










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.3 San José Sep. 2014
Marine Protected Area monitoring in the nearshore waters of Grenada, Eastern Caribbean: benthic cover and fish populations
Monitoreo en aguas cercanas al área marina protegida en la costa de Granada, Caribe Oriental: cobertura bentónica y poblaciones de peces
Monitoreo en aguas cercanas al área marina protegida en la costa de Granada, Caribe Oriental: cobertura bentónica y poblaciones de peces
Robert Anderson1*, Clare Morrall2*, Jonathan Jossart3*, Steve Nimrod2, Emily Bolda1, Katie Musser1, Craig Berg4* & Robert Balza1
Abstract
Grenada is highly dependent on coral reefs as a source of food and to support tourism. Local and global environmental stressors threaten these reefs. Legislation was created for this MPA in 2001, permanent mooring buoys were deployed in 2009 and enforcement of fishing restrictions began in 2010. Initiatives to address point and nonpoint source pollution from the land have recently begun, aimed at reducing stress on reef area. This study documents benthic cover and fish populations associated with reefs in a Marine Protected Area (MPA) along Grenada’s southwest coast from 2008 through 2012. Both Point Line Intercept and Photo Quadrat methods were used to assess benthic cover along permanent 30m transects (8 in and 12 outside the area) annually. Fish and Diadema antillarum urchin relative abundance were determined based on 2m wide belt surveys along the same transects. The predominant substrate cover was algae, ranging from 41% in 2009 to 74.2% in 2011. A general trend of increasing algal cover was noted. Combined annual survey results prior (2008-2010) and after controls were implemented (2011-2012) showed a significant increase in algal cover. The predominant algal form was macroalgae comprising 65.4% - 90.8% of total algae. Live hard coral percent cover ranged from 8.7% to 21.1%. Little annual variation was observed in percent live coral cover. Branching corals (34.1% - 52.3% of total living hard coral) were the most common. Of the 19 living hard coral species identified, Porites porites (21%-23%) and Porites astreoides (20%) dominated percentage composition. Madracis mirabilis contributed 21% of total live hard coral outside the MPA but only 8.7% in the MPA. Of the 63 species of fish identified in the study areas Chromis spp. (71.5% - 46%) was the dominant group. Wrasse had a significant increase from 6.9% in 2008 to 21.5% in 2010 inside the MPA with a similar increase peaking in 2011 outside the MPA. There was a noticeable (though not statistically significant) increase in piscivorous fishes in the MPA in 2012. This is a promising indication that fishing restrictions in the MPA may be having an effect. Diadema antillarum density was low, ranging from 4.58 to 0.21 urchins/100m2 outside and 0.28 to 0.10 urchins/100m2 inside despite a stocking attempt in the area in 2011.
Key words: benthic cover, reef fish, monitoring, Grenada, Eastern Caribbean, marine protected area.
Resumen
Granada es muy dependiente de los arrecifes coralinos como fuente de alimento y apoyo al turismo. Factores estresantes locales y globales amenazan con estos arrecifes. La legislación fue creada para esta área protegida en el 2001, boyas de amarre fueron colocadas en 2009 y las restricciones a la pesca comenzaron a implementarse a partir del 2010. Iniciativas para reducir el estrés por contaminación terrestre, de fuentes puntuales y no-puntuales, en la zona comenzaron recientemente. Este estudio documenta la cobertura bentónica y poblaciones de peces asociados a los arrecifes del área marina Protegida (MPA) a lo larfo de la costa suroeste de Granada del 2008 al 2012. Se utilizaron los métodos de Punto Línea Intercepción (PLI) y Cuadrantes Fotográficos (PQ) para evaluar anualmente la cobertura bentónica a lo largo de transectos permanentes de 30m (ocho dentro y 12 fuera del MPA). La abundancia relativa de peces y del erizo negro Diadema antillarum a través de censos visuales de 2m de ancho a lo largo de los mismos transectos lineales. La cobertura de sustrato estuvo dominada por algas con 41% en 2009 y 74.2% en 2011. Se notó una tendencia general de aumento en la cobertura algal. La combinación de los resultados antes de las encuestas (2008-2010) y después de la implementación de los controles del MPA (2011-2012) mostró un aumento significativo en la cobertura algal. La forma algal predominante fue macroalgas que consta el 65.4% y el 90.8% del total de algas. El porcentaje de cobertura de coral durovarió entre un 8.7% y un 21.1%. Se observó una pequeña variación anual en el porcentaje de cobertura de coral vivo. Los corales ramificados (34.1%-52.3% del total de corales duros) fueron la forma de coral más común encontrada durante el estudio. De las 19 especies de corales duros identificados, Porites porites (21% - 23%) y Porites astreoides (20%) dominaron el porcentaje de composición. Madracis mirabilis contribuyó en un 21% del total de corales duros fuera del MPA pero solo un 8.7% en el MPA. De las 63 especies de peces identificadas en el área de estudio Chromis spp. (71.5% - 46%) fue el grupo dominante. Los lábridos (Labridae) mostraron un aumento significativo de su abundancia de un 6.9% en 2008 a un 21.5% en 2010 dentro del MPA con un pico de incremento similar en el 2011 fuera de la MPA. Hubo un aumento notable (aunque no significativo estadísticamente) en los peces piscívoros dentro del MPA en 2012. Esto es una indicación prometedora de que las restricciones de pesca en la MPA pueden estar dando efecto. La densidad de Diadema antillarum fue baja, osciló entre 4.58 y 0.21 erizos/100m2 fuera del MPA y entre 0.28 y 0.10 erizos/100m2 dentro del MPA a pesar de la existencia de un programa de repoblación de la especie llevado a cabo en el 2011.
Palabras clave: cubierta bentónica, peces de arrecife, monitoreo, Granada, área protegida del Caribe, Marina oriental.
The Reefs at Risk Revisited report (Burke, Reytar, Spalding & Perry, 2011) documents Grenada as a country with high exposure to reef threats and high reef dependence. Many countries have established Marine Protected Areas (MPAs) to conserve coral reef systems thereby addressing problems associated with coral community decline (Kelleher, 1999; Guarderas, Hacker & Lubchenco, 2008; Graham, Barrett & Stuart-Smith, 2009; Selig & Bruno, 2010; Alvarado, Cortés, Esquivel & Salas, 2012; Crabbe, 2013). It is hoped that by protecting these areas they will recover and serve as a source of biodiversity for adjacent communities (Kelleher, 1999). In addition, MPAs help raise community awareness of the economic value of coral reefs and provide opportunities to educate the public about the threats to coral communities (Angulo-Valdes & Hatcher, 2010; Sala et al., 2013).
Since tourism is the primary industry of Grenada and the nearshore reefs are important tourist attractions, the Grenadian government established legislation for the Moliniere-Beausejour MPA on the southwest coast of the island in 2001 (Byrne, 2007; Turner, 2009). Permanent mooring buoys were established in 2009 and in 2010 warden patrols began enforcing newly established fishing and anchoring restrictions. Annual monitoring of fish and coral communities within Grenada’s MPA and at similar coral communities outside the MPA began in 2008 (Anderson et al., 2012). Monitoring is an essential component of the success of this MPA (Convention on Biological Diversity, 2012). The biological data collected over the past five years can be used to inform adaptive management of the Moliniere-Beausejour MPA. The current study provides a detailed analysis of substrate cover including relative abundance of coral species as well as two years of monitoring data since implementation of MPA restrictions.
Methods and Materials
Study Area: The Moliniere-Beausejour MPA is located along Grenada’s southwest coast. Two study sites are in the MPA (Dragon Bay, 12°5’6.00”N, 61°45’45.36”W and Flamingo Bay, 12°5’30.36”N, 61°45’30.60”W) and three sites on nearby reef areas outside the MPA (Northern Exposure Shallow, 12°1’57.30”N, 61°46’14.28”W; Northern Exposure Deep, 12°2’22.14”N, 61°46’4.74”W and Quarter Wreck, 12°1’40.98”N, 61°47’0.84”W). Water depth at the study sites ranged from 5.2m-12.2m. All sites were established in 2008; in water surveys were conducted during May or June each year.
Methods: Both Point Line Intercept (PLI) and Photo Quadrat (PQ) methods were used to assess substrate type inside and outside the Moliniere-Beausejour MPA. Relative abundance of major substrate types along with fish species and Diadema antillarum were estimated based on a revision to the Crosby and Reese (1996) PLI method developed by Crosby and Bruckner in 2002. Three algal forms were identified in the sampling protocol: macro algae, turf algae and coralline algae. In addition live hard coral was grouped into branching, massive, plate and encrusting forms. Four 30m parallel permanent transects were set up at each of the five sampling sites. Substrate type was recorded at points at 50cm intervals along the 30m transects. Fish species and D. antillarum observed within a two meter wide belt along the transect tape and throughout the water column during a 10 minute scan were recorded. In order to enhance the reliability of these observations digital photographs were taken with a Canon EOS Digital Rebel XTI, with EF-S 60mm f/2.8 Macro USM lens and dual Ikelite DS160 strobe lights each 50cm along the transect tape immediately after the PLI transects were completed. An “L” shaped PVC pipe attached to the camera housing was used to maintain a 60cm distance from the substrate and provide a standard scale for size calibration. CPCe v.3.6 and v.4.0 (Kohler & Gill, 2006) was used to estimate substrate cover types in a 20cm by 20cm square within each photo. The substrate under eight randomly generated points was identified providing a total of 480 points per transect. An approximately six minute video scan of each transect at each study site was also made with a Sony HDR-SR8 in an Amphibico housing to provide a general perspective of the coral community.
Coral species encountered along transects were identified and relative abundance determined based on occurrence within 50cm by 30cm rectangles created on 2011 transect photographs using CPCe v.3.6. Scleractinian corals were identified to species level and octocorals were identified to genus unless picture quality would not allow sufficient detail for identification. Identifications were based on Humann (1993) and Sprung (1999).
Statistical Analysis: For the photo quadrat, point line intercept, and fish data a Repeated Measure Analysis of Variance (ANOVAR) with two factors, time and protection, was used to determine if the category varied by year or location (inside and outside of the MPA). All data was tested for normality, equal variance, and sphericity, and an ArcSine Root or Log transformation was used to satisfy the assumptions of normality and equal variance. A Greenhouse-Geisser correction was used for data that did not meet the assumption of sphericity. When significant differences were detected a follow up one-way ANOVA for time and a T-Test for protection were used to determine when the differences occurred. If the data did not meet the assumptions of equal variance or normality, despite transformations, the ranked values were analyzed instead. Non-normally distributed data, if deviation from normality was not severe, was still analyzed as ANOVA tests are robust to deviations from normality (Sokal & Rohlf, 1995). Additionally, before (2008-2010) and after (2011-2012) analysis using a T-Test or a non-parametric Wilcoxon test was done on the various categories to determine if enforcement measures may have had an effect. For the 2011 coral species abundance data a non-parametric Wilcoxon test was also used to determine significant differences between a particular coral species inside and outside the Marine Protected Area.
Results
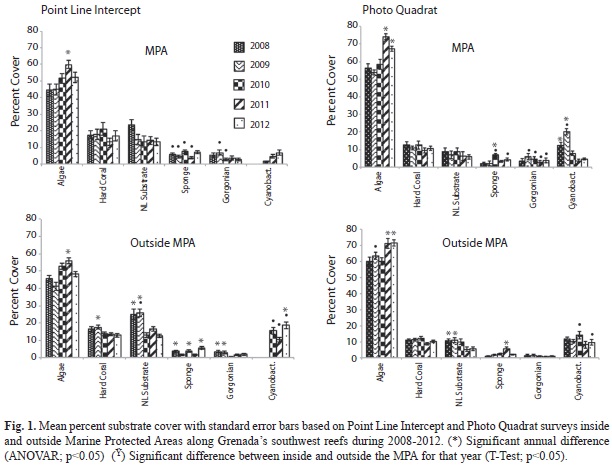
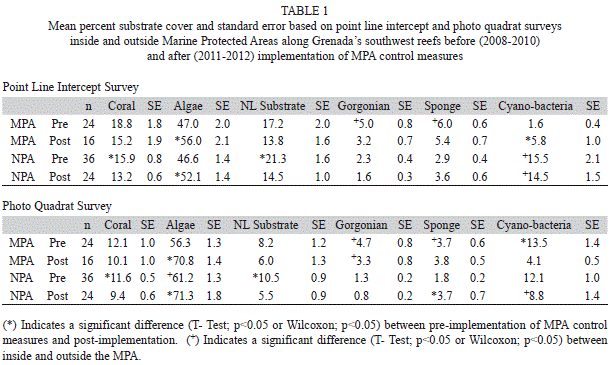
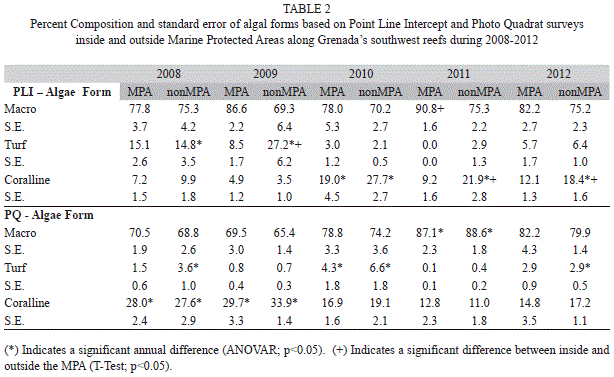
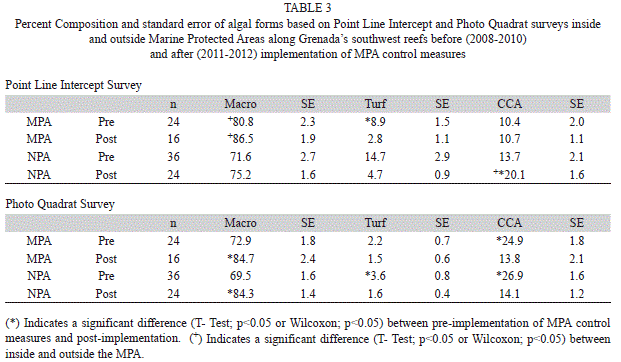
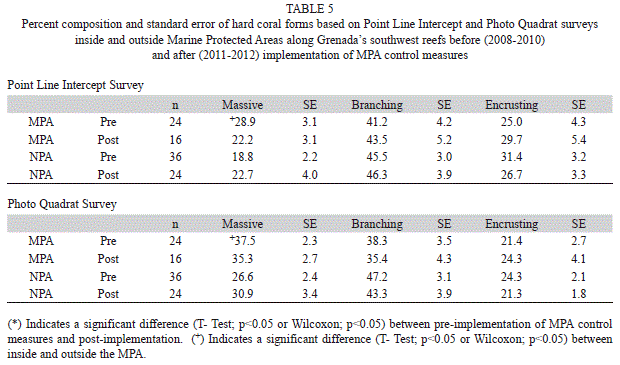
Substrate: Algae was the dominant substrate cover at survey sites inside and outside the MPA. Both the Point Line Intercept (PLI) and Photo Quadrat (PQ) survey methods showed a trend of increasing algal cover reaching a peak in 2011 (Fig. 1). Combining survey results prior to implementation of MPA controls (2008-2010) and comparing this to combined data after MPA controls were implemented (2011-2012) showed a significant increase in algal cover for both PLI and PQ after controls were implemented both inside and outside the MPA (Table 1) (T-Test, p<0.05). Algal cover ranged from 41.1% (SE=2.3, n=60) outside the MPA in 2009 to 74.2% (1.6, 40) inside the MPA in 2011 and was not significantly different (T-Test, p>0.05) inside and outside the MPA except in 2009 PQ surveys (T-Test, p<0.05). Three algal forms were identified in the sampling protocol: macro algae, turf algae and coralline algae. Macro algae dominated ranging from 65.4% (3.1, 60) to 90.8% (1.6, 40) of the total algae found (Table 2). Both PLI and PQ surveys indicated an increasing trend for macro algae across the years although these differences were not statistically significant (ANOVAR, p>0.05). A significant difference between percent composition of macro algae inside and outside the MPA was seen only in 2011 (T-Test, p<0.05) when it reached its highest level of 90.8% in the MPA while outside the MPA it was 75.3% (2.2, 60). Percent composition of macro algae inside and outside the MPA was not significantly different (T-Test, p>0.05) after implementation of MPA controls relative to prior to implementation however it was significantly higher in the MPA in both time frames (Table 3) (T-Test, p<0.05). Turf algae had significant annual variation (ANOVAR, p<0.05) with the highest percent composition of 27.2% (6.2, 60) occurring during 2009 outside the MPA. That same year turf algae was significantly less (T-Test, p<0.05) inside the MPA only reaching 8.5% (1.7, 40) of total algae found. The contribution of turf algae to total algal composition since 2009 has been less than 7% at all sites (Table 2). The combined years comparison revealed that the proportion of turf algae decreased significantly after implementation of controls in the MPA (T-Test, p<0.05). Outside the MPA turf algae also decreased however this was not significant (Wilcoxon, p=0.089), although it was very close to being significant. Photo Quadrat results did show significantly less turf algae outside the MPA after fishing controls were implemented in the MPA (Table 3) (T-Test, p<0.05). Percent cover of coralline algae was significantly higher in 2008 and 2009 than in 2010 through 2012 (ANOVAR, p<0.05) in the PQ surveys. Percent contribution of coralline algae in the PLI surveys for 2008 and 2009 were much lower than the PQ results for the same transects possibly due to under estimates by the divers. This shows the benefit of photo quadrat sampling on the same transects as the PLI surveys. The coralline algae contribution inside and outside the MPA was significantly higher before MPA controls were implemented based on PQ data (T-Test, p<0.05) however PLI results showed that coralline algae was significantly higher after MPA controls outside the MPA (Table 3) (Wilcoxon, p<0.05).
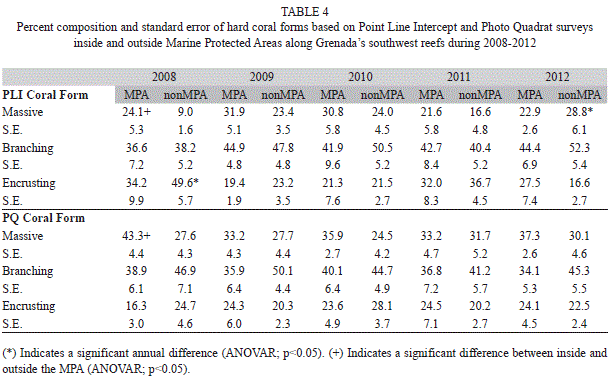
Live hard coral percent cover (Fig. 1) ranged from 8.7% (0.8, 60) outside the MPA in 2011 to 21.1% (3.9, 40) inside the MPA in 2010 with little annual variation inside or outside the MPA (ANOVAR p>0.05). Percent cover for live hard coral was somewhat greater inside than outside the MPA but this difference was not significant (T-Test, p>0.05). Combined year comparisons show in both PLI and PQ results that percent live hard coral cover did not change significantly in the MPA since implementation of control measures (T-Test, p>0.05). In addition there was no significant difference between live hard coral cover inside and outside the MPA (T-Test, p>0.05). Interestingly, live hard coral cover decreased significantly after MPA controls outside the MPA (Table 1) (T-Test, p<0.05). Branching coral was the predominant form of coral found on the transects ranging from 34.1% (5.3, 40) to 52.3% (5.4, 60) of all coral. The percent composition of this coral form varied little from year to year and comprised a slightly greater portion of the hard coral outside the MPA compared to within the MPA (Table 4). Combined annual results before and after implementation of MPA controls (Table 5) showed that the percent composition of branching coral did not change significantly (T-Test, p>0.05). Massive coral percent composition on the other hand was significantly greater inside than outside the MPA (T-Test, p<0.05) before but not after controls were implemented based on both the PLI and PQ surveys (Table 5). Massive and encrusting coral were similar in percent composition of the total hard coral community with no significant annual variation (ANOVAR, p>0.05) except for a significantly higher percent composition (T-Test, p<0.05) seen outside the MPA in 2012 compared to 2008 (Table 4). Percent composition of massive coral was significantly higher in the MPA in 2008 (T-Test, p<0.05) than outside the MPA but the difference declined somewhat through the years to the point that massive coral percent composition was higher outside the MPA in the 2012 PLI survey (Table 4).
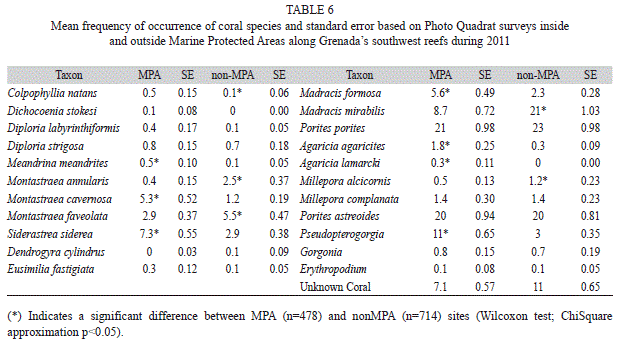
A total of 22 coral taxa were identified (19 hard coral species and three octocoral genera) in the nearshore waters of Grenada. In the MPA Porites porites and Porites astreoides dominated the surveys (Table 6) making up 21% (1.0, 478) and 20% (0.9, 478) of the live coral cover respectively and no significant difference in percent composition outside the MPA (Wilcoxon, p>0.05) was found. Madracis mirabilis was also a major coral species outside the MPA making up 21% (1.0, 714) of the live coral cover however it was significantly lower (Wilcoxon, p<0.05) at 8.7% (0.7, 478) in the MPA. Massive corals Montastraea cavernosa and Siderastrea siderea occurred more frequently in the MPA (Wilcoxon, p<0.05) while Montastraea annularis and Montastraea faveolata occurred more often outside the MPA (Wilcoxon, p<0.05). Soft coral, Pseudopterogorgia spp., was more prevalent in the MPA where it made up 11% (0.7, 478) of the total substrate cover compared to the 3% (0.4, 714) outside the MPA (Wilcoxon, p<0.05). The species Dichocoenia stokesi and Agaricia lamarcki were only found in the MPA.
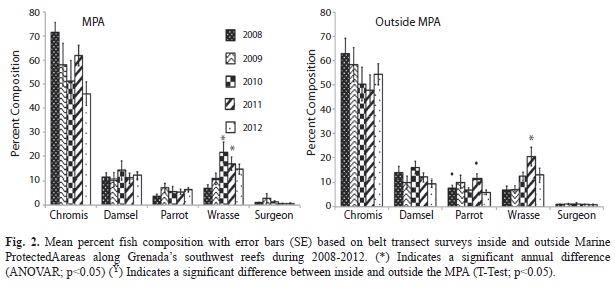
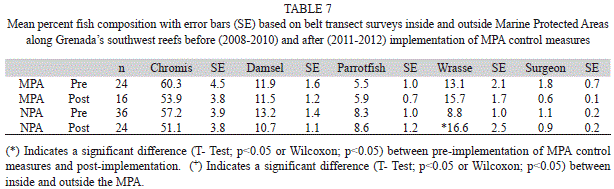
Fish: Of the 63 species of fish observed along transects inside and outside the MPA (Table 8) Chromis spp. was the dominant group (Fig. 2). Through the five years of the study the relative composition of Chromis spp. declined from 71.5% (4.2, 40) of the total fish recorded to 46.0% (4.9, 40) in the MPA. Outside the MPA Chromis spp. also declined from 2008 through 2011 but increased to nearly the 2009 level in 2012. None of these annual variations were statistically significant (ANOVAR, p>0.05). The proportion of Wrasses increased significantly in the MPA from 6.9% (1.5, 40) in 2008 to 21.5% (4.4, 40) in 2010 (ANOVA p<0.05) but declined in 2011 and 2012. Outside the MPA the proportion of wrasses increased significantly from 6.9% (1.6, 60) in 2008 to 20.4% (3.9, 60) in 2011 (ANOVA, p<0.05) but declined to 12.8% (2.7, 60) in 2012. Percent composition of territorial Damselfishes has remained fairly constant across years in the 10% range (ANOVAR p>0.05). Parrotfishes and Surgeonfishes also remained steady from year to year around the 5% (ANOVAR, p>0.05) and 1% (ANOVAR, p>0.05) range respectively in the MPA. A similar annual pattern for Parrotfishes and Surgeonfishes was seen outside the MPA; however in 2008 (T-Test, p<0.05) and 2011 (T-Test, p<0.05) percent composition of Parrotfishes was significantly higher outside compared to inside the MPA (Fig. 2). Analysis of combined annual results prior to implementation of fishing controls and after implementation showed no significant change for the major groups of fishes observed along transects inside and outside the MPA (T-Test, p>0.05). The only exception to this was a significant increase in wrasse outside the MPA after implementation of fishing controls (T-Test, p<0.05) (Table 7).
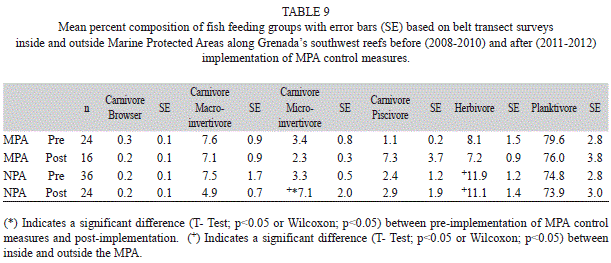
The fish observed along transects were grouped based on their feeding habits following Sandin, Sampayo and Vermeij (2008). Planktivores, comprised mainly of Blue and Brown Chromis as well as Bicolor Damselfish, dominated the feeding groups’ percent composition (Fig. 3). Percent composition of planktivores was significantly greater inside than outside the MPA in 2011 and significantly higher in 2008 and 2011(T-Test, p<0.05). Herbivores, made up of Parrotfishes, territorial Damselfishes, and Surgeonfishes, ranked second among the percent composition of feeding groups and were significantly greater outside the MPA in 2008 (T-Test, p<0.05) and 2011 (T-Test, p<0.05). There was a large increase in the piscivore feeding group in the MPA during 2012; however this increase was not significant due to the high variation in the data (ANOVAR, p>0.05). Combining percent composition of fish feeding groups results revealed that carnivorous microinvertivore feeders were significantly more abundant outside the MPA than inside after implementation of controls in the MPA (T-Test, p<0.05). Herbivorous fishes were significantly more abundant outside the MPA than inside before and after implementation of MPA controls (Table 9) (T-Test, p<0.05).
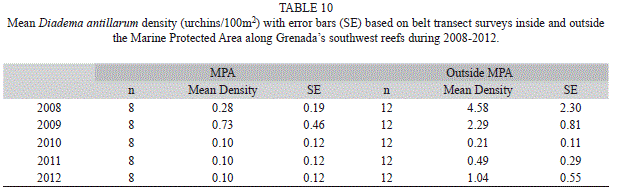
Diadema antillarum: Density of Diadema antillarum has been consistently greater outside than inside the MPA during 2008 through 2012. Density ranged from 4.58 urchins/100m2 (2.3; 24) outside the MPA in 2008 to 0.1 urchins/100m2 (0.1; 16) in the MPA during the 2010 - 2012 surveys. There is a general decline in density across the years (Table 10).
Discussion
Nearshore coral reefs are suffering from local as well as global environmental impacts. Local impacts such as overfishing, nutrient and soil runoff from farms, municipal pollution and damage from tourist activities combine with the impacts of global climate change to cause degradation of coral communities (Gardner, Côte´, Gill, Grant & Watkinson, 2003; Burke & Maidens, 2004; Knowlton & Jackson, 2008; Burke et al., 2011; Jackson et al., 2012). Changes in global climate have resulted in ocean temperatures high enough to cause bleaching and stress to corals (Baker, Glynn & Riegl, 2008; Bruckner & Hill, 2009; Cantin et al., 2010; Eakin et al., 2010; Buddemeier, lane & Martinich, 2011; Riegl, Berumen & Bruckner, 2013). Local impacts such as increased nutrient levels have now been shown to increase sensitivity of corals to bleaching (Wiedemann et al., 2013). Increased CO2 levels in the atmosphere drive higher CO2 levels in ocean water reducing pH that in turn hampers formation of the calcium carbonate skeletons of some coral species (Ries, Stanley & Hardie, 2006, Fine & Tchernov, 2007; Riegl, Bruckner, Coles, Renaud & Dodge, 2009). Marine Protected Areas have the potential to offer some relief from the local stressors thereby increasing potential resilience of the coral community to impacts associated with climate change (Riegl et al., 2009; Selig & Bruno, 2010; Riegl et al., 2013). The purpose of this study is to monitor the coral reef community in Grenada’s MPA as various management practices are implemented.
The five years of data compiled in this study to date confirm concerns that Grenada’s nearshore reefs are at risk as indicated in Burke et al. (2011). The general increase in macro algal cover and low percent live coral cover are clear indicators of continued local as well as global stresses. High relative percent of macro algae compared to turf and coralline algae both inside and outside the MPA are indicators of high nutrient levels and overfishing (Littler & Littler, 2007; Sandin et al., 2008). The greater proportion of branching corals compared to massive corals and the high relative percent of non-framework building corals (Gardner et al., 2003) such as Porites porites, Madracis mirabilis, and Porites astreoides (Table 4) are not indicative of a resilient coral reef system. The predominance of planktivorous fishes and low percent composition of piscivores are likely a result of selective overfishing and nutrient loading (Knowlton & Jackson, 2008; Sandin, 2008; Jackson et al., 2012).
Grenada’s Moliniere-Beausejour MPA encompasses an important portion of the reefs along Grenada’s southwest coast and improvement in the reef community in the MPA has the potential for improving all of the coral reef communities along the southwest shore (Angulo-Valdes & Hatcher, 2009; Crabbe, 2013; Sala et al., 2013). Fishing restrictions and required usage of permanent mooring structures have been implemented in the MPA. The MPA is patrolled and information campaigns highlighting the role and importance of the MPA have targeted fishermen as well as the general public on the island. An attempt to reduce macro algae in the MPA by introducing D. antillarum in 2011 (Nimrod, 2012) did not result in a detectible increase in urchin density during surveys in 2011 or 2012 (Table 10) and in fact D. antillarum density in the MPA was actually lower in 2010 through 2012 than in the previous two years. Comparison of combined annual survey data before MPA controls were implemented (2008-2010) to surveys after implementation (2011-2012 show that algal cover on the reef has increased significantly both inside and outside the MPA. This algal increase along with some reduction in live hard coral cover suggests that protection measures have not yet impacted the benthic reef community. To date no significant changes in the benthic or fish community have been recorded in the MPA. It is interesting to note however that there was an increase in piscivores and a slight decline in planktivores in the MPA in 2012 (Fig. 3, Table 9). These results may be an indication that additional measures need to be taken to enhance the MPA. Measures implemented thus far target fishing and physical damage to the reef. Since excess nutrient runoff from shore is a potential driver of algal growth this may be an important issue to be addressed by all concerned with marine resources.
Studies of local nutrient runoff from point and nonpoint sources on local farmland and municipal areas are under development. In addition the government of Grenada Fisheries Division will soon be implementing a modified version of the Great Barrier Reef Marine Park Authority ‘Reef Guardian’ educational program for groups including farmers (pers. comm. C. Andrew 2013). It is hoped that follow through on these initiatives may help to reduce the stresses posed to this coral community.
This ongoing nearshore monitoring study is focused on those reefs used most extensively by Grenada’s dive and snorkel industry. It is hoped that the data collected will assist in raising public awareness of conditions on the reefs and also assist MPA managers in applying effective management practices. Enhanced reefs in the nearshore waters of Grenada’s southwest shore could result in a boost to the island’s economy through increased interest in dive tourism. With appropriate regulation of tourist and local user activities this area will be a sustainable resource for Grenada.
Acknowledgments
Funding for this project was provided by the Fischer Family Foundation and Gary Stimac and is greatly appreciated. Thanks are also offered to Jacob Krause, Jillian Groeschel, Kyle Foster, Svetlana Bornschlegl, Victoria Krueger, Thomas Dietrich, Ben Hermanson, Laurelyn Dexter, Angela Majeskie, Allison Page, and Angela Blasezyk, Andrew Dunlop, Nate Sorum, Joe DuFore, Courtney Moll, Arielle Schildback, Heather McLain and Henry Tews for data collection. Billie Harrison of the Milwaukee County Zoo is thanked for expert field work.
References
Alvarado, J. J., Cortés J., Esquivel M. F., & Salas, E. (2012). Costa Rica’s Marine Protected Areas: status and perspectives. Revista de Biologia Tropical, 60, 129-142. [ Links ]
Anderson, R., Morrall C., Nimrod S., Balza R., Berg C., & Jossart, J. (2012). Benthic and fish population monitoring in the nearshore waters of Grenada, Eastern Caribbean. Revista de Biologia Tropical, 60, 71-87. [ Links ]
Angulo-Valdes, J. A., & Hatcher, B. C. (2010). A new typology of benefit derived from marine protected areas. Marine Policy, 34, 635-644. [ Links ]
Baker, A. C., Glynn P. W., & Riegl, B. (2008). Climate change and coral reef bleaching: an ecological assessment of long-term impacts, recovery trends and future outlook. Estuarine, Coastal and Shelf Science, 80, 435-471. [ Links ]
Bruckner, A.W., & Hill, R. (2009). Ten years of change to coral communities off Mona and Desecheo Islands, Puerto Rico from disease and bleaching. Diseases of Aquatic Organisms, 87, 19-31. [ Links ]
Buddemeier, R.W., Lane, D.R., & Martinich, J. A. (2011). Modeling regional coral reef responses to global warming and changes in ocean chemistry: Caribbean case study. Climatic Change, 109, 375-397. [ Links ]
Burke, L., & Maidens J. (2004). Reefs at risk in the Caribbean. Washington, D.C., USA: World Resources Institute. [ Links ]
Burke, L., Reytar, K., Spalding M., & Perry, A. (2011). Reefs at Risk Revisited. Washington, D.C., USA: World Resources Institute. [ Links ]
Byrne, J. (2007). Grenada Gap Analysis. Christiansted, St. Croix, USVI. 25: The Nature Conservancy. [ Links ]
Cantin, N. E., Cohen, A. L., Karnauskas, K. B., Tarrant, A. M., &. McCorkle, D. C. (2010). Ocean warming slows coral growth in the central Red Sea. Science, 329, 322-325. [ Links ]
Convention on Biological Diversity. (2012). Action Plan for Implementing the Convention on Biological Diversity’s Programme of Work on Protected Areas (Grenada). Retrieved from http://www.cbd.int/protected/implementation/actionplans/country/?country=gd. [ Links ]
Crabbe, M. (2013). Coral Reef Populations in the Caribbean: Is There a Case for Better Protection against Climate Change? American Journal of Climate Change, 2(2), 97-105. [ Links ]
Crosby, M. P., & Reese E. S. (1996). A manual for monitoring coral reefs with indicator species: butterfly fishes as indicators of change on the Indo-Pacific reefs. NOAA, Silver Spring, Maryland, USA: Office of Ocean and Coastal Resource Management, [ Links ]
Eakin, C. M., Morgan J. A., Heron S. F., Smith T. B, Liu G., Alvarez-Filip, L., Baca, B., Bartels, E. Bastidas, C., …, & Yusuf, Y. (2010). Caribbean corals in crisis: record thermal stress, bleaching, and mortality in 2005. PLoS ONE, 5, e13969. [ Links ]
Fine, M. & Tchernov, D. (2007). Scleractinian coral species survive and recover from decalcification. Science, 315, 1811. [ Links ]
Gardner, T. A., Côte´, I. M., Gill, J. A., Grant, A., & Watkinson A. R. (2003). Long term region-wide declines in Caribbean corals. Science, 301, 958-960. [ Links ]
Graham, J. E., Barrett, N. S., & Stuart-Smith R. D. (2009). Exploited reefs protected from fishing transform over decades into conservation features otherwise absent from seascapes. Ecological Application, 19, 1967-1974. [ Links ]
Guarderas, A. P., Hacker, S. D., & Lubchenco, J. (2008). Current Status of Marine Protected Areas in Latin America and the Caribbean. Conservation Biology, 22, 1630-1640. [ Links ]
Humann, P. (1993). Reef Coral Identification (Florida, Caribbean, Bahamas). Jacksonville: New World Publications. [ Links ]
Jackson, J., Cramer, K. Donovan, M., Friedlander A., Hooten, A., & Lam, V. (2012). Tropical Americas Coral Reef Resilience Workshop - 2012. GCRMN Technical Report. Panama City, Panama: Smithsonian Tropical Research Institute. [ Links ]
Kelleher, G. (1999). Guidelines for Marine Protected Areas. IUCN - The World Conservation Union. Series 3. [ Links ]
Knowlton, N., & Jackson J. (2008). Shifting Baselines, Local Impacts, and Global Change on Coral Reefs. PLoS Biology, 6(2), e54. [ Links ]
Kohler, K. E., & Gill, S.M. (2006). Coral Point Count with excel extensions (CPCe): A visual basic program for the determination of coral and substrate coverage using random point count methodology. Computers & Geosciences, 32, 1259-1269. [ Links ]
Littler, M. M., & Littler, D. S. (2007). Assessment of coral reefs using herbivory, nutrient assays and indicator groups of benthic primary producers: a critical synthesis, proposed protocols, and critique of management strategies. Aquatic Conservation: Marine and Freshwater Ecosystems, 17, 195-215. [ Links ]
Nimrod, S. (2012). The effectiveness of Diadema in triggering a phase shift reversal. Mini-Symposia presentation 12th ICRS, Cairns, Australia. [ Links ]
Riegl, B., Bruckner, A., Coles, S., Renaud, P., & Dodge, R. E. (2009). Coral Reefs - Threats and conservation in an era of global change. Ann. NY Academic of Science, 1162, 136-186. [ Links ]
Riegl, B., Berumen, M., & Bruckner, A. (2013). Coral population trajectories, increased disturbance and management intervention: a sensitivity analysis. Ecology & Evolution, 3(4), 1050-1064. [ Links ]
Ries, J. B., Stanley S. M., & Hardie, L. A. (2006). Scleractinian corals produce calcite, and grow more slowly, in artificial Cretaceous seawater. Geology, 34, 525-528. [ Links ]
Sala, E., Costello, C., Dougherty, D., Heal, G., Kelleher, K., Murray, J., Rosenberg, A., & Sumaila, R. (2013). A General Business Model for Marine Reserves. PLoS ONE, 8(4), e58799. [ Links ]
Sandin, S. A., Sampayo E. M., & Vermeij, M. J. (2008). Coral reef fish and benthic community structure of Bonaire and Curaçao, Netherlands Antilles. Caribbean Journal of Science, 44, 137-144. [ Links ]
Selig, E. R., & Bruno, J. F. (2010). A Global Analysis of the Effectiveness of Marine Protected Areas in Preventing Coral Loss. PLoS ONE, 5(2), e9278. [ Links ]
Sokal, R. R., & Rohlf, F. J. (1995). Biometry: The principles and practice of statistics in biological research. 3rd edition. New York: W.H. Freeman. [ Links ]
Sprung, J. (1999). Oceanographic Series Corals: A Quick Reference Guide. Miami: Ricordia Publishing. [ Links ]
Turner, M. (2009). Draft Grenada Protected Area System Plan Part 1 - Identification and Designation of Protected Areas. Prepared for the Environment and Sustainable Development Unit (ESDU) of the Organization of Eastern Caribbean States (OECS) Protected Areas and Associated Livelihoods (OPAAL) Project. [ Links ]
Wiedenmann, J., D’Angelo, C., Smith, E. G., Hunt, A. N., Legiret, F. E., Postle, A. D., & Achterberg, E. P. (2013). Nutrient enrichment can increase the susceptibility of reef corals to bleaching. Nature and Climate Change, 3, 160-164. [ Links ]
Anderson, R., Morrall C., Nimrod S., Balza R., Berg C., & Jossart, J. (2012). Benthic and fish population monitoring in the nearshore waters of Grenada, Eastern Caribbean. Revista de Biologia Tropical, 60, 71-87. [ Links ]
Angulo-Valdes, J. A., & Hatcher, B. C. (2010). A new typology of benefit derived from marine protected areas. Marine Policy, 34, 635-644. [ Links ]
Baker, A. C., Glynn P. W., & Riegl, B. (2008). Climate change and coral reef bleaching: an ecological assessment of long-term impacts, recovery trends and future outlook. Estuarine, Coastal and Shelf Science, 80, 435-471. [ Links ]
Bruckner, A.W., & Hill, R. (2009). Ten years of change to coral communities off Mona and Desecheo Islands, Puerto Rico from disease and bleaching. Diseases of Aquatic Organisms, 87, 19-31. [ Links ]
Buddemeier, R.W., Lane, D.R., & Martinich, J. A. (2011). Modeling regional coral reef responses to global warming and changes in ocean chemistry: Caribbean case study. Climatic Change, 109, 375-397. [ Links ]
Burke, L., & Maidens J. (2004). Reefs at risk in the Caribbean. Washington, D.C., USA: World Resources Institute. [ Links ]
Burke, L., Reytar, K., Spalding M., & Perry, A. (2011). Reefs at Risk Revisited. Washington, D.C., USA: World Resources Institute. [ Links ]
Byrne, J. (2007). Grenada Gap Analysis. Christiansted, St. Croix, USVI. 25: The Nature Conservancy. [ Links ]
Cantin, N. E., Cohen, A. L., Karnauskas, K. B., Tarrant, A. M., &. McCorkle, D. C. (2010). Ocean warming slows coral growth in the central Red Sea. Science, 329, 322-325. [ Links ]
Convention on Biological Diversity. (2012). Action Plan for Implementing the Convention on Biological Diversity’s Programme of Work on Protected Areas (Grenada). Retrieved from http://www.cbd.int/protected/implementation/actionplans/country/?country=gd. [ Links ]
Crabbe, M. (2013). Coral Reef Populations in the Caribbean: Is There a Case for Better Protection against Climate Change? American Journal of Climate Change, 2(2), 97-105. [ Links ]
Crosby, M. P., & Reese E. S. (1996). A manual for monitoring coral reefs with indicator species: butterfly fishes as indicators of change on the Indo-Pacific reefs. NOAA, Silver Spring, Maryland, USA: Office of Ocean and Coastal Resource Management, [ Links ]
Eakin, C. M., Morgan J. A., Heron S. F., Smith T. B, Liu G., Alvarez-Filip, L., Baca, B., Bartels, E. Bastidas, C., …, & Yusuf, Y. (2010). Caribbean corals in crisis: record thermal stress, bleaching, and mortality in 2005. PLoS ONE, 5, e13969. [ Links ]
Fine, M. & Tchernov, D. (2007). Scleractinian coral species survive and recover from decalcification. Science, 315, 1811. [ Links ]
Gardner, T. A., Côte´, I. M., Gill, J. A., Grant, A., & Watkinson A. R. (2003). Long term region-wide declines in Caribbean corals. Science, 301, 958-960. [ Links ]
Graham, J. E., Barrett, N. S., & Stuart-Smith R. D. (2009). Exploited reefs protected from fishing transform over decades into conservation features otherwise absent from seascapes. Ecological Application, 19, 1967-1974. [ Links ]
Guarderas, A. P., Hacker, S. D., & Lubchenco, J. (2008). Current Status of Marine Protected Areas in Latin America and the Caribbean. Conservation Biology, 22, 1630-1640. [ Links ]
Humann, P. (1993). Reef Coral Identification (Florida, Caribbean, Bahamas). Jacksonville: New World Publications. [ Links ]
Jackson, J., Cramer, K. Donovan, M., Friedlander A., Hooten, A., & Lam, V. (2012). Tropical Americas Coral Reef Resilience Workshop - 2012. GCRMN Technical Report. Panama City, Panama: Smithsonian Tropical Research Institute. [ Links ]
Kelleher, G. (1999). Guidelines for Marine Protected Areas. IUCN - The World Conservation Union. Series 3. [ Links ]
Knowlton, N., & Jackson J. (2008). Shifting Baselines, Local Impacts, and Global Change on Coral Reefs. PLoS Biology, 6(2), e54. [ Links ]
Kohler, K. E., & Gill, S.M. (2006). Coral Point Count with excel extensions (CPCe): A visual basic program for the determination of coral and substrate coverage using random point count methodology. Computers & Geosciences, 32, 1259-1269. [ Links ]
Littler, M. M., & Littler, D. S. (2007). Assessment of coral reefs using herbivory, nutrient assays and indicator groups of benthic primary producers: a critical synthesis, proposed protocols, and critique of management strategies. Aquatic Conservation: Marine and Freshwater Ecosystems, 17, 195-215. [ Links ]
Nimrod, S. (2012). The effectiveness of Diadema in triggering a phase shift reversal. Mini-Symposia presentation 12th ICRS, Cairns, Australia. [ Links ]
Riegl, B., Bruckner, A., Coles, S., Renaud, P., & Dodge, R. E. (2009). Coral Reefs - Threats and conservation in an era of global change. Ann. NY Academic of Science, 1162, 136-186. [ Links ]
Riegl, B., Berumen, M., & Bruckner, A. (2013). Coral population trajectories, increased disturbance and management intervention: a sensitivity analysis. Ecology & Evolution, 3(4), 1050-1064. [ Links ]
Ries, J. B., Stanley S. M., & Hardie, L. A. (2006). Scleractinian corals produce calcite, and grow more slowly, in artificial Cretaceous seawater. Geology, 34, 525-528. [ Links ]
Sala, E., Costello, C., Dougherty, D., Heal, G., Kelleher, K., Murray, J., Rosenberg, A., & Sumaila, R. (2013). A General Business Model for Marine Reserves. PLoS ONE, 8(4), e58799. [ Links ]
Sandin, S. A., Sampayo E. M., & Vermeij, M. J. (2008). Coral reef fish and benthic community structure of Bonaire and Curaçao, Netherlands Antilles. Caribbean Journal of Science, 44, 137-144. [ Links ]
Selig, E. R., & Bruno, J. F. (2010). A Global Analysis of the Effectiveness of Marine Protected Areas in Preventing Coral Loss. PLoS ONE, 5(2), e9278. [ Links ]
Sokal, R. R., & Rohlf, F. J. (1995). Biometry: The principles and practice of statistics in biological research. 3rd edition. New York: W.H. Freeman. [ Links ]
Sprung, J. (1999). Oceanographic Series Corals: A Quick Reference Guide. Miami: Ricordia Publishing. [ Links ]
Turner, M. (2009). Draft Grenada Protected Area System Plan Part 1 - Identification and Designation of Protected Areas. Prepared for the Environment and Sustainable Development Unit (ESDU) of the Organization of Eastern Caribbean States (OECS) Protected Areas and Associated Livelihoods (OPAAL) Project. [ Links ]
Wiedenmann, J., D’Angelo, C., Smith, E. G., Hunt, A. N., Legiret, F. E., Postle, A. D., & Achterberg, E. P. (2013). Nutrient enrichment can increase the susceptibility of reef corals to bleaching. Nature and Climate Change, 3, 160-164. [ Links ]
1. Wisconsin Lutheran College, 8800 W. Bluemound Rd., Milwaukee, WI 53226, USA; banderson@wlc.edu, ebolda@hotmail.com, katie.musser@mail.wlc.edu, rob.balza@wlc.edu
2. St. George’s University, P.O. BOX 7, St. George’s, Grenada, West Indies; cmorrall@sgu.edu, snimrod@sgu.edu
3. University of the Virgin Islands; jossart1@gmail.com
4. Milwaukee County Zoo, 10001 W Bluemound Road, Milwaukee, WI 53226, USA; craig.berg@milwcnty.com
Received 07-IX-2013 Corrected 23-II-2014 Accepted 24-III-2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}