










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.2 San José Apr. 2014
Downstream effects of hydropower production on aquatic macroinvertebrate assemblages in two rivers in Costa Rica
Abstract
Despite the fact that little is known about the consequences of hydropower production in tropical areas, many large dams (>15m high) are currently under construction or consideration in the tropics. We researched the effects of large hydroelectric dams on aquatic macroinvertebrate assemblages in two Costa Rican rivers. We measured physicochemical characteristics and sampled aquatic macroinvertebrates from March 2003 to March 2004 in two dammed rivers, Peñas Blancas and San Lorenzo, as well as in the undammed Chachagua River. Sites above and below the dam had differences in their physicochemical variables, with wide variation and extreme values in variables measured below the dam in the San Lorenzo River. Sites below the dams had reduced water discharges, velocities, and depths when compared with sites above the dams, as well as higher temperatures and conductivity. Sites above dams were dominated by collector-gatherer-scrapers and habitat groups dominated by swimmer-clingers, while sites below dams had a more even representation of groups. In contrast, a comparison between two sites at different elevation in the undammed river maintained a similar assemblage composition. Tributaries might facilitate macroinvertebrate recovery above the turbine house, but the assemblage below the turbine house resembled the one below the dam. A massive sediment release event from the dam decreased the abundance per sample and macroinvertebrate taxa below the dam in the Peñas Blancas River. Our study illustrates the effects of hydropower production on neotropical rivers, highlighting the importance of using multiple measures of macroinvertebrate assemblage structure for assessing this type of environmental impact. Rev. Biol. Trop. 62 (Suppl. 2): 179-201. Epub 2014 April 01.
Key words: Functional feeding groups, habitat groups, hydropower, tropical river ecology, turbine house.
Resumen
Existe poca información publicada sobre el efecto de grandes represas hidroeléctricas (>15m alto) en ríos tropicales. Investigamos los efectos de estas represas en las comunidades de macroinvertebrados acuáticos en dos ríos de Costa Rica. Medimos características físico-químicas y recolectamos macroinvertebrados a lo largo del año en dos ríos represados, Peñas Blancas y San Lorenzo, así como en el río Chachagua, el cual no era utilizado para la producción hidroeléctrica. Los sitios arriba y debajo de las represas presentaron diferencias físico-químicas, con amplia variación y valores extremos en las variables medidas debajo de la represa en el Río San Lorenzo. Los sitios arriba de las represas estaban dominados por colectores-recolectores-raspadores y nadadores-colgadores, mientras que debajo de las represas hubo una representación más equitativa de grupos funcionales. En contraste, las comunidades fueron semejantes en dos sitios a diferente elevación en el río sin represa. La presencia de afluentes podría haber facilitado la recuperación de macroinvertebrados arriba de la casa de máquinas, pero las comunidades fueron similares bajo la casa y abajo de la represa. Una liberación masiva de sedimentos disminuyó la abundancia y el número de táxones bajo la represa en el Río Peñas Blancas.
Palabras clave: grupos tróficos funcionales, grupos de hábitat, energía hidroeléctrica, ecología de ríos tropicales, casa de máquinas.
Due to rising energy demands, hydropower production is increasingly advocated and implemented in developing, tropical countries (Revenga, Brunner, Henninger, Kassem & Payne, 2000; Anderson, Pringle & Rojas, 2006; Finer & Jenkins, 2012). While the world-wide construction rate of large hydroelectric dams (>15m, World Commission on Dams (WCD), 2000) peaked in the 1960s, the construction rate in tropical areas is still increasing (Petts, 1990; Pringle, Freeman & Freeman, 2000; Regalado, 2011). This disparity is partially due to the exhaustion of suitable undammed rivers for hydropower production in temperate, but not tropical, areas (Revenga et al., 2000; Finer & Jenkins, 2012). This rising development in hydropower generation comes with environmental and socio-economical costs for the countries involved (Pringle, 2000; WCD, 2000; Tollefson, 2011).
Large hydroelectric dams affect biological communities through changes in stream physicochemical characteristics. Large dams reduce river connectivity (Pringle, 2001; Finer & Jenkins, 2012), cause habitat loss (Dudgeon et al., 2006; Freeman, Pringle & Jackson, 2007; Rodrigues & Silva, 2012), alter water temperature and oxygen (Ward & Stanford, 1983; Viana, 2002; Foto Menbohan, Koji, Ajeagah, Bilong Bilong & Njiné, 2012), and interrupt water flow, movement of sediments, nutrients and fauna (Ward & Stanford, 1983; Pringle, 1997; Bunn & Arthington, 2002; Greathouse, Pringle & Holmquist, 2006a; Williams, Armstrong, Katopodis, Lariniere & Travade, 2012). Streams affected by large dams can have decreased macrophyte total biomass, as well as macroinvertebrate abundance, richness, and biomass (Troelstrup & Hergenrader, 1990; Dejalon & Sanchez, 1994; Vinson, 2001). Additionally, due to changes in food sources and substrate, the composition of macroinvertebrate functional groups changes downstream from hydroelectric dams, favoring functional groups that can exploit plankton drifting from the reservoir, and that prefer habitats with low sediment deposition (Dejalon & Sanchez, 1994; Doi et al., 2008; Katano, Negishi, Minagawa, Doi, Kawaguchi & Kayaba, 2009). Furthermore, species assemblages of invertebrates and fish are also affected below dams, favoring organisms that are tolerant to the physicochemical characteristics imposed by hydroelectric production (Dejalon & Sanchez, 1994; Vinson, 2001; Foto Menbohan et al., 2012).
Until recently, most of our knowledge on the effects of hydroelectric dams on stream biota came from studies in temperate areas (Pringle et al., 2000). However in the last decade there has been an increased effort to understand the effects of hydroelectric dams on tropical stream biota. Most previous studies on large hydroelectric dams on tropical aquatic organisms have focused on the consequences of flow regulation on fish assemblages (e.g. Albrecht, Caramaschi & Horn, 2009; Terra, dos Santos & Araujo, 2010). Connectivity disruption along streams and changes in physicochemical characteristics associated with hydropower production alter the composition and structure of tropical fish assemblages, and lowers fish diversity downstream from dams (Pringle et al., 2000; Agostinho, Pelicice & Gomes, 2008; Araujo, Pinto & Teixeira, 2009; Cooney & Kwak, 2013). Despite the importance of tropical aquatic macroinvertebrates as a food source for higher trophic levels (Chan, Zhang & Dudgeon, 2007; Ortaz, Martín & Lopez-Ordaz, 2011) and as a functional group in streams (Benstead, 1996; Wright & Covich, 2005), little is known about the consequences of dams on their assemblages.
The little work that has been published on the consequences of hydroelectric dams on tropical aquatic macroinvertebrates comes from particular geographic areas, limiting our general understanding of the consequences of dams on macroinvertebrates. Continental tropical rivers show a decrease in taxa richness and changes in species assemblages in rivers associated with large reservoirs (Jorcin & Nogueira, 2008; Jorcin, Nogueira & Belmont, 2009; Foto Menbohan et al., 2012). Additionally, island streams, which are shorter and have a prevalence of diadromy (March, Benstead, Pringle & Scatena, 2003; McLarney, Mafla, Arias & Bouchonnet, 2010), show negative effects of large dams on migratory tropical fauna, such as shrimps and snails, with consequences for the entire assemblage of aquatic macroinvertebrates (Greathouse et al., 2006a; Greathouse, Pringle, McDowell & Holmquist, 2006b). Little is known, however, about the effects of hydroelectric dams on rivers in isthmuses, including Central America.
Despite the proposed expansion of hydroelectricity in Central America (Esselman & Opperman, 2010), to our knowledge there are no published studies on the effects of dams on benthic fauna in Central American rivers. Studying the consequences of hydroelectricity on aquatic macroinvertebrates in Central America can advance our understanding of the ecological effects of hydroelectricity in tropical streams as a whole. In addition, it can inform comprehensive management techniques, such as a minimal instream flow to protect aquatic biota (known as the minimum environmental flow), that can be used to mitigate the impacts of this type of energy production in Central American rivers (Scatena, 2004; Esselman & Opperman, 2010). We studied the effects of large hydroelectric dams on aquatic macroinvertebrates in two rivers in the San Carlos River basin in Costa Rica. Our goal was to evaluate the impact of dams on physicochemical characteristics downstream the dam and the turbine house, as well as the impacts on aquatic macroinvertebrate abundance, richness and assemblages. Based on previous studies, we predicted that large hydroelectric dams in our study area will change the physicochemical characteristics of streams and decrease aquatic macroinvertebrate abundance and richness below the dam and the turbine house. Additionally, we predicted changes in the macroinvertebrate assemblages downstream from the dam. We specifically expected an increase in the relative abundance of functional groups that are favored by conditions of low sedimentation and high plankton availability, which are common downstream from dams.
Materials and methods
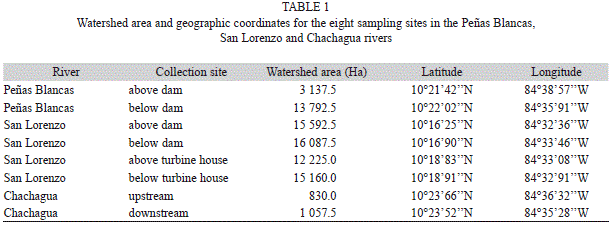
Study site: This work was conducted in three rivers on the northern Caribbean slope of Costa Rica, the dammed rivers Peñas Blancas and San Lorenzo, as well as the undammed Chachagua River (Fig. 1). The San Lorenzo, Peñas Blancas and Chachagua rivers are located in comparable life zones and have similar land use in their watersheds (Barrantes, Allan & Gallardo, 1990; Gutiérrez, 1999). The San Lorenzo River has an average annual flow of 10.40m³s-1 on dry years, and of 15.50m³s-1 on rainy years (Barrantes et al., 1990). This hydroelectric project collects water from the San Lorenzo and the Jamaical rivers in a 63 000m3 reservoir, behind a 21m high dam, and maintains a minimum flow of 0.5m³s-1 below the dam (Barrantes et al., 1990; Conelectricas, 2013; L. Torre Conelectricas, pers. comm.). The Peñas Blancas River has an average annual flow of 35.60m³s-1 at the Peñas Blancas gauging station, and of 18.20m³s-1 in the Pocosol gauging station (Gutiérrez, 1999). This hydroelectric project collects water from the Peñas Blancas River in a 2x106m3 reservoir, behind a 47m high dam, and maintains a minimum flow of 1.0m³s-1 below the dam (Gutiérrez, 1999). At both hydroelectric projects water is diverted and returned to the river after passing through the turbine house about 3km downstream from the dam (Barrantes et al., 1990; Gutiérrez, 1999). Hydropower generation in San Lorenzo and Peñas Blancas occurs year-round, and at times of highest electrical demand, normally from 1 000 to 1 400hr. This type of electricity generation causes daily fluctuation in river flow below the turbine house. The San Lorenzo hydroelectric project receives input from two streams downstream from the dam; these streams are not used for energy production (Fig. 1).
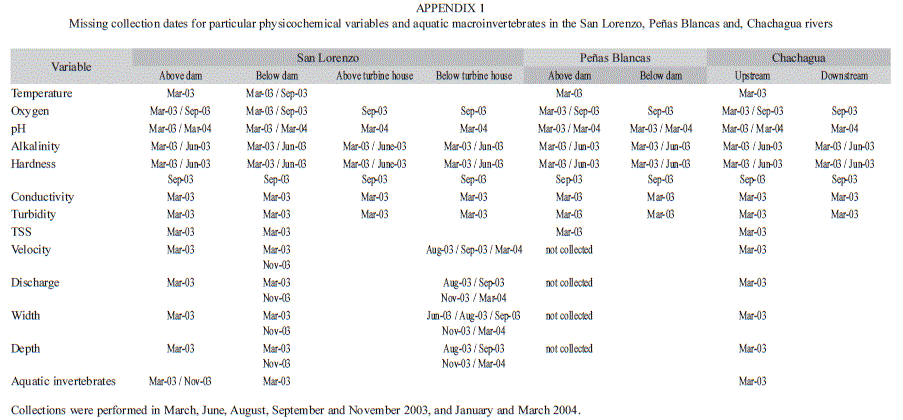
To test for effects of hydroelectric operations on the physicochemical characteristics and aquatic macroinvertebrate assemblages, we used a paired design where each comparable site in each river was sampled seven times throughout one year. We sampled above and below the dam in the Peñas Blancas and the San Lorenzo River, and above and below the turbine house in the San Lorenzo River (Table 1, Fig. 1). Sites above the dam were free from dam effects (Fig. 1). Additionally, we chose two sampling sites in the Chachagua River for reference purposes, which were comparable in elevation to sampling sites in dammed rivers (Table 1, Fig. 1). We collected samples on five dates in 2003 –March, June, August, September and November–, and two dates in 2004 –January and March–. However, due to logistic constraints we were unable to collect data for all the study variables on every collection date (Appendix 1).
Physicochemical sampling: At each site we measured temperature and dissolved oxygen (DO) using a YSI® model 52, pH, alkalinity and hardness using Hach® test kits, and conductivity with a La Motte® conductimeter. We also collected water samples to measure total suspended solids (TSS) by weight difference of the retained material in pre-weighed glass fiber filters (GF/C) and turbidity through the determination of the absorbance at 450nm in a Shimadzu® UV-160A spectrophotometer (Lind, 1985). Additionally, we measured mean water velocity with an electronic flow meter (Global Water® FP111), and depth and channel width using a tape measure. Mean water velocity and depth were measured at 1m intervals across the channel. Mean water velocity readings were taken at 60% of water depth at each measuring point. Width was measured by holding the tape measure across the channel from bank to bank on the water surface. We measured discharge using the midsection method (Hauer and Lamberti, 2007). Mean water velocity, depth, width, and water discharge were not measured upstream the dam in the Peñas Blancas River, however we obtained these measures from the hydrology department at Instituto Costarricense de Electricidad (ICE).
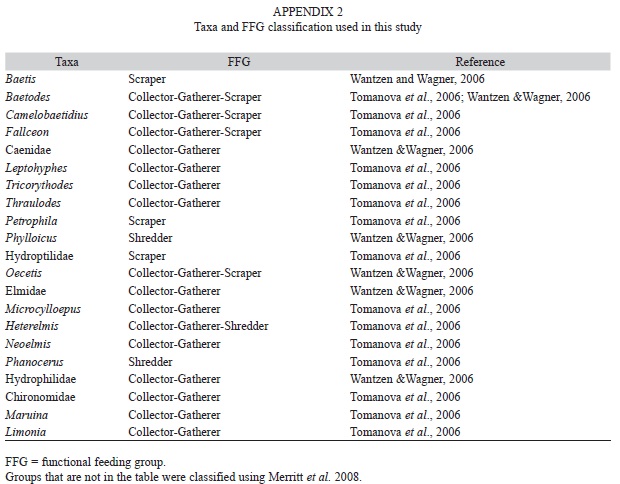
Aquatic macroinvertebrate sampling: Our sampling method for aquatic macroinvertebrates represents a capture per unit effort (CPUE). To estimate abundance per sample we performed multi-habitat collections using a 500µm mesh sized hand-net for one hour along a ~25m reach of the river at each site. We pooled samples by collection date, preserved in 70% EtOH and counted and identified all the individuals to the maximum taxonomical resolution, mostly genera, using various taxonomic keys (Merritt & Cummins, 1996; Roldán, 1996; Springer, Ramírez & Hanson, 2010). Specimens were deposited at the Museum of Zoology, University of Costa Rica. We used tropical functional feeding group (FFG) classification when available (Cummins, Merritt & Andrade, 2005; Tomanova, Goitia & Helesic, 2006; Wantzen & Wagner, 2006; Appendix 2). Groups not included in tropical studies were classified according to Merritt, Cummins & Berg (2008). Additionally, we classified the insects in habitat groups according to Merritt et al. (2008).
We collected additional aquatic macroinvertebrate samples from the Peñas Blancas River, on November 7 and December 22, 2003, in order to assess the effect of a massive sediment release event from the Peñas Blancas reservoir on October 31, 2003 (La Nación, 2003). Flushing sediments from the reservoir is a common practice in hydroelectric facilities in Costa Rica, since it prolongs the useful life of the dam.
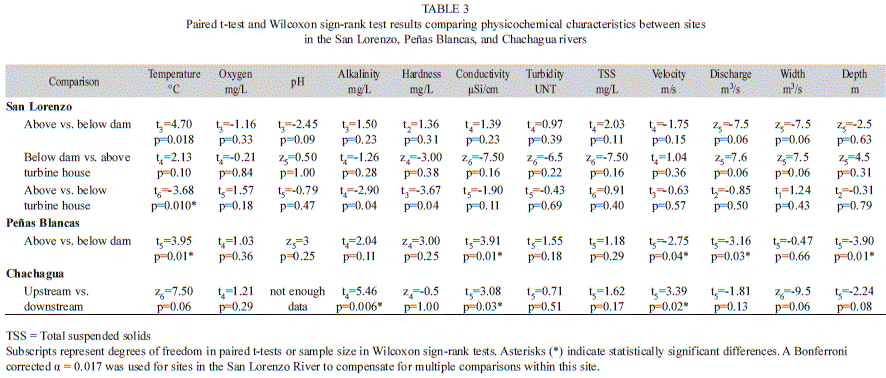
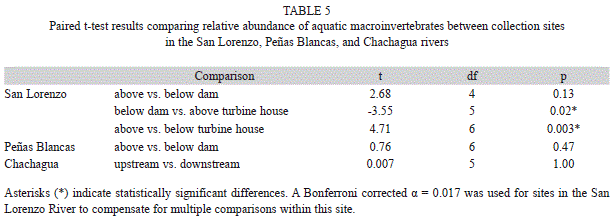
To test for differences in physicochemical characteristics, as well as in macroinvertebrate abundance per sample between sites within each of the streams, we used paired statistical analyses. To ensure that temporal autocorrelations did not bias P-values, we confirmed that there were no significant correlations among sequential sampling dates by using the Durbin-Watson statistic test. In the case of the San Lorenzo River, we performed the paired tests for adjacent sections of the river (above dam vs. below dam, below dam vs. above turbine house, and above turbine house vs. below turbine house) and used a Bonferroni-corrected alpha of 0.017. Correlations and paired tests were performed using JMP software (SAS institute Inc., 2009).
To account for effects of differences in macroinvertebrate abundance on estimates of taxa richness, we rarified richness estimates based on the abundance of the least-abundant sample in each site per month to obtain an unbiased expected taxa richness. Rarified taxa richness was estimated using EcoSim software version 7.0 (Gotelli & Colwell, 2001) and all randomizations were repeated 1 000 times. We also rarified estimates of total dominance (Simpson dominance index (S)). To statistically compare taxa richness and dominance between sites, we used the low and high bound for a 95% confidence interval calculated by EcoSim.
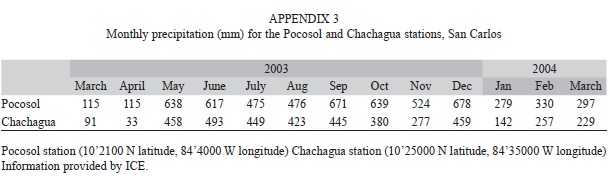
To test for effects of seasonal rainfall on the response variables collected during the study period we performed simple linear regressions between precipitation and the physicochemical variables, macroinvertebrate abundance per sample, and rarified richness measured at the different sites. The measures of precipitation for the study period were obtained from the Pocosol and the Chachagua stations (Appendix 3). Variables measured in the Peñas Blancas and the San Lorenzo rivers were regressed against precipitation data from the Pocosol station, while variables measured in the Chachagua River were regressed against precipitation data from the Chachagua station. We selected the precipitation data set used in each analysis based on distance of each station to the study sites, as well as similarities in elevation between the sites and each station. The data used in the analyses fulfilled the assumptions of regression. Regressions were performed using JMP software (SAS institute Inc., 2009).
To assess differences in macroinvertebrate assemblages among study sites, we performed a non-metric multidimensional scaling (nMDS) analysis on a Bray-Curtis dissimilarity matrix. nMDS is a distance-based approach that relies on a similarity matrix, maximizing rank-order correlation between distance measures and distance in ordination space (Clarke & Warwick, 2001). The stress value obtained from the NMDS analysis is a measure of distortion between the positions of real data points and their graphical representation (Clarke & Warwick, 2001). Thus, a low stress value is associated with a graph that more accurately represents the dissimilarities. The nMDS was performed using the vegan package (Oksanen et al., 2010) in R statistical software (R development core team, 2009).
We used chi-square tests to test for differences in percentages of the different FFGs and habitat groups among sites within streams, using JMP software (SAS institute Inc., 2009). Only months when all compared sites were sampled were included in the nMDS, FFGs and habitat group analyses. Finally, to test the effects of the massive sediment liberation in Peñas Blancas we performed a t-test using JMP software (SAS institute Inc., 2009). We tested for a difference in the number of taxa below the dam in the Peñas Blancas River before and after the massive sediment liberation. The data used in this analysis fulfilled the assumptions of t-test.
Results
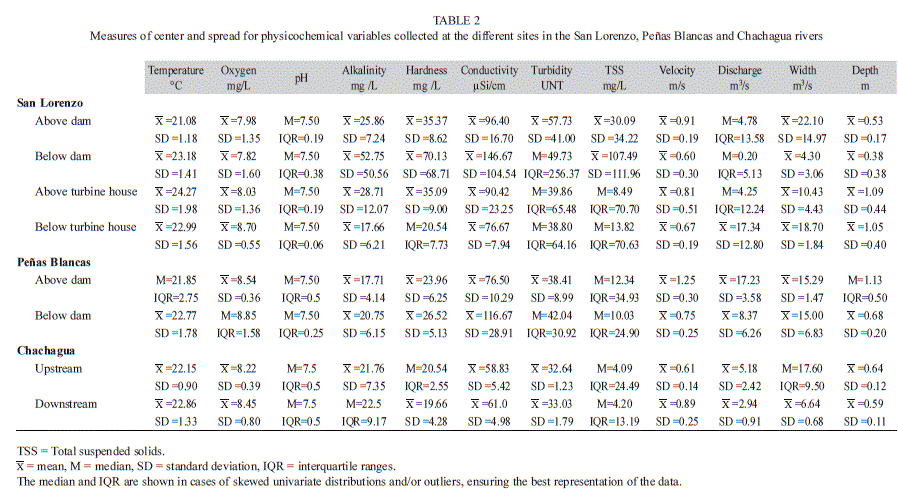
Physicochemical characteristics: The site below the dam in the San Lorenzo River had extreme values for several physicochemical variables. Alkalinity, hardness, and conductivity were higher immediately below the dam than in the other sites in this river, with alkalinity and hardness being >80% higher than in the other sites (Table 2). Additionally, water discharge was ~95% lower below the dam than in the other studied sites, and velocity, width, and depth were lowest below the dam (Table 2). None of these differences were statistically significant (Table 3). These physicochemical variables varied widely throughout the study period; however none of this variation had a relationship with precipitation (Appendix 4). Relative to above the turbine house, the downstream site had four times more water discharge and twice the width (Table 2), however these differences were not statistically significant (Table 3). Temperature was the only physicochemical variable that was statistically different between sites in the San Lorenzo River, with temperatures in average 1.28°C higher above the turbine house than below (Table 2 and 3). Although not statistically significant, the site below the dam had temperatures on average 2.10°C higher than above the dam in the San Lorenzo River (Table 2 and 3).
Similar patterns as those found above vs. below the dam in the San Lorenzo River were observed for physicochemical comparisons in the Peñas Blancas River above vs. below the dam. Conductivity was 53% higher below the dam than above the dam, and temperature was 0.92°C higher below dam (Table 2). Additionally, water discharge, velocity, and depth were more than 40% lower below the dam (Table 2 and 3).
There was variation in physicochemical variables between upstream and downstream sites in the Chachagua River. Alkalinity and conductivity were 3% and 4% higher respectively downstream than upstream (Table 2). Water discharge and stream width were 76% and 165% higher upstream than downstream respectively, however only the difference in width was statistically significant (Table 3). Additionally, water velocity was 46% higher downstream than upstream (Table 2 and 3).
Most of the physicochemical variables in this study did not show a relationship with precipitation (Appendix 4). Depth decreased as precipitation increased in the site below the dam in the Peñas Blancas River and above the turbine house in the San Lorenzo River. Discharge decreased as precipitation increased in the Peñas Blancas River below the dam, while velocity increased in the site above the dam. Above the turbine house in the San Lorenzo River pH decreased with precipitation.
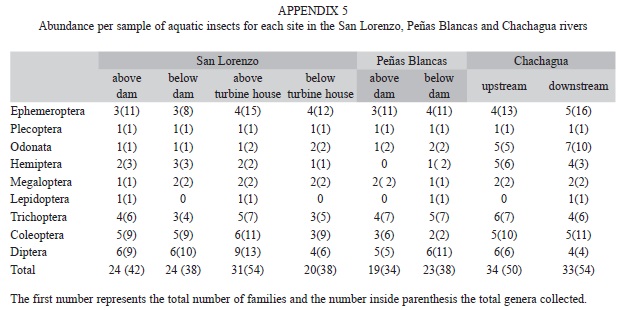
Aquatic macroinvertebrate assemblage: A total of 5 803 individuals from 98 genera or morphogenera in 46 families from nine orders of aquatic insects were collected. At each study site the number of families ranged between 19 and 34, and between 34 and 54 genera, with the highest numbers found at the reference sites in Chachagua River and above the turbine house in San Lorenzo (Appendix 5). Few taxa contained most of the abundance (Table 4). Non-insect taxa were represented by Acari, Hirudinea and Gastropoda.
The overall average abundance per sample in the San Lorenzo River was 191±104 individuals above the dam, 182±69 individuals above the turbine house, 51±42 individuals below the dam, and 61±35 individuals below the turbine house. On average the site above the dam had four times higher abundances than the site below the turbine house, however differences in abundance were only statistically significant between sites below the dam and turbine house, and above the turbine house (Table 5). In the Peñas Blancas River, the site above the dam had an average abundance per sample of 92±35 individuals, while the site below had 69±66 individuals; these differences were not statistically significant (Table 5). Both sites in the undammed river Chachagua had similar abundances per sample (Chachagua upstream=132±46, Chachagua downstream=137±50; Table 5). The only collection site that had a relationship between abundance and precipitation was the site above the turbine house, with abundance decreasing as precipitation increased (Appendix 6).
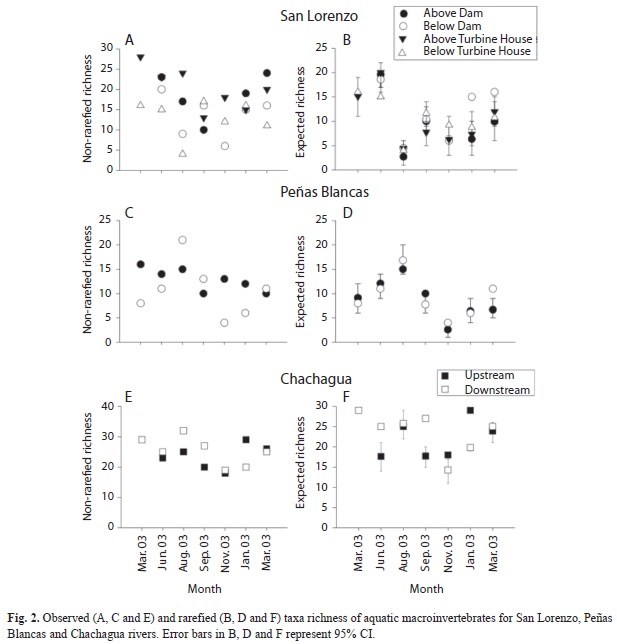
Among sites in the San Lorenzo River, taxa richness was lower below the dam for all months except September 2003, and the site below the turbine house had lower observed taxa richness in all collection months except September 2003 and January 2004 (Fig. 2A). Rarified taxa richness was significantly higher below the dam in January (72%) and March (34%) 2004 (Fig. 2B). Additionally, below the turbine house had significantly lower (19%) rarified richness in June 2003 (Fig. 3B). In the Peñas Blancas River observed taxa richness was lower in the site below the dam than above the dam in all collection months except August and September 2003, while in March 2004 the observed richness for both sites was very similar (Fig. 2C). Rarified richness was significantly higher (65%) for the site below the dam than above the dam in March 2004 (Fig. 2D). Observed richness in Chachagua upstream was higher than downstream only in January 2004, while November 2003 and March 2004 the observed richness was very similar (Fig. 2E). Rarified richness was significantly higher in Chachagua downstream than upstream in June (42%) and September (52%) 2003, while it was significantly higher in Chachagua upstream in November 2003 (26%) and January 2004 (46%) (Fig. 2F). None of the collection sites had a relationship between macroinvertebrate rarified richness and precipitation (Appendix 6).
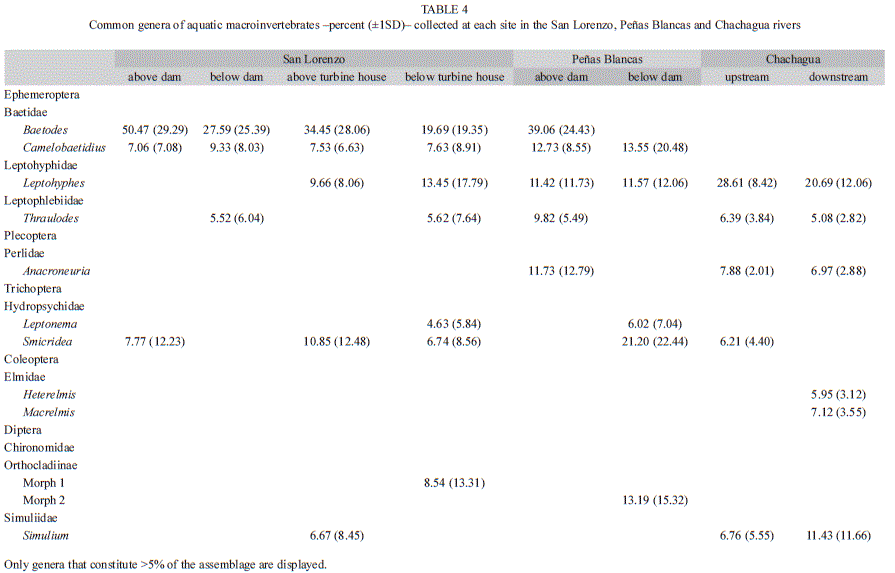
When comparing composition among sites, the non-metric multidimensional scaling ordination (stress=0.09) separated the sites above both the dam and the turbine house from sites below both the dam and the turbine house, while both sites in the Chachagua River were closer together (Fig. 3). Sites above the dam were dominated by Baetodes (Ephemeroptera: Baetidae) (SSan Lorenzo above dam=0.57, 95% CI=0.52-0.61; SPeñas Blancas above dam=0.45, 95% CI=0.42-0.47; Table 4), and above the turbine house this genus was over one-third of the taxa collected (S=0.33, 95% CI=0.29-0.38; Table 4). Sites below the dam and the turbine house in the San Lorenzo River did not have a dominant taxa (SSan Lorenzo below dam=0.23, 95% CI=0.23 to 0.23; SSan Lorenzo below turbine house=0.18, 95% CI=0.16-0.20; Table 4). Below the dam in the Peñas Blancas River had high abundance per sample of Smicridea (Trichoptera: Hydropsychidae) (S=0.35, 95% CI=0.35 to 0.35; Table 4). Although upstream Chachagua had higher dominance than downstream (SChachagua upstream=0.30, 95% CI=0.30 to 0.30; SChachagua downstream=0.20, 95% CI=0.19-0.22), both sites had high abundance per sample of Leptohyphes (Ephemeroptera: Leptohyphidae) (Table 4).
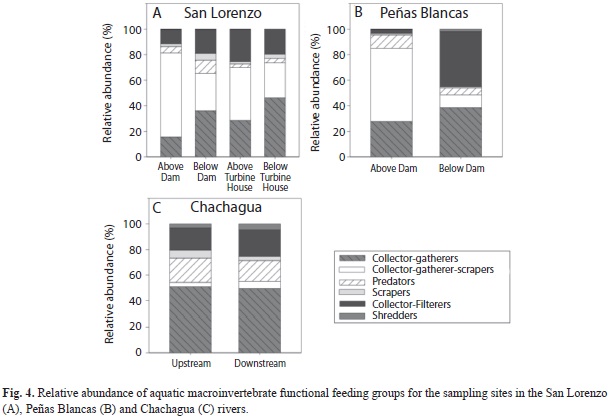
We found differences in FFG composition between sites above the dam and the turbine house in the San Lorenzo River, and the sites below these structures (χ²15=342.74, p<0.0001). There was a shift from assemblages with higher relative abundance of collector-gatherer-scrapers (66%) in the site above the dam, to assemblages with high abundance of collector-gatherers (36%), collector-gatherers-scrapers (30%), and collector-filterers (19%) below the dam (Fig. 4A). The site above the turbine house had higher abundance of collector-gatherers-scrapers (41%) and collector-filterers (25%), and a lower of collector-gatherers (29%) relative to the site below the dam (Fig. 4A). The site below turbine house had fewer collector-gatherers-scrapers (27%) and collector-filterers (20%) relatively to the site above the turbine house, but more collector-gatherers (46%) (Fig. 4A). Similarly to the San Lorenzo River, the site above the dam in Peñas Blancas River had higher relative abundance of collector-gatherers-scrapers (57%), while the site below the dam had high abundances of collector-gatherers (39%) and collector- filterers (43%) (χ²5=458.29, p<0.0001; Fig. 4B). In contrast, both sites in the undammed river had higher abundance of collector-gatherers (51% upstream and 49% downstream); Chachagua upstream had more predators and scrapers, and less collector-filterers, collector-gatherers-scrapers and shredders than downstream (χ²5=20.15, p=0.001; Fig. 4C).
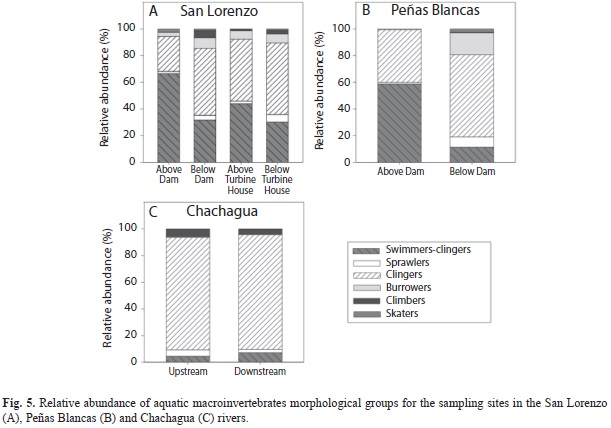
Habitat groups differed between sites above the dam and turbine house and sites below the dam and turbine house in the San Lorenzo River (χ²15=301.29, p<0.0001). The site above the dam had high relative abundance of swimmer-clingers (66%), while the site below the dam had a decrease in swimmer-clingers (32%) and an increase in clingers (50%) (Fig. 5A). Swimmer-clingers increased (44%) in the site above the turbine house in San Lorenzo relatively to the site below the dam, but decreased in the site below the turbine house (30%) (Fig. 5A). Similarly to the San Lorenzo River, above the dam in the Peñas Blancas River had high relative abundance of swimmer-clingers (59%), while the site below the dam had a decrease in swimmer-clingers (11%) and an increase in clingers (62% ) (χ²5=381.25, p<0.0001; Fig. 5B). Both sites in the reference stream were dominated by clingers (84% upstream and 86% downstream); upstream had more climbers and sprawlers, while downstream had more swimmer-clingers (χ²5=14.52, p=0.01; Fig. 5C).
The massive sediment liberation on October 31st, 2003, in the Peñas Blancas River decreased the number of taxa below the dam from an average of 13.25 (SE=2.78) in the months previous to the sediment liberation, to 7.2 (SE=1.24) after the liberation and until the end of this study (t4.19=1.99, p=0.06). The only specimens collected on November 7th were one individual from the Veliidae and four individuals in the Chironomidae.
Discussion
To the best of our knowledge this is the first published study documenting the effects of large dams on physicochemical variables and aquatic macroinvertebrates in Central America. In general, the results agree with our hypotheses on the effects of hydroelectric dams on physicochemical variables and macroinvertebrate assemblages of the studied streams. We found that sites above and below the dam had evident differences in their physicochemical variables, with wide variation and extreme values in variables measured below the dam in the San Lorenzo River. Additionally, we found changes in the assemblage structure of aquatic macroinvertebrates above and below the dams and the turbine house that were not observed in the undammed, reference river. Although sites above the dam had high dominance of a single species, sites below the dam showed more even compositions. Composition above the turbine house resembles that of above the dam, likely due to the confluence of tributaries above the sites, but the assemblage composition below the turbine house resembles that from below the dam. Although composition was affected by damming, taxa richness and abundance were not consistently affected.
The results of our study reflect the effects of hydroelectric production on stream hydrology and further consequences on the water quality of rivers (Poff et al., 1997; Pringle et al., 2000; Bunn & Arthington, 2002). Sites below the dams had a clear trend for reduced water discharge, velocities, and depths when compared to sites above the dams. Although both of the hydroelectric plants maintained a minimum environmental flow below the dam, this flow is a reduced version of what the natural flow was before streams were dammed, and it does not necessarily reflect the habitat preferences of stream biota (Zeledón, 2001). These observed reductions in water flow below the dams could explain differences in water quality parameters found between above and below the dams. Increases in water temperature below dams are a known consequence of water abstraction, reducing the water’s thermal capacity (Sinokrot & Gulliver, 2000). Moreover, higher conductivities below dams could be a consequence of higher temperatures in these sites (Kalff, 2002). Interestingly, there was a lack of statistical support for the observed differences in physicochemical variables between the site above and below the dam in the San Lorenzo River. This lack of statistical support is related with the wide variation in these parameters below the dam, which could be due to the nature of hydroelectric generation in this plant, where water is stored and released based on the electricity demands. Additionally, in August 2003 a flashflood blocked the minimum ecological flow, and this flow had not been re-established by the end of this study. Our results might be reflecting the importance of maintaining this minimum ecological flow.
Differences in the assemblage structure of aquatic macroinvertebrates between sites above and below the dam could be a consequence of the observed changes in hydrology and water quality. Previous studies have found that changes in hydrology and disruptions of the stream’s natural connectivity affect the transportation of nutrients, energy, and sediments to sites below the dam, with effects on macroinvertebrate assemblages (Ward & Stanford, 1983; Petts, 1984; Petts, Armitage & Castella, 1993; Ligon, Dietrich & Trush, 1995; Finer & Jenkins, 2012). Increases in the abundance of collector-filterers below dams are often a consequence of increased transport of phytoplankton and zooplankton from the reservoir through the hydroelectric facility to the river downstream, creating a lake-derived subsidy for food webs below the dam (Ward & Stanford, 1983; Doi et al., 2008; Foto Menbohan et al., 2012). This increase in plankton availability could explain the observed shift from communities dominated by Baetodes, a collector-gatherer-scraper, above dams in our study, to assemblages with higher relative abundance of collector-filterers in sites below the dams. Interestingly, the increase in collector-filterers was more obvious in the Peñas Blancas River, the river with the largest reservoir in our study. The lower relative abundance of swimmer-clingers downstream of the dam might be due to this group’s active drifting behavior to find better sites (Merritt et al., 2008). Studies have found an increase in macroinvertebrate drifting after changes in flow discharge (Minshall & Winger, 1968; Brooker & Hemsworth, 1978; Scullion & Scinton, 1983). Furthermore, the increase of clingers downstream of the dam could be explained by the release of clear water from the reservoir, which removes fine sediment particles creating sites of low sediment deposition below the dam (Ward & Stanford, 1983; Stevens, Shannon & Blinn, 1997; Rabeni, Doisy & Zweig, 2005; Katano et al., 2009).
The resemblance in composition between the site above the turbine house and the site above the dam in the San Lorenzo River could be explained by the confluence of tributaries upstream from the turbine house. The confluence of tributaries on impounded rivers attenuates the effects of hydropower production by restoring particle size, organic debris, and increasing diversity downstream from the confluence (Petts et al., 1993; Takao, Kawaguchi, Minagawa, Kayaba & Morimoto, 2008; Katano et al., 2009; Foto Menbohan et al., 2012). The role of tributaries facilitating the recovery of the aquatic macroinvertebrate assemblages could explain the increase in the relative abundance of collector-gatherer-scrapers and swimmer-clingers above the turbine house, relative to the site below the dam in the San Lorenzo River. Below the turbine house, however, the assemblage was similar to the one below the dam. According to WCD (2000) even though water is restored after electricity production below the turbine house, the amount and periodicity of discharge is very different from the natural flow regime, affecting benthic macroinvertebrates that are adapted to natural changes in water discharge (Cushman, 1985). This change in the natural flow regime below the turbine house was evidenced by the higher water discharges found below the turbine house. In comparison with the rest of the sites in the San Lorenzo River, these discharges were variable.
Results from the undammed Chachagua River exemplify how longitudinal changes occur in natural rivers, and are of lower magnitude than the changes measured when a dam is present. Although water discharge between Chachagua upstream and downstream did not vary significantly, width was higher upstream while water velocity was higher downstream. Additionally, the variation in alkalinity and conductivity between these two sites was not as evident as differences found between sites above and below dams, and could be a reflection of differences in geology between the sites. Furthermore, the changes in relative abundance of FFGs and morphological groups between upstream and downstream of the reference sites were less strong than the changes seen in the sites affected by hydroelectric production; both sites in the reference stream were dominated by collector-gatherers and clingers.
Despite existing monitoring efforts in dammed rivers in Central America, the studies typically remain as technical studies and there are few publications on the effects of dams on aquatic fauna. Our study highlights the importance of including community composition, especially comparisons of functional groups, in monitoring programs. We recommend future studies in tropical rivers to continue exploring the mechanisms responsible for changes in aquatic macroinvertebrates due to hydropower production. These studies should include continuous measures of physicochemical variables that allow capturing daily variation in discharge and peak flows, as well as rare, catastrophic events such as the massive sediment release that occurred in the Peñas Blancas River. These variations might be important in hydroelectric plants where water is stored and released based on electricity demands. Additionally, we recommend including measures on substrate size since this factor can be important in affecting assemblages of aquatic macroinvertebrates. Understanding the mechanisms affecting macroinvertebrates will facilitate development of mitigation techniques in existing and future hydroelectricity projects in the tropics. Furthermore, future studies should examine the consequences of changes in community composition due to hydropower production. Changes in species composition could potentially affect aquatic and terrestrial food webs since aquatic macroinvertebrates constitute an important food source for aquatic and terrestrial predators (e.g. Nakano & Murakami, 2001). Moreover, lower abundances or shifts in dominant species, as the ones seen in this study, could influence the invasion success of non-native species (Gilbert, Turkington & Srivastava, 2009) and have implications for ecosystem function since single species can be important in maintaining ecosystem functioning in tropical ecosystems (Taylor, Flecker & Hall, 2006).
Acknowledgments
This research was supported by the Center for Research of Marine Science and Limnology (CIMAR) at the University of Costa Rica (808-A3-065). Dartmouth College provided support for Chaves-Ulloa during the manuscript preparation. We are grateful with ICE and Coneléctricas; especially with J. Granados, A. Madrigal and J.A. Zúñiga from ICE and L. Torre from Coneléctricas. MINAET-SINAC provided collection permits. Thanks to Y. Gutiérrez, R. Lara, and J. Sibaja who assisted with field work. We also want to acknowledge L. Paniagua and R. Pizarro for preparation of Figure 1, and K. Cottingham, Z.J. Gezon, and M. Sasa for advice on data analysis. Y. Astorga, L.E. Culler, M.C. Duryea, S.B. Fey, Z.J. Gezon, E. Larson, S.M. McDermott, M. Overstrom-Coleman, M.A. McPeek and B.W. Taylor provided insightful comments on drafts of this manuscript. Z.J. Gezon was very helpful with the translation of the final document.
References
Agostinho, A. A., Pelicice, F. M. & Gomes, L. C. (2008). Dams and the fish fauna of the Neotropical region: impacts and management related to diversity and fisheries. Brazilian Journal of Biology, 68, 1119-1132. [ Links ]
Albrecht, M. P., Caramaschi, E. P. & Horn, M. H. (2009). Population responses of two omnivorous fish species to impoundment of a Brazilian tropical river. Hydrobiologia, 627, 181-193. doi: 10.1007/s10750-009-9727-7 [ Links ]
Anderson, E. P., Pringle, C. M. & Rojas, M. (2006). Transforming tropical rivers: an environmental perspective on hydropower development in Costa Rica. Aquatic Conservation-Marine and Freshwater Ecosystems, 16, 679-693. doi: 10.1002/aqc.806 [ Links ]
Araujo, F. G., Pinto, B. C. T. & Teixeira, T. P. (2009). Longitudinal patterns of fish assemblages in a large tropical river in southeastern Brazil: evaluating environmental influences and some concepts in river ecology. Hydrobiologia, 618, 89-107. doi: 10.1007/s10750-008-9551-5 [ Links ]
Barrantes, C., Alan, E. & Gallardo, R. (1990). Estudio de impacto ambiental proyecto hidroeléctrico San Lorenzo. San José: Consorcio de empresas de generación hidroeléctrica. Alajuela: Conelectrica R.L. [ Links ]
Benstead, J. P. (1996). Macroinvertebrates and the processing of leaf litter in a tropical stream. Biotropica, 28, 367-375. doi: 10.2307/2389200 [ Links ]
Brooker, M. P. & Hemworth, R. J. (1978). The effect of the release of an artificial discharge of water on invertebrate drift in the R. Wye, Wales. Hydrobiologia, 59, 155-163. [ Links ]
Bunn, S. E. & Arthington, A. H. (2002). Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environmental Management, 30, 492-507. doi: 10.1007/s00267-002-2737-0 [ Links ]
Chan, E. K. W., Zhang, Y. X. & Dudgeon, D. (2007). Contribution of adult aquatic insects to riparian prey availability along tropical forest streams. Marine and Freshwater Research, 58, 725-732. [ Links ]
Clarke, K. R. & Warwick, R. M. (2001). Change in marine communities: an approach to statistical analysis and interpretation (2nd ed.) Plymouth: P. [ Links ]
Conelectricas. (2013). Central hidroeléctrica San Lorenzo. Retrieved from http://conelectricas.com/centrales-hidroelectricas/central-hidroelectrica-san-lorenzo [ Links ]
Cooney, P. B & Kwak, T. J. (2013). Spatial extent and dynamics of dam impacts on tropical island freshwater fish assemblages. BioScience, 63, 176-190. [ Links ]
Cummins, K. W., Merritt, R. W. & Andrade, P. C. N. (2005). The use of invertebrate functional groups to characterize ecosystem attributes in selected streams and rivers in south Brazil. Studies on Neotropical Fauna and Environment, 40, 69-89. doi: 10.1080/01650520400025720 [ Links ]
Cushman, R. M. (1985). Review of ecological effects of rapidly varying flows downstream from hydroelectric facilities. North American Journal of Fisheries Management, 5, 330-339. [ Links ]
Dejalon, D. G. & Sanchez, P. (1994). Downstream effects of a new hydropower impoundment on macrophyte, macroinvertebrate and fish communities. Regulated Rivers-Research & Management, 9, 253-261. doi: 10.1002/rrr.3450090406 [ Links ]
Doi, H., Chang, K. H., Ando, T., Imai, H., Nakano, S. I., Kajimoto, A. & Katano, I. (2008). Drifting plankton from a reservoir subsidize downstream food webs and alter community structure. Oecologia, 156, 363-371. doi: 10.1007/s00442-008-0988-z [ Links ]
Dudgeon, D., Arthington, A. H., Gessner, M. O., Kawabata, Z. I., Knowler, D. J., Lévêque, C., Naiman, R. J., Prieur-Richard, A., Soto, D., Stiassny, M. L. J. & Sullivan, C. A. (2006). Freshwater biodiversity: importance, threats, status and conservation challenges. Biological Reviews, 81, 163-182. doi: 10.1017/s1464793105006950 [ Links ]
Esselman, P. C. & Opperman, J. J. (2010). Overcoming Information Limitations for the Prescription of an Environmental Flow Regime for a Central American River. Ecology and Society, 15, 6. [ Links ]
Finer, M. & Jenkins, C. N. (2012). Proliferation of Hydroelectric Dams in the Andean Amazon and Implications for Andes-Amazon Connectivity. Plos One 7, e35126. doi: 10.1371/journal.pone.0035126 [ Links ]
Foto Menbohan, S., Koji, E., Ajeagah, G., Bilong Bilong, C. F. & Njiné, T. (2012). Impact of dam construction on the diversity of benthic macroinvertebrates community in a periurban stream in Cameroon. International Journal of Biosciences, 2, 137-145. [ Links ]
Freeman, M. C., Pringle, C. M. & Jackson, C. R. (2007). Hydrologic connectivity and the contribution of stream headwaters to ecological integrity at regional scales. Journal of the American Water Resources Association, 43, 5-14. doi: 10.1111/j.1752-1688.2007.00002.x [ Links ]
Gilbert, B., Turkington, R. & Srivastava, D. S. (2009). Dominant Species and Diversity: Linking Relative Abundance to Controls of Species Establishment. American Naturalist, 174, 850-862. doi: 10.1086/647903 [ Links ]
Gotelli, N. J. & Colwell, R. K. (2001). Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters, 4, 379-391. doi: 10.1046/j.1461-0248.2001.00230.x [ Links ]
Greathouse, E. A., Pringle, C. M. & Holmquist, J. G. (2006a). Conservation and management of migratory fauna: dams in tropical streams of Puerto Rico. Aquatic Conservation-Marine and Freshwater Ecosystems, 16, 695-712. doi: 10.1002/aqc.804 [ Links ]
Greathouse, E. A., Pringle, C. M., McDowell, W. H. & Holmquist, J. G. (2006b). Indirect upstream effects of dams: Consequences of migratory consumer extirpation in Puerto Rico. Ecological Applications, 16, 339-352. doi: 10.1890/05-0243 [ Links ]
Gutiérrez, D. (1999). Estudio de impacto ambiental proyecto hidroeléctrico Peñas Blancas y la línea de transmisión. San José: Universidad de Costa Rica. [ Links ]
Hauer, F. R. & Lamberti, G. A. 2007. Methods in stream ecology. London: Elsevier. [ Links ]
Jorcin, A. & Nogueira, M. G. (2008). Benthic macroinvertebrates in the Paranapanema reservoir cascade (southeast Brazil). Brazilian Journal of Biology, 68, 1013-1024. [ Links ]
Jorcin, A., Nogueira, M. G. & Belmont, R. (2009). Spatial and temporal distribution of the zoobenthos community during the filling up period of Porto Primavera Reservoir (Parana River, Brazil). Brazilian Journal of Biology, 69, 19-29. [ Links ]
Kalff, J. (2002). Limnology. London: Prentice-Hall. [ Links ]
Katano, I., Negishi, J. N., Minagawa, T., Doi, H., Kawaguchi, Y. & Kayaba, Y. (2009). Longitudinal macroinvertebrate organization over contrasting discontinuities: effects of a dam and a tributary. Journal of the North American Benthological Society, 28, 331-351. doi: 10.1899/08-010.1 [ Links ]
La Nación. (2003, november 14th). ICE acepta responsabilidad en la muerte de peces. La Nación. Retrieved from http://wvw.nacion.com/ln_ee/2003/noviembre/14/pais7.html [ Links ]
Ligon, F. K., Dietrich, W. E. & Trush, W. J. (1995). Downstream ecological effects of dams. Bioscience, 45, 183-192. doi: 10.2307/1312557 [ Links ]
Lind, O. T. (1985). Handbook of common methods in limnology. Iowa: Kendall/Hunt Publishing Company. [ Links ]
March, J. G., Benstead, J. P., Pringle, C. M. & Scatena, F. N. (2003). Damming tropical island streams: Problems, solutions, and alternatives. Bioscience, 53, 1069-1078. doi: 10.1641/0006-3568(2003)053[1069:dtisps]2.0.co;2 [ Links ]
McLarney, W. O., Mafla, M. H., Arias, A. M. & Bouchonnet, D. (2010). The threat to biodiversity and ecosystem function of proposed hydroelectric dams in the La Amistad world heritage site, Panama and Costa Rica. (Report submitted to UNESCO World Heritage Committee). San José: Asociación ANAI (Costa Rica). [ Links ]
Merritt, R. W. & Cummins, K. W. (1996). An introduction to the aquatic insects of North America. Iowa: Kendall/Hunt. [ Links ]
Merritt, R. W., Cummins, K. W. & Berg, M. B. (2008). An introduction to the aquatic insects of North America (4th ed.). Iowa: Kendall/Hunt. [ Links ]
Minshall, W. & Winger, P. V. (1968). The Effect of Reduction in Stream Flow on Invertebrate Drift. Ecology, 49, 580-582. [ Links ]
Nakano, S. & Murakami, M. (2001). Reciprocal subsidies: Dynamic interdependence between terrestrial and aquatic food webs. Proceedings of the National Academy of Sciences of the United States of America, 98, 166-170. [ Links ]
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., O’hara, R. B., Simpson, G. L. & Wagner, H. (2010). Vegan: Community Ecology Package. R package version 1.17-4. Retrieved from http://CRAN.R-project.org/package=vegan [ Links ]
Ortaz, M., Martín, R. & Lopez-Ordaz, A. (2011). Variación espacial y temporal en la composición de la dieta de peces invertívoros en un río neotropical, Venezuela. Revista Biología Tropical, 59, 1217-1231. [ Links ]
Petts, G., Armitage, P. & Castella, E. (1993). Physical habitat changes and macroinvertebrate response to river regulation- the River Rede, UK. Regulated Rivers-Research & Management, 8, 167-178. doi: 10.1002/rrr.3450080119 [ Links ]
Petts, G. E. (1984). Impounded rivers: Perspectives for ecological management. New York: John-Wiley and sons. [ Links ]
Petts, G. E. (1990). Regulation of large rivers- Problems and possibilities for environmentally-sound river development in South-America. Interciencia, 15, 388-395. [ Links ]
Poff, N. L., Allan, J. D., Bain, M. B., Karr, J. R., Prestegaard, K. L., Richter, B. D., Sparks, R. E. & Stromberg, J. C. (1997). The natural flow regime. Bioscience, 47, 769-784. doi: 10.2307/1313099 [ Links ]
Pringle, C. M. (1997). Exploring how disturbance is transmitted upstream: Going against the flow. Journal of the North American Benthological Society, 16, 425-438. doi: 10.2307/1468028 [ Links ]
Pringle, C. M. (2000). Riverine conservation in tropical versus temperate regions: Ecological and socioeconomic considerations. In Boon, P. J., Davis B. R. & Petts G. E. (Eds.), Global perspectives on river conservation: science, policy and practice (pp. 367-378). New York, USA: John Wiley and Sons Ltd. [ Links ]
Pringle, C. M., Freeman, M. C. & Freeman, B. J. (2000). Regional effects of hydrologic alterations on riverine macrobiota in the New World: Tropical-temperate comparisons. Bioscience, 50, 807-823. doi: 10.1641/0006-3568(2000)050[0807:reohao]2.0.co;2 [ Links ]
Pringle, C. M. (2001). Hydrologic connectivity and the management of biological reserves: A global perspective. Ecological Applications, 11, 981-998. doi: 10.2307/3061006 [ Links ]
R development core team. (2009). R: A language and environment for statistical computing. Vienna, Austria ISBN 3-900051-07-0, URL R foundation for statistical computing. [ Links ]
Rabeni, C. F., Doisy, K. E. & Zweig, L. D. (2005). Stream invertebrate community functional responses to deposited sediment. Aquatic Sciences, 67, 395-402. doi: 10.1007/s00027-005-0793-2 [ Links ]
Regalado, A. (2011). Brazil gives green light to ‘Monster Dam’: Science Insider (Ecology). Science. Retrieved from http://news.sciencemag.org/scienceinsider/2011/06/brazil-gives-green-light-to-monster-dam.html [ Links ]
Revenga, C., Brunner, J., Henninger, N., Kassem, K. & Payne, R. (2000). Pilot analysis of global ecosystems: freshwater systems. Retrieved from http://www.wri.org/wr2000: World Resources Institute. [ Links ]
Rodrigues, S. C. & Silva, T. I. (2012). Dam construction and loss of geodiversity in the Araguari River basin, Brazil. Land Degradation & Development, 23, 419-426. doi: 10.1002/ldr.2157 [ Links ]
Roldán, G. (1996). Guía para el estudio de los macroinvertebrados acuáticos del Departamento de Antioquia. Antioquia: Editorial Presencia Ltda. [ Links ]
SAS institute Inc. (2009). JMP® version 8. Cary, NC, USA. [ Links ]
Scatena, F. N. (2004). A survey of methods for setting minimum instream flow standards in the Caribbean Basin. River Research and Applications, 20, 127-135. doi: 10.1002/rra.738 [ Links ]
Scullion, N. J. & Scinton A. (1983). Effects of artificial freshets on substratum composition, benthic invertebrate fauna and invertebrate drift in two impounded rivers in mid-Wales. Hydrobiologia, 107, 261-269. [ Links ]
Sinokrot, B. A. & Gulliver, J. S. (2000). In-stream flow impact on river water temperatures. Journal of Hydraulic Research, 38, 339-349. [ Links ]
Springer, M., Ramírez, A. & Hanson, P. (2010). Macroinvertebrados de agua dulce de Costa Rica. Revista Biologia Tropical, 58, 1-240. [ Links ]
Stevens, L. E., Shannon, J. P., & Blinn, D. W. (1997). Colorado River benthic ecology in Grand Canyon, Arizona, USA: Dam, tributary and geomorphological influences. Regulated Rivers-Research & Management, 13, 129-149. doi: 10.1002/(sici)1099-1646(199703)13:2<129::aid-rrr431>3.0.co;2-s [ Links ]
Takao, A., Kawaguchi, Y., Minagawa, T., Kayaba, Y. & Morimoto, Y. (2008). The relationchips between benthic macroinvertebrates and biotic and abiotic environmental characteristics downstream of the Yahagi dam, Central Japan, and the state chage caused by inflow from a tributary. River Research and Applications, 24, 580-597. [ Links ]
Taylor, B. W., Flecker, A. S. & Hall, R. O. (2006). Loss of a harvested fish species disrupts carbon flow in a diverse tropical river. Science, 313, 833-836. doi: 10.1126/science.1128223 [ Links ]
Terra, B. D., dos Santos, A. B. I. & Araujo, F. G. (2010). Fish assemblage in a dammed tropical river: an analysis along the longitudinal and temporal gradients from river to reservoir. Neotropical Ichthyology, 8, 599-606. [ Links ]
Tollefson, J. (2011). Worth a dam? Nature, 474, 430-430. [ Links ]
Tomanova, S., Goitia, E. & Helesic, J. (2006). Trophic levels and functional feeding groups of macroinvertebrates in neotropical streams. Hydrobiologia, 556, 251-264. doi: 10.1007/s10750-005-1255-5 [ Links ]
Troelstrup, N. H. & Hergenrader, G. L. (1990). Effect of hydropower peaking flow fluctuations on community structure and feeding guilds of invertebrates colonizing artificial substrates in a large impounded river. Hydrobiologia, 199, 217-228. doi: 10.1007/bf00006354 [ Links ]
Viana, J. P. (2002). Physical and chemical post-dam alterations in the Jamari River, a hydroelectric-developed river of the Brazilian Amazon. Hydrobiologia, 472, 235-247. doi: 10.1023/a:1016389919574 [ Links ]
Vinson, M. R. (2001). Long-term dynamics of an invertebrate assemblage downstream from a large dam. Ecological Applications, 11, 711-730. doi: 10.2307/3061112 [ Links ]
Wantzen, K. M. & Wagner, R. (2006). Detritus processing by invertebrate shredders: a neotropical-temperate comparison. Journal of the North American Benthological Society, 25, 216-232. doi: 10.1899/0887-3593(2006)25[216:dpbisa]2.0.co;2 [ Links ]
Ward, J. V. & Stanford, J. A. (1983). The serial discontinuity concept of lotic ecosystems. In Fontaine T. D. & Bartell S. M. (Eds.), Dynamics of lotic ecosystems (pp. 29-42). Ann Arbor, Mich: Ann Arbor Scientific Publishers. [ Links ]
Williams, J. G., Armstrong, G., Katopodis, C., Lariniere, M. & Travade, F. (2012). Thinking like a fish: a key ingredient for development of effective fish passage facilities at river obstructions. River Research and Applications, 28, 407-417. doi: 10.1002/rra.1551 [ Links ]
World Commission on Dams (WCD). (2000). Dams and development: a new framework for decision- making. London: Earthscan Publications. [ Links ]
Wright, M. S., & Covich, A. P. (2005). The effect of macroinvertebrate exclusion on leaf breakdown rates in a tropical headwater stream. Biotropica, 37, 403-408. doi: 10.1111/j.1744-7429.2005.00053.x [ Links ]
Zeledón, J. M. (2001). Aprovechamiento Hidroeléctrico en Costa Rica: Experiencia en la apertura de la generación hidroeléctrica y el desarrollo sostenible. Paper presented at the III Encuentro de las Aguas, Santiago de Chile, 24-26 oct. 2001. Santiago de Chile. [ Links ]
Albrecht, M. P., Caramaschi, E. P. & Horn, M. H. (2009). Population responses of two omnivorous fish species to impoundment of a Brazilian tropical river. Hydrobiologia, 627, 181-193. doi: 10.1007/s10750-009-9727-7 [ Links ]
Anderson, E. P., Pringle, C. M. & Rojas, M. (2006). Transforming tropical rivers: an environmental perspective on hydropower development in Costa Rica. Aquatic Conservation-Marine and Freshwater Ecosystems, 16, 679-693. doi: 10.1002/aqc.806 [ Links ]
Araujo, F. G., Pinto, B. C. T. & Teixeira, T. P. (2009). Longitudinal patterns of fish assemblages in a large tropical river in southeastern Brazil: evaluating environmental influences and some concepts in river ecology. Hydrobiologia, 618, 89-107. doi: 10.1007/s10750-008-9551-5 [ Links ]
Barrantes, C., Alan, E. & Gallardo, R. (1990). Estudio de impacto ambiental proyecto hidroeléctrico San Lorenzo. San José: Consorcio de empresas de generación hidroeléctrica. Alajuela: Conelectrica R.L. [ Links ]
Benstead, J. P. (1996). Macroinvertebrates and the processing of leaf litter in a tropical stream. Biotropica, 28, 367-375. doi: 10.2307/2389200 [ Links ]
Brooker, M. P. & Hemworth, R. J. (1978). The effect of the release of an artificial discharge of water on invertebrate drift in the R. Wye, Wales. Hydrobiologia, 59, 155-163. [ Links ]
Bunn, S. E. & Arthington, A. H. (2002). Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environmental Management, 30, 492-507. doi: 10.1007/s00267-002-2737-0 [ Links ]
Chan, E. K. W., Zhang, Y. X. & Dudgeon, D. (2007). Contribution of adult aquatic insects to riparian prey availability along tropical forest streams. Marine and Freshwater Research, 58, 725-732. [ Links ]
Clarke, K. R. & Warwick, R. M. (2001). Change in marine communities: an approach to statistical analysis and interpretation (2nd ed.) Plymouth: P. [ Links ]
Conelectricas. (2013). Central hidroeléctrica San Lorenzo. Retrieved from http://conelectricas.com/centrales-hidroelectricas/central-hidroelectrica-san-lorenzo [ Links ]
Cooney, P. B & Kwak, T. J. (2013). Spatial extent and dynamics of dam impacts on tropical island freshwater fish assemblages. BioScience, 63, 176-190. [ Links ]
Cummins, K. W., Merritt, R. W. & Andrade, P. C. N. (2005). The use of invertebrate functional groups to characterize ecosystem attributes in selected streams and rivers in south Brazil. Studies on Neotropical Fauna and Environment, 40, 69-89. doi: 10.1080/01650520400025720 [ Links ]
Cushman, R. M. (1985). Review of ecological effects of rapidly varying flows downstream from hydroelectric facilities. North American Journal of Fisheries Management, 5, 330-339. [ Links ]
Dejalon, D. G. & Sanchez, P. (1994). Downstream effects of a new hydropower impoundment on macrophyte, macroinvertebrate and fish communities. Regulated Rivers-Research & Management, 9, 253-261. doi: 10.1002/rrr.3450090406 [ Links ]
Doi, H., Chang, K. H., Ando, T., Imai, H., Nakano, S. I., Kajimoto, A. & Katano, I. (2008). Drifting plankton from a reservoir subsidize downstream food webs and alter community structure. Oecologia, 156, 363-371. doi: 10.1007/s00442-008-0988-z [ Links ]
Dudgeon, D., Arthington, A. H., Gessner, M. O., Kawabata, Z. I., Knowler, D. J., Lévêque, C., Naiman, R. J., Prieur-Richard, A., Soto, D., Stiassny, M. L. J. & Sullivan, C. A. (2006). Freshwater biodiversity: importance, threats, status and conservation challenges. Biological Reviews, 81, 163-182. doi: 10.1017/s1464793105006950 [ Links ]
Esselman, P. C. & Opperman, J. J. (2010). Overcoming Information Limitations for the Prescription of an Environmental Flow Regime for a Central American River. Ecology and Society, 15, 6. [ Links ]
Finer, M. & Jenkins, C. N. (2012). Proliferation of Hydroelectric Dams in the Andean Amazon and Implications for Andes-Amazon Connectivity. Plos One 7, e35126. doi: 10.1371/journal.pone.0035126 [ Links ]
Foto Menbohan, S., Koji, E., Ajeagah, G., Bilong Bilong, C. F. & Njiné, T. (2012). Impact of dam construction on the diversity of benthic macroinvertebrates community in a periurban stream in Cameroon. International Journal of Biosciences, 2, 137-145. [ Links ]
Freeman, M. C., Pringle, C. M. & Jackson, C. R. (2007). Hydrologic connectivity and the contribution of stream headwaters to ecological integrity at regional scales. Journal of the American Water Resources Association, 43, 5-14. doi: 10.1111/j.1752-1688.2007.00002.x [ Links ]
Gilbert, B., Turkington, R. & Srivastava, D. S. (2009). Dominant Species and Diversity: Linking Relative Abundance to Controls of Species Establishment. American Naturalist, 174, 850-862. doi: 10.1086/647903 [ Links ]
Gotelli, N. J. & Colwell, R. K. (2001). Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters, 4, 379-391. doi: 10.1046/j.1461-0248.2001.00230.x [ Links ]
Greathouse, E. A., Pringle, C. M. & Holmquist, J. G. (2006a). Conservation and management of migratory fauna: dams in tropical streams of Puerto Rico. Aquatic Conservation-Marine and Freshwater Ecosystems, 16, 695-712. doi: 10.1002/aqc.804 [ Links ]
Greathouse, E. A., Pringle, C. M., McDowell, W. H. & Holmquist, J. G. (2006b). Indirect upstream effects of dams: Consequences of migratory consumer extirpation in Puerto Rico. Ecological Applications, 16, 339-352. doi: 10.1890/05-0243 [ Links ]
Gutiérrez, D. (1999). Estudio de impacto ambiental proyecto hidroeléctrico Peñas Blancas y la línea de transmisión. San José: Universidad de Costa Rica. [ Links ]
Hauer, F. R. & Lamberti, G. A. 2007. Methods in stream ecology. London: Elsevier. [ Links ]
Jorcin, A. & Nogueira, M. G. (2008). Benthic macroinvertebrates in the Paranapanema reservoir cascade (southeast Brazil). Brazilian Journal of Biology, 68, 1013-1024. [ Links ]
Jorcin, A., Nogueira, M. G. & Belmont, R. (2009). Spatial and temporal distribution of the zoobenthos community during the filling up period of Porto Primavera Reservoir (Parana River, Brazil). Brazilian Journal of Biology, 69, 19-29. [ Links ]
Kalff, J. (2002). Limnology. London: Prentice-Hall. [ Links ]
Katano, I., Negishi, J. N., Minagawa, T., Doi, H., Kawaguchi, Y. & Kayaba, Y. (2009). Longitudinal macroinvertebrate organization over contrasting discontinuities: effects of a dam and a tributary. Journal of the North American Benthological Society, 28, 331-351. doi: 10.1899/08-010.1 [ Links ]
La Nación. (2003, november 14th). ICE acepta responsabilidad en la muerte de peces. La Nación. Retrieved from http://wvw.nacion.com/ln_ee/2003/noviembre/14/pais7.html [ Links ]
Ligon, F. K., Dietrich, W. E. & Trush, W. J. (1995). Downstream ecological effects of dams. Bioscience, 45, 183-192. doi: 10.2307/1312557 [ Links ]
Lind, O. T. (1985). Handbook of common methods in limnology. Iowa: Kendall/Hunt Publishing Company. [ Links ]
March, J. G., Benstead, J. P., Pringle, C. M. & Scatena, F. N. (2003). Damming tropical island streams: Problems, solutions, and alternatives. Bioscience, 53, 1069-1078. doi: 10.1641/0006-3568(2003)053[1069:dtisps]2.0.co;2 [ Links ]
McLarney, W. O., Mafla, M. H., Arias, A. M. & Bouchonnet, D. (2010). The threat to biodiversity and ecosystem function of proposed hydroelectric dams in the La Amistad world heritage site, Panama and Costa Rica. (Report submitted to UNESCO World Heritage Committee). San José: Asociación ANAI (Costa Rica). [ Links ]
Merritt, R. W. & Cummins, K. W. (1996). An introduction to the aquatic insects of North America. Iowa: Kendall/Hunt. [ Links ]
Merritt, R. W., Cummins, K. W. & Berg, M. B. (2008). An introduction to the aquatic insects of North America (4th ed.). Iowa: Kendall/Hunt. [ Links ]
Minshall, W. & Winger, P. V. (1968). The Effect of Reduction in Stream Flow on Invertebrate Drift. Ecology, 49, 580-582. [ Links ]
Nakano, S. & Murakami, M. (2001). Reciprocal subsidies: Dynamic interdependence between terrestrial and aquatic food webs. Proceedings of the National Academy of Sciences of the United States of America, 98, 166-170. [ Links ]
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., O’hara, R. B., Simpson, G. L. & Wagner, H. (2010). Vegan: Community Ecology Package. R package version 1.17-4. Retrieved from http://CRAN.R-project.org/package=vegan [ Links ]
Ortaz, M., Martín, R. & Lopez-Ordaz, A. (2011). Variación espacial y temporal en la composición de la dieta de peces invertívoros en un río neotropical, Venezuela. Revista Biología Tropical, 59, 1217-1231. [ Links ]
Petts, G., Armitage, P. & Castella, E. (1993). Physical habitat changes and macroinvertebrate response to river regulation- the River Rede, UK. Regulated Rivers-Research & Management, 8, 167-178. doi: 10.1002/rrr.3450080119 [ Links ]
Petts, G. E. (1984). Impounded rivers: Perspectives for ecological management. New York: John-Wiley and sons. [ Links ]
Petts, G. E. (1990). Regulation of large rivers- Problems and possibilities for environmentally-sound river development in South-America. Interciencia, 15, 388-395. [ Links ]
Poff, N. L., Allan, J. D., Bain, M. B., Karr, J. R., Prestegaard, K. L., Richter, B. D., Sparks, R. E. & Stromberg, J. C. (1997). The natural flow regime. Bioscience, 47, 769-784. doi: 10.2307/1313099 [ Links ]
Pringle, C. M. (1997). Exploring how disturbance is transmitted upstream: Going against the flow. Journal of the North American Benthological Society, 16, 425-438. doi: 10.2307/1468028 [ Links ]
Pringle, C. M. (2000). Riverine conservation in tropical versus temperate regions: Ecological and socioeconomic considerations. In Boon, P. J., Davis B. R. & Petts G. E. (Eds.), Global perspectives on river conservation: science, policy and practice (pp. 367-378). New York, USA: John Wiley and Sons Ltd. [ Links ]
Pringle, C. M., Freeman, M. C. & Freeman, B. J. (2000). Regional effects of hydrologic alterations on riverine macrobiota in the New World: Tropical-temperate comparisons. Bioscience, 50, 807-823. doi: 10.1641/0006-3568(2000)050[0807:reohao]2.0.co;2 [ Links ]
Pringle, C. M. (2001). Hydrologic connectivity and the management of biological reserves: A global perspective. Ecological Applications, 11, 981-998. doi: 10.2307/3061006 [ Links ]
R development core team. (2009). R: A language and environment for statistical computing. Vienna, Austria ISBN 3-900051-07-0, URL R foundation for statistical computing. [ Links ]
Rabeni, C. F., Doisy, K. E. & Zweig, L. D. (2005). Stream invertebrate community functional responses to deposited sediment. Aquatic Sciences, 67, 395-402. doi: 10.1007/s00027-005-0793-2 [ Links ]
Regalado, A. (2011). Brazil gives green light to ‘Monster Dam’: Science Insider (Ecology). Science. Retrieved from http://news.sciencemag.org/scienceinsider/2011/06/brazil-gives-green-light-to-monster-dam.html [ Links ]
Revenga, C., Brunner, J., Henninger, N., Kassem, K. & Payne, R. (2000). Pilot analysis of global ecosystems: freshwater systems. Retrieved from http://www.wri.org/wr2000: World Resources Institute. [ Links ]
Rodrigues, S. C. & Silva, T. I. (2012). Dam construction and loss of geodiversity in the Araguari River basin, Brazil. Land Degradation & Development, 23, 419-426. doi: 10.1002/ldr.2157 [ Links ]
Roldán, G. (1996). Guía para el estudio de los macroinvertebrados acuáticos del Departamento de Antioquia. Antioquia: Editorial Presencia Ltda. [ Links ]
SAS institute Inc. (2009). JMP® version 8. Cary, NC, USA. [ Links ]
Scatena, F. N. (2004). A survey of methods for setting minimum instream flow standards in the Caribbean Basin. River Research and Applications, 20, 127-135. doi: 10.1002/rra.738 [ Links ]
Scullion, N. J. & Scinton A. (1983). Effects of artificial freshets on substratum composition, benthic invertebrate fauna and invertebrate drift in two impounded rivers in mid-Wales. Hydrobiologia, 107, 261-269. [ Links ]
Sinokrot, B. A. & Gulliver, J. S. (2000). In-stream flow impact on river water temperatures. Journal of Hydraulic Research, 38, 339-349. [ Links ]
Springer, M., Ramírez, A. & Hanson, P. (2010). Macroinvertebrados de agua dulce de Costa Rica. Revista Biologia Tropical, 58, 1-240. [ Links ]
Stevens, L. E., Shannon, J. P., & Blinn, D. W. (1997). Colorado River benthic ecology in Grand Canyon, Arizona, USA: Dam, tributary and geomorphological influences. Regulated Rivers-Research & Management, 13, 129-149. doi: 10.1002/(sici)1099-1646(199703)13:2<129::aid-rrr431>3.0.co;2-s [ Links ]
Takao, A., Kawaguchi, Y., Minagawa, T., Kayaba, Y. & Morimoto, Y. (2008). The relationchips between benthic macroinvertebrates and biotic and abiotic environmental characteristics downstream of the Yahagi dam, Central Japan, and the state chage caused by inflow from a tributary. River Research and Applications, 24, 580-597. [ Links ]
Taylor, B. W., Flecker, A. S. & Hall, R. O. (2006). Loss of a harvested fish species disrupts carbon flow in a diverse tropical river. Science, 313, 833-836. doi: 10.1126/science.1128223 [ Links ]
Terra, B. D., dos Santos, A. B. I. & Araujo, F. G. (2010). Fish assemblage in a dammed tropical river: an analysis along the longitudinal and temporal gradients from river to reservoir. Neotropical Ichthyology, 8, 599-606. [ Links ]
Tollefson, J. (2011). Worth a dam? Nature, 474, 430-430. [ Links ]
Tomanova, S., Goitia, E. & Helesic, J. (2006). Trophic levels and functional feeding groups of macroinvertebrates in neotropical streams. Hydrobiologia, 556, 251-264. doi: 10.1007/s10750-005-1255-5 [ Links ]
Troelstrup, N. H. & Hergenrader, G. L. (1990). Effect of hydropower peaking flow fluctuations on community structure and feeding guilds of invertebrates colonizing artificial substrates in a large impounded river. Hydrobiologia, 199, 217-228. doi: 10.1007/bf00006354 [ Links ]
Viana, J. P. (2002). Physical and chemical post-dam alterations in the Jamari River, a hydroelectric-developed river of the Brazilian Amazon. Hydrobiologia, 472, 235-247. doi: 10.1023/a:1016389919574 [ Links ]
Vinson, M. R. (2001). Long-term dynamics of an invertebrate assemblage downstream from a large dam. Ecological Applications, 11, 711-730. doi: 10.2307/3061112 [ Links ]
Wantzen, K. M. & Wagner, R. (2006). Detritus processing by invertebrate shredders: a neotropical-temperate comparison. Journal of the North American Benthological Society, 25, 216-232. doi: 10.1899/0887-3593(2006)25[216:dpbisa]2.0.co;2 [ Links ]
Ward, J. V. & Stanford, J. A. (1983). The serial discontinuity concept of lotic ecosystems. In Fontaine T. D. & Bartell S. M. (Eds.), Dynamics of lotic ecosystems (pp. 29-42). Ann Arbor, Mich: Ann Arbor Scientific Publishers. [ Links ]
Williams, J. G., Armstrong, G., Katopodis, C., Lariniere, M. & Travade, F. (2012). Thinking like a fish: a key ingredient for development of effective fish passage facilities at river obstructions. River Research and Applications, 28, 407-417. doi: 10.1002/rra.1551 [ Links ]
World Commission on Dams (WCD). (2000). Dams and development: a new framework for decision- making. London: Earthscan Publications. [ Links ]
Wright, M. S., & Covich, A. P. (2005). The effect of macroinvertebrate exclusion on leaf breakdown rates in a tropical headwater stream. Biotropica, 37, 403-408. doi: 10.1111/j.1744-7429.2005.00053.x [ Links ]
Zeledón, J. M. (2001). Aprovechamiento Hidroeléctrico en Costa Rica: Experiencia en la apertura de la generación hidroeléctrica y el desarrollo sostenible. Paper presented at the III Encuentro de las Aguas, Santiago de Chile, 24-26 oct. 2001. Santiago de Chile. [ Links ]
1. Biological Sciences Dept., Dartmouth College, 78 College St. Hanover, NH 03755 USA. Escuela de Biología, Universidad de Costa Rica, 2060 San José, Costa Rica; Ramsa.Chaves.Ulloa.GR@dartmouth.edu
2. Escuela de Biología, Universidad de Costa Rica, 2060 San José, Costa Rica. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Universidad de Costa Rica, 2060 San José, Costa Rica; gerardo.umana@ucr.ac.cr
3. Escuela de Biología, Universidad de Costa Rica, 2060 San José, Costa Rica. Centro de Investigación en Ciencias del Mar y Limnología (CIMAR), Universidad de Costa Rica, 2060 San José, Costa Rica; monika.springer@ucr.ac.cr
Received 12-XII-2013. Corrected 20-I-2014. Accepted 13-II-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}