










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.3 San José Sep. 2014
Sulfate reducing bacteria as secondary and necessary pathogens in black band disease of corals
Bacterias reductoras de sulfato como patógenos secundarios y necesarios en la enfermedad de la banda negra de coral
Bacterias reductoras de sulfato como patógenos secundarios y necesarios en la enfermedad de la banda negra de coral
Abstract
Black band disease (BBD) is a complex, polymicrobial disease that consists of cyanobacteria, sulfide-oxidizing and sulfate-reducing bacteria (SRB), and heterotrophic bacteria. The cyanobacterium Roseofilum reptotaenium has been implicated as the primary pathogen of BBD, but other consortium members may be secondary pathogens that are necessary to the development of the disease. It is known that populations of the sulfate-reducing bacterium Desulfovibrio are present in BBD and that these populations generate sulfide within the band as a byproduct of dissimilatory sulfate reduction. It is also known that exposure of healthy corals to sulfide leads to cell lysis and coral tissue death. Previous work showed that when freshly collected BBD, which easily infects healthy corals, is exposed to sodium molybdate, a specific inhibitor of sulfate reduction, infection does not occur. In this study we examined the effect of sodium molybdate on infection of corals by a unialgal culture of R. reptotaenium. Coral fragments of Montastraea cavernosa and Siderastrea siderea were transferred into two experimental aquaria, one a control with only artificial seawater (ASW) and the second containing ASW and 2mM sodium molybdate. Small mats of cultured R. reptotaenium were inoculated onto the surface of experimental coral fragments. Both M. cavernosa (n = 6) and S. siderea (n=4) became infected and developed BBD-like infections in the control tank, while there were temporary attachments to, but no successful infection of M. cavernosa (n=3) or S. siderea (n=2) in the experimental tank containing sodium molybdate. The results of this study reveal that a secondary pathogen is essential to the infection process and development of BBD in scleractinian corals. Specifically, SRB such as Desulfovibrio are required for the development of BBD on the coral host. This is the first step in understanding the roles of secondary pathogens in a complex, polymicrobial coral disease.
Key words: Black band disease, sulfate-reducing bacteria, coral.
Resumen
La enfermedad de la Banda Negra (BBD) es una enfermedad polimicrobiana compleja, que consiste en las cianobacterias, sulfuro oxidante y bacterias reductoras de sulfato (SRB) y bacterias heterotróficas. La cianobacteria Roseofilum reptotaenium se considera como el principal patógeno de BBD, pero otros miembros del consorcio pueden ser patógenos secundarios que son necesarios para el desarrollo de la enfermedad. Se conoce que las poblaciones de la bacteria reductora de sulfato Desulfovibrio están presentes en BBD y que estas poblaciones generan sulfuro dentro de la banda como un subproducto de la reducción del sulfato. También se conoce que la exposición de los corales sanos a sulfuro conduce a la lisis celular y la muerte del tejido de coral. Trabajos previos muestran que cuando recién colectado el tejido con BBD, que infecta fácilmente corales sanos, se expone al molibdato de sodio, un inhibidor específico de la reducción del sulfato, la infección no se produce. En este estudio analizamos el efecto del molibdato de sodio sobre la infección de los corales en un cultivo unialgal de R. reptotaenium. Fragmentos de coral de Montastraea cavernosa y Siderastrea siderea se transfirieron a dos acuarios experimentales, uno control con solo agua de mar artificial (ASW) y el segundo que contenía ASW y molibdato de sodio 2mM. Pequeñas esteras de cultivo R. reptotaenium se inocularon en la superficie de fragmentos experimentales de coral. Tanto M. cavernosa (n=6) y S. siderea (n=4) se infectaron y desarrollaron infecciones BBD- como en el tanque de control, mientras que había adjuntos temporales, sin infección exitosa en M. cavernosa (n=3) o S. siderea (n=2) en el tanque experimental con molibdato de sodio. Los resultados de este estudio revelan que un patógeno secundario es esencial para el proceso de infección y el desarrollo de BBD en corales escleractíneos. Específicamente, SRB como Desulfovibrio son necesarios para el desarrollo de BBD en el coral hospedero. Este es el primer paso en la comprensión de las funciones de los patógenos secundarios en una enfermedad polimicrobiana compleja de coral.
Palabras clave: Enfermedad de banda, sulfato reductor de bacterias, coral.
Coral diseases were first reported in the 1970s on reefs of the Caribbean (Antonius, 1976; Garret & Ducklow, 1975; Dustan, 1977; Gladfelter, Gladfelter, Monahan, Ogden & Dill, 1977). Since then, the number of documented coral diseases has increased significantly both in the Caribbean and on reefs worldwide. Coral diseases are correlated with many anthropogenic factors including elevated sea surface-temperature, sewage pollution, eutrophication, and sedimentation (Rogers, 1990; Harvell et al., 2002; Voss & Richardson, 2006). Such anthropogenic factors have also been shown to increase pathogen virulence and disease severity. For example, Voss & Richardson (2006) found that in situ nutrient dosing increased the migration rate of the band in black band disease (BBD) infected corals and accelerated coral tissue loss. Although several drivers of coral disease have been identified, the causative agents of most coral diseases are unknown. Of the 27 coral diseases documented worldwide, pathogens of only seven have been proposed, and Koch’s postulates have only been fulfilled for five –white plague type II, aspergillosis, white pox, and two cases of bacterial bleaching (Harvell et al., 2002; Weil, 2004; Harvell et al., 2007).
A wide range of diseases afflict corals on reefs in the wider Caribbean (Weil, 2004). Of these, BBD is of particular interest because it kills large, reef-building coral. BBD is comprised of by a polymicrobial consortium of cyanobacteria, sulfate-reducing and sulfide-oxidizing bacteria, and a diverse assemblage of heterotrophic bacteria that together form a dark, band-shaped microbial mat (Antonius, 1976; Garret & Ducklow, 1975; Ducklow & Mitchell, 1979; Cooney et al., 2002; Frias-Lopez, Zerkle, Bonheyo & Fouke, 2002; Frias-Lopez, Klaus, Bonheyo & Fouke, 2004; Sekar, Mills, Remily, Voss & Richardson, 2006; Sekar, Kaczmarsky & Richardson, 2008). The BBD mat migrates across the surface of infected scleractinian, as well as gorgonian, coral hosts, lysing coral tissue and leaving behind bare coral skeleton (Antonius, 1981). At rates of up to 1cm a day, BBD can denude an entire coral colony in as little as one summer season (Richardson, 1996). The pathogenicity of BBD is caused by a combination of factors: anoxia at the BBD mat-coral interface, high levels of the toxicant sulfide, the presence of the cyanotoxin microcystin, and a synergistic interaction between sulfide and microcystin (Richardson, Kuta, Schnell & Carlton, 1997; Gantar, Sekar & Richardson, 2009; Glas, Sato, Ulstrup & Bourne, 2012, Miller & Richardson, 2012).
BBD infection can be easily modeled in a laboratory setting due to the highly infectious nature of the disease. In the model system, infection can be initiated when an inoculum of naturally-occurring BBD mat from an infected coral is placed onto a healthy coral fragment (Antonius, 1985). The inoculum can be observed to attach to the coral surface, penetrate and invade tissue, and develop into a BBD lesion that migrates across the coral surface, lysing coral tissue and eventually killing the coral, leaving a bare coral skeleton. In this model the inoculum contains a fully developed BBD-consortium: cyanobacteria, sulfide-oxidizing and sulfate-reducing bacteria, as well as associated heterotrophic bacteria. While several different members of the consortium have been proposed to be the primary pathogen of BBD, Koch’s postulates have yet to be fulfilled for any member. It may be that BBD requires the entire consortium to be pathogenic –in this case it would not be possible to identify a primary pathogen and fulfill Koch’s postulates.
There has been a lot of interest in identifying a primary pathogen of BBD. The major target has been BBD cyanobacteria, since the biomass of the band is always dominated by filamentous cyanobacteria (Antonius, 1976). The other members of the consortium –sulfate reducing bacteria (SRB) and sulfide oxidizing bacteria (Ducklow & Mitchell, 1979), as well as heterotrophic bacteria (Cooney et al., 2002)– have also been implicated. However, to date investigations of potential BBD pathogens have mainly focused on BBD cyanobacteria.
A critical member of the BBD consortium is Roseofilum reptotaenium (Casamatta, Stanić, Gantar & Richardson, 2012). Roseofilum reptotaenium is a gliding, filamentous, phycoerythrin-rich cyanobacterium detected in BBD mats of tropical and sub-tropical coral reefs around the world (Miller & Richardson, 2011). This cyanobacterium forms the matrix of the mat and is adapted to the harsh BBD conditions because of its ability to conduct sulfide-insensitive oxygenic photosynthesis (Myers, Sekar & Richardson, 2007). The ability to conduct photosynthesis in the presence of sulfide is a rare trait among cyanobacteria since sulfide poisons electron flow in Photosystem II (Cohen, Jorgensen, Revsbech & Poplawski, 1986). However, this trait has been found in all BBD cyanobacterial strains tested to date (Myers & Richardson, 2009). Roseofilum reptotaenium can perform sulfide-insensitive oxygenic photosynthesis in the presence of 0.5mM sulfide (Myers & Richardson, 2009), comparable to sulfide levels measured in intact Caribbean BBD (Carlton & Richardson, 1995).
In 2009, samples of BBD were collected from BBD-infected Diploria strigosa and Siderastrea siderea on reefs in St. Croix, U.S. Virgin Islands. Two strains of R. reptotaenium were isolated from the mat, designated as strain 101-1 (isolated from D. strigosa) and 100-1 (isolated from S. siderea). Stanić (2010) found that exposure of apparently healthy D. strigosa and S. siderea coral fragments to both strains led to BBD-like lesions that ultimately killed the fragments. The inoculum of each strain attached to scleractinian coral tissue, caused an infection, and eventually developed into a BBD-like lesion.
Additional work on BBD pathogenicity has targeted SRB associated with the disease, since the population of SRB is the source of toxic sulfide present in the band (Carlton & Richardson, 1995; Glas et al., 2012). To investigate the role of SRB in BBD pathogenesis, Richardson et al. (2009) used the coral fragment model of BBD infection to assess the effect of sodium molybdate on infection. Na molybdate is a specific metabolic inhibitor of the energy yielding, anaerobic respiratory pathway of dissimilatory sulfate reduction, which produces sulfide as a byproduct of the reaction. Na molybdate effectively stops this physiological process at concentrations of 2mM. In a series of experiments in flow through aquaria containing natural, sand-filtered sea-water, Richardson et al. (2009) inoculated fragments of Montastraea annularis and S. siderea with freshly collected BBD from infected colonies on the reef. The inoculum consisted of the fully developed consortium of BBD microbes. Coral fragments that were inoculated with BBD that was exposed to 2mM of Na molybdate prior to inoculation did not develop BBD infections (Richardson et al., 2009). Conversely, coral fragments infected with inocula that were not exposed to Na molybdate all developed actively migrating BBD lesions. When these infected fragments were then exposed to 2mM Na molybdate there was no effect on disease progression. These results demonstrated that when the entire BBD community is present, SRB are required for infection but not continued disease activity. Thus, the overall implication is that SRB are directly involved in the first stages of BBD pathogenesis.
The specific role of SRB as potential primary and/or secondary pathogens of BBD infection and development remains unknown. It may be that SRB are acting together with other, as yet unidentified, BBD bacteria to initiate the disease. The goal of this study was to further assess the role of SRB in BBD infection using a laboratory culture of R. reptotaenium for the inoculum. We hypothesized that the biomass of R. reptotaenium creates an anaerobic environment on the coral surface; that this environment enriches for SRB; and that the development of an SRB population is required for BBD infection.
Materials and Methods
Fragments of two Caribbean scleractinian coral species, Montastraea cavernosa and S. siderea, were collected from the Florida Keys National Marine Sanctuary in Key West (Permit number FKMNS-2012-153). After collection, fragments were maintained at ambient (reef water) temperature during transport to Florida International University where they were placed in a 340L flow-through holding tank. After acclimation in the large tank for eight weeks, fragments approximately 4 to 7cm in diameter were then transferred to two 21L experimental aquaria for further acclimation (seven days) prior to beginning the experiment. One (control) aquarium contained only artificial seawater (ASW) at 34ppt salinity while the second (experimental) aquarium contained ASW (34ppt) as well as 2mM Na molybdate. Aquaria temperatures were kept between 27.5 and 29ºC on a 12/12 light-dark cycle using cool-white fluorescent light and metal halide bulbs. Light intensity, measured using a Biospherical Quantum Scalar Irradiance meter (model QSL100), was 1x1015quanta∙s-1∙cm-2. Experimental inoculations of the control fragments (ASW) and experimental fragments (ASW and Na molybdate) were run in parallel.
Two unialgal cultures of R. reptotaenium were used as inocula for the experiments. Strains were originally isolated using the gliding method (Castenholz, 1988) from BBD samples collected on a reef in Frederiksted, St. Croix, U.S. Virgin Islands (Stanić, 2010). The cultures were maintained in BG-11 and ASW at room temperature in filtered, natural light. Strains 101-1 and 100-1 were used to inoculate M. cavernosa and S. siderea respectively. While strain 101-1 was not isolated from BBD on M. cavernosa, preliminary results proved it was infectious for this Caribbean coral species. To inoculate the experimental coral fragments, fragments were taken out of the water and a small amount of R. reptotaenium biomass was placed on the fragment surface between polyps. A sterile glass pipette was used to hold the biomass onto the coral surface. Coral fragments were then returned to the aquaria with the pipette in place. Once the biomass attached to the surface of the coral, 24-48 hours later, the pipette was removed. Following pipette removal, if the biomass was sloughed from the surface, a reinfection was attempted in the same area. Reinfections were attempted at least twice on coral fragments with failed inoculations. When infection occurred, progression of the lesion was documented by photographing the fragments a minimum of once a week.
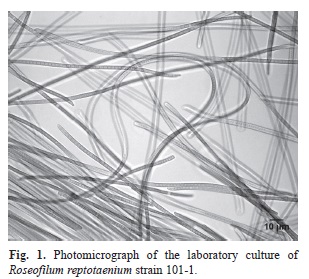
The laboratory culture of R. reptotaenium strain 101-1 was photographed using a Leica Application Suite (version 2.8.1) imaging system with a Leica/Leitz DMRB light microscope. The image was analyzed using ImageJ processing.
Results
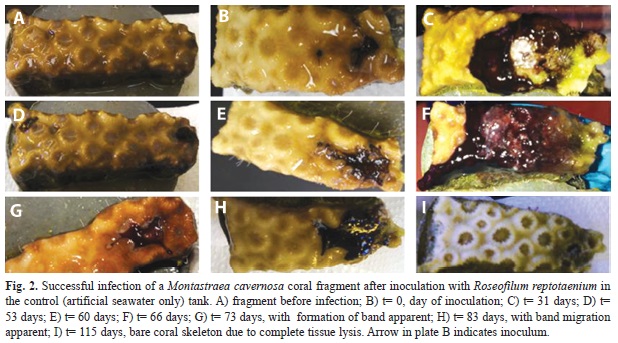
In total, 21 experiments were conducted using 18 coral fragments. Six of eight M. cavernosa fragments and four of five S. siderea fragments from the control (ASW only) aquarium were successfully infected using R. reptotaenium strain 101-1 (Fig. 1) and 100-1 respectively. In infected coral fragments, the inoculum of R. reptotaenium filaments was observed to attach to the surface of the coral tissue within seven days (Fig. 2). The small amount of inoculum developed into a BBD-like lesion as the cyanobacterial biomass increased and began to spread (Fig. 2C-F). Thin tendrils of groups of cyanobacterial filaments could be observed to migrate between and into the coral polyps ahead of the advancing microbial mat (Fig. 2C). Over a period of several weeks, the infection grew into a BBD lesion that migrated across the coral surface (Fig. 2G), lysing tissue (Fig. 2H), and leaving a bare coral skeleton (Fig. 2I). Similar to naturally occurring BBD, the mat did not tightly attach to the surface of the coral and could be easily disturbed or dislodged from the coral surface.
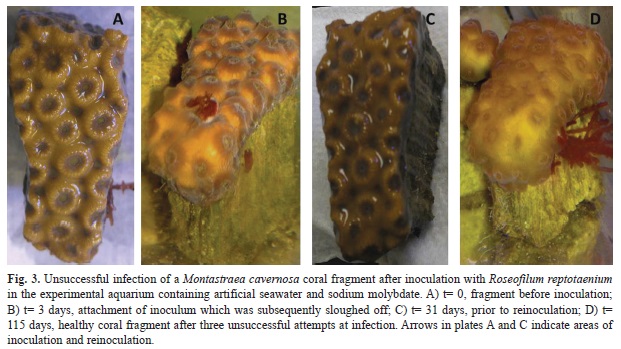
In contrast to the controls, none of the experimental (ASW plus Na molybdate) fragments –three M. cavernosa fragments and two S. siderea fragments– became infected (Fig. 3). While the R. reptotaenium inoculum was observed to attach to the surface of the coral initially (Fig. 3A), within 72 hours following attachment the filaments were sloughed off of the surface of the coral (Fig. 3B). None of the five fragments exposed to R. reptotaenium in the presence of Na molybdate exhibited permanent attachment to the coral tissue or infection of the fragments, and there was no development of BBD lesions. Reinfection of each of the five coral fragments in the experimental tank containing Na molybdate was attempted at least twice, with parallel control (ASW only) reinfections of additional fragments to serve as controls. In the Na molybdate tanks, fresh clumps of R. reptotaenium were placed in the same area on the fragments as previous attempted infections. There were no successful reinfections of the coral fragments in the experimental tank. However, reinfection of coral fragments within the control tank was successful.
Discussion
The results of this study revealed that blocking the growth of SRB using the metabolic inhibitor Na molybdate prevented the development of BBD when using unialgal cultures of the BBD cyanobacterium R. reptotaenium as the inoculum. Previous work showed that cultured R. reptotaenium can produce BBD in controlled laboratory settings and that populations of sulfur-cycle bacteria develop in the resulting BBD mat (Stanić, 2010). Additional previous work (Richardson et al., 2009) demonstrated that use of Na molybdate prevented BBD infection using freshly collected BBD from infected corals in situ. In those experiments the inoculum contained the intact polymicrobial BBD community which always includes active populations of SRB (Miller & Richardson, 2011; Glas et al., 2012).
The two strains of R. reptotaenium used in the experiments presented here are unialgal (each derived from a single cyanobacterial filament), however neither culture is axenic. In each case heterotrophic bacteria are growing in close association with the mucilaginous, polysaccharide-rich, filament sheaths. As is common with many cyanobacterial cultures, R. reptotaenium does not remain viable in an axenic culture for an extended period of time. Stanić (2010) was able to isolate strain 101-1 into axenic culture using the gliding method (Castenholz, 1988), however the culture died within six months. It is generally thought that bacteria associated with the mucilaginous sheath cyanobacteria may provide a necessary vitamin or co-factor for growth and survival. Whatever the underlying reason, the associated bacteria preclude fulfillment of Koch’s postulates, which require infection using a pure (axenic) culture.
The bacterial contaminants in the R. reptotaenium cultures were isolated, their 16S rRNA genes sequenced, and it was found that no SRB were present (Stanić, 2010). Therefore, we can conclude that SRB must be recruited from the environment into the BBD consortium. In any oxygenated environment with the presence of organic carbon, the dominant microbial metabolic mode is aerobic respiration, since oxygen is the most preferred (highest energy yielding) electron acceptor. When such environments become anaerobic, any bacterium that is capable of anaerobic energy-yielding metabolic pathways has a competitive advantage for growth. For metabolically flexible SRB, a variety of conditions present in freshwater and marine mats and sediments provide an optimal microenvironment for anaerobic respiration using sulfate as the electron acceptor, specifically anoxia and the availability of sulfate (Plugge, Zhang, Scholten & Stams, 2011). In the case of marine environments, high concentrations (28 mM) of sulfate are present in seawater, providing SRB with an immediate alternate electron acceptor (Stal, 2002). It has been shown that a transition to anaerobic conditions in the marine environment, in the presence of organic carbon released by photosynthetic cyanobacteria, always enriches for SRB (Frund & Cohen, 1992).
In our experiments, no SRB were added to the artificial sea water of the control and experimental aquaria. Two potential reservoirs for SRB, however, were the coral fragments and the live-rock used to establish the tank for two weeks prior to the experiment. We theorize that as the cyanobacterial biomass increased at the site of infection on the coral surface, it created an anaerobic environment at the mat-coral interface. The anaerobic environment then enriched for SRB from the reservoirs in the tank, allowing them to proliferate in the anaerobic environment by using sulfate as a terminal electron acceptor and releasing toxic sulfide as a byproduct. Because prevention of sulfate reduction effectively blocked BBD initiation in the aquarium with Na molybdate, the presence of SRB –thus sulfide production– appears to be crucial for the development of the band.
The presence of high levels of sulfide in BBD has proven to be critical for the pathogenicity of the disease. Using microelectrodes, sulfide levels within BBD have been measured at >800µM on the Caribbean reefs and ~5mM on the Great Barrier Reef (Carlton & Richardson, 1995; Glas et al., 2012). Sulfide is highly toxic to eukaryotes and most photosynthetic cyanobacteria at levels lower than 0.5mM (Padan, 1979), making conditions within BBD harmful to coral tissue. According to a tissue degradation rating, developed by Miller & Richardson (2012), coral fragments exposed to 0.5mM sulfide for 22 hours rated as 2.80 out of 5.0; a rating two-fold higher than the control. In the presence of sulfide the coral epidermis visibly thinned and zooxanthellae were only loosely held in the gastrodermis (Miller & Richardson, 2012), leading to disintegration of coral tissue integrity as the band travels across the surface.
In a series of experiments aimed at studying BBD pathogenesis on the Great Barrier Reef, researchers documented an increase in populations of SRB during a transition of cyanobacterial patches, shown to be precursors of BBD, to fully developed BBD infections (Sato, Willis & Bourne, 2010). In cyanobacterial patches, SRB accounted for less than 1% of the total population but that number increased beyond 7% in BBD lesions (Bourne, Muirhead & Sato, 2011). The changes in the microbial community, specifically increasing SRB, lead to enhanced production of sulfide, and sulfide concentrations within the mat increased as the cyanobacterial patch transitioned into a BBD lesion (Bourne et al., 2011). Five-fold faster migration rates were observed with increased numbers of SRB in BBD-mats as opposed to the cyanobacterial patches (Sato et al., 2010); likely due to greater sulfide production in BBD.
While different genera of SRB have been identified by sequencing the BBD microbial community, members of the genus Desulfovibrio have been most consistently documented in BBD samples worldwide (Frias-Lopez et al., 2002; Viehman, Mills, Meichel & Richardson, 2006; Sekar et al., 2006; Sekar et al., 2008; Sato et al., 2010). Viehman et al. (2006) cultured and sequenced eight strains of Desulfovibrio from BBD collected from six coral colonies of four species in the Florida Keys. Desulfovibrio was also detected by Sato et al. (2010) as the SRB in BBD lesions rather than cyanobacterial patches. The presence of Desulfovibrio and other SRB in BBD on different species of coral worldwide strengthens the proposal that SRB are required for all BBD infections.
When comparing our laboratory results using R. reptotaenium as the BBD inoculum with the results of Richardson et al. (2009) using the fully developed BBD community, marked differences in infection rate between the cyanobacterial culture and the fully developed BBD inoculum are apparent. While infection of scleractinia coral using naturally occurring BBD occurred within two days, with formation of an actively migrating band within two to three additional days (Richardson et al., 2009), infection using R. reptotaenium took place over a period of weeks. This time lag may be due to the need to recruit SRB from the aquarium environment. These results are in agreement with the results of Sato et al. (2010), who determined that the transition from a cyanobacterial patch to a BBD lesion in situ on the Great Barrier Reef (GBR) occurred over a period of 62±5 days. This time period may similarly be based on the need to recruit SRB from the environment. Alternatively, the much longer time required for BBD to fully develop from inoculation with R. reptotaenium in the aquarium experiments, or from cyanobacterial patches on the GBR, may be based on the necessity to recruit additional BBD bacteria in addition to the BBD cyanobacterium and SRB.
The work presented here, together with the body of research to date on the pathogenesis of BBD, strongly suggests that BBD is induced by a sequential community of pathogens, beginning with the cyanobacterium R. reptotaenium, followed by recruitment of SRB, and perhaps involving incorporation of additional BBD bacteria. Based on these findings we propose that SRB are secondary and necessary pathogens of black band disease. This is the first step in understanding the role of a secondary pathogen in a complex, polymicrobial coral disease.
Acknowledgments
This work was supported by the National Science Foundation (NSF grant no. 1208784) and Florida International University.
References
Antonius, A. (1976). New observations on coral destruction in reefs. Tenth Meeting of the Association of Island Marine Laboratories of the Caribbean, 10, 3. [ Links ]
Antonius, A. (1981). The ‘band’diseases in coral reefs. Proceedings from the 4th International. Coral Reef Symposium, 2, 7-14. [ Links ]
Antonius, A. (1985). Black band disease infection experiments on hexacorals and octocorals. Proceedings from the 5th International. Coral Reef Symposium, 6, 155-160. [ Links ]
Bourne, D. G., Muirhead A., & Sato, Y. (2011). Changes in sulfate-reducing bacterial populations during the onset of black band disease. The International Society of Microbial Ecology Journal, 53, 559-564. [ Links ]
Carlton, R., & Richardson L. L. (1995). Oxygen and sulfide dynamics in a horizontally migrating cyanobacterial mat- black band disease of corals. Federation of European Microbioligical Society, Microbiology. Ecology, 18, 155-162. [ Links ]
Casamatta, D., Stanić D., Gantar M., & Richardson L. L. (2012). Characterization of Roseofilum reptotaenium (Oscillatoriales, Cyanobacteria) gen. et sp. nov. isolated from Caribbean black band disease. Phycologia, 51, 489-499. [ Links ]
Castenholz, R. (1988). Culturing methods for cyanobacteria. Methods in Enzymol, 167, 68-93. [ Links ]
Cohen, Y., Jorgensen, B., Revsbech N., & Poplawski, R. (1986). Adaptation to hydrogen-sulfide of oxygenic and anoxygenic photosynthesis among cyanobacteria. Applied Environmental Microbiology, 51, 398-407. [ Links ]
Cooney, R., Pantos O., Le Tissier M., Barer M., O’Donnell, A., & Bythell, J. (2002). Characterization of the bacterial consortium associated with black band disease in coral using molecular microbiological techniques. Environmental Microbiology, 4, 401-413. [ Links ]
Ducklow, H., & Mitchell, R. (1979). Observations on naturally and artificially diseased tropical corals - scanning electron-microscope study. Microbial Ecology, 5, 215-223. [ Links ]
Dustan, P. (1977). Vitality of reef coral populations off Key Largo, Florida - recruitment and mortality. Environmental Geology, 2, 51-58. [ Links ]
Frias-Lopez, J., Zerkle, A. L., Bonheyo, G. T., & Fouke, B. W. (2002). Partitioning of bacterial communities between seawater and healthy, black band diseased, and dead coral surfaces. Applied and Environmental Microbiology, 68, 2214-2228. [ Links ]
Frias-Lopez, J., Klaus, J. S., Bonheyo, G. T., & Fouke, B. W. (2004). Bacterial community associated with black band disease in corals. Applied and Environmental Microbiology, 70, 5955-5962. [ Links ]
Frund., C., & Cohen, Y. (1992). Diurnal cycles of sulfate reduction under oxic conditions in cyanobacterial mats. Applied and Environmental Microbiology, 58, 70-77. [ Links ]
Gantar, M., Sekar, R., & Richardson, L. L. (2009). Cyanotoxins from black band disease of corals and from other coral reef environments. Microbiology Ecology, 58, 856-864. [ Links ]
Garrett, P., & Ducklow, H. (1975). Coral diseases in Bermuda. Nature, 253, 349-350. [ Links ]
Gladfelter, W., Gladfelter, E., Monahan, R., Ogden, J., & Dill, R. (1977). Environmental studies of Buck Island Reef National Monument. St. Croix: U.S.V.I. National Park Service. [ Links ]
Glas, M. S., Sato Y., Ulstrup K. E., & Bourne, D. G. (2012). Biogeochemical conditions determine virulence of black band disease in corals. The International Society of Microbial Ecology Journal, 6, 1526-1534. [ Links ]
Harvell, C., Mitchell, C., Ward, J., Altizer, S., Dobson, A., Ostfeld, R., & Samuel, M. (2002). Ecology - climate warming and disease risks for terrestrial and marine biota. Science, 296, 2158-2162. [ Links ]
Harvell, D., Jordan-Dahlgren, E., Merkel, S., Rosenberg, E., Raymundo, L., & Smith, G. (2007). Coral disease, environmental drivers, and the balance between coral and microbial associates. Oceanography, 20, 172-195. [ Links ]
Miller, A. W., & Richardson, L. L. (2011). A meta-analysis of 16S rRNA gene clone libraries from the polymicrobial black band disease of corals. Federation of European Microbioligical Society, Microbiology Ecology, 75, 231-241. [ Links ]
Miller, A. W., & Richardson, L. L. (2012). Fine structure analysis of black band disease (BBD) infected coral and coral exposed to the BBD toxins microcystin and sulfide. Journal of Invertebrate Pathology, 109, 27-33. [ Links ]
Myers, J. L., Sekar, R., & Richardson, L. L. (2007). Molecular detection and ecological significance of the cyanobacterial genera Geitlerinema and Leptolyngbya in black band disease of corals. Applied and Environmental Microbiology, 73, 5173-5182. [ Links ]
Myers, J. L. & Richardson L.L. (2009). Adaptation of cyanobacteria to the sulfide-rich microenvironment of black band disease of coral. Federation of European Microbioligical Society, Microbiology Ecology, 67, 242-251. [ Links ]
Padan, E. (1979). Facultative anoxygenic photosynthesis in cyanobacteria. Annual Review of Plant Physiology, 30, 27-40. [ Links ]
Plugge, C. M., Zhang, W., Scholten, J. C. M., & Stams, A. J. M.(2011). Metabolic flexibility of sulfate-reducing bacteria. Frontiers in Microbiology, 2, 1. [ Links ]
Richardson, L. L. (1996). Horizontal and vertical migration patterns of Phormidium corallyticum and Beggiatoa spp. associated with black-band disease of corals. Microbial Ecology, 32, 323-335. [ Links ]
Richardson, L. L, Kuta, K., Schnell, S., & Carlton, R. (1997). Ecology of the black band disease microbial consortium. Proceedings from the 8th International. Coral Reef Symposium, 1, 597-600. [ Links ]
Richardson, L. L., Miller, A. W., Broderick, E., Kaczmarsky, L., Gantar, M., Stanić, D., & Sekar, R. (2009). Sulfide, microcystin, and the etiology of black band disease. Diseases of Aquatic Organisms, 87, 79-90. [ Links ]
Rogers, C. (1990). Responses of coral reefs and reef organisms to sedimentation. Marine Ecology Progress Series, 62, 185-202. [ Links ]
Sato, Y., Willis B. L., & Bourne, D.G. (2010). Successional changes in bacterial communities during the development of black band disease on the reef coral, Montipora hispida. The International Society of Microbial Ecology Journal, 4, 203-214. [ Links ]
Sekar, R., Mills, D. K., Remily, E. R., Voss, J. D., & Richardson, L. L. (2006). Microbial communities in the surface mucopolysaccharide layer and the black band microbial mat of black band-diseased Siderastrea siderea. Applied and Environmental Microbiology, 72, 5963-5973. [ Links ]
Sekar, R., Kaczmarsky, L. T., & Richardson, L. L. (2008). Microbial community composition of black band disease on the coral host Siderastrea siderea from three regions of the wider Caribbean. Marine Ecology Progress Series, 362, 85-98. [ Links ]
Stal, L. J. (2002). Cyanobacterial mats and stromatolites, pp. 62-111. In Whitton, B. A., & Potts, M.(eds.). The Ecology of Cyanobacteria: Their Diversity in Time and Space. New York: Kluwer Academic Publishers. [ Links ]
Stanić, D. (2010). Characterization of Oscillatoria spp. and their role in black band disease of coral. M S. Thesis, Florida International University, Miami, Florida. [ Links ]
Viehman, S., Mills, D. K., Meichel, G., & Richardson, L. L. (2006). Culture and identification of Desulfovibrio spp. from corals infected by black band disease on Dominican and Florida Keys reefs. Diseases of Aquatic Organisms, 69, 119-127. [ Links ]
Voss, J. D., & Richardson, L. L. (2006). Nutrient enrichment enhances black band disease progression in corals. Coral Reefs, 25, 569-576. [ Links ]
Weil, E. (2004). Coral reef diseases in the wider Caribbean, pp. 35-68. In Loya, Y., & Rosenburg, E.(eds.). Coral Health and Disease. Berlin: Springer-Verlag. [ Links ]
Antonius, A. (1981). The ‘band’diseases in coral reefs. Proceedings from the 4th International. Coral Reef Symposium, 2, 7-14. [ Links ]
Antonius, A. (1985). Black band disease infection experiments on hexacorals and octocorals. Proceedings from the 5th International. Coral Reef Symposium, 6, 155-160. [ Links ]
Bourne, D. G., Muirhead A., & Sato, Y. (2011). Changes in sulfate-reducing bacterial populations during the onset of black band disease. The International Society of Microbial Ecology Journal, 53, 559-564. [ Links ]
Carlton, R., & Richardson L. L. (1995). Oxygen and sulfide dynamics in a horizontally migrating cyanobacterial mat- black band disease of corals. Federation of European Microbioligical Society, Microbiology. Ecology, 18, 155-162. [ Links ]
Casamatta, D., Stanić D., Gantar M., & Richardson L. L. (2012). Characterization of Roseofilum reptotaenium (Oscillatoriales, Cyanobacteria) gen. et sp. nov. isolated from Caribbean black band disease. Phycologia, 51, 489-499. [ Links ]
Castenholz, R. (1988). Culturing methods for cyanobacteria. Methods in Enzymol, 167, 68-93. [ Links ]
Cohen, Y., Jorgensen, B., Revsbech N., & Poplawski, R. (1986). Adaptation to hydrogen-sulfide of oxygenic and anoxygenic photosynthesis among cyanobacteria. Applied Environmental Microbiology, 51, 398-407. [ Links ]
Cooney, R., Pantos O., Le Tissier M., Barer M., O’Donnell, A., & Bythell, J. (2002). Characterization of the bacterial consortium associated with black band disease in coral using molecular microbiological techniques. Environmental Microbiology, 4, 401-413. [ Links ]
Ducklow, H., & Mitchell, R. (1979). Observations on naturally and artificially diseased tropical corals - scanning electron-microscope study. Microbial Ecology, 5, 215-223. [ Links ]
Dustan, P. (1977). Vitality of reef coral populations off Key Largo, Florida - recruitment and mortality. Environmental Geology, 2, 51-58. [ Links ]
Frias-Lopez, J., Zerkle, A. L., Bonheyo, G. T., & Fouke, B. W. (2002). Partitioning of bacterial communities between seawater and healthy, black band diseased, and dead coral surfaces. Applied and Environmental Microbiology, 68, 2214-2228. [ Links ]
Frias-Lopez, J., Klaus, J. S., Bonheyo, G. T., & Fouke, B. W. (2004). Bacterial community associated with black band disease in corals. Applied and Environmental Microbiology, 70, 5955-5962. [ Links ]
Frund., C., & Cohen, Y. (1992). Diurnal cycles of sulfate reduction under oxic conditions in cyanobacterial mats. Applied and Environmental Microbiology, 58, 70-77. [ Links ]
Gantar, M., Sekar, R., & Richardson, L. L. (2009). Cyanotoxins from black band disease of corals and from other coral reef environments. Microbiology Ecology, 58, 856-864. [ Links ]
Garrett, P., & Ducklow, H. (1975). Coral diseases in Bermuda. Nature, 253, 349-350. [ Links ]
Gladfelter, W., Gladfelter, E., Monahan, R., Ogden, J., & Dill, R. (1977). Environmental studies of Buck Island Reef National Monument. St. Croix: U.S.V.I. National Park Service. [ Links ]
Glas, M. S., Sato Y., Ulstrup K. E., & Bourne, D. G. (2012). Biogeochemical conditions determine virulence of black band disease in corals. The International Society of Microbial Ecology Journal, 6, 1526-1534. [ Links ]
Harvell, C., Mitchell, C., Ward, J., Altizer, S., Dobson, A., Ostfeld, R., & Samuel, M. (2002). Ecology - climate warming and disease risks for terrestrial and marine biota. Science, 296, 2158-2162. [ Links ]
Harvell, D., Jordan-Dahlgren, E., Merkel, S., Rosenberg, E., Raymundo, L., & Smith, G. (2007). Coral disease, environmental drivers, and the balance between coral and microbial associates. Oceanography, 20, 172-195. [ Links ]
Miller, A. W., & Richardson, L. L. (2011). A meta-analysis of 16S rRNA gene clone libraries from the polymicrobial black band disease of corals. Federation of European Microbioligical Society, Microbiology Ecology, 75, 231-241. [ Links ]
Miller, A. W., & Richardson, L. L. (2012). Fine structure analysis of black band disease (BBD) infected coral and coral exposed to the BBD toxins microcystin and sulfide. Journal of Invertebrate Pathology, 109, 27-33. [ Links ]
Myers, J. L., Sekar, R., & Richardson, L. L. (2007). Molecular detection and ecological significance of the cyanobacterial genera Geitlerinema and Leptolyngbya in black band disease of corals. Applied and Environmental Microbiology, 73, 5173-5182. [ Links ]
Myers, J. L. & Richardson L.L. (2009). Adaptation of cyanobacteria to the sulfide-rich microenvironment of black band disease of coral. Federation of European Microbioligical Society, Microbiology Ecology, 67, 242-251. [ Links ]
Padan, E. (1979). Facultative anoxygenic photosynthesis in cyanobacteria. Annual Review of Plant Physiology, 30, 27-40. [ Links ]
Plugge, C. M., Zhang, W., Scholten, J. C. M., & Stams, A. J. M.(2011). Metabolic flexibility of sulfate-reducing bacteria. Frontiers in Microbiology, 2, 1. [ Links ]
Richardson, L. L. (1996). Horizontal and vertical migration patterns of Phormidium corallyticum and Beggiatoa spp. associated with black-band disease of corals. Microbial Ecology, 32, 323-335. [ Links ]
Richardson, L. L, Kuta, K., Schnell, S., & Carlton, R. (1997). Ecology of the black band disease microbial consortium. Proceedings from the 8th International. Coral Reef Symposium, 1, 597-600. [ Links ]
Richardson, L. L., Miller, A. W., Broderick, E., Kaczmarsky, L., Gantar, M., Stanić, D., & Sekar, R. (2009). Sulfide, microcystin, and the etiology of black band disease. Diseases of Aquatic Organisms, 87, 79-90. [ Links ]
Rogers, C. (1990). Responses of coral reefs and reef organisms to sedimentation. Marine Ecology Progress Series, 62, 185-202. [ Links ]
Sato, Y., Willis B. L., & Bourne, D.G. (2010). Successional changes in bacterial communities during the development of black band disease on the reef coral, Montipora hispida. The International Society of Microbial Ecology Journal, 4, 203-214. [ Links ]
Sekar, R., Mills, D. K., Remily, E. R., Voss, J. D., & Richardson, L. L. (2006). Microbial communities in the surface mucopolysaccharide layer and the black band microbial mat of black band-diseased Siderastrea siderea. Applied and Environmental Microbiology, 72, 5963-5973. [ Links ]
Sekar, R., Kaczmarsky, L. T., & Richardson, L. L. (2008). Microbial community composition of black band disease on the coral host Siderastrea siderea from three regions of the wider Caribbean. Marine Ecology Progress Series, 362, 85-98. [ Links ]
Stal, L. J. (2002). Cyanobacterial mats and stromatolites, pp. 62-111. In Whitton, B. A., & Potts, M.(eds.). The Ecology of Cyanobacteria: Their Diversity in Time and Space. New York: Kluwer Academic Publishers. [ Links ]
Stanić, D. (2010). Characterization of Oscillatoria spp. and their role in black band disease of coral. M S. Thesis, Florida International University, Miami, Florida. [ Links ]
Viehman, S., Mills, D. K., Meichel, G., & Richardson, L. L. (2006). Culture and identification of Desulfovibrio spp. from corals infected by black band disease on Dominican and Florida Keys reefs. Diseases of Aquatic Organisms, 69, 119-127. [ Links ]
Voss, J. D., & Richardson, L. L. (2006). Nutrient enrichment enhances black band disease progression in corals. Coral Reefs, 25, 569-576. [ Links ]
Weil, E. (2004). Coral reef diseases in the wider Caribbean, pp. 35-68. In Loya, Y., & Rosenburg, E.(eds.). Coral Health and Disease. Berlin: Springer-Verlag. [ Links ]
1. Department of Biological Sciences, Florida International University, Miami, Florida 33199 USA; abrownel@fiu.edu
Received 02-XI-2013 Corrected 02-II-2014 Accepted 24-III-2014


{kind=link}
{kind=link}