










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.3 San José Sep. 2014
The planktonic communities of the Jamaican south-east coast; a comparison of harbor, shelf and oceanic areas
Comunidades plactónicas de la costa sudoriental Jamaiquina; una comparación de puerto, estante y áreas oceánicas
Abstract
Few studies have compared water quality and plankton along the eutrophication gradient from Kingston Harbour to oceanic waters around Jamaica. To compare the planktonic community along the expected nutrient gradient, we sampled every two weeks at four stations, from eutrophic Kingston Harbour to oceanic California Bank. Phytoplankton was assessed from whole water Niskin bottle casts and zooplankton by vertical hauls with plankton nets of three different mesh sizes: 64µm, 200µm, and 600µm. Total phytoplankton biomass declined sharply away from the harbour (1.0 μg L-1 at the Harbour Shoal Beacon to 0.2 μg L-1 at California Bank). Characteristic estuarine phytoplankton genera -such as Ceratium, Gonyaulax, Gyrodinium and Rhizosolenia- dominated harbour samples while genera characteristic of offshore locations -such as Asterionelliopsis, Navicula, Nitzschia, Rhizosolenia and Thalassionema- dominated California Bank. Highest phytoplankton densities (mean values of 34 174 cells L-1) were found at the harbor mouth. Mean zooplankton abundances ranged from maximum (5 858.5m-3) at Beacon to minimum (2 124.2 m-3) at California; 171 species of zooplankton were identified and copepods dominated with 76 species. Overall, 75 species of zooplankton were identified from Beacon, 107 from Port Royal Cays- South East Cay, 110 from the exposed shelf edge- Windward Edge, and 95 from the oceanic California Bank. Larval forms dominated; copepod nauplii, fish eggs and echinoderm larvae occurred at all sites. Lucifer faxoni and Penilia avirostris were indicative of harbor waters and Microsetella sp. and Farranula carinata of offshore waters. Some zooplankton taxa, like L. faxoni, Paracalanus parvus and copepod nauplii, despite showing gradual decline with distance from Beacon to the Edge, increased in abundance at the furthest station, California. California Bank clearly experiences enrichment which at times can be as high as near-shore areas, but the planktonic community is unique and similar only to those at other offshore station. To be reliable, characterization of water masses must involve a range of physicochemical and biological parameters.
Key words: Plankton, Water quality, Kingston Harbour, South-east coast, Jamaica.
Resumen
Pocos estudios han comparado la calidad de agua y plancton a lo largo de un gradiente de eutrofización en el Puerto Kingston con aguas oceánicas en Jamaica. Para comparar la comunidad de plancton a lo largo del gradiente de nutrientes esperado, muestreamos cada dos semanas en cuatro estaciones, desde el eutrófico Puerto Kingston hasta el Banco oceánico California. Evaluamos el fitoplancton de agua entera en botellas de molde tipo Niskin usando la biomasa total y y zooplancton por arrastres verticales con redes de plancton de tres tamaños de malla diferente: 64µm, 200µm y 600µm. La biomasa total de fitoplancton mostró una fuerte caída al aumentar la distancia desde el puerto Kingston (1.0µg L-1 en el Puerto Shoal Beacon a 0.2µg L-1 en el Banco California). Géneros característicos estuarinos como Ceratium, Gonyaulax, Gyrodinium y Rhizosolenia dominaron en las muestras del Puerto Kingston mientras que géneros característicos de costa afuera como Asterionelliopsis, Navicula, Nitzschia, Rhizosolenia y Thalassionema dominaron en el Banco California. Las densidades más altas de fitoplancton (valores promedio de 34 174 células L-1) se encontraron en la boca del puerto. Valores promedio de zooplancton oscilaban entre un máximo (5 858.5m-3) en el Puerto Shoal Beacon (HSB) a un mínimo (2 124.19m-3) en el Banco California (CB), 171 especies del zooplancton fueron identificadas, de los cuales los copépodos dominaban con 76 especies. En general, se identificaron 75 especies del zooplancton en HSB, 107 de los Cayos Port Royal (SEC), 110 del borde de la plataforma expuesta (WE) y 95 del Banco oceánico (CB). Las formas larvarias eran numéricamente dominantes con nauplios de copépodos, huevos de peces y larvas de equinodermos que ocurren en todos los sitios. La dominancia de especies individuales fue utilizada para caracterizar las masas de agua. Lucifer faxoni y Penilia avirostris fueron indicativos de las aguas del puerto y Microsetella sp. y Farranula carinata fueron identificados como indicadores de aguas costa afuera. Taxones seleccionados de de zooplancton, como L. faxoni, Paracalanus parvus y nauplios de copépodos, a pesar de mostrar un decline gradual con la distancia desde el puerto de Kingston a la otra estación costa afuera, aumentan su abundancia en la estación más lejana, Banco California. Banco California es claramente una zona que experimenta un enriquecimiento que a veces puede ser tan alto como las áreas cerca de la costa, pero la comunidad planctónica es única y similar únicamente a las estaciones costa afuera. Para ser confiable, la caracterización de masas de agua debe implicar una serie de parámetros físico-químicos y biológicos.
Palabras clave: Plancton, costa sur-este, calidad de agua, Puerto Kingston, Jamaica.
There is now a great deal of information concerning the planktonic communities of the Kingston Harbour and the nearby coastal shelf areas (Moore & Sander, 1979; Webber & Roff, 1996; Webber & Webber, 1998; Dunbar & Webber, 2003; Campbell, Manning, Webber & Webber, 2008). Some of these studies have been quite extensive covering distances from Kingston Harbour to as far as 5km to the shelf edge (Moore & Sander, 1979) or parallel to the coast for 15km (Lindo, 1991). Other studies have covered extensive areas of the Port Royal cays (Webber et al., 1996; Webber & Roff, 1996, Persad, Hopcroft, Webber & Roff, 2003) in an effort to indicate the extent of Kingston Harbour influence and to identify indicator species of the water masses in the area. In most cases, the Kingston Harbour has been cited as a source of eutrophication with polluted waters leaving the mouth and being diluted by oceanic waters as the distance increased. However, it is believed that the outflow from Kingston Harbour, mainly affects the Hellshire coastline and proximal areas of the Port Royal cays and so progressively improved water quality and pristine conditions should be experienced with increasing distance form Kingston Harbour and the south coast shelf.
Few studies have adequately compared water quality and plankton along this eutrophication gradient from Kingston Harbour to oceanic waters around Jamaica. Indeed, the only previous study was conducted in 1964 and compared the zooplankton community and water quality between Kingston Harbour, the Port Royal Cays and an offshore station located 5 km away from the coastline of Jamaica (Moore & Sander, 1979). Moore and Sander (1979) indicated a gradient of conditions including, increased zooplankton abundances, phytoplankton biomass and nutrients across the stations towards the harbor. The increase in zooplankton abundances and phytoplankton biomass observed was found to be due to various factors: upwelling, run-off of nutrient rich waters, freshwater and local current dynamics. These factors had the effect of enhancing the nutrient levels thereby increasing productivity.
The area beyond the southern Jamaican shelf although influenced by predominantly oceanic sources, Grant and Wyatt (1980), is very productive in some locations. These locations include the offshore banks, e.g. California Bank and the edges of the Jamaican shelf and are considered as a moderate fisheries resource (Munro, 1983).
The paucity of studies comparing oceanic, shelf and harbor waters on the south coast is due primarily to the extent of the south coast shelf (28km at its widest point) and the local unavailability of vessels capable of safely embarking on voyages to areas far offshore. Compounding this problem is the difficulty in the actual data collection and processing while offshore. In general, work on water quality and plankton in Jamaica’s offshore locations has been limited to occasional cruises on large scientific vessels or to the opportunistic trips on Jamaican Coast Guard vessels. Thus these voyages provide inadequate descriptions of the offshore planktonic communities and little opportunity for contemporaneous collections in near-shore waters that would allow for adequate examination of an inshore to offshore gradient.
The primary aim of this investigation was therefore to compare water quality and plankton along a gradient from expectedly pristine areas well beyond the shelf with areas close to shore and ultimately high nutrient areas like Kingston Harbour. The study was intended to:
1. Identify and confirm a eutrophication gradient between Kingston Harbour, the Port Royal Cays and offshore areas,
2. Compare the planktonic communities along the gradient and
3. Identify planktonic indicator species or communities characteristic of the areas along the gradient.
Materials and Methods
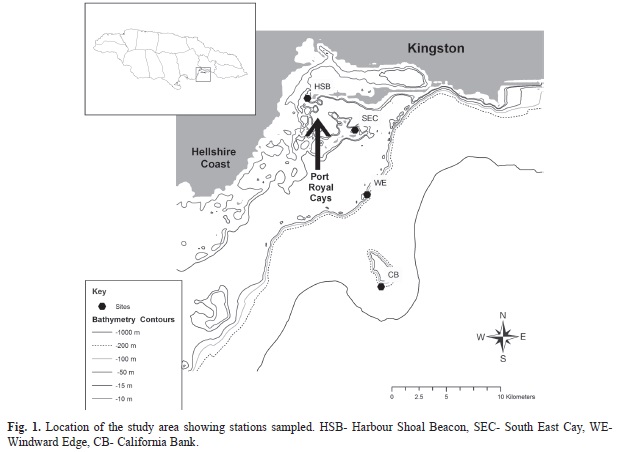
Study sites: Four sites were selected (Fig. 1) to represent four contrasting marine environments with increasing linear distance from Kingston Harbour (Table 1). The linear distance represented a nutrient gradient from the eutrophic Kingston Harbour to oligotrophic California Bank (furthest away). The first site (Harbour Shoal Beacon - HSB) was located at the mouth of Kingston Harbour which has been characterized as eutrophic since the 1970’s (Goodbody, 2003; Wade, Antonio & Mahon, 1972). Kingston Harbour receives agricultural runoff and partially treated sewage effluent which serve as a source of nutrient enrichment. The second site (South East Cay - SEC) was located 5 km from HSB. This location represents a coral cay and is one of a number of cays on the south coast shelf of Jamaica referred to as the Port Royal Cays. The Port Royal Cays are important for near-shore fishery and recreational sites for locals and visitors (Steele, 1998). The third location (Windward Edge - WE) was located 7km from the SEC, on the south-coast shelf edge and as such is exposed to oceanic influences. The coral reefs of this location are deep, patchy and often senescent (Munro, 1983). The fourth sampling location (California Bank - CB) was the furthest being ten 10km from the third site (WE) or 22km from Kingston Harbour. California Bank represents a small offshore bank, 8.6km2 in area with immediate surrounding waters of approximately 700km depth. The water depth on the bank ranges between 35-45m. Munro (1983) also describes the reefs of this location to be sparse in coral cover and with sills that are not well developed.
Sampling programme: Sampling was carried out every two weeks from April to December 2004. Table 1 has the G.P.S coordinates of the stations that were occupied for sampling. On each occasion, sampling commenced from the station furthest offshore (California Bank) and progressed towards the more eutrophic station (Harbour Shoal Beacon). All four stations were sampled between 0830 and 1300 hours on the same day.
Water quality and phytoplankton assessment: Physical and chemical parameters including: temperature, salinity, oxidative redox potential, turbidity and light were collected in situ on each sampling occasion using a hydro-lab multi-parameter probe. Light values at specific depths (starting from 1.0m) were used to calculate the extinction coefficient (Poole & Atkins, 1929). Sampling from California Bank, Windward Edge and South East Cay was done at 5m intervals starting at the surface (0.1m) to a maximum depth of 20m. However, collections from Harbour Shoal Beacon were done at 1 m intervals starting at the surface to a maximum of 5m due to the shallow nature of that site.
Whole water samples, collected using a 6L Niskin water bottle, were used to determine total and size fractionated phytoplankton biomass, phytoplankton species composition and abundance as well as nitrate and phosphate concentrations. Whole water samples from California Bank, Windward Edge and South East Cay were collected with the Niskin bottle at depths of 20m, 10m and at the surface 0.5m; while at Harbour Shoal Beacon samples were collected from 5m and 0.5m depths. Phytoplankton samples were kept in the dark to protect them from light shock and processed within a maximum holding time of 5h (Parsons, Maita & Lalli, 1984).
A 250mL portion of each sample was immediately preserved (poured directly from the Niskin bottle into 250mL plastic bottle) containing 2.5mL of acidized Lugol’s solution for counting and identification while the remainder of the sample was filtered through Nalgene size fractionating tower into three size classes. Separation was done using the appropriate size filters as follows: Nitex screening - 20µm; Whatman glassfibre filters, GDF - 2.7µm and Whatman glassfibre filters, GDF - 0.7µm.
Chlorophyll a content was determined using 6mL of 90% acetone to extract the chlorophyll (Arar & Collins, 1997). The quantity of Chlorophyll a in the extract was determined using a Turner Design Fluorometer TD700. Lugol’s preserved phytoplankton samples were gently homogenized and 100mL portions used to fill settling chambers which were allowed to stand for 24 hours before examination using a Carl Zeiss inverted microscope with phase contrast (Model # 401672). Where cell numbers were low, the entire sample was enumerated (e.g. from California Bank, Windward Edge and South East Cay). However, cells from 30 random fields of view (Paerl, 1978) were counted from the Harbour Shoal Beacon. The number of phytoplankton genera and the total number of cells per liter were determined from the analysis.
Nitrates and phosphate were determined from the filtrate produced after the phytoplankton filtering. The filtrate was frozen as soon as possible after collection. Nitrate values were determined using a modified cadmium reduction column method. Phosphate analysis was done using the Molybdate Colorimetric method (Parsons et al., 1984). Ammonium was not determined as the detection limit of the methods (APHA-AWWA-WPCF, 1980) was exceeded by background values, thus making sample values negligible and very unreliable.
Zooplankton assessment: SCOR, WP2 pattern plankton nets (UNESCO, 1968) of three different mesh sizes: 600µm, 200µm and 64µm were used to collect zooplankton samples in replicate (n=2) vertical hauls. The hoop diameter of the 600µm net was 1 m while the diameter of the 200µm and 64µm net was 0.5. Samples were collected from a depth of 20m from California Bank, Windward Edge and South East Cay but from a 5m depth at Harbour Shoal Beacon. The net was hauled at a speed of approximately 0.5ms-1 using a manual winch. Plankton nets were washed between stations to prevent artificial transfer of species.
Zooplankton samples were fixed upon collection by pouring the contents of the cod end into 1 l containers containing 10mL of full strength formalin. All samples were later preserved in 10% formalin. Zooplankton species identification and enumeration were conducted on a sub-sample obtained using a modification of the beaker split method (Van Guelphen, Markle & Duggan, 1982). The sub-sample was transferred into a Bogorov tray and analyzed using a Wild (M7) binocular microscope. Zooplankton organisms were enumerated and identified to genus and species where possible using guides including: Owre and Foyo (1967), Newell and Newell (1977), Michel (1984) and Todd, Laverack & Boxshall (1996).
Results
A total of 10 physicochemical, five phytoplankton and 13 zooplankton parameters were collected and analyzed each as 105-108 discrete samples during this investigation. All data were assessed for normal distribution using STATISTICA 7 and Log (x +1) transformations applied where necessary. Temporal differences were not significant (MANOVA, p>0.05) but spatial variability was significant for eight of the physicochemical, all of the phytoplankton (size fractionated biomass (pico, nano and netplankton), abundance and richness, and eight of the zooplankton parameters (total abundance, species richness, abundance of copepods, calanoids, larvae, decapods, cladocerans and cnidarians). All significant variables were used in stepwise variable selection multiple regression models.
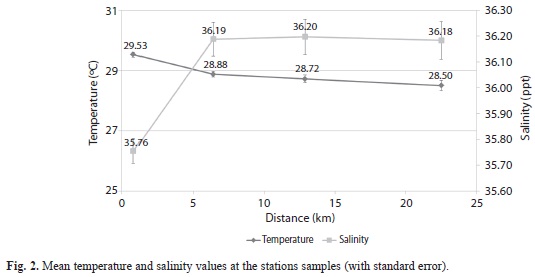
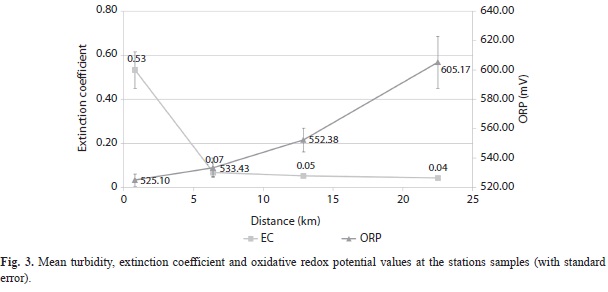
Physicochemical parameters: Mean temperature values showed a slight decrease with increasing distance from Harbour Shoal beacon (HSB). Values ranged from 29.53oC at HSB to 28.50oC at California Bank (Fig. 2). Nevertheless, temperatures were significantly different between the stations (MANOVA, P=0.003). Mean salinity values (MANOVA, P < 0.001) increased sharply from Harbour Shoal Beacon (35.76o/oo) to South East Cay (36.19o/oo) and remained almost constant as distance increased from South East Cay (SEC) to offshore stations (Fig. 2). Significant differences in turbidity values were not observed (MANOVA, P=0.165) and there was no trend in the distribution of turbidity as distance increased. However, extinction coefficient (MANOVA, P< 0.001) initially declined from 0.53 at Harbour Shoal Beacon to 0.068 at South East Cay, before gradually decreasing as distance offshore increased, reaching 0.044 at California Bank (Fig. 3).
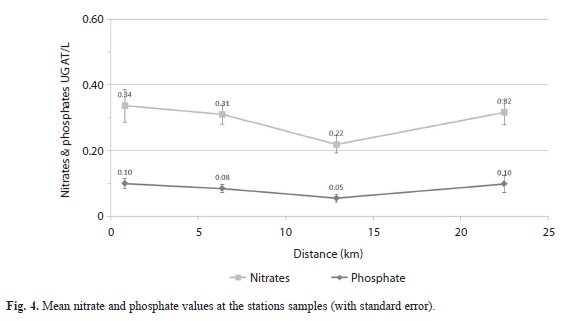
Oxidative redox potential (ORP) values (MANOVA, p = 0.011) increased as distance offshore increased (Fig. 3) but Phosphate (MANOVA, P = 0.027) showed similar values at the Harbour Shoal Beacon as at California Bank. Phosphate values ranged from a low of 0.055 µg at L-1 at Windward Edge to a high of 0.099µg at L-1 at Harbour Shoal Beacon with California Bank being a close second with high values of 0.098µg at L-1 (Fig. 4). However, nitrate values showed no significant differences between stations (MANOVA, P = 0.079).
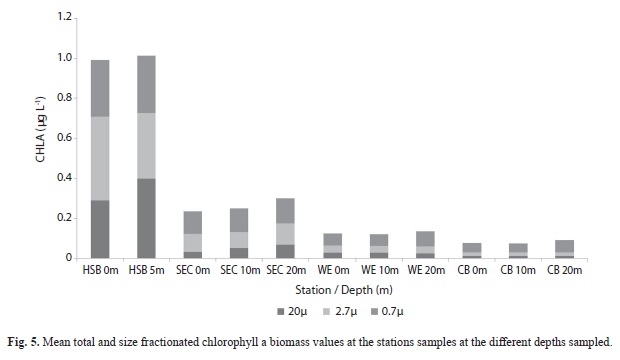
Phytoplankton: A severe decline in mean chlorophyll a total biomass (MANOVA p = < 0.001) was seen from Harbour Shoal Beacon (1.25µg L-1) to South East Cay (0.25µg L-1). A more gradual decline was observed towards Windward Edge (0.2µg L-1) then to California Bank (0.125µg L-1). Also as depth increased, chlorophyll a values showed small increases at all stations and were found to not be significant (Fig. 5).
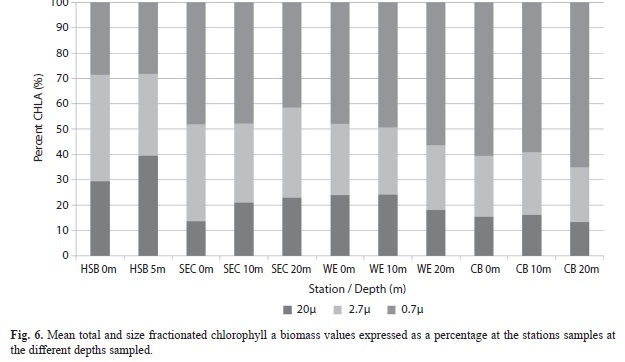
Examination of the proportion contributed by each phytoplankton size fraction (Fig. 6) showed that the picoplankton (0.7µm) contributed the dominant portion of the biomass at California Bank, Windward Edge and South East Cay while the netplankton (20µm) dominated only at Harbour Shoal Beacon. Generally the picoplankton fraction increased and netplankton decreased with distance from Kingston Harbour. The nanoplankton fraction (2.7µm) showed the least pattern across all stations (Fig. 6). Chlorophyll a values for all size classes of phytoplankton showed significant spatial variation (MANOVA P <0.001) after log transformation.
Stepwise, variable selection multiple regression tests showed that 56% of the variation in netplankton was related to variation in distance, temperature, pH and dissolved oxygen (R2=55.75%, P<0.005), while 72% of the variation in nanoplankton was related to variation in distance, temperature, specific conductivity and phosphates (R2=71.38%, P<0.005). Variation in distance, dissolved oxygen and salinity were attributable for 54% of the variation in picoplankton (R2=53.29, P<0.005).
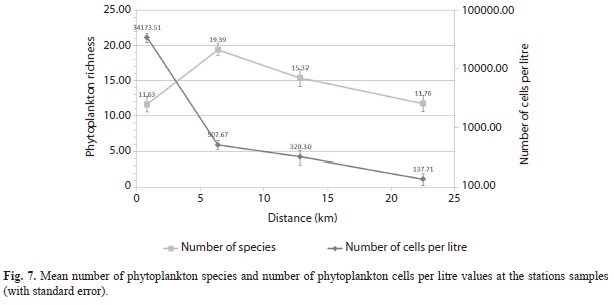
The number of phytoplankton taxa (richness) increased from Harbour Shoal Beacon (mean of 12) to South East cay with a mean of 19 (MANOVA, P < 0.001). However, as distance increased further offshore a general decline was then observed with Windward Edge having a richness of 15 genera, the same as California Bank (Fig. 7). Stepwise variable selection multiple regression showed that variations of 13% in the number of phytoplankton genera could be directly attributable to variation in nitrates and dissolved oxygen (R2=12.58%, P<0.05), with both having a negative relationship to richness.
Phytoplankton abundance (# of cells l-1) followed a similar spatial distribution pattern (MANOVA, P<0.001) where a severe decrease was seen from Harbour Shoal Beacon (34 173.51 cells l-1) to South East Cay (507.67 cells l-1). A gradual decrease was then obtained from South East Cay as distance offshore increased. Windward Edge had mean abundance values of 320.30 cells l-1 while California Bank had 137.71 cells l-1 (Fig. 6). Stepwise variable selection multiple regression showed that 61% of the variation in number of cells was attributable to distance and total dissolved solids (R2=60.79%, P<0.05), both having negative relationships to the number of cells l-1.
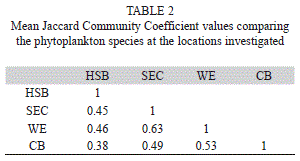
Jaccard community coefficient (JCC) analysis indicated that the communities at Harbour Shoal Beacon and South East cay were 45% similar, Harbour Shoal Beacon and Windward Edge were 46% similar while Harbour Shoal Beacon and California Bank were 38% similar. South East Cay and Windward Edge were 63% similar and South East Cay and California Bank 49% similar. Finally, phytoplankton communities at Windward Edge and California Bank were 53% similar (Table 2).
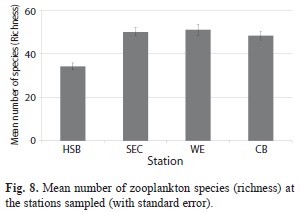
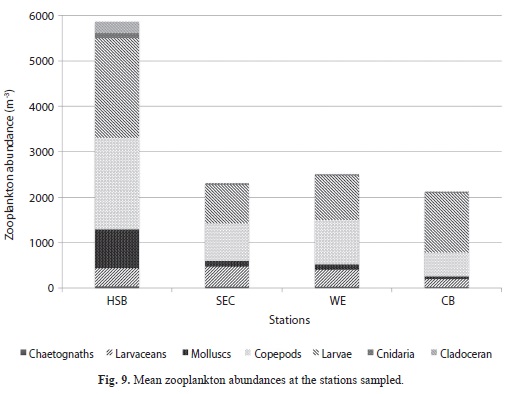
Zooplankton species richness and total abundances: The overall number of zooplankton species encountered during the sampling period (total richness) ranged from 81 species at Harbour Shoal Beacon (HSB) to 115 species at Windward Edge (MANOVA, P<0.001). HSB also consistently had the lowest mean richness (Fig. 8). Contrastingly, HSB had the greatest mean zooplankton abundance (Nos. m-3) when compared to the other three stations (MANOVA, P<0.001). Mean zooplankton abundance at HSB was 5 963 individuals m-3 while South East Cay (SEC), Windward Edge (WE) and California Bank (CB) had abundance values of approximately 2 000 individuals m3 (Fig. 9).
When all net collections were combined, larvae (invertebrate and vertebrate) were the most abundant group being the greatest proportion of the collections from all four stations. Larvae accounted for more than half of the total abundance at CB (Fig. 9). Of the larval group, copepod nauplii were numerically the most dominant taxon. These occurred universally across all stations and sampling occasions with highest numbers observed at HSB followed by CB. The other two stations (SEC and WE) showed similar low numbers. Adult copepods were the second most abundant taxonomic group (after larvae). Cnidarians made the smallest contribution at each station, but were still most abundant at Harbour Shoal Beacon.
Potential zooplankton indicator species were examined in terms of their relative abundance across the four stations. Species like Penilia avirostris and Lucifer faxoni had been identified as Kingston harbor indicators (Lindo, 1991; Webber, Roff, Chisholm & Clarke, 1996). P. avirostris showed greatest abundance at Harbour Shoal Beacon (Table 4) and then declined exponentially to the stations further offshore (MANOVA P > 0.001). L. faxoni was similar with maximum abundances at HSB, followed by a severe decline at SEC and WE. However, abundances showed a slight increase at CB, above values recorded at SEC and WE.
Microsetella sp. occurred at all four stations investigated and abundances gradually increased with distance (MANOVA, P<0.001) from the Harbour Shoal Beacon, where mean abundances were less than 5 individuals m-3. However it was present as 30 and 37 individuals m-3 at Windward Edge and California Bank, respectively. F. carinata (MANOVA, P= 0.001) seemed to have similar distribution (increase in the numbers of this species as distance offshore increased) but it was only observed at South East Cay, Windward Edge and California Bank. It was not observed at Harbour Shoal Beacon over all the sampling occasions.
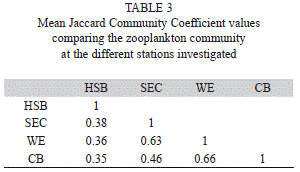
Using the Jaccard Community Coefficient (JCC), the zooplankton community at the stations increased in similarity from Harbour Shoal Beacon (HSB) towards California Bank (CB). HSB was the least similar compared to the other stations with similarities of 38% and less (Table 3). The highest community similarities were observed between CB and WE (66% similarity). Communities at South East Cay (SEC) and Windward Edge (WE) showed 63% similarity while SEC and CB were 46% similar (Table 4). This pattern was almost identical to the phytoplankton community.
Discussion
Comparison of water quality and plankton across areas spanning coastal bays or harbors and offshore, oceanic banks are rare as intensive plankton and water quality sampling requires relatively short intervals between collecting and processing events. Alternatives include availability of large enough vessels for shipboard analysis or restricting attention to few parameters (Moore & Sander, 1979). This study achieved comparative analysis of 11 physicochemical parameters, 13 zooplankton parameters and five phytoplankton parameters sampled from different depths at stations up to 22km apart across an expected eutrophication gradient.
Thus it was expected that the physicochemical parameters would show severe changes from the eutrophic waters at Harbour Shoal Beacon (HSB) at the mouth of Kingston harbor to South East Cay (SEC), Windward Edge (WE) and California Bank (CB) 22km away, along the predicted eutrophication gradient. However, with the exception of Extinction coefficient and Salinity, the change in water quality parameters with increasing distance off shore was not as severe as expected. In fact the gradually declining nutrient gradient was disrupted by values at the furthest station (CB) being higher than those from its next nearest neighbor. This pattern was also demonstrated for the zooplankton parameters: species richness, numbers of L. faxoni, Paracalanus parvus and copepod nauplii. Values at California bank for these parameters indicated unexpected enrichment of the area. Offshore banks and shelf areas may receive enrichment from upwelling which is aided by horizontal circulation patterns like gyres that concentrate nutrients and retain planktonic communities (Genin, 2004; Rakhesh, Raman & Sudarsan, 2006). Such circulation is said to lead to increased numbers in especially larval forms, phytoplankton and other plankton with limited means of movement (Carleton, Brinkman & Doherty, 2001).
While it was unexpected for the offshore bank to have nitrate values as high as at the mouth of Kingston Harbour, upwelling has been shown to promote significant enrichment of surface waters (Caldeira et al., 2002). Nevertheless, it is important to note that nutrient values between the eutrophic Kingston harbor and relatively pristine offshore sites were not as different as expected; clearly indicating the deficiency in relying on nutrients as indicators in such conditions.
The identification of indicator species and groups from the plankton was more reliable at characterizing different water masses (McGehee, Derner & Warren, 2004; Fehling et al., 2012; Tseng, Dahms, Chen & Hwang, 2013). Netplankton (>20µm) is known to be dominant in high nutrient waters because of the low surface area to volume ratio causing them to be best suited and therefore allowing them to persist in high nutrient concentrations (Hopcroft, 1988; Webber, 1990), such as is available at HSB. Contrastingly, the picoplankton size class, having a high surface area to volume ratio would dominate oligotrophic areas such as South East Cay, Windward Edge and California Bank (Hopcroft, 1988; Webber, 1990). The last size fraction, the nanoplankton, are known to remain fairly consistent whether near or far from the coast (Vant & Safi, 1996). This is corroborated by the findings of Webber and Roff (1996) where small picoplankton dominated nutrient poor oceanic waters and netplankton in coastal waters nutrient rich waters.
The taxonomic richness (number of zooplankton species and phytoplankton genera) did not follow the expected gradient as lowest number of species occurred at both Harbour Shoal Beacon and California Bank. Furthermore, while total abundances (zooplankton Nos. m-3 and Chlorophyll a L-1) followed the expected gradient, individual species and groups such as copepod nauplii showed secondary increases at the station located furthest offshore due to enrichment associated with the changes in circulation associated with the submerged bank.
While abundances may be unpredictable, community composition has been shown to be better when identifying distinct water masses (Campbell et al., 2008). Thus greater percentage similarity was expected and obtained from stations closer to each other along the gradient. Even in cases where the nutrient concentrations were similar, (e.g. Harbor Shoal Beacon and California Bank), the planktonic communities were most different and reflected the differing conditions of the associated water mass. This is also supported by the dominant species (indicators) in each water body. In terms of the phytoplankton, coastal and oceanic water bodies were separated into two general groups with, the coastal stations having dinoflagellattes (also shown by Matsuoka, Joyce, Katoni & Matsuyma, 2003) such as: Gonyaulax, Gyrodinium and Perdinium being dominant while oceanic stations having diatoms such as: Navicula, Nitzschia, Rhixosloenia and Thallassionema being dominant; irrespective of nutrient levels.
Throughout the study, zooplankton groups were dominated by copepods and larvae which accounted for more than 70% of the abundances at each station. However, copepods were expected to be the dominant group as was previously reported by other authors (Youngbluth, 1980; Moore & Sander, 1979; Webber & Roff, 1995; Dunbar & Webber, 2003). The larval group contained representatives from different groups notably copepod nauplii, fish eggs and echinoderms. The high larval presence at the offshore stations (California Bank and Windward Edge), was further evidence that they are productive areas with reduced flushing and water retention processes that lead to increased numbers of especially larval forms with limited means of movement (Cowen & Castro, 1994; Carleton et al., 2001).
Within each group/taxa a few species displayed high abundances and occurrence at the stations and therefore have the potential for use as indicators of different water masses/conditions. L. faxoni has been identified as an indicator of Kingston Harbour waters in previous studies (Lindo, 1991; Webber et al., 1996; Dunbar & Webber, 2003). L. faxoni, clearly displayed highest numbers at the Harbour station but did not have lowest mean values at the furthest offshore station. The distribution of the species appears to be less affected by salinity levels and more by nutrients.
The Cladoceran, P. avirostris has often been paired with L. faxoni. As shown in the present investigation, it also tends to have highest numbers at the harbor station and low numbers at offshore areas. P. avirostris has been previously reported in high abundances in the Kingston Harbour and at its mouth (Moore & Sander, 1979; Dunbar & Webber, 2003; Webber et al., 2005). The distribution of the species appears to be affected by salinity concentrations and food availability. Moraitou-Apostolopoulou and Kiortsis (1973) found that higher salinities and pelagic nature of the waters had an effect on P. avirostris’ distribution, thereby limiting its presence to areas with shallow low-salinity waters.
The copepod Microsetella sp. was the dominant harpacticoid at the offshore stations and Moore and Sander (1979) reported this species as only occurring at stations outside of the Harbour and while Webber et al. (1996) did not report it from their study of the Port Royal Cays, it was found offshore Discovery Bay, north coast of Jamaica (Webber & Roff 1995). The cyclopoid, F. carinata was important at the offshore stations, while being absent from the Harbour Shoal Beacon. This species has also been found to be important in oceanic waters off the north coast at Discovery Bay, Jamaica by Webber & Roff (1995). Microsetella sp. and F. carinata therefore could be indicators of offshore waters.
Larvae followed the general trend of highest numbers at the Harbour mouth; however the numbers at station California Bank were second highest. Moore and Sander (1979) had reported relatively low mean numbers (135m-3) at their offshore station, a magnitude lower than reported by the present study. In using a single mesh (~200µm) for their study they would have underestimated copepodites by a factor of three to six, and also underestimated copepods by a factor of five (Webber & Roff, 1995). The relatively high larval abundance at California Bank would suggest a mechanism or processes that retain larvae associated with this area. Similar effects have been seen by Heywood, Barton & Simpson (1990) and Hernández-León (1990), where they found an increase in the biological production and attributed it to the presence of an offshore bank.
In general there was a change in biological parameters with increasing distance from the harbor towards the open sea. Based on the indices investigated, the identification of a gradient of conditions with different levels of eutrophication was partially achieved with confirmation that Harbour Shoal Beacon was the most eutrophic site and Windward Edge most oligotrophic. South East Cay, due to its position on the south coast shelf, should be classified as mesotrophic (Vollenweider et al., 1998) along with California Bank. California Bank while having oceanic planktonic communities, is not oligotrophic, but is influenced by processes that reflect mesotrophic conditions hence its pairing with South East Cay. The findings of this study indicate that accurate characterization of water masses and their trophic status must involve a range of physicochemical and biological parameters.
References
APHA-AWWA-WPFC. (1980). Standard methods for examination of water and wastewater. 15th ed. New York: Public Health Association Inc. [ Links ]
Arar, J. E., & Collins, G. B. (1997). In vitro determination of chlorophyll a and pheophytin a in marine and freshwater algae by fluorescence. National Exposure Research Laboratories Office of Research Development. USEPA Method 445.0. [ Links ]
Caldeira, R. M. A., Gordon, S., Miller, P., Pilgrim, D., & Nezlin, N. P. (2002). Sea-surface signatures of the island mass effect phenomenon around Madeira island, northeastern Atlantic. Remote Sensing of Environments, 80, 336 - 360. [ Links ]
Campbell, P., Manning J., Webber, M., & Webber, D. (2008). Planktonic communities as indicators of water quality in mangrove lagoons: a Jamaican case study. Transitional Waters Bulletin, 3, 39 - 63. [ Links ]
Carleton, J. H., Brinkman, R., & Doherty, P. J. (2001). Zooplankton community structure and water flow in the lee of Helix reef (Great Barrier Reef, Australia). Marine Biology, 139, 705-763. [ Links ]
Cowen, R. K., & Castro, L. R. (1994). Relation of coral reef fish larval distributions to island scale circulation around Barbados, West Indies. Bulletin of Marine Science, 54, 228-244. [ Links ]
Dunbar, F. N., & Webber, M. K. (2003). zooplankton distribution in the eutrophic Kingston Harbour, Jamaica. Bulletin of Marine Science, 73, 343-359. [ Links ]
Fehling, J., Davisdon, K., Bolch, C., Brand, T., & Narayanaswamy, B. (2012). The relationship between phytoplankton distribution and water column characteristics in north western European shelf sea waters. PLoS ONE, 7(3), e34098. doi:10.1371/journal.pone.0034098. [ Links ]
Genin, A. (2004). Bio-physical coupling in the formation of zooplankton and fish aggregations over abrupt topographies. Journal of Marine Systems, 50, 3-20. [ Links ]
Goodbody, I. (2003). Kingston harbor Jamaica: an overview. Bulletin of Marine Science, 73, 249 - 255. [ Links ]
Grant, C. J., & Wyatt, J. R. (1980). Surface currents in the eastern Cayman and western Caribbean seas. Bulletin of Marine Sciences, 30, 613 - 622. [ Links ]
Hernández-León, S. (1990). Accumulation of mesozooplankton in a wake area as a causative mechanism of the “island-mass effect”. Marine Biology, 109, 141-147. [ Links ]
Heywood, K. J., Barton, E. D., & Simpson, J. H. (1990). The effects of flow disturbance by an oceanic island. Journal of Marine Research, 48, 55 -73. [ Links ]
Hopcroft, R. R. (1988). Size fractionated biomass and primary production of tropical neretic phytoplankton near Kingston Jamaica. M.Sc. Thesis, University of Guelph, Jamaica. [ Links ]
Lindo, M. (1991). The effect of Kingston Harbour outflow on the zooplankton populations of Hellshire, south-east coast, Jamaica. Estuarine, Coastal and Shelf Science, 32, 597 - 608. [ Links ]
Matsouka, K., Joyce, L. B., Katoni, Y., & Matsuyma, Y. (2003). Modern dinoflagellate cysts in hypertrophic coastal waters off Tokyo Bay, Japan. Journal of Plankton Research, 25, 1461-1470. [ Links ]
McGehee, D. E., Derner, D. A., & Warren, J. D. (2004). Zooplankton in the Ligurian Sea: Part 1. Characterization of their dispersion, relative abundance and environment during summer 1999. Journal of Plankton Research, 26, 1409 - 1418. [ Links ]
Michel, H. B. (1984). Chaetognaths of the Caribbean sea and adjacent waters. NOAA Technical Report of the North-East Marine Fisheries Service, 15, 25. [ Links ]
Moore, E., & Sander, F. (1979). A comparative study of zooplankton from oceanic, shelf, and harbour waters of Jamaica. Biotropica, 11, 196-206. [ Links ]
Moraitou-Apostolopoulou, M., & Kiortsis, V. (1973). The Cladocerans of the Aegan Sea: Occurrence and seasonal variation. Marine Biology, 20, 137-143. [ Links ]
Munro, J. C. (1983). Caribbean coral reef fishery report. ODA/ UWI Fisheries Ecology Research Project 1969-1973. [ Links ]
Newell, G. E., & Newell, R. C. (1977). Marine plankton: a practical guide. London: Huthinson and Co. Ltd. [ Links ]
Owre, H. B., & Foyo, M. (1967). Copepods of the Florida current. Fauna Caribaea No. 1.Crustacea, Part1: Copepoda. Miami: Institute of marine science University of Miami. [ Links ]
Paerl, H. W. (1978). Effectiveness of various counting methods in detecting viable plankton. Journal of Marine and Freshwater Research, 12, 67-72. [ Links ]
Parsons, T. R., Maita, Y., & Lalli, C. M. (1984). A manual of chemical and biological methods for seawater analysis. UK: Pergamon Press. [ Links ]
Persad, G., Hopcroft, R. R., Webber, M. K., & Roff, J. C. (2003). Abundance, biomass and production of ctenophores and medusae off Kingston, Jamaica. Bulletin of Marine Sciences, 73, 379-396. [ Links ]
Poole, H. H., & Atkins, W. R. C. (1929). Photoelectric measurements of submarine illumination throughout the year. Journal of the Marine Biological Association of the U.K., 16, 297 - 324. [ Links ]
Rakhesh, M., Raman, A. V., & Sudarsan, D. (2006). Discriminating zooplankton assemblages in neritic and oceanic waters: A case for the northeast coast of India, Bay of Bengal. Marine Environmental Research, 61, 93-109. [ Links ]
Steele, R. D. (1998). NRCA - Demo Palisadoes and Port Royal Protected area project. Report on the Palisadoes Tombolo and the Port Royal Cays. The Department of Life Sciences, University of the West Indies Mona. [ Links ]
Todd, C. D., Laverack, M. S., & Boxshall, G. A. (1996). Coastal marine zooplankton. A practical manual for students. Oxford: Cambridge Univ. Press. [ Links ]
Tseng, L., Dahms, H., Chen, Q., & Hwang, J. (2013). Geospatial variability in the autumn community structure of epipelagic zooplankton in the upper layer of the northern South China Sea. Zoological Studies, 52, 1-12. [ Links ]
UNESCO. (1968). Monographs on oceanic methodology: zooplankton sampling. Paris: UNESCO. [ Links ]
Van Guelph, L., Markle, D. F., & Duggan, D. J. (1983). An evaluation of the accuracy, precision, and speed of several zooplankton sub sampling techniques. J. Cons. Perm. Int. Explor. Mer., 40, 226-236. [ Links ]
Vant, W. N., & Safi, K. A. (1996). Size - fractionated phytoplankton biomass and photosynthesis in Manukau Harbour, New Zealand. New Zealand Journal of Marine and Freshwater Research, 30, 115 -125. [ Links ]
Vollenweider, R. Giovanardi, A. F. Montanari, G., & Rinaldi, A. (1998). Characterization of the trophic conditions of the marine coastal waters with special reference to the NW Adriatia sea: proposal for a trophic scale, turbidity and generalized water quality index. Environmetrics, 9, 329-357. [ Links ]
Wade, B., Antonio, L., & Mahon, R. (1972). Increasing organic pollution in Kingston Harbour, Jamaica. Marine Pollution Bulletin, 3, 106 - 110. [ Links ]
Webber, D. F. (1990). Phytoplankton populations of the coastal zone and nearshore waters of Hellshire: St. Catherine, Jamaica. Ph.D. Thesis, University of the West Indies, Mona. [ Links ]
Webber, M. K., & Roff, J. C. (1995). Annual structure of the copepod community and associated pelagic environment off Discovery Bay, Jamaica. Marine Biology, 123, 467-479. [ Links ]
Webber, M. K., Roff, J. C., Chisholm, L. A., & Clarke, C. (1996). Zooplankton distributions and community structure in an area of the south coast shelf of Jamaica. Bulletin of Marine Sciences, 59, 259-270. [ Links ]
Webber, D. F., & Roff, J. C. (1996). Influence of Kingston Harbour on the phytoplankton community of the nearshore Hellshire coast, southeast Jamaica. Bulletin of Marine Sciences, 59, 245 - 258. [ Links ]
Webber, D. F., & Webber, M. K. (1998). The water quality of Kingston Harbour: Evaluating the use of the planktonic community and traditional water quality indices. Chemistry and Ecology, 14, 357-374. [ Links ]
Webber, M. K., Edward-Myers, E., Campbell, C., & Webber, D. (2005). Phytoplankton and zooplankton as indicators of water quality in Discovery Bay, Jamaica. Hydrobiologia, 545, 177-193. [ Links ]
Youngbluth, M. J. (1980). Daily, seasonal, and annual fluctuations among zooplankton populations in an unpolluted tropical embayment. Estuarine, Coastal and Marine Science, 10, 265-287. [ Links ]
Arar, J. E., & Collins, G. B. (1997). In vitro determination of chlorophyll a and pheophytin a in marine and freshwater algae by fluorescence. National Exposure Research Laboratories Office of Research Development. USEPA Method 445.0. [ Links ]
Caldeira, R. M. A., Gordon, S., Miller, P., Pilgrim, D., & Nezlin, N. P. (2002). Sea-surface signatures of the island mass effect phenomenon around Madeira island, northeastern Atlantic. Remote Sensing of Environments, 80, 336 - 360. [ Links ]
Campbell, P., Manning J., Webber, M., & Webber, D. (2008). Planktonic communities as indicators of water quality in mangrove lagoons: a Jamaican case study. Transitional Waters Bulletin, 3, 39 - 63. [ Links ]
Carleton, J. H., Brinkman, R., & Doherty, P. J. (2001). Zooplankton community structure and water flow in the lee of Helix reef (Great Barrier Reef, Australia). Marine Biology, 139, 705-763. [ Links ]
Cowen, R. K., & Castro, L. R. (1994). Relation of coral reef fish larval distributions to island scale circulation around Barbados, West Indies. Bulletin of Marine Science, 54, 228-244. [ Links ]
Dunbar, F. N., & Webber, M. K. (2003). zooplankton distribution in the eutrophic Kingston Harbour, Jamaica. Bulletin of Marine Science, 73, 343-359. [ Links ]
Fehling, J., Davisdon, K., Bolch, C., Brand, T., & Narayanaswamy, B. (2012). The relationship between phytoplankton distribution and water column characteristics in north western European shelf sea waters. PLoS ONE, 7(3), e34098. doi:10.1371/journal.pone.0034098. [ Links ]
Genin, A. (2004). Bio-physical coupling in the formation of zooplankton and fish aggregations over abrupt topographies. Journal of Marine Systems, 50, 3-20. [ Links ]
Goodbody, I. (2003). Kingston harbor Jamaica: an overview. Bulletin of Marine Science, 73, 249 - 255. [ Links ]
Grant, C. J., & Wyatt, J. R. (1980). Surface currents in the eastern Cayman and western Caribbean seas. Bulletin of Marine Sciences, 30, 613 - 622. [ Links ]
Hernández-León, S. (1990). Accumulation of mesozooplankton in a wake area as a causative mechanism of the “island-mass effect”. Marine Biology, 109, 141-147. [ Links ]
Heywood, K. J., Barton, E. D., & Simpson, J. H. (1990). The effects of flow disturbance by an oceanic island. Journal of Marine Research, 48, 55 -73. [ Links ]
Hopcroft, R. R. (1988). Size fractionated biomass and primary production of tropical neretic phytoplankton near Kingston Jamaica. M.Sc. Thesis, University of Guelph, Jamaica. [ Links ]
Lindo, M. (1991). The effect of Kingston Harbour outflow on the zooplankton populations of Hellshire, south-east coast, Jamaica. Estuarine, Coastal and Shelf Science, 32, 597 - 608. [ Links ]
Matsouka, K., Joyce, L. B., Katoni, Y., & Matsuyma, Y. (2003). Modern dinoflagellate cysts in hypertrophic coastal waters off Tokyo Bay, Japan. Journal of Plankton Research, 25, 1461-1470. [ Links ]
McGehee, D. E., Derner, D. A., & Warren, J. D. (2004). Zooplankton in the Ligurian Sea: Part 1. Characterization of their dispersion, relative abundance and environment during summer 1999. Journal of Plankton Research, 26, 1409 - 1418. [ Links ]
Michel, H. B. (1984). Chaetognaths of the Caribbean sea and adjacent waters. NOAA Technical Report of the North-East Marine Fisheries Service, 15, 25. [ Links ]
Moore, E., & Sander, F. (1979). A comparative study of zooplankton from oceanic, shelf, and harbour waters of Jamaica. Biotropica, 11, 196-206. [ Links ]
Moraitou-Apostolopoulou, M., & Kiortsis, V. (1973). The Cladocerans of the Aegan Sea: Occurrence and seasonal variation. Marine Biology, 20, 137-143. [ Links ]
Munro, J. C. (1983). Caribbean coral reef fishery report. ODA/ UWI Fisheries Ecology Research Project 1969-1973. [ Links ]
Newell, G. E., & Newell, R. C. (1977). Marine plankton: a practical guide. London: Huthinson and Co. Ltd. [ Links ]
Owre, H. B., & Foyo, M. (1967). Copepods of the Florida current. Fauna Caribaea No. 1.Crustacea, Part1: Copepoda. Miami: Institute of marine science University of Miami. [ Links ]
Paerl, H. W. (1978). Effectiveness of various counting methods in detecting viable plankton. Journal of Marine and Freshwater Research, 12, 67-72. [ Links ]
Parsons, T. R., Maita, Y., & Lalli, C. M. (1984). A manual of chemical and biological methods for seawater analysis. UK: Pergamon Press. [ Links ]
Persad, G., Hopcroft, R. R., Webber, M. K., & Roff, J. C. (2003). Abundance, biomass and production of ctenophores and medusae off Kingston, Jamaica. Bulletin of Marine Sciences, 73, 379-396. [ Links ]
Poole, H. H., & Atkins, W. R. C. (1929). Photoelectric measurements of submarine illumination throughout the year. Journal of the Marine Biological Association of the U.K., 16, 297 - 324. [ Links ]
Rakhesh, M., Raman, A. V., & Sudarsan, D. (2006). Discriminating zooplankton assemblages in neritic and oceanic waters: A case for the northeast coast of India, Bay of Bengal. Marine Environmental Research, 61, 93-109. [ Links ]
Steele, R. D. (1998). NRCA - Demo Palisadoes and Port Royal Protected area project. Report on the Palisadoes Tombolo and the Port Royal Cays. The Department of Life Sciences, University of the West Indies Mona. [ Links ]
Todd, C. D., Laverack, M. S., & Boxshall, G. A. (1996). Coastal marine zooplankton. A practical manual for students. Oxford: Cambridge Univ. Press. [ Links ]
Tseng, L., Dahms, H., Chen, Q., & Hwang, J. (2013). Geospatial variability in the autumn community structure of epipelagic zooplankton in the upper layer of the northern South China Sea. Zoological Studies, 52, 1-12. [ Links ]
UNESCO. (1968). Monographs on oceanic methodology: zooplankton sampling. Paris: UNESCO. [ Links ]
Van Guelph, L., Markle, D. F., & Duggan, D. J. (1983). An evaluation of the accuracy, precision, and speed of several zooplankton sub sampling techniques. J. Cons. Perm. Int. Explor. Mer., 40, 226-236. [ Links ]
Vant, W. N., & Safi, K. A. (1996). Size - fractionated phytoplankton biomass and photosynthesis in Manukau Harbour, New Zealand. New Zealand Journal of Marine and Freshwater Research, 30, 115 -125. [ Links ]
Vollenweider, R. Giovanardi, A. F. Montanari, G., & Rinaldi, A. (1998). Characterization of the trophic conditions of the marine coastal waters with special reference to the NW Adriatia sea: proposal for a trophic scale, turbidity and generalized water quality index. Environmetrics, 9, 329-357. [ Links ]
Wade, B., Antonio, L., & Mahon, R. (1972). Increasing organic pollution in Kingston Harbour, Jamaica. Marine Pollution Bulletin, 3, 106 - 110. [ Links ]
Webber, D. F. (1990). Phytoplankton populations of the coastal zone and nearshore waters of Hellshire: St. Catherine, Jamaica. Ph.D. Thesis, University of the West Indies, Mona. [ Links ]
Webber, M. K., & Roff, J. C. (1995). Annual structure of the copepod community and associated pelagic environment off Discovery Bay, Jamaica. Marine Biology, 123, 467-479. [ Links ]
Webber, M. K., Roff, J. C., Chisholm, L. A., & Clarke, C. (1996). Zooplankton distributions and community structure in an area of the south coast shelf of Jamaica. Bulletin of Marine Sciences, 59, 259-270. [ Links ]
Webber, D. F., & Roff, J. C. (1996). Influence of Kingston Harbour on the phytoplankton community of the nearshore Hellshire coast, southeast Jamaica. Bulletin of Marine Sciences, 59, 245 - 258. [ Links ]
Webber, D. F., & Webber, M. K. (1998). The water quality of Kingston Harbour: Evaluating the use of the planktonic community and traditional water quality indices. Chemistry and Ecology, 14, 357-374. [ Links ]
Webber, M. K., Edward-Myers, E., Campbell, C., & Webber, D. (2005). Phytoplankton and zooplankton as indicators of water quality in Discovery Bay, Jamaica. Hydrobiologia, 545, 177-193. [ Links ]
Youngbluth, M. J. (1980). Daily, seasonal, and annual fluctuations among zooplankton populations in an unpolluted tropical embayment. Estuarine, Coastal and Marine Science, 10, 265-287. [ Links ]
1. Department of Life Sciences, University of the West Indies, Mona, Kingston 7, Jamaica; hugh.small@uwimona.edu.jm, mona.webber@uwimona.edu.jm. dale.webber@uwimona.edu.jm, krislue@gmail.com.
Received 19-IX-2013 Corrected 31-III-2014 Accepted 01-IV-2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}