










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.2 San José Apr. 2014
Effects of insect and decapod exclusion and leaf litter species identity on breakdown rates in a tropical headwater stream
Abstract
High species richness of tropical riparian trees influences the diversity of organic detritus entering streams, creating temporal variability in litter quantity and quality. We examined the influence of species of riparian plants and macroinvertebrate exclusion on leaf-litter breakdown in a headwater stream in the Luquillo Mountains of Puerto Rico. Leaf litter of Dacryodes excelsa (Burseraceae), Guarea guidonia (Meliaceae), Cecropia scheberiana (Moraceae), Manilkara bidentata (Sapotaceae), and Prestoea acuminata (Palmae) were incubated in litter bags in a pool of Quebrada Prieta. Fine mesh bags were used to exclude macroinvertebrates during leaf breakdown, and coarse mesh bags allowed access to decapod crustaceans (juvenile shrimps and crabs) and aquatic insects (mainly mayflies, chironomids, and caddisflies). D. excelsa and G. guidonia (in coarse- and fine-mesh bags) had significantly higher breakdown rates than C. scheberiana, M. bidentata, and P. acuminata. Breakdown rates were significantly faster in coarse-mesh bag treatments for all leaf types, thus indicating a positive contribution of macroinvertebrates in leaf litter breakdown in this headwater stream. After 42 days of incubation, densities of total invertebrates, mayflies and caddisflies, were higher in bags with D. excelsa and G. guidonia, and lower in P. acuminata, C. scheberiana y M. bidentata. Decay rates were positively correlated to insect densities. Our study highlights the importance of leaf identity and macroinvertebrate exclusion on the process of leaf litter breakdown in tropical headwater streams. Rev. Biol. Trop. 62 (Suppl. 2): 143-154. Epub 2014 April 01.
Key words: leaf breakdown, aquatic insects, tropical stream, macroconsumers, detritivores, Puerto Rico, tropical rainforest.
Resumen
La gran variedad de especies vegetales junto a los ríos tropicales afecta la diversidad de detritos orgánicos que llegan a los arroyos, creando variaciones temporales en cantidad y calidad de hojarasca. Examinamos la influencia de las especies vegetales y de la exclusión de macroinvertebrados sobre la descomposición de hojarasca en la parte alta de un arroyo en las montañas Luquillo de Puerto Rico. Incubamos hojarasca de Dacryodes excelsa (Burseraceae), Guarea guidonia (Meliaceae), Cecropia scheberiana (Moraceae), Manilkara bidentata (Sapotaceae) y Prestoea acuminata (Palmae) en bolsas descomposición que colocamos en una poza de Quebrada Prieta. Usamos bolsas de malla fina para excluir los macroinvertebrados y de malla gruesa para permitir la entrada a crustáceos decápodos (camarones y cangrejos jóvenes) e insectos acuáticos (principalmente efemerópteros, quironómidos y tricópteros). D. excelsa y G. guidonia (en ambos tipos de malla) tuvieron tasas de descomposición más altas que C. scheberiana, M. bidentata y P. acuminata. La descomposición fue más rápida para todas las especies de hoja con la malla gruesa, resaltando el papel de los descomponedores en este hábitat. Tras 42 días de esta incubación, las densidades totales de invertebrados, efemerópteros y tricópteros fueron mayores en las bolsas con hojas de D. excelsa y G. guidonia, e inferiores en P. acuminata, C. scheberiana y M. bidentata. A mayor densidad de insectos hubo mayor tasa de descomposición. Nuestros resultados indican la importancia de la especie de las hojas y de la presencia de macroinvertebrados en el proceso de descomposición de hojarasca en partes altas de arroyos tropicales.
Palabras clave: descomposición de hojarasca, insectos acuáticos, arroyo tropical, macroconsumidores, detritívoros, Puerto Rico, bosque lluvioso tropical.
Riparian plants provide essential inputs of organic detritus that alter food-web dynamics in streams by determining the quantity and quality of organic litter inputs (Naiman & Décamps, 1997; Wallace, Eggert, Meyer & Webster, 1999). Species-specific differences in leaf breakdown rates are especially well documented for a large number of tree species in temperate streams (e.g., Petersen & Cummins, 1974; Webster & Benfield, 1986; Feio & Graça, 2000). Moreover, rates of litter breakdown in temperate-zone forested streams are also known to differ in response to differences in diversity of both riparian tree species and benthic detritivore species (Jonsson & Malmqvist, 2000; Quinn, Smith, Burrell & Parkyn, 2000; Jonsson, Malmqvist & Hoffsten, 2001; Usio & Townsend, 2001). However, much less is known about leaf litter processing in tropical streams (e.g., Irons, Oswood, Stout & Pringle, 1994; Benstead, 1996; Rosemond, Pringle & Ramirez, 1998; Graça, 2001; Boulton et al., 2008; Boyero et al., 2011; Boyero et al. 2012) and even less about species-specific effects on rates of litter processing (Covich, Palmer & Crowl, 1999; Mathooko, M’Erimba & Leichtfried, 2000; Crowl, McDowell, Covich & Johnson, 2001; Graça et al., 2001; Covich et al., 2004).
High species richness of tropical riparian trees influences the diversity of organic detritus entering streams, creating also temporal variability in litter quantity and quality (Covich 1988; Dudgeon 1994; Covich & McDowell 1996; Dobson, Mathooko, Ndegwa & M’erimba, 2003). Tropical riparian tree species differ greatly in terms of the thickness, fiber content, and secondary chemistry (e.g., Stout, 1989; Irons et al., 1994; Campbell & Fuchshuber, 1995; Wantzen, Wagner, Suetfeld & Junk, 2002; Ardón & Pringle, 2008; Ardón, Pringle & Eggert, 2009) and nutrient content (Gonçalves, Graça & Callisto, 2006) of their leaves. These chemical and physical differences may partially explain the observed species-specific differences in litter processing rates for tropical streams (Ardón et al., 2009).
In headwater streams of the Luquillo Mountains (Puerto Rico), terrestrially produced leaf litter, small twigs, and fruits constitute a relatively continuous input of detritus-based energy (Zou, Zucca, Waide & McDowell, 1995; Covich & McDowell, 1996; Vogt et al., 1996). Recently, Zalamea & González (2008) showed a bimodal pattern of leaf fall in this Puerto Rican forest. Rates of decomposition of these riparian inputs can be rapid, but different tree species have distinct patterns of decay that depend on the chemical contents and structure of their leaves (Padgett, 1976; Vogt et al., 1996; Wright & Covich, 2005; Crowl, Welsh, Heartsill-Scalley & Covich, 2006).
The rain forest in the El Verde Research Area (Puerto Rico) represents a mosaic of past land-use history (Zimmerman et al., 1995; Zou et al., 1995). Thus, the effect of forest deforestation on forest tree composition is still evident after 60 years of secondary succession. The mid-successional forest (MSF) was dominated by species like Cecropia scheberiana, and abundant occurrences of early/middle successional stages (Smith, 1970; Crow, 1980; Brokaw et al., 2012). In contrast, the mature tabonuco forest (MTF) was dominated by Dacryodes excelsa (tabonuco) and Guarea guidonia, species that are characteristic of a typical mature tabonuco forest (Crow, 1980). It has been suggested that decomposition in the MSF was faster than in the MTF, probably due to differences in leaf litter chemistry (Zou et al., 1995). Differences in leaf decay rates between successional species and primary forest species was correlated with lignin and fiber content of leaf litter in the same tabonuco forest (La Caro & Rudd, 1985). These differences in leaf decay rates between early successional species and primary forest species have important implications for energy flow and nutrient cycling in tropical streams following regrowth of secondary forests after disturbances (Wantzen et al., 2002; Beard et al., 2005; Wright & Covich, 2005; Zimmerman & Covich, 2007; Brokaw et al., 2012). Recent studies indicated that total literfall in the Luquillo Experimental Forest (LEF) was 7 504kg/ha/year and leaves accounted for 65.4% of total litterfall with the period with major leaf fall in April-June and August-September (Zalamaea & Gonzalez, 2008). The late succession species Dacryodes excelsa and Guarea guidonia showed a bimodal pattern with peaks in May-June and September-October, conversely Manilkara bidentata exhibited an unimodal pattern with a peak in September. On the other hand, the pioneer species Cecropia scheberiana showed a nonmodal pattern, although a December peak in leaf fall was associated to this species (Zalamea & González, 2008).
Our main objective was to examine how different riparian tree species that are representative of early and late stages of forest succession differ in their rates of detrital processing in a typical Puerto Rican headwater stream, Quebrada Prieta. Secondarily, we studied the effect of excluding decapods and insects on leaf litter breakdown. Previous field and lab studies on Luquillo headwater streams and their food web have typically focused on the influence of either insects (Ramírez & Hernández-Cruz, 2004; Bobeldyk & Ramírez, 2007) or shrimps (Crowl et al., 2001; Crowl et al., 2006; March, Benstead, Pringle & Ruebel, 2001) but very few on leaf litter species effects (Wright & Covich, 2005).
Materials and methods
Study area: Quebrada Prieta is a second-order tributary of the Quebrada Sonadora in the Río Espíritu Santo catchment that drains the steep hillslopes of the Luquillo Experimental Forest (18º18’N, 65º47’W) of northeastern Puerto Rico. We studied a reach at 490masl, which is representative of the steeply sloped stream channels. Riparian vegetation is dominated by Dacryodes excelsa Vahl, large patches of Cecropia scheberiana Miq., and the sierra palm, Prestoea acuminata (Graham) G. Nicholson. The distribution of these species along the riparian forest represents different patches of early and late succesional dynamics resulting from a long history of land use and tropical storms (Zou et al., 1995; Beard et al., 2005; Zimmerman & Covich, 2007). Stream water temperatures are consistently between 20 and 26ºC, and the average annual precipitation is 3 600mm, with May through December generally having greater monthly average rainfall than January-April (Garcia-Martino, Warner, Scatena & Civco, 1996; McDowell et al., 2012). High flows can occur at any time of the year, but the streams rapidly return to base-flow levels.
Plant species selection and decomposition experiment: Following the successional status described by Uriarte et al. (2012) and Brokaw et al. (2012), we used recently abscised dead leaves of three species of late successional trees: Dacryodes excelsa Vahl. (Burseraceae), Manilkara bidentata (A.DC.) A. Chev. (Sapotaceae), and Guarea guidonia (L.) Sleumer (Meliaceae); one pioneer successional tree, Cecropia scheberiana Miq. (Moraceae); and one species that occurs in both early and late successional communities forming nearly monospecific stands in hill slopes facing hurricanes tracks (Heartsill, Crowl & Thompson, 2009), P. acuminata var. montana (Palmae). Leaves were air dried and stored in plastic bags before use (Boulton & Boon, 1991). Approximately 3g of leaves of each tree species were placed into fine (0.3mm mesh, 15x15cm) and coarse (1x1.2cm mesh, 15x15cm) mesh nylon bags. Fine-mesh bags were used to exclude most macroinvertebrates and coarse-mesh bags allowed access of both macroconsumers (juvenile shrimps and crabs) and small aquatic insects (mainly mayflies, chironomids, and caddisflies). Sets of fine- and coarse-mesh bags of leaves were tethered to tree trunks along the riverbank. Three sets of litter bags (containing triplicates for both treatments and all five tree species) were returned to the laboratory to estimate initial fresh mass and the initial dry mass. A total of 180 litter bags were submerged in Q. Prieta on 6 March, 2000. We collected litter bags (n=3) of the five leaf species in each of the two bag types from the stream after 7, 14, 21, 28, 35, and 42 days after immersion. Litter bags were placed individually in plastic bags and transported to the laboratory in an ice chest. Immediately following retrieval, leaves were gently rinsed and macroinvertebrates and detritus were removed. Leaves were oven-dried at 60°C (48hrs) and then were weighed to the nearest 0.001g for the determination of dry mass. Macroinvertebrates were kept in ethanol (80%) and sorted, identified, and counted.
Effects of leaf type and macroinvertebrate exclusion on litter decomposition: Percent mass remaining through time was adjusted to an exponential model: mt/m0=e-k t; where mt is the mass remaining at time t, m0 is the initial mass, and k is the breakdown rate constant. Breakdown rates (-k) were estimated by non-linear regression (k ± asymptotic SE). The effect of leaf type and mesh size on breakdown rates was assessed using a Repeated Measures ANOVA and a Bonferroni pos hoc test was performed on data of remaining mass (loge). Invertebrate densities (total individuals, mayflies, and caddisflies) on leaf bags were expressed as numbers per unit mass of litter. Relationships between invertebrate density and breakdown rates were investigated using nonlinear regression analysis to fit original data to a straight-line model. Original data were tested for normality and homogeneity of variance before performing regression analysis and analysis of variance, and transformation was implemented when necessary. Statistical analyses were performed using Origin 6.0 (Microcal Software Inc. 1999), Prism 5.01 (GraphPad Software, Inc. 2007) and Statistica 4.3 (Stat-Soft, Inc. 1993).
Results
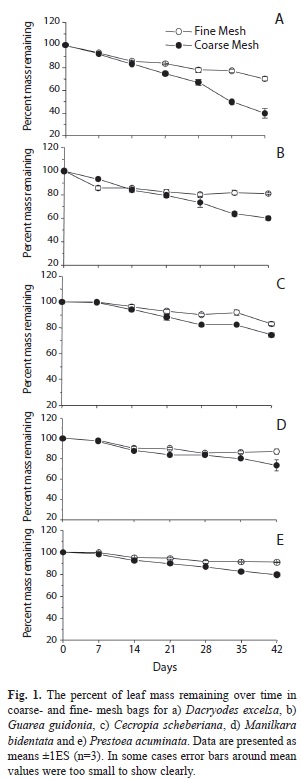
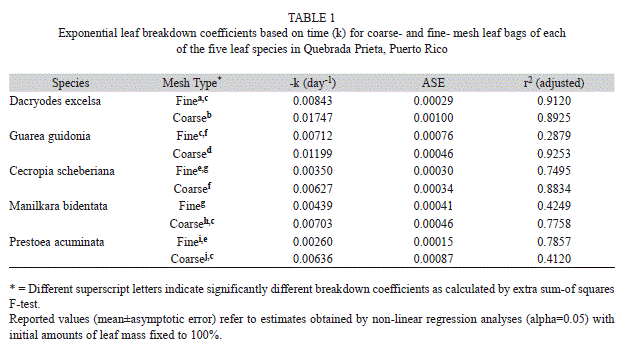
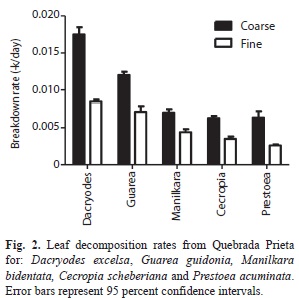
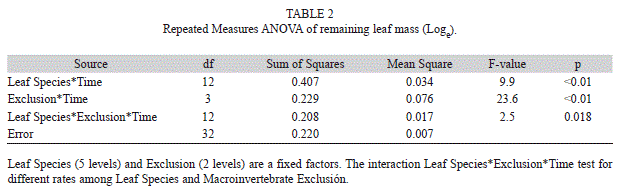
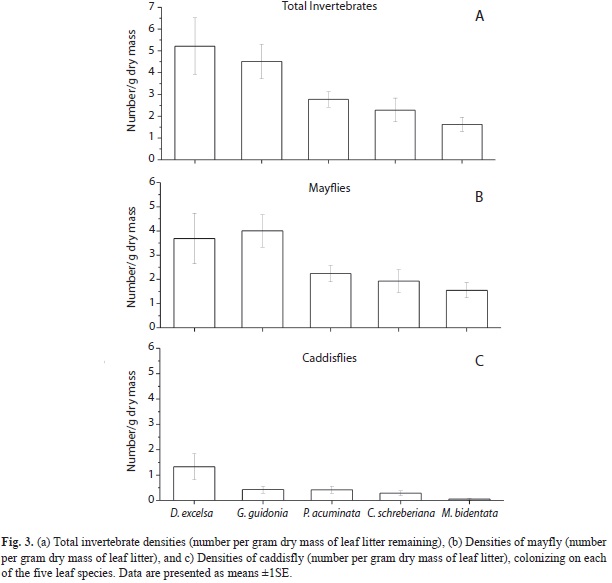
During the 42 days of incubation D. excelsa and G. guidonia showed the fastest breakdown rates in coarse-mesh bags (k=-0.01747±0.0010 and k=-0.01199±0.00033, 40% and 60% of mass remaining, respectively), whereas P. acuminata and C. shreberiana presented the slowest rates (k=-0.00636±0.00087 and k=-0.00627±0.00033; 80% and 74% of mass remaining, respectively; Fig. 1, Table 1). Similarly, in fine-mesh bags D. excelsa and G. guidonia exhibited the fastest rates (k=-0.007813±0.00030 and k=-0.007122±0.00076; 70% and 81% of mass remaining, respectively) in contrast with the slowest rates showed by C. shreberiana and P. acuminata (k=-0.0035±0.00030 and k=0.0026±0.00015). Leaf breakdown of M. bidentata was intermediate in both coarse- and fine-mesh bags (k=-0.007031±0.00046 and k=-0.004396±0.00041, 74% and 87% of mass remaining, respectively). Repeated Measures ANOVA indicated that the breakdown rate was significantly affected by mesh size and leaf type, but there was also a significant interaction between mesh size and leaf type (Fig. 2, Table 2). Breakdown rates of D. excelsa and G. guidonia in coarse-mesh bags were the fastest. Breakdown rates of C. scheberiana and P. acuminata in fine-mesh bags were slower than the other species. Breakdown rates of M. bidentata in coarse-mesh bags were not significantly different to rates of D. excelsa and G. guidonia in fine-mesh bags, and P. acuminata and C. sheberiana in coarse-mesh bags. Overall, breakdown rates in coarse-mesh bags were significantly faster than fine-mesh for all leaf types (Fig. 2, Table 2).
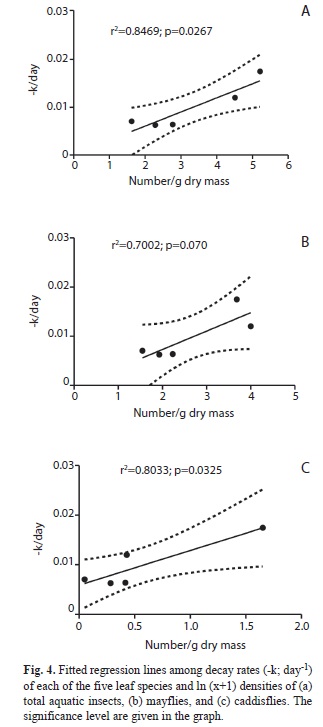
After 42 days of incubation, densities of total invertebrates and mayflies were highest in bags with D. excelsa and G. guidonia and lowest in P. acuminata, C. scheberiana and M. bidentata (Fig. 3). Mayflies dominated the invertebrate fauna colonizing litter bags. Larval Leptophlebiidae were the dominant mayfly group. Phylloicus pulchrus, a caddisfly and shredder, also colonized litter bags in lower densities and at slower rates compared to mayflies. Mayflies colonized leaves during the first seven days, increasing their densities through the time. Caddisflies colonized the litter bags after 14-21 days of incubation. The atyid shrimp, Atya lanipes, was found in coarse-mesh leaf packs of C. scheberiana 23 days after the beginning of the experiment. Determining shrimp abundance associated with bags was not possible, as they moved rapidly away from the coarse-mesh bags during periods of day-light observations and are especially active at night. However, previous studies have found high densities of both A. lanipes and X. elongate in this stream reach for more than a decade (Covich, Crowl & Scatena, 2003). Visual observation on leaf bags indicated that shrimps fed on litter from coarse mesh bags. Also some juvenile crabs were observed inside the coarse mesh bags. A linear relationship was found between decay rates and total invertebrate density (r2=0.80, p<0.05) and caddisfly density (r2=0.73, p<0.05), but not for mayfly density (r2=0.60, p>0.05) (Fig. 4).
Discussion
According to litter decay rates, we classified our leaves in coarse mesh bags (macroinvertebrate access) as follows: Dacryodes excelsa and Guarea guidonia are fast (kd=0.010-0.015); and Manilkara bidentata, Cecropia scheberiana, and Prestoea acuminata are in the medium category (kd=0.005-0.010). These processing coefficients were comparable to those measured in other tropical streams (e.g., Stout, 1989; Mathooko, 2000; Dobson et al., 2003; Wantzen, Yule, Mathooko & Pringle, 2008) and for these same riparian species in more open, wider channels (13 m) in this same drainage catchment (March et al., 2001). However, our k-values estimates for D. excelsa and C. scheberiana are lower than those reported by Wright & Covich (2005) in this same stream and this can likely be explained for the longer duration (84 days) and for the incubation of leaf discs instead of the whole leaf in their study.
Leaf litter breakdown differed significantly among species. Initial leaf chemical composition as reported by Sanchez, López & Lugo (1997) may have accounted for this species-specific difference in breakdown rates. Previous studies in the Luquillo Experimental Forest showed that D. excelsa has a lower content of crude fiber and lignin than C. scheberiana (LaCaro & Rudd, 1985). Furthermore, the palm P. acuminata is characterized by fronds with high content of fiber and lignocellullosic compounds that break down relatively slowly in streams (Vogt et al., 1996). Thus, those species with higher values of slow-degradation compounds may have high leaf toughness and consequently may retard the leaching process, providing an initial physical barrier against microbes and invertebrate detritivores. Early/middle successional species, as C. scheberiana and P. acuminata, seem to have developed different strategies against herbivores and nutrient limitation. A higher content of slow-degradation compounds in early successional trees may provide a nutrient-conserving function and a more effective defense against herbivory during early stages of forest succession (Coley & Barone, 2001). Overall, this difference in the content of slow-degradation compounds may explain, in part, the observed species-specific differences in leaf litter breakdown in our study. These species-specific differences have implications for energy flow and nutrient cycling in these tropical headwater food webs (Wright & Covich, 2005). For example, disturbances may change tree species composition and leaf litter chemistry, and consequently alter the leaf decomposition processes.
In this study, macroinvertebrates access to mesh bags accelerated breakdown rates for all leaf types. In concordance with Bobeldyk & Ramírez (2007), these results roughly indicate an important contribution of shrimps and insects in accelerating breakdown rates of leaf litter in this headwater stream. Although we did not quantify shrimp abundance, previous evidence from this stream suggests that shrimp may have accounted for differences in leaf breakdown between coarse- and fine-mesh bags in our study. Shrimps (A. lanipes and Xiphocaris elongata) are important breaking down leaf-litter in headwater streams in the Luquillo Mountains (Covich & McDowell, 1996; Crowl et al., 2006; Cross, Covich, Crowl, Benstead & Ramirez, 2008). March et al. (2001) experimentally attributed a fast rate of leaf decay primarily to shredding by shrimps (particularly X. elongata). They suggested that aquatic insects played a minor role in leaf decay at their high-elevation (300m) site on a larger stream (13m wide, 69% canopy cover), Quebrada Sonadora, Puerto Rico. Furthermore, Crowl et al. (2001) experimentally demonstrated that X. elongata increase rates of leaf-litter breakdown and that A. lanipes increase upstream retention of suspended, leaf-litter derived, particulates in Quebrada Prieta. Recently, it has been shown that, although this tropical island stream is dominated by decapods, there was a contribution (6%) of small, less abundant of detritivorous insects, on leaf breakdown (Bobeldyk & Ramírez, 2007).
We found the shredder Phylloicus pulchrus (Trichoptera, Calamoceratidae) colonizing coarse-mesh bags and, due to the shredder feeding habit of this insect, we suggest that this calamoceratid could be an additional component contributing to detrital processing despite their relatively low densities. The widely distributed tropical calamoceratid caddisfly (Phylloicus) cuts disks of leaves for case construction (Rincón & Martínez, 2006) and occurred in natural leaf packs in Quebrada Prieta. These shredding and case-building activities may have facilitated microbial access to the leaves. As aquatic insects scrape leaf surfaces or shred leaf edges, rates of leaf decay can be accelerated when aquatic fungi increase rates of penetration into the opened areas of the damaged leaf tissue (Jonsson & Malmqvist, 2000; Covich et al., 2004). This set of interactions among insect detritivores is more complex than that described for the two dominant species of shrimps that shred and filter feed (Crowl et al., 2001; Crowl et al., 2006). Other species, such as amphibious river crabs (e.g., Epilobocera sinuatifrons), can provide additional complexity to these processing chains by shredding leaves and also consuming aquatic insects associated with decomposing leaf packs (Zimmerman & Covich, 2003; Bobeldyk & Ramírez, 2007).
Although the importance of leaf-litter shredding by benthic detritivores in sustaining instream ecosystem processes is widely recognized (Covich et al., 1999; 2004), the current uncertainty regarding the relative values of dietary sources of carbon and the potential paucity of shredders in tropical streams likely results from several gaps of knowledge (Crowl et al., 2001; Dobson et al., 2002; Cheshire, Boyero & Pearson, 2005). Recently, Boyero et al. (2011) found that the shredder trophic guild is generally more prevalent and diverse in temperate streams, although variation at the regional scale within the tropics could be important. For example, the importance of freshwater crabs as shredders in some tropical streams requires wider documentation of their roles (Zimmerman & Covich, 2003; Dobson et al., 2003). Stable isotope analysis demonstrated a mixed diet for these crabs in nearby Quebrada Sonadora with reliance on leaf-derived carbon and some animal-derived carbon and nitrogen (March & Pringle, 2003). More comparisons of the quality of leaf-litter inputs and algae, as well as more complete sampling and observations on specific feeding behaviors of different detritivores are needed before regional generalizations are possible.
In summary, our results suggest that rates of organic matter processing may change depending on the interactive contribution of: 1) leaf species identity and associated quality and 2) the presence of macroinvertebrate shredders. Both aspects have been poorly studied in tropical streams. More focused studies using stable isotopes and intensive sampling of various types of decapods, as the crab Epilobocera sinuatifrons, and the caddisfly calamoceratid Phylloicus should be investigated.
Acknowledgments
We thank Fondo Nacional para la Investigación Científica y Tecnológica de Venezuela (FONACYT) (Grant Nº PI-99001007) and the University of Puerto Rico’s Institute for Tropical Ecosystem Studies (ITES) for partial funding of this research. Additional support was provided by the U.S. Forest Service, Colorado State University, and Universidad del Zulia. We thank Manuel Graça and Alonso Ramírez for providing helpful comments. We also appreciate the help and facilities provided by ITES at El Verde Field Station, Puerto Rico.
References
Ardon, M., & Pringle, C. M. (2008). Do secondary compounds inhibit microbial- and insect-mediated leaf breakdown in a tropical stream? Oecologia, 155, 311-323. [ Links ]
Ardon, M., Pringle, C.M., & Eggert, S.L. (2009). Does leaf chemistry differentially affect leaf breakdown in tropical versus temperate streams?: Importance of standardized analytical techniques to measure leaf chemistry. Journal of the North American Benthological Society, 28(2), 440-453 [ Links ]
Beard, K. H., Vogt, K. A., Vogt, D. J., Scatena, F. N., Covich, A. P., Siccama, T. C., Sigurdardottir, R., & Crowl, T. A. (2005). The response of structural and functional characteristics of a tropical forested ecosystem to multiple, natural disturbances in legacy environments. Ecological Monographs, 75, 345-361. [ Links ]
Benstead, J. P. (1996). Macroinvertebrates and the processing of leaf litter in a tropical stream. Biotropica, 28, 367-375. [ Links ]
Bobeldyk, A. M., & Ramírez, A. (2007). Leaf breakdown in a tropical headwater stream (Puerto Rico): The role of freshwater shrimps and detritivorous insects. Journal of Freshwater Ecology, 22, 581-590. [ Links ]
Boulton, A. J., & Boon, P. J. (1991). A review of methodology used to measure leaf litter decomposition in lotic environments: time to turn over a new leaf? Australian Journal of Marine and Freshwater Research, 42, 1-43. [ Links ]
Boulton, A. J., Boyero, L., Covich, A. P., Dobson, M., Lake, S., & Pearson, R. (2008) Are tropical streams ecologically different from temperate streams? In D. Dudgeon (Ed.), Tropical stream ecology (pp. 257-284). San Diego: Academic Press. [ Links ]
Boyero, L., Pearson, R. G., Dudgeon, D., Graca, M. A. S., Gessner, M. O., Albariño, R. J., Ferreira, V., Yule, C. M., Boulton, A. J., Arunachalam, M., Callisto, M., Chauvet, E., Ramirez, A., Chara, J., Moretti, M. S., Goncalves, J. F., Helson, J. E., Chara, J., Encalada, A. C., Davies, S. J., Lamothe, S., Cornejo, A., Li, Y., Buria, L. M., Villanueva, N., Zuniga, M. C., & Pringle, C. M. (2011). Global distribution of a key trophic guild contrasts with common latitudinal diversity patterns. Ecology, 92, 1839-1848. [ Links ]
Boyero, L., Richard, G., Pearson, R. G., Dudgeon, D., Ferreira, V., Graça, M. A. S., Gessner, M. O., Boulton, A. J., Chauvet, E., Yule, C. M., Albariño, R. J., Ramírez, A., Helson, J. E., Callisto, M., Arunachalam, M., Chará, J., Figueroa, R., Mathooko, J. M., Gonçalves, J. F., Moretti, M. S., Chará-Serna, A. M., Davies, J. N., Encalada, A., Lamothe, S., Buria, L. M., Castela, J., Cornejo, A., Li, A. O. Y., M’Erimba, C., Díaz Villanueva, V., Zúñiga, M. C., Swan, C. M., & Barmuta, L. A. (2012). Global patterns of stream detritivore distribution: implications for biodiversity loss in changing climates. Global Ecology and Biogeography, 21 (2), 134-141. [ Links ]
Brokaw, N., Zimmerman, J. K., Willig, M. R., Camilo, G. R., Covich, A. P., Crowl, T. A., Fetcher, N., Haines, B. L., Lodge, D. J., Lugo, A. E., Myster, R. W., Pringle, C. M., Sharpe, J. M., Scatena, F. N., Schowalter, T. D., Silver, W. L., Thompson, J., Vogt, D. J., Vogt, K. A., Waide, R. B., Walker, L. R., Woolbright, L. L., Wunderle, J. M., & Zou, X. (2012). Response to Disturbance. In: N. Brokaw, T. A. Crowl, A. E. Lugo, W. H. Mcdowell, F. N. Scatena, R. B. Waide & M. R. Willig (Eds.), A Caribbean Forest Tapestry: The Multidimensional Nature of Disturbance and Response (pp. 201-271). New York: Oxford University Press, Inc. [ Links ]
Campbell, I. C., & Fuchshuber, L. (1995). Polyphenols, condensed tannins, and processing rates of tropical and temperate leaves in an Australian stream. Journal of the North American Benthological Society, 14, 174-182. [ Links ]
Cheshire, K., Boyero, L., & Pearson, R. G. (2005). Food webs in tropical Australian streams: Shredders are not scarce. Freshwater Biology, 50, 748-769. [ Links ]
Coley, P. D., & Barone, J. A. (1996). Herbivory and plant defenses in tropical forests. Annual Review of Ecology and Systematics, 27, 305-335. [ Links ]
Coley, P. D., & Barone, J. A. (2001). Ecology of Defenses. In S. Levin (Ed.), Encyclopedia of Biodiversity (Vol. 2) (pp. 11-21). Amsterdam, The Netherlands: Academic Press. [ Links ]
Covich, A. P. (1988). Geographical and historical comparisons of Neotropical streams: Biotic diversity and detrital processing in highly variable habitats. Journal of the North American Benthological Society, 7, 361-386. [ Links ]
Covich, A. P., & McDowell, W. H. (1996). The stream community. In D. P. Reagan & R. B. Waide (Eds.), The food web of a tropical rain forest (pp. 433-459). Chicago: University of Chicago Press. [ Links ]
Covich, A. P., Palmer, M. A., & Crowl, T. A. (1999). Role of benthic invertebrate species in freshwater ecosystems. Bioscience, 49, 119-127. [ Links ]
Covich, A. P., Crowl, T. A., & Scatena, F. N. (2003). Effects of extreme low flows on freshwater shrimps in a perennial tropical stream. Freshwater Biology, 48, 1199-1206. [ Links ]
Covich, A. P., Austen, M. C., Barlocher, F., Chauvet, E., Cardinale, B. J., Biles, C. L., Inchausti, P., Dangles, O., Solan, M., Gessner, M. O., Statzner, B., Moss, B., & Moss, B. (2004). The role of biodiversity in the functioning of freshwater and marine benthic ecosystems. Bioscience, 54, 767-775. [ Links ]
Cross, W. F., Covich, A. P., Crowl, T. A., Benstead, J. P., & Ramirez, A. (2008). Secondary production, longevity, and consumption rates of freshwater shrimps in two tropical streams with contrasting geomorphology and foodweb structure. Freshwater Biology, 53, 2504-2519. [ Links ]
Crow, T. (1980). A rain forest chronicle: A 30 year record of change in structure and composition at El Verde, Puerto Rico. Biotropica, 12, 42-55. [ Links ]
Crowl, T. A., McDowell, W. H., Covich, A. P., & Johnson, S. L. (2001). Freshwater shrimp effects on detrital processing and localized nutrient dynamics in a montane, tropical rain forest stream. Ecology, 82, 775-783. [ Links ]
Crowl, T. A., Welsh, V., Heartsill-Scalley, T., & Covich, A. P. (2006). Effects of different types of conditioning on rates of leaf-litter shredding by Xiphocaris elongata, a Neotropical freshwater shrimp. Journal of the North American Benthological Society, 25, 196-206. [ Links ]
Dobson, M., Mathooko, J. M., Magana, A., & Ndegwa, F. K. (2002). Macroinvertebrate assemblages and detritus processing in Kenyan highland streams: More evidence for the paucity of shredders in the tropics? Freshwater Biology, 47, 909-919. [ Links ]
Dobson, M., Mathooko, J. M., Ndegwa, F. K., & M’erimba, C. (2003). Leaf litter processing rates in a Kenyan stream, the Njoro River. Hydrobiologia, 519, 207-210. [ Links ]
Dudgeon, D. (1994). The influence of riparian vegetation on macroinvertebrate community structure and functional organization in 6 New Guinea streams. Hydrobiologia, 292, 65-85. [ Links ]
Feio, M. J., & Graça, M. A. S. (2000). Food consumption by the larvae of Sericostoma vittatum (Trichoptera), and endemic species from the Iberian Peninsula. Hydrobiologia, 439, 7-11. [ Links ]
Garcia-Martino, A. R., Warner, G. S., Scatena, F. N., & Civco, D. L. (1996). Rainfall, runoff relationships in the Luquillo Mountains of Puerto Rico. Caribbean Journal of Science, 32, 413-424. [ Links ]
Gonçalves, J. F., Graça, M. A. S., & Callisto, M. (2006). Leaf-litter breakdown in 3 streams in temperate, Mediterranean and tropical Cerrado climates. Journal of the North American Benthological Society, 25, 344-355. [ Links ]
Graça, M. A. S. (2001). The role of invertebrates on leaf litter decomposition in streams – a review. International Review of Hydrobiology, 86, 383-393. [ Links ]
Graça, M. A. S., Cressa, C., Gessner, M. O., Feio, M. J., Callies, K. A., & Barrios, C. (2001). Food quality, feeding preferences, survival and growth of shredders from temperate and tropical streams. Freshwater Biology, 46, 947-957. [ Links ]
Heartsill, S. T., Crowl, T. A., & Thompson, J. (2009). Tree species distributions in relation to stream distance in a midmontane wet forest, Puerto Rico. Caribbean Journal of Science, 45, 52–63. [ Links ]
Irons, J. G. III, Oswood, M. W., Stout, R. J., & Pringle, C. M. (1994). Latitudinal patterns in leaf litter breakdown: is temperature really important? Freshwater Biology, 32, 401-411. [ Links ]
Jonsson, M., & Malmqvist, B. (2000). Ecosystem process rate increases with animal species richness: evidence from leaf-eating aquatic insects. Oikos, 89, 519-523. [ Links ]
Jonsson, M., Malmqvist, B., & Hoffsten, P. O. (2001). Leaf litter breakdown rates in boreal streams: Does shredder species richness matter? Freshwater Biology, 46, 161-171. [ Links ]
La Caro, F., & Rudd, R. L. (1985). Leaf litter disappearance rates in Puerto Rican montane rain forest. Journal of Tropical Ecology, 17, 269-276. [ Links ]
March, J. G., Benstead, J. P., Pringle, C. M., & Ruebel, M. W. (2001). Linking shrimp assemblages with rates of detrital processing along an elevational gradient in a tropical stream. Canadian Journal of Fisheries and Aquatic Sciences, 58, 470-478. [ Links ]
March, J. G., & Pringle, C. M. (2003). Food web structure and basal resource utilization along a tropical island stream continuum, Puerto Rico. Biotropica, 35, 84-93. [ Links ]
McDowell, W. H., Scatena, F. N., Waide, R. B., Brokaw, N., Camilo, G. R., Covich, A. P., Crowl, T. A., González, G., Greathouse, E. A., Klawinski, P., Lodge, D. J., Lugo, A. E., Pringle, C. M., Richardson, B. A., Richardson, M. J., Schaefer, D. A., Silver, W. L., Thompson, J., Vogt, D. J., Vogt, K. A., Willig, M. R., Woolbright, L. L., Zou, X., Zimmerman, J. K. (2012). Geographic and Ecological Setting of the Luquillo Mountains. In N. Brokaw, T. A. Crowl, A. E. Lugo, W. H. Mcdowell, F. N. Scatena, R. B. Waide & M. R. Willig (Eds.), A Caribbean Forest Tapestry: The Multidimensional Nature of Disturbance and Response (pp. 72-163). New York, NY: Oxford University Press, Inc. [ Links ]
Mathooko, J. M., M’Erimba, C. M., & Leichtfried, M. (2000). Decomposition of leaf litter of Dombeya goetzenii in the Njoro River, Kenya. Hydrobiologia, 418, 147-152. [ Links ]
Naiman, R. J., & Décamps, H. (1997). The ecology of interfaces: Riparian zones. Annual Review of Ecology and Systematics, 28, 621-658. [ Links ]
Padgett, D. E. (1976). Leaf decomposition by fungi in a tropical rainforest stream. Biotropica, 8, 166-178. [ Links ]
Petersen, R. C., & Cummins, K. W. (1974). Leaf processing in a woodland stream. Freshwater Biology, 4, 343-368. [ Links ]
Quinn, J. M., Smith, B. J., Burrell, G. P., & Parkyn, S. M. (2000). Leaf litter characteristics affect colonisation by stream invertebrates and growth of Olinga feredayi (Trichoptera: Conoesucidae). New Zealand Journal of Marine and Freshwater Research, 34, 273-287. [ Links ]
Ramirez, A., & Hernandez-Cruz, L. R. (2004). Aquatic insect assemblages in shrimp-dominated tropical streams, Puerto Rico. Biotropica, 36, 259-266. [ Links ]
Rincón, J. E., & Martínez, I. (2006). Food quality and feeding preferences of Phylloicus sp. (Trichoptera: Calamoceratidae). Journal of the North American Benthological Society, 25, 207-213. [ Links ]
Rosemond, A. D., Pringle, C. M., & Ramirez, A. (1998). Macroconsumer effects on insect detritivores and detritus processing in a tropical stream. Freshwater Biology, 39, 515-523. [ Links ]
Rosemond, A. D., Pringle, C. M., Ramirez, A., & Paul, M. (2001). A test of top-down and bottom-up control in a detritus-based food web. Ecology, 82, 2279-2293. [ Links ]
Sanchez, M. J., López, E., & Lugo, A. E. (1997). Chemical and physical analysis of selected plants and soils from Puerto Rico (1981-1990). Research Note IITF-RN-1. Ríos Piedras, Puerto Rico: U.S. Department of Agriculture, Forest Service, International Institute of Tropical Forestry. [ Links ]
Smith, R. F. (1970). The vegetation structure of a Puerto Rican rain forest before and after short-term gamma irradiation. In H. T. Odum & R. T. Pigeon (Eds.), A tropical rain forest (pp. H101-H104). Springfield, VA: USAEC, NTIS. [ Links ]
Stout, R. J. (1989). Effects of condensed tannins on leaf processing in mid-latitude and tropical streams: A theoretical approach. Canadian Journal of Fisheries and Aquatic Science, 46, 1097-1106. [ Links ]
Uriarte, M., Clark, J. S., Zimmerman, J. K., Comita, L. S., Forero-Montaña, J., & Thompson, J. (2012). Multidimensional trade-offs in species responses to disturbance: implications for diversity in a subtropical forest. Ecology, 93, 191–205. [ Links ]
Usio, N., & Townsend, C. R. (2001). The significance of the crayfish Paranephrops zealandicus as shredders in a New Zealand headwater stream. Journal of Crustacean Biology, 21, 354-359. [ Links ]
Vogt, K. A., Vogt, D. J., Boon, P., Covich, A., Scatena, F. N., Asbjornsen, H., O’Hara, J. L., Pérez, J., Siccama, T. G., Bloomfield, J., & Ranciato, J. F. (1996). Litter dynamics along stream, riparian and upslope areas following Hurricane Hugo, Luquillo Experimental Forest, Puerto Rico. Biotropica, 28, 458-470. [ Links ]
Wallace, J. B., Eggert, S. L., Meyer, J. L., & Webster, J. R. (1999). Effects of resource limitation in a detrital-based ecosystem. Ecological Monographs, 69, 409-442. [ Links ]
Wantzen, K. M., Wagner, R., Suetfeld, R., & Junk, W. J. (2002). How do plant-herbivore interactions of trees influence coarse detritus processing by shredders in aquatic ecosystems of different latitudes? Internationale Vereinigung fur Theoretische und Angewandte Limnologie, 28, 815-821. [ Links ]
Wantzen, K. M., & Wagner, R. (2006). Detritus processing by shredders: a tropical-temperate comparison. Journal of the North American Benthological Society, 25, 214–230. [ Links ]
Wantzen, K. M., Yule, C. M., Mathooko, J. M., & Pringle, C. M. (2008). Organic Matter Processing in Tropical Streams. In D. Dudgeon (Ed.), Tropical Stream Ecology (pp. 43-64). London: Academic Press. [ Links ]
Webster, J. R., & Benfield, E. F. (1986). Vascular plant breakdown in freshwater ecosystems. Annual Review of Ecology and Systematics, 17, 567-594. [ Links ]
Wright, M. S., & Covich, A. P. (2005). The effect of macroinvertebrate exclusion on leaf breakdown rates in a tropical headwater stream. Biotropica, 37, 403-408. [ Links ]
Zalamea, M., & González, G. (2008). Leaf fall phenology in a subtropical wet forest in Puerto Rico: from species to community patterns. Biotropica, 40, 295–304. [ Links ]
Zar, J. H. (1999). Biostatistical Analysis (4th ed.). New Jersey: Prentice Hall, Inc. [ Links ]
Zimmerman, J. K., Aide, T. M., Herrera, L., Rosario, M., Rosario, M., & Serrano, M. (1995). Effects of land management and a recent hurricane on forest structure and composition in the Luquillo Experimental Forest, Puerto Rico. Forest Ecology and Management, 78, 147-157. [ Links ]
Zimmerman, J. K. H., & Covich, A. P. (2003). Distribution of juvenile crabs (Epilobocera sinuatifrons) in two Puerto Rican headwater streams: Effects of pool morphology and past land-use legacies. Archiv für Hydrobiologie, 158, 343-357. [ Links ]
Zimmerman, J. K. H., & Covich, A. P. (2007). Damage and recovery of riparian sierra palms (Prestoea acuminata var. montana) after Hurricane Georges: Influence of topography, land use, and biotic characteristics. Biotropica, 39, 43-49. [ Links ]
Zou, X., Zucca, C. P., Waide, R. B., & McDowell, W. H. (1995). Long-term influence of deforestation on tree species composition and litter dynamics of a tropical rain forest in Puerto Rico. Forest Ecology and Management, 78, 147-157. [ Links ]
Ardon, M., Pringle, C.M., & Eggert, S.L. (2009). Does leaf chemistry differentially affect leaf breakdown in tropical versus temperate streams?: Importance of standardized analytical techniques to measure leaf chemistry. Journal of the North American Benthological Society, 28(2), 440-453 [ Links ]
Beard, K. H., Vogt, K. A., Vogt, D. J., Scatena, F. N., Covich, A. P., Siccama, T. C., Sigurdardottir, R., & Crowl, T. A. (2005). The response of structural and functional characteristics of a tropical forested ecosystem to multiple, natural disturbances in legacy environments. Ecological Monographs, 75, 345-361. [ Links ]
Benstead, J. P. (1996). Macroinvertebrates and the processing of leaf litter in a tropical stream. Biotropica, 28, 367-375. [ Links ]
Bobeldyk, A. M., & Ramírez, A. (2007). Leaf breakdown in a tropical headwater stream (Puerto Rico): The role of freshwater shrimps and detritivorous insects. Journal of Freshwater Ecology, 22, 581-590. [ Links ]
Boulton, A. J., & Boon, P. J. (1991). A review of methodology used to measure leaf litter decomposition in lotic environments: time to turn over a new leaf? Australian Journal of Marine and Freshwater Research, 42, 1-43. [ Links ]
Boulton, A. J., Boyero, L., Covich, A. P., Dobson, M., Lake, S., & Pearson, R. (2008) Are tropical streams ecologically different from temperate streams? In D. Dudgeon (Ed.), Tropical stream ecology (pp. 257-284). San Diego: Academic Press. [ Links ]
Boyero, L., Pearson, R. G., Dudgeon, D., Graca, M. A. S., Gessner, M. O., Albariño, R. J., Ferreira, V., Yule, C. M., Boulton, A. J., Arunachalam, M., Callisto, M., Chauvet, E., Ramirez, A., Chara, J., Moretti, M. S., Goncalves, J. F., Helson, J. E., Chara, J., Encalada, A. C., Davies, S. J., Lamothe, S., Cornejo, A., Li, Y., Buria, L. M., Villanueva, N., Zuniga, M. C., & Pringle, C. M. (2011). Global distribution of a key trophic guild contrasts with common latitudinal diversity patterns. Ecology, 92, 1839-1848. [ Links ]
Boyero, L., Richard, G., Pearson, R. G., Dudgeon, D., Ferreira, V., Graça, M. A. S., Gessner, M. O., Boulton, A. J., Chauvet, E., Yule, C. M., Albariño, R. J., Ramírez, A., Helson, J. E., Callisto, M., Arunachalam, M., Chará, J., Figueroa, R., Mathooko, J. M., Gonçalves, J. F., Moretti, M. S., Chará-Serna, A. M., Davies, J. N., Encalada, A., Lamothe, S., Buria, L. M., Castela, J., Cornejo, A., Li, A. O. Y., M’Erimba, C., Díaz Villanueva, V., Zúñiga, M. C., Swan, C. M., & Barmuta, L. A. (2012). Global patterns of stream detritivore distribution: implications for biodiversity loss in changing climates. Global Ecology and Biogeography, 21 (2), 134-141. [ Links ]
Brokaw, N., Zimmerman, J. K., Willig, M. R., Camilo, G. R., Covich, A. P., Crowl, T. A., Fetcher, N., Haines, B. L., Lodge, D. J., Lugo, A. E., Myster, R. W., Pringle, C. M., Sharpe, J. M., Scatena, F. N., Schowalter, T. D., Silver, W. L., Thompson, J., Vogt, D. J., Vogt, K. A., Waide, R. B., Walker, L. R., Woolbright, L. L., Wunderle, J. M., & Zou, X. (2012). Response to Disturbance. In: N. Brokaw, T. A. Crowl, A. E. Lugo, W. H. Mcdowell, F. N. Scatena, R. B. Waide & M. R. Willig (Eds.), A Caribbean Forest Tapestry: The Multidimensional Nature of Disturbance and Response (pp. 201-271). New York: Oxford University Press, Inc. [ Links ]
Campbell, I. C., & Fuchshuber, L. (1995). Polyphenols, condensed tannins, and processing rates of tropical and temperate leaves in an Australian stream. Journal of the North American Benthological Society, 14, 174-182. [ Links ]
Cheshire, K., Boyero, L., & Pearson, R. G. (2005). Food webs in tropical Australian streams: Shredders are not scarce. Freshwater Biology, 50, 748-769. [ Links ]
Coley, P. D., & Barone, J. A. (1996). Herbivory and plant defenses in tropical forests. Annual Review of Ecology and Systematics, 27, 305-335. [ Links ]
Coley, P. D., & Barone, J. A. (2001). Ecology of Defenses. In S. Levin (Ed.), Encyclopedia of Biodiversity (Vol. 2) (pp. 11-21). Amsterdam, The Netherlands: Academic Press. [ Links ]
Covich, A. P. (1988). Geographical and historical comparisons of Neotropical streams: Biotic diversity and detrital processing in highly variable habitats. Journal of the North American Benthological Society, 7, 361-386. [ Links ]
Covich, A. P., & McDowell, W. H. (1996). The stream community. In D. P. Reagan & R. B. Waide (Eds.), The food web of a tropical rain forest (pp. 433-459). Chicago: University of Chicago Press. [ Links ]
Covich, A. P., Palmer, M. A., & Crowl, T. A. (1999). Role of benthic invertebrate species in freshwater ecosystems. Bioscience, 49, 119-127. [ Links ]
Covich, A. P., Crowl, T. A., & Scatena, F. N. (2003). Effects of extreme low flows on freshwater shrimps in a perennial tropical stream. Freshwater Biology, 48, 1199-1206. [ Links ]
Covich, A. P., Austen, M. C., Barlocher, F., Chauvet, E., Cardinale, B. J., Biles, C. L., Inchausti, P., Dangles, O., Solan, M., Gessner, M. O., Statzner, B., Moss, B., & Moss, B. (2004). The role of biodiversity in the functioning of freshwater and marine benthic ecosystems. Bioscience, 54, 767-775. [ Links ]
Cross, W. F., Covich, A. P., Crowl, T. A., Benstead, J. P., & Ramirez, A. (2008). Secondary production, longevity, and consumption rates of freshwater shrimps in two tropical streams with contrasting geomorphology and foodweb structure. Freshwater Biology, 53, 2504-2519. [ Links ]
Crow, T. (1980). A rain forest chronicle: A 30 year record of change in structure and composition at El Verde, Puerto Rico. Biotropica, 12, 42-55. [ Links ]
Crowl, T. A., McDowell, W. H., Covich, A. P., & Johnson, S. L. (2001). Freshwater shrimp effects on detrital processing and localized nutrient dynamics in a montane, tropical rain forest stream. Ecology, 82, 775-783. [ Links ]
Crowl, T. A., Welsh, V., Heartsill-Scalley, T., & Covich, A. P. (2006). Effects of different types of conditioning on rates of leaf-litter shredding by Xiphocaris elongata, a Neotropical freshwater shrimp. Journal of the North American Benthological Society, 25, 196-206. [ Links ]
Dobson, M., Mathooko, J. M., Magana, A., & Ndegwa, F. K. (2002). Macroinvertebrate assemblages and detritus processing in Kenyan highland streams: More evidence for the paucity of shredders in the tropics? Freshwater Biology, 47, 909-919. [ Links ]
Dobson, M., Mathooko, J. M., Ndegwa, F. K., & M’erimba, C. (2003). Leaf litter processing rates in a Kenyan stream, the Njoro River. Hydrobiologia, 519, 207-210. [ Links ]
Dudgeon, D. (1994). The influence of riparian vegetation on macroinvertebrate community structure and functional organization in 6 New Guinea streams. Hydrobiologia, 292, 65-85. [ Links ]
Feio, M. J., & Graça, M. A. S. (2000). Food consumption by the larvae of Sericostoma vittatum (Trichoptera), and endemic species from the Iberian Peninsula. Hydrobiologia, 439, 7-11. [ Links ]
Garcia-Martino, A. R., Warner, G. S., Scatena, F. N., & Civco, D. L. (1996). Rainfall, runoff relationships in the Luquillo Mountains of Puerto Rico. Caribbean Journal of Science, 32, 413-424. [ Links ]
Gonçalves, J. F., Graça, M. A. S., & Callisto, M. (2006). Leaf-litter breakdown in 3 streams in temperate, Mediterranean and tropical Cerrado climates. Journal of the North American Benthological Society, 25, 344-355. [ Links ]
Graça, M. A. S. (2001). The role of invertebrates on leaf litter decomposition in streams – a review. International Review of Hydrobiology, 86, 383-393. [ Links ]
Graça, M. A. S., Cressa, C., Gessner, M. O., Feio, M. J., Callies, K. A., & Barrios, C. (2001). Food quality, feeding preferences, survival and growth of shredders from temperate and tropical streams. Freshwater Biology, 46, 947-957. [ Links ]
Heartsill, S. T., Crowl, T. A., & Thompson, J. (2009). Tree species distributions in relation to stream distance in a midmontane wet forest, Puerto Rico. Caribbean Journal of Science, 45, 52–63. [ Links ]
Irons, J. G. III, Oswood, M. W., Stout, R. J., & Pringle, C. M. (1994). Latitudinal patterns in leaf litter breakdown: is temperature really important? Freshwater Biology, 32, 401-411. [ Links ]
Jonsson, M., & Malmqvist, B. (2000). Ecosystem process rate increases with animal species richness: evidence from leaf-eating aquatic insects. Oikos, 89, 519-523. [ Links ]
Jonsson, M., Malmqvist, B., & Hoffsten, P. O. (2001). Leaf litter breakdown rates in boreal streams: Does shredder species richness matter? Freshwater Biology, 46, 161-171. [ Links ]
La Caro, F., & Rudd, R. L. (1985). Leaf litter disappearance rates in Puerto Rican montane rain forest. Journal of Tropical Ecology, 17, 269-276. [ Links ]
March, J. G., Benstead, J. P., Pringle, C. M., & Ruebel, M. W. (2001). Linking shrimp assemblages with rates of detrital processing along an elevational gradient in a tropical stream. Canadian Journal of Fisheries and Aquatic Sciences, 58, 470-478. [ Links ]
March, J. G., & Pringle, C. M. (2003). Food web structure and basal resource utilization along a tropical island stream continuum, Puerto Rico. Biotropica, 35, 84-93. [ Links ]
McDowell, W. H., Scatena, F. N., Waide, R. B., Brokaw, N., Camilo, G. R., Covich, A. P., Crowl, T. A., González, G., Greathouse, E. A., Klawinski, P., Lodge, D. J., Lugo, A. E., Pringle, C. M., Richardson, B. A., Richardson, M. J., Schaefer, D. A., Silver, W. L., Thompson, J., Vogt, D. J., Vogt, K. A., Willig, M. R., Woolbright, L. L., Zou, X., Zimmerman, J. K. (2012). Geographic and Ecological Setting of the Luquillo Mountains. In N. Brokaw, T. A. Crowl, A. E. Lugo, W. H. Mcdowell, F. N. Scatena, R. B. Waide & M. R. Willig (Eds.), A Caribbean Forest Tapestry: The Multidimensional Nature of Disturbance and Response (pp. 72-163). New York, NY: Oxford University Press, Inc. [ Links ]
Mathooko, J. M., M’Erimba, C. M., & Leichtfried, M. (2000). Decomposition of leaf litter of Dombeya goetzenii in the Njoro River, Kenya. Hydrobiologia, 418, 147-152. [ Links ]
Naiman, R. J., & Décamps, H. (1997). The ecology of interfaces: Riparian zones. Annual Review of Ecology and Systematics, 28, 621-658. [ Links ]
Padgett, D. E. (1976). Leaf decomposition by fungi in a tropical rainforest stream. Biotropica, 8, 166-178. [ Links ]
Petersen, R. C., & Cummins, K. W. (1974). Leaf processing in a woodland stream. Freshwater Biology, 4, 343-368. [ Links ]
Quinn, J. M., Smith, B. J., Burrell, G. P., & Parkyn, S. M. (2000). Leaf litter characteristics affect colonisation by stream invertebrates and growth of Olinga feredayi (Trichoptera: Conoesucidae). New Zealand Journal of Marine and Freshwater Research, 34, 273-287. [ Links ]
Ramirez, A., & Hernandez-Cruz, L. R. (2004). Aquatic insect assemblages in shrimp-dominated tropical streams, Puerto Rico. Biotropica, 36, 259-266. [ Links ]
Rincón, J. E., & Martínez, I. (2006). Food quality and feeding preferences of Phylloicus sp. (Trichoptera: Calamoceratidae). Journal of the North American Benthological Society, 25, 207-213. [ Links ]
Rosemond, A. D., Pringle, C. M., & Ramirez, A. (1998). Macroconsumer effects on insect detritivores and detritus processing in a tropical stream. Freshwater Biology, 39, 515-523. [ Links ]
Rosemond, A. D., Pringle, C. M., Ramirez, A., & Paul, M. (2001). A test of top-down and bottom-up control in a detritus-based food web. Ecology, 82, 2279-2293. [ Links ]
Sanchez, M. J., López, E., & Lugo, A. E. (1997). Chemical and physical analysis of selected plants and soils from Puerto Rico (1981-1990). Research Note IITF-RN-1. Ríos Piedras, Puerto Rico: U.S. Department of Agriculture, Forest Service, International Institute of Tropical Forestry. [ Links ]
Smith, R. F. (1970). The vegetation structure of a Puerto Rican rain forest before and after short-term gamma irradiation. In H. T. Odum & R. T. Pigeon (Eds.), A tropical rain forest (pp. H101-H104). Springfield, VA: USAEC, NTIS. [ Links ]
Stout, R. J. (1989). Effects of condensed tannins on leaf processing in mid-latitude and tropical streams: A theoretical approach. Canadian Journal of Fisheries and Aquatic Science, 46, 1097-1106. [ Links ]
Uriarte, M., Clark, J. S., Zimmerman, J. K., Comita, L. S., Forero-Montaña, J., & Thompson, J. (2012). Multidimensional trade-offs in species responses to disturbance: implications for diversity in a subtropical forest. Ecology, 93, 191–205. [ Links ]
Usio, N., & Townsend, C. R. (2001). The significance of the crayfish Paranephrops zealandicus as shredders in a New Zealand headwater stream. Journal of Crustacean Biology, 21, 354-359. [ Links ]
Vogt, K. A., Vogt, D. J., Boon, P., Covich, A., Scatena, F. N., Asbjornsen, H., O’Hara, J. L., Pérez, J., Siccama, T. G., Bloomfield, J., & Ranciato, J. F. (1996). Litter dynamics along stream, riparian and upslope areas following Hurricane Hugo, Luquillo Experimental Forest, Puerto Rico. Biotropica, 28, 458-470. [ Links ]
Wallace, J. B., Eggert, S. L., Meyer, J. L., & Webster, J. R. (1999). Effects of resource limitation in a detrital-based ecosystem. Ecological Monographs, 69, 409-442. [ Links ]
Wantzen, K. M., Wagner, R., Suetfeld, R., & Junk, W. J. (2002). How do plant-herbivore interactions of trees influence coarse detritus processing by shredders in aquatic ecosystems of different latitudes? Internationale Vereinigung fur Theoretische und Angewandte Limnologie, 28, 815-821. [ Links ]
Wantzen, K. M., & Wagner, R. (2006). Detritus processing by shredders: a tropical-temperate comparison. Journal of the North American Benthological Society, 25, 214–230. [ Links ]
Wantzen, K. M., Yule, C. M., Mathooko, J. M., & Pringle, C. M. (2008). Organic Matter Processing in Tropical Streams. In D. Dudgeon (Ed.), Tropical Stream Ecology (pp. 43-64). London: Academic Press. [ Links ]
Webster, J. R., & Benfield, E. F. (1986). Vascular plant breakdown in freshwater ecosystems. Annual Review of Ecology and Systematics, 17, 567-594. [ Links ]
Wright, M. S., & Covich, A. P. (2005). The effect of macroinvertebrate exclusion on leaf breakdown rates in a tropical headwater stream. Biotropica, 37, 403-408. [ Links ]
Zalamea, M., & González, G. (2008). Leaf fall phenology in a subtropical wet forest in Puerto Rico: from species to community patterns. Biotropica, 40, 295–304. [ Links ]
Zar, J. H. (1999). Biostatistical Analysis (4th ed.). New Jersey: Prentice Hall, Inc. [ Links ]
Zimmerman, J. K., Aide, T. M., Herrera, L., Rosario, M., Rosario, M., & Serrano, M. (1995). Effects of land management and a recent hurricane on forest structure and composition in the Luquillo Experimental Forest, Puerto Rico. Forest Ecology and Management, 78, 147-157. [ Links ]
Zimmerman, J. K. H., & Covich, A. P. (2003). Distribution of juvenile crabs (Epilobocera sinuatifrons) in two Puerto Rican headwater streams: Effects of pool morphology and past land-use legacies. Archiv für Hydrobiologie, 158, 343-357. [ Links ]
Zimmerman, J. K. H., & Covich, A. P. (2007). Damage and recovery of riparian sierra palms (Prestoea acuminata var. montana) after Hurricane Georges: Influence of topography, land use, and biotic characteristics. Biotropica, 39, 43-49. [ Links ]
Zou, X., Zucca, C. P., Waide, R. B., & McDowell, W. H. (1995). Long-term influence of deforestation on tree species composition and litter dynamics of a tropical rain forest in Puerto Rico. Forest Ecology and Management, 78, 147-157. [ Links ]
1. Departamento de Biología. Facultad de Ciencias, Universidad del Zulia. Apartado Postal 4001, Maracaibo, Venezuela; jerincon04@gmail.com
2. Institute of Ecology, Odum School of Ecology, University of Georgia, Athens, GA 30602-2202 USA; alanc@uga.edu
Received 12-XII-2013. Corrected 20-I-2014. Accepted 13-II-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}