25fish/100m2). Sparisoma aurofrenatum (redband parrotfish) and Scarus iseri (striped parrotfish) were most abundant, but all parrotfish were exceptionally small (mean=12cm total length) including terminal phase fish (18% were 21-29cm and 3% were 31-40cm). Other abundant species were surgeonfish (11fish/100m2), wrasses (9fish/100m2), and grunts (6fish/100m2); most other functional groups had a density of <1 fish/100m2.
All species of fish were small; over 78% were less than 20cm and only 3.5% were over 30cm. Overall biomass of reef fishes was low; herbivores had the greatest biomass, at 5 500g/100m2 followed by invertivores (2 965g/100m2). Triggerfish contributed most to the biomass of invertivores, with the abundance of these species increasing with distance from the Cays. Many functional groups of fishes were rare or absent including all snappers, large serranids (populations consisted only of hinds and graysby), barracuda, morays, grunts, and angelfish. Commercially significant species showed the lowest numbers and biomass overall, emphasizing the heavy fishing pressure occurring on Pedro Bank.
Although fish populations remain healthier on Pedro Bank than that observed off mainland Jamaica, fish community structure has shown substantial changes since 2 005 surveys (Fig. 7). Most importantly, the abundance and biomass of surgeonfishes has declined quite substantially. Parrotfish abundances are slightly higher than that recorded in 2005, but the biomass for the two time periods is virtually the same, suggesting the average size of individual fish has declined over this period. In sites closest to the fishing village (0 to 10mi) the total biomass declined by 36% and the biomass of herbivores declined by 46% between 2005 and 2012. The biomass also markedly increased at a distance of 10mi from the fishing village in 2005, while the biomass in 2012 showed no increase except at distances of 20mi or more from the Cays.
Motile invertebrates: Commercially important (Panulirus, Strombus) and ecologically important motile invertebrates (Diadema, sea cucumbers, large crabs, octopus) were present in extremely low abundances. Queen conch were observed on 9 reefs at a very low density, with slightly higher abundances in six locations (0.1animal/m2). This is not necessarily indicative of the population size on Pedro Bank, however, as key Strombus habitats were not examined. Lobsters were rarely observed on reefs, occurring at 12 sites at densities of 2lobster/100m2); much higher numbers were seen in landings examined on shore. Prominent Diadema populations occurred in two locations (12-20/100m2); low densities (<0.5/100m2) in other sites suggests they have shown limited recovery since the die-off in the 1980s.
Drivers of community structure: With exception of a few sites, dominant taxa were similar in size structure and abundance, and the condition of the substrate and health of corals showed minimal differences. Sites could be delineated into three groups based on benthic attributes, with macroalgae, crustose coralline algae (CCA) and non-living substrate contributing most to the differences (Fig. 8). Sites also subdivided into eight groups based on the contribution of different fish taxa to biomass, with groupings differentiated mostly due to relative amounts of parrotfish, surgeonfish and/or triggerfish. Most other variables were fairly homogeneous between locations. For instance, coral cover, fish abundance and biomass, and Diadema abundance were uniformly low among all sites. Analysis of Similarity (ANOSIM) testing did not reveal any significant relationships between coral abundance and macroalgal cover (R2=0.155, p=0.161); coral abundance was related to fish biomass (all species pooled), but the R-value was very low (R2=0.249; p=0.006). Fish abundance (all species pooled) was not related to coral cover (R2=0.011; p=0.444) or vertical relief (height between the substrate and the tops of the corals; R2=0.059; p=0.778), but it was related to macroalgal cover, although the R value again was low (R2=0.428; p=0.013). Herbivorous fish populations (biomass and abundance of pooled species, parrotfish and surgeonfish) did not appear to be related to algae (CCA, fleshy macroalgae or turf algae) or coral (cover or abundance) populations. The lack of identifiable drivers of community structure may be due to the fact that all sites were overfished, coral cover was uniformly low, and algal communities were fairly homogeneous.
Resilience and health of reefs: The sites examined in this study exhibited Reef Health Index (RHI) scores that varied from 1-3 (critical to fair condition) for seven parameters (coral cover, coral recruitment, coral disease, fleshy algae biomass, herbivore abundance (Diadema, parrotfishes and surgeonfishes), and commercially important reef fish) and a cumulative RHI of 2 to 3.5 (Fig. 9).
Discussion
Pedro Bank is a unique area that supports a wide variety of shallow marine habitats including seagrass beds, gorgonian hardgrounds, rubble fields, deep algal meadows, sand flats and coral reefs. Coral reef habitats occupied only a small percentage of the bank overall, and were mostly concentrated at the eastern and southeastern edge of the bank, adjacent to the drop-off. The new fishery reserve contains some of the healthiest coral reefs, including extensive shallow Acropora palmata framework and the largest healthy stand of A. cervicornis found on Pedro Bank. Other important high relief coral habitats and seagrass beds were adjacent but outside of the reserve boundaries and around Banner Rock, Blowers Rock and D and C shoal.
In all locations, coral cover was fairly low (<20%) and dominated by small corals, especially early colonizing species (Agaricia and Porites) as well as Siderastrea, Diploria, and Meandrina. Of note were several populations of larger massive framework corals such as Montastraea annularis (complex), as well as stands of Madracis mirabilis, and numerous large Dendrogyra colonies. Montastraea annularis (complex), once the most important frame builders in the Western Atlantic, has declined throughout its range since the mid-1990s, primarily from recent bleaching events and disease (white plague and yellow band disease) (Miller, Atkinson & Witcher, 2009; Bruckner, 2012a). These corals formerly made up 50-80% of the live coral cover, but recently have been reduced to 2-5% cover; what were once large (several meter) colonies are now skeletons colonized by other corals, algae and invertebrates, with small M. annularis tissue remnants. On Pedro Bank, most of the larger M. annularis and M. faveolata colonies were completely ‘live”. Very little disease, bleaching and recent mortality was noted on these species. Extensive Acropora framework with scattered live colonies surrounded many of the cays and emergent shoals, and small patches of A. cervicornis were found on many of the deeper reefs (10-30m depth). Acroporid corals have become rare in most Caribbean locations as a result of white band disease outbreaks in the 1980s, recent bleaching events, storm damage and a host of other factors and were recently listed on the U.S. Endangered Species Act (Bruckner, 2003). The lack of disease, few corallivores and presence of small colonies with little mortality on Pedro Bank is indicative of ongoing recovery of these species.
One of the most positive findings among the stony corals examined on Pedro Bank was the low level of partial mortality. AGRRA surveys that have been conducted throughout the Caribbean over the last 15 years (Lang, 2003; Bruckner & Hill, 2009; Bruckner, 2012b), as well as the Caribbean surveys completed during the Global Reef Expedition, have recorded a mean of about 20-30% partial mortality on all species pooled and upwards of 50-80% partial mortality in M. annularis complex and other larger massive corals. On Pedro Bank most colonies were intact; partial mortality was usually less than 10% and colonies of M. annularis (complex) showed about 20% partial mortality. The presence of healthy, undamaged colonies may be an indication of higher resilience, better environmental conditions and a greater potential for successful reproduction, as corals have not been reduced below the minimal size threshold necessary for gametogenesis (Szmant-Froelich, 1985; Szmant, 1991).
The benthos also appeared to be in fairly good condition. Even though Diadema were rare in most locations, and cover of macroalgae exceeded the cover of living coral, there was a general impression that fleshy macroalgae was less abundant than that reported from many other Caribbean localities (Lang 2003). The substrate had a prominent cover of crustose coralline algae, with small patches of erect coralline algae and some macroalgae, most of which were concentrated at the margins of corals and not on open reef substrates. Unlike other areas in the Caribbean, the reef was not being smothered by thick mats of Dictyota, Microdictyon (completely absent) and Lobophora (occurred in low abundances), only on a few corals showed partial mortality, and cyanobacterial mats were rare.
The largest concern to Pedro Bank reefs is overexploitation of the fishery resources. Exploitation of Pedro Bank began in the 1960s, but has progressively increased as the densities of humans on the cays increased (Munro, 1983). Most fishing effort is concentrated on the southeast (SE) portions of the bank, near the cays, which is also the location of some of the best developed coral reefs. As a result, the biomass of reef fish was lowest surrounding the fishing village with higher biomass (mainly invertivores) seen at distances of 20mi or further from the village. This is probably related to the distance that a fisherman can realistically travel from the fishing village. Fishers tend to go to their farthest point (under ten miles in one day) and then work their way back, fishing at different spots as they return (Meggs pers. comm.).
Intensive fishing pressure appears to have affected most species as all reefs had a fairly low biomass of fishes and many key species were extremely rare or absent. The current targets of the fishery appeared to be two families of herbivores which made up the bulk of the catch, scaridae and acanthuridae. Very few schools of surgeonfish were documented and individuals were small (5-15cm) and large-bodied scarids were infrequently seen. In addition, the dominant parrotfish on these reefs (and in the catch) was the red band parrotfish followed by stoplight parrotfish. While parrotfish were still fairly abundant, both species were very small (most <15cm). These species were changing sex from initial phase (females) to terminal phase (males) at a much smaller size than that seen historically (TP fish of 10cm were fairly common). The smaller than average size in sex change may be a survival mechanism that is a direct outcome of high fishing pressure (Hawkins & Roberts, 2003). However, the decline in the average size of the population will cause a disproportionate decline in the reproductive output, because there is an exponential relationship between body length and egg production (Bohnsack, 1990), which may have negative consequences on the long term persistence of these fish.
Two main fishery approaches are now undertaken on Pedro Bank, trap (pot) fishing and spearfishing using hookah (Aiken & Kong, 2000; Gustavson, 2002). The Antillean z-traps were observed on or adjacent (in sand) to coral reef habitats in many of the sites we surveyed. Traps typically contained a very low biomass of fish, but the fish consisted of a broad spectrum including piscivores (small serranids, lutjanids, grunts, balistids, pomacanthids), as well as herbivores (scarids), but surgeonfish were the dominant species noted. These traps seemed to favor smaller species and individuals (partially because no large fish remain), which may contribute to growth overfishing. Further, the depletion of large predatory fishes has triggered a shift from high value species to a high diversity of lower value species, especially species that formerly were considered non-target species and are important indicators of a healthy reef system (e.g. butterflyfish, angelfish); this is suggestive of ecosystem overfishing (Pauly, 1979). As traps have become less economical, there has been expanded use of hookah, primarily targeting parrotfish with incidental catch of other “large” fish and lobsters, which may lead to Malthusian overfishing (Pauly, 1990). A continued decline of herbivores will also cause ancillary damage as reef substrates become monopolized by increasing amounts of fleshy algae, reducing the area available for encrusting coralline algae, and inhibiting coral settlement. Over the longer term, the reefs will be less likely to resist future disturbances and their potential to rebound may be reduced.
At this time, the overall condition of Pedro Bank’s coral reefs remains far better than most reefs off mainland Jamaica, but these reefs are at a tipping point. The establishment of the South West Cay Special Fishery Conservation Area is a key step forward in conservation of Pedro Bank’s coral reef resources, as it will allow fish to grow larger, potentially producing more offspring and spill over into adjacent fished areas. In particular it can help increase the abundance and size of parrotfish which represent the primary target of fishermen and concurrently species responsible for one of the most important processed in maintaining ecological balance, herbivory. The newly established, measuring 16.3 km2, is far too small to adequately protect fishery resources as this represents <0.1% of the total area (9 247km2; Zenny, 2006) of Pedro Bank. Furthermore, the increasing fishing effort and destructive types of fishing are unsustainable and are likely to trigger deleterious changes to the reefs. Additional conservation measures that emphasize an expanded network of protected areas may help maintain and restore Pedro banks precious coral reefs.
Recommendations
Significant benefits could be achieved by expanding the boundaries of the newly established Fishery Reserve such that it encompasses extensive and healthiest high relief Montastraea dominated coral habitat found to the south and west, and it includes an adjacent large seagrass bed. Banner reef should be considered a candidate site for an MPA as it contained the largest number of different marine habitats including a very diverse shallow water reef dominated by large Montastraea faveolata and M. annularis colonies intermixed with extensive stands of Porites porites, A. cervicornis patches, large colonies of Dendrogyra cylindricus and high relief coral bommies in deeper sandy areas, extensive Acropora framework, and an elongate ridge with steeply sloping sides, caves, and ledges that provided considerable habitat for large crabs, lobsters, octopus, groupers and other high value species. Banner reef also had the largest population of Diadema seen on Pedro Bank, the highest diversity of fish families, and a high biomass of fish composed of schools of grunts, parrotfishes and surgeonfishes. Blowers Rock is also another candidate MPA site. It had the largest high-relief Acropora palmata framework seen on Pedro Bank with extensive patches of live A. palmata. A very unusual Montastraea dominated reef was also identified in 15-20m depth. This reef had unusually large (2-4m diameter/height) and old Montastraea faveolata colonies, the largest seen on the bank. Because these areas are east (and upcurrent) of the inhabited Cays and the Fish Sanctuary, the endangered corals that occur on these reefs (Montastraea and Acropora) could serve as seed stock for other areas on the bank to the west. They also provide considerable habitat for other species as a result of their high relief. Finally, C Shoal and D Shoal are additional candidates for inclusion in an MPA network. It had extensive Acropora palmata framework with small stands of living A. palmata in shallow (2-5m depth) areas and extensive diverse deeper coral habitats composed of large Montastraea-dominated communities, patches of Acropora cervicornis and the only large stands of Madracis mirabilis and Porites porites seen in the region.
Acknowledgments
The data presented in this manuscript were collected during the fifth coral reef research mission completed during the Global Reef Expedition (GRE). All research was performed under a permit obtained from the National Environment and Planning Agency (NEPA) (ref #18/27, 8 December, 2011). Financial support for the surveys and the research vessel, M/Y Golden Shadow was provided by His Royal Highness Prince Khaled bin Sultan. We are grateful for the involvement of the Nature Conservancy, Jamaica, the University of West Indies, the Ministry of Agriculture & Fisheries, the National Environment and Planning Agency; and Natural Resource Management and Environmental Planning Department (NRMEP), Nova Southeastern University, Atlantic and Gulf Rapid Reef Assessment, and Reef Environmental Education Foundation. Special thanks to Judy Lang, Ken Marks, David Grenda, Alex Dempsey, Llewyn Meggs, Nathalie Zenny and Rachel D’Silva for their assistance with the coral, algae, fish and invertebrate surveys.
References
Aiken, K., & Kong, G. A. (2000). The marine fisheries of Jamaica. The ICLARM Quarterly, 23, 29-35. [ Links ]
Bohnsack, J. A. (1990). The potential of marine fishery reserves for reef fish management in the US Southern Atlantic. NOAA Tech. Memo NMFS-SEFC-261, Miami. [ Links ]
Bruckner, A.W. (2003). Proceedings of the Caribbean Acropora workshop: potential application of the Endangered Species Act as a conservation strategy. NOAA Tech. Memo. NMFSOPR-24. [ Links ]
Bruckner, A. (2012a). Factors contributing to the regional decline of Montastraea annularis (complex). Proc. 12th Int. Coral Reef Symp., Cairns 11B_1, Phase shifts and alternative states on coral reefs. [ Links ]
Bruckner, A. (2012b). Static measures of the resilience of Caribbean coral populations. Revista de Biologia Tropical, 60(Supl. 1), 39-57. [ Links ]
Bruckner, A. W., & Hill, R. (2009). Ten years of change to coral communities off Mona and Desecheo Islands, Puerto Rico from disease and bleaching. Diseases of Aquatic Organisms, 87, 19-31. [ Links ]
Clarke, K., & Gorley, R. N. (2006). PRIMER v6 - User Manual/Tutorial. Plymouth: Primer-e Ltd. [ Links ]
Clarke, K., & Warwick, R. (2001). Changes in marine communities: an approach to statistical analysis and interpretation. Plymouth: Primer-e Ltd. [ Links ]
Espeut, P. (2006). The Wild Frontier: Living and fishing on the Pedro Cays of Jamaica: a Socio-economic Assessment. Kingston: The Nature Conservancy. [ Links ]
Gustavson, K. (2002). Economic production from the artisanal fisheries of Jamaica. Fisheries Research, 57, 103-115. [ Links ]
Hawkins, J. P., & Roberts, C. M. (2003). Effects of fishing on sex-changing Caribbean parrotfishes. Biological Conservation, 115, 213-216. [ Links ]
Hay, D. B. (2006). Biological survey of the Pedro Cays. Kingston: The Nature Conservancy. [ Links ]
HRI. (2008). Healthy reefs for healthy people. A guide to indicators of reef health and social well-bieng in the Mesoamerican reef region. Retrieved from www.healthyreefs.org. [ Links ]
Hughes, T. P. (1994). Catastrophes, phase shifts, and large scale degradation of a Caribbean coral reef. Science, 265, 1547-1551. [ Links ]
Koslow, J. A., Hanley, F., & Wicklund, R. (1988). Effects of fishing on reef fish communities at Pedro Bank and Port Royal Cays, Jamaica. Marine Ecology Progress Series, 43, 201-212. [ Links ]
Kramer, P. R. (2006). Pedro Bank Coral Reefs: Status of coral reef and reef fishes. Report for the Nature Conservancy. Miami, FL: The Ocean Research and Education Foundation. [ Links ]
Lang, J. L. (2003). Status of coral reefs in the western Atlantic: results of initial surveys, Atlantic and Gulf Rapid Reef Assessment (AGGRA) program. Atoll Research Bulletin, 496, 1- 630. [ Links ]
Miller, J., Atkinson, A., & Witcher, B. (2009). Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs, 28, 925-937. [ Links ]
Munro, J. L. (1983). Caribbean coral reef fishery resources. A second edition of ‘The Biology, ecology, exploitation and management of Caribbean reef fishes: scientific report of the ODA/UWI Fisheries Ecology Research Project, 1969-1973’. University of the West Indies, Jamaica. ICLARM Studies Review, 7, 1-276. [ Links ]
Nicholson, W., & Hartsuijker, L. (1982). The State of the Fisheries Resources of the Pedro Bank and South Jamaica Shelf. FAO Fisheries Report, 278, 215-254. [ Links ]
Pauly, D. (1979). Theory and management of tropical multispecies stocks: a review with emphasis on the Southeast Asian demersal fisheries. ICLARM Studies Review, 1,1-35. [ Links ]
Pauly, D. (1990). On Malthusian overfishing. Naga. ICLARM Quarterly, 13(1), 3-4. [ Links ]
Szmant, A. M. (1991). Sexual reproduction by the Caribbean reef corals Montastraea annularis and M. cavernosa. Marine Ecology Progress Series, 74,13-25. [ Links ]
Szmant-Froelich, A. M. (1985). The effect of colony size on the reproductive ability of the Caribbean coral Montastraea annularis (Ellis and Solander). Proc. 5th Int. Coral Reef Symp., Tahiti, 4, 295-300. [ Links ]
Zenny, N. (2006). Technical Summary of the Jamaica Ecoregional Planning (JERP), Marine Analysis. Draft. Kingston: The Nature Conservancy Jamaica Country Programme. [ Links ]
Bohnsack, J. A. (1990). The potential of marine fishery reserves for reef fish management in the US Southern Atlantic. NOAA Tech. Memo NMFS-SEFC-261, Miami. [ Links ]
Bruckner, A.W. (2003). Proceedings of the Caribbean Acropora workshop: potential application of the Endangered Species Act as a conservation strategy. NOAA Tech. Memo. NMFSOPR-24. [ Links ]
Bruckner, A. (2012a). Factors contributing to the regional decline of Montastraea annularis (complex). Proc. 12th Int. Coral Reef Symp., Cairns 11B_1, Phase shifts and alternative states on coral reefs. [ Links ]
Bruckner, A. (2012b). Static measures of the resilience of Caribbean coral populations. Revista de Biologia Tropical, 60(Supl. 1), 39-57. [ Links ]
Bruckner, A. W., & Hill, R. (2009). Ten years of change to coral communities off Mona and Desecheo Islands, Puerto Rico from disease and bleaching. Diseases of Aquatic Organisms, 87, 19-31. [ Links ]
Clarke, K., & Gorley, R. N. (2006). PRIMER v6 - User Manual/Tutorial. Plymouth: Primer-e Ltd. [ Links ]
Clarke, K., & Warwick, R. (2001). Changes in marine communities: an approach to statistical analysis and interpretation. Plymouth: Primer-e Ltd. [ Links ]
Espeut, P. (2006). The Wild Frontier: Living and fishing on the Pedro Cays of Jamaica: a Socio-economic Assessment. Kingston: The Nature Conservancy. [ Links ]
Gustavson, K. (2002). Economic production from the artisanal fisheries of Jamaica. Fisheries Research, 57, 103-115. [ Links ]
Hawkins, J. P., & Roberts, C. M. (2003). Effects of fishing on sex-changing Caribbean parrotfishes. Biological Conservation, 115, 213-216. [ Links ]
Hay, D. B. (2006). Biological survey of the Pedro Cays. Kingston: The Nature Conservancy. [ Links ]
HRI. (2008). Healthy reefs for healthy people. A guide to indicators of reef health and social well-bieng in the Mesoamerican reef region. Retrieved from www.healthyreefs.org. [ Links ]
Hughes, T. P. (1994). Catastrophes, phase shifts, and large scale degradation of a Caribbean coral reef. Science, 265, 1547-1551. [ Links ]
Koslow, J. A., Hanley, F., & Wicklund, R. (1988). Effects of fishing on reef fish communities at Pedro Bank and Port Royal Cays, Jamaica. Marine Ecology Progress Series, 43, 201-212. [ Links ]
Kramer, P. R. (2006). Pedro Bank Coral Reefs: Status of coral reef and reef fishes. Report for the Nature Conservancy. Miami, FL: The Ocean Research and Education Foundation. [ Links ]
Lang, J. L. (2003). Status of coral reefs in the western Atlantic: results of initial surveys, Atlantic and Gulf Rapid Reef Assessment (AGGRA) program. Atoll Research Bulletin, 496, 1- 630. [ Links ]
Miller, J., Atkinson, A., & Witcher, B. (2009). Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs, 28, 925-937. [ Links ]
Munro, J. L. (1983). Caribbean coral reef fishery resources. A second edition of ‘The Biology, ecology, exploitation and management of Caribbean reef fishes: scientific report of the ODA/UWI Fisheries Ecology Research Project, 1969-1973’. University of the West Indies, Jamaica. ICLARM Studies Review, 7, 1-276. [ Links ]
Nicholson, W., & Hartsuijker, L. (1982). The State of the Fisheries Resources of the Pedro Bank and South Jamaica Shelf. FAO Fisheries Report, 278, 215-254. [ Links ]
Pauly, D. (1979). Theory and management of tropical multispecies stocks: a review with emphasis on the Southeast Asian demersal fisheries. ICLARM Studies Review, 1,1-35. [ Links ]
Pauly, D. (1990). On Malthusian overfishing. Naga. ICLARM Quarterly, 13(1), 3-4. [ Links ]
Szmant, A. M. (1991). Sexual reproduction by the Caribbean reef corals Montastraea annularis and M. cavernosa. Marine Ecology Progress Series, 74,13-25. [ Links ]
Szmant-Froelich, A. M. (1985). The effect of colony size on the reproductive ability of the Caribbean coral Montastraea annularis (Ellis and Solander). Proc. 5th Int. Coral Reef Symp., Tahiti, 4, 295-300. [ Links ]
Zenny, N. (2006). Technical Summary of the Jamaica Ecoregional Planning (JERP), Marine Analysis. Draft. Kingston: The Nature Conservancy Jamaica Country Programme. [ Links ]
1. Khaled bin Sultan Living Oceans Foundation, 8181 Professional Place, Landover, MD 20785 USA, 301-577-1288; bruckner@lof.org
Received 04-IX-2013 Corrected 02-I-2014 Accepted 24-III-2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}