










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.1 San José Feb. 2014
Densidad y estructura poblacional de Cebus capucinus curtus (Primates: Cebidae) y Bradypus variegatus gorgon (Pilosa: Bradypodidae), en Isla Gorgona, Colombia
Density and structure of populations of Cebus capucinus curtus (Primates: Cebidae) and Bradypus variegatus gorgon (Pilosa: Bradypodidae) in Gorgona Island, Colombia
Density and structure of populations of Cebus capucinus curtus (Primates: Cebidae) and Bradypus variegatus gorgon (Pilosa: Bradypodidae) in Gorgona Island, Colombia
Mario Fernando Garcés-Restrepo1*, Pamela Carvajal-Nieto1, Jurany Milena Astorquiza1,2*, Elizabeth Sánchez-Vélez1 & Alan Giraldo1
Abstract
Gorgona is a continental island in the Pacific of Colombia. Two subspecies of arboreal mammals inhabit the island: the White-throated Capuchin (Cebus capucinus curtus) and the Brown-throated Sloth (Bradypus variegatus gorgon). These organisms are important for conservation since they are endemic and perform an essential role as seed dispersers in the National Park. This paper presents information on the population ecology of these two subspecies by using the transects-of-variable-width method, based on perpendicular detection distance, to establish density and age structure. The density of C. capucinus curtus in Gorgona Island was 170.6ind/km2 (95% CI=122-238.4), while that of B. variegatus gorgon was 2.6ind/km2 (IC 95%= 1.3 to 4.9ind/km2). The density of C. capucinus curtus in Isla Gorgona is the highest recorded for this species throughout its geographic range, while that of B. variegatus gorgon is the lowest reported for the species. The high density of C. curtus capucinus could be related to a synergistic effect between low predation and continuous availability of food, while the low density of B. variegates gorgon could be related to past hunting pressure, to the low reproductive rate of the species, and to a pandemic occurred in 2005. We recommend constant monitoring of populations of both subspecies, and population health studies for B. variegatus gorgon. Rev. Biol. Trop. 62 (Suppl. 1): 447-456. Epub 2014 February 01.
Key words: Population density, Island effect, key resources, Bradypus variegatus gorgon, Cebus capucinus curtus, Gorgona.
Resumen
En Isla Gorgona se registran dos subespecies endémicas de mamíferos arbóreos, el Mono capuchino de pecho blanco (Cebus capucinus curtus) y el Perezoso de tres dedos de garganta marrón (Bradypus variegatus gorgon), especies importante para la conservación debido a su carácter endémico y papel ecológico como dispersores de semillas en el PNN Gorgona. En este trabajo se presenta información sobre la ecología poblacional de estas dos subespecies, utilizando el método de muestreo por distancia con transectos lineales para establecer la densidad, además se describió la estructura etaria general de cada población con base en los muestreos y observaciones directas. La densidad de C. capucinus curtus en isla Gorgona fue de 170,6 ind/km2 (IC 95%=122,0-238,4 ind/km2) mientras que para B. variegatus gorgon fue de 2,6 ind/km2 (IC 95%=1,3-4,9 ind/km2). El registro de densidad de C. capucinus curtus en isla Gorgona es el más alto para la especie en todo su rango de distribución geográfica, mientras que el de B. variegatus gorgon es el más bajo reportado para la especie. La alta densidad de C. capucinus curtus estaría relacionada con un efecto sinérgico entre la baja depredación natural y la continua disponibilidad de alimento, mientras que la baja densidad de B. variegatus gorgon estaría relacionada con la presión de caza realizada en el pasado, la baja tasa reproductiva de la especie y una pandemia ocurrida en el año 2005. Se recomienda el monitoreo constante de las poblaciones y estudios de salud poblacional para B. variegatus gorgon.
Palabras clave: Densidad poblacional, efecto isla, recursos claves, Cebus capucinus curtus, Bradypus variegatus gorgon, Gorgona, Colombia.
Isla Gorgona, es reconocida en el Pacífico Oriental Tropical (POT) principalmente por albergar una alta variedad de ambientes marino costeros (Barrios & López-Victoria, 2001; Giraldo & Valencia, 2012), sin embargo en el ambiente terrestre se desarrolla una frondosa selva pluvial tropical que presenta asociaciones vegetales únicas (Aguirre & Rangel, 1990). A nivel faunístico, esta localidad presenta una baja riqueza de especies de vertebrados, como resultado del “efecto isla” (MacArthur & Wilson, 1967; Lack, 1976; Williamson, 1981), no obstante contiene una alta tasa de endemismo (Von Prahl, Guhl & Grogl, 1979; Alberico, 1986) a causa del aislamiento reproductivo y a las presiones selectivas diferenciales que se desarrollan en estos ambientes insulares (Case, 1978; Lomolino, 1986; Boback, 2003).
El ecosistema terrestre de isla Gorgona ha sido alterado por actividad humana durante más de 300 años. Inicialmente sirvió como sitio de abastecimiento y descanso para las embarcaciones que navegaron por la ruta del Pacífico desde Panamá hacia Perú, posteriormente se desarrollaron actividades agrícolas y ganaderas (De Robledo, 1997), y finalmente se estableció una prisión, que durante 25 años, utilizó de manera intensa los recursos naturales disponibles en esta localidad, alterando la estructura y composición de cerca del 80% de los bosques naturales y probablemente llevando a la extinción de una subespecie endémica de guatín, Dasyprocta punctata pandora (Alberico, 1986; 1987; Torres, 1986; Aguirre & Rangel, 1990; De Robledo, 1997; Díaz, López-Victoria & Barrios, 2001a; Giraldo & Valencia, 2012). Sin embargo, después de la declaratoria de isla Gorgona como Parque Nacional en 1984, la mayoría de las zonas intervenidas han tenido un proceso de restauración natural de alrededor de tres décadas (UAESPNN, 1998).
Entre las especies de vertebrados terrestres que habitan en isla Gorgona, el Mono capuchino de pecho blanco (Cebus capucinus curtus, Bangs, 1905) y el Perezoso de tres dedos de garganta marrón (Bradypus variegatus gorgon, Thomas, 1926), se consideran subespecies endémicas. Aunque existe mucha discusión acerca de la validez de estos taxa, debido a la contaminación genética por parte de individuos llevados desde el continente y la falta de análisis genéticos y morfológicos apropiados; sin embargo las últimas revisiones taxonómicas realizadas por Defler (2004) y Haysen (2009), reconocen su validez. En este contexto, las poblaciones residentes en la isla, representan la totalidad de estos linajes únicos, por lo que su conservación dependerá únicamente de las acciones que se realicen en este lugar.
En un contexto ecológico, el mono cariblanco y el perezoso de tres dedos de garganta marrón podrían estar cumpliendo un papel clave en procesos de dispersión activa y pasiva de semillas de algunas especies vegetales en la isla Gorgona (Howe & Smallwood, 1982; Estrada & Fleming, 1986; Aguirre & Rangel, 1990; Defler, 2004). Además, estas especies generalmente son un enlace efectivo para el flujo de la energía entre las diferentes secciones verticales del bosque (Clark, Poulsen & Parker, 2001; Carvajal-Nieto, Giraldo & Plese, 2013). Sin embargo, la magnitud de la presión humana por transformación del hábitat y cacería (Alberico, 1986; 1987; Valencia et al., 2012) durante el funcionamiento de la prisión, pueden haber generado algún efecto sobre estas poblaciones.
El propósito de este trabajo fue establecer el estado de las poblaciones de C. capucinus curtus y B. variegatus gorgon en isla Gorgona con base en la cuantificación de su densidad y descripción de su estructura etaria, generando una línea base que permita a mediano plazo identificar tendencias y evaluar la efectividad de las medidas de manejo que puedan llegar a ser implementadas. Adicionalmente, se evaluó el grado de variación de la densidad poblacional entre la zona primitiva y la zona de recuperación natural de isla Gorgona, zonas que fueron definidas por la unidad de Parques Nacionales Naturales con base en el grado de transformación del hábitat terrestre (UAESPNN, 2010).
Materiales y métodos
Área de estudio: El Parque Nacional Natural Gorgona es un área protegida representada por 61 680ha y un territorio insular que corresponde a 1 568.4ha. Se localiza en el Océano Pacífico al suroccidente colombiano a 35km del lugar más cercano del continente que corresponde a Punta Reyes, en la localidad de Bazán (Díaz et al., 2001a). La principal porción emergida, isla Gorgona, tiene un área de 1 333.3ha de las cuales el 85% presentan una topografía escarpada con una pendiente entre 50-75%, y el 15% son planas con pendiente que no sobrepasan el 12% (Chamorro, 1990), siendo el punto más alto sobre el nivel del mar el cerro La Trinidad, con 338m (Díaz, Pinzón, Perdomo, Barrios & López-Victoria, 2001b).
Se registran dos máximos de precipitación a lo largo del año: de mayo a junio, y de septiembre a octubre, con un periodo de menor precipitación entre enero y abril. La precipitación anual supera los 6 500mm de lluvia. Las temperaturas medias anuales son de 26.0ºC, con una desviación máxima de 1.4º, y la humedad relativa promedio de 85.2%, enmarcando la localidad dentro de un clima súper húmedo tropical (Rangel & Rudas, 1990; Blanco, 2009).
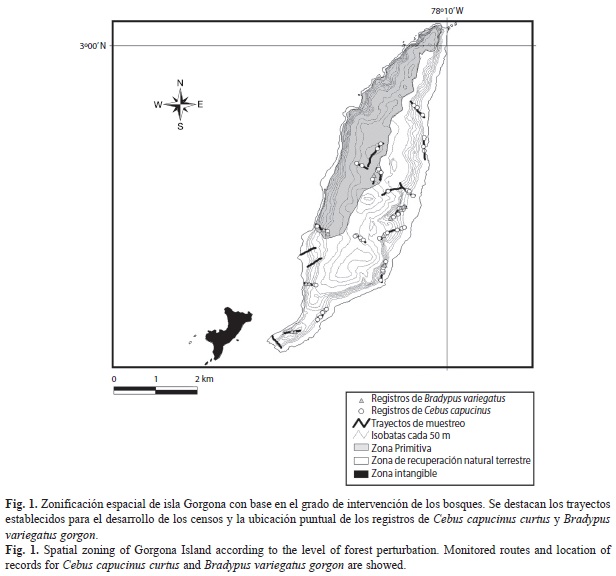
La vegetación del sistema insular de Gorgona hace parte de la provincia del Chocó biogeográfico, con una distribución de franjas de vegetación generadas por efecto de la intervención humana. Se identifican tres franjas principales: una franja a lo largo de los bordes de playa altamente intervenida, una franja intermedia con vegetación sucesional en diferentes estados de desarrollo y como respuesta a diferentes grados de intervención, y una franja interior de vegetación nativa con bajo grado de intervención y en mejor estado de conservación (Rangel & Rivera-Díaz, 2004) (Fig. 1). Para el desarrollo de este trabajo se consideró la franja interior como la zona de bosque primitivo, y la franja costera e intermedia como la zona de bosque en recuperación natural (UAESPNN, 1998).
Métodos: Debido a la topografía del terreno, se dispusieron cinco trayectos lineales de ancho y longitud variable (Bibby, Burguess, Hill & Mustoe, 2000, Buckland, Anderson, Burnham & Laake, 2001) en la zona primitiva y 14 trayectos lineales en la zona de recuperación natural terrestre. Cada trayecto fue recorrido por dos observadores a una velocidad constante de 0.5km/h, equipados con binoculares, entre octubre 2010 y julio 2011 en intervalos de dos a tres días. Todas las observaciones se realizaron entre las 06:00 y las 18:00h. Cada observación de C. capucinus curtus y B. variegatus gorgon fue tabulada, registrando la hora del encuentro, el número de individuos observados, la distancia perpendicular hasta el centroide del grupo o el individuo al trayecto, el sexo, edad (adulto, joven o cría) y la actividad realizada.
Para establecer el tamaño y variación de los grupos de C. capucinus curtus se realizaron seguimientos intensivos de una hora a grupos detectados en las zonas de muestreo (Defler & Pintor, 1985; Pruetz & Leasor, 2002). Adicionalmente, durante los seguimientos se estableció la estructura de los grupos considerando el número de hembras, machos, adultos, sub adultos y crías presentes. La estructura poblacional para B. variegatus gorgon se estableció con base en los registros realizados durante los censos, complementados por observaciones fortuitas, durante los desplazamientos entre trayectos de muestreo.
Análisis estadístico: La densidad poblacional de C. capucinus curtus y B. variegatus gorgon se estableció utilizando el programa Distance 6.0 (Thomas et al., 2010). Las densidades fueron determinadas teniendo en cuenta la distribución de distancias perpendiculares que mejor se ajusta al modelo teórico, escogiendo la combinación de la función clave y la serie de expansión con el valor más bajo en el criterio de información de Akaike (Buckland et al., 2001). Como en los muestreos en trayectos es poco probable observar los grupos completos de primates debido a las acciones evasivas de los individuos (Defler & Pintor, 1985; Pruetz & Leasor, 2002), la densidad de C. capucinus curtus fue calculada introduciendo como constantes el promedio del tamaño de grupo estimado en los seguimientos a las tropas, el error estándar y los grados de libertad, de tal manera que se integró la variación del tamaño de grupo en el estimado de la variación de la densidad poblacional.
Debido a la compleja topografía del área, no se pudo acceder a muchas zonas de la isla, por lo tanto los estimativos corresponden a valores asociados al área de muestreo y no se extrapolan a la totalidad de la isla. Considerando el bajo número de registros de B. variegatus gorgon en la zona de estudio, el cálculo de la variación de la densidad se asumió ajustando los registros a una distribución de Poisson (Buckland et al., 1993). Se compararon las densidades estimadas de C. capucinus curtus y B. variegatus gorgon entre la zona primitiva y la zona de recuperación natural, comparando los promedios de la densidad y su desviación estándar a través de una prueba T para parámetros (Zar, 1999) utilizando el programa PAST 2.16 (Hammer, Harper & Ryan, 2001).
Resultados
Se recorrió una distancia total de 134.8km en los trayectos: 106.8km en zona de recuperación natural terrestre y 28.0km en la zona primitiva. Se registraron 412 individuos de C. capucinus curtus, en 62 eventos de detección (Fig. 1). Se realizó el seguimiento a 14 grupos, y se calculó un promedio de 15 individuos por tropa (s=1.09). La densidad estimada para esta especie en la zona de estudio fue de 170.6ind/km2 (IC 95%=122,0-238.4ind/km2), siendo significativamente más alta (T=3.49, p=0.002) en la zona de recuperación natural terrestre (D=184.0 ind/km2, IC 95%=123.3-274.8 ind/km2) que en la zona primitiva (D=122.4ind/km2, IC 95%=75,6-198,2ind/km2) (Cuadro 1).
Para el mismo esfuerzo de recorrido se registraron 14 individuos solitarios de B. variegatus gorgon. La densidad estimada para esta especie en la zona de estudio fue de 2.6ind/km2 (IC 95%=1.3-4.9ind/km2), sin detectarse diferencias estadísticamente significativas (T=0.433, p=0.192) para la densidad entre la zona de recuperación natural terrestre (D=2.6 ind/km2; IC 95%=1.2-5.3ind/km2) y la zona primitiva (D=2.3ind/km2, IC 95%=0.6-9.2ind/km2) (Tabla 1).
Los grupos de C. capucinus curtus en isla Gorgona estuvieron conformados principalmente por jóvenes (59%), seguido de hembras adultas (17%), machos adultos (13%) y crías (10%), estableciéndose una proporción machos:hembras de 1.3:1 y de hembras adultas:inmaduros de 1.0:4.0. Todas las observaciones de B. variegatus gorgon realizadas durante el desarrollo de los muestreos fueron de individuos adultos, siendo la proporción machos:hembras de 2:1.
Discusión
Mono capuchino de pecho blanco (Cebus capucinus curtus): Con base en los registros bibliográficos sobre densidad de C. capucinus curtus en su rango de distribución geográfica, la densidad estimada para isla Gorgona corresponde al registro más alto para esta especie. En otras localidades los estimados de densidad se encuentran entre 5 y 60ind/km2 (Fedigan, Fedigan & Chapman, 1985; Robinson & Janson, 1987; Oppenheimer, 1990; Pruetz & Leasor, 2002; Roncancio & Gómez-Posada, 2009) en ambientes continentales y entre 18 y 24ind/km2 (Robinson & Janson, 1987) en ambientes insulares (Cuadro 2).
La alta densidad registrada para C. capucinus curtus en la isla Gorgona puede estar explicada en gran medida por la disminución de competencia interespecífica y depredadores, como consecuencia de ser una localidad insular (MacArthur & Wilson, 1967; Nilsson, Bjorkman, Forslund & Hoglund, 1985; George, 1987; Tilman, 1994). Es notario en esta localidad la ausencia de depredadores naturales, con excepción de la Boa común (Boa constrictor) y la Babilla (Caiman cocrodilus) que podrían tener un efecto de control sobre neonatos y juveniles. El efecto de la depredación como agente modulador de la densidad de vertebrados ha sido ampliamente estudiado (Krebs, 1995; Korpimaki & Krebs, 1996). En términos generales la depredación tiene un efecto directo sobre la población al incrementar la tasa de mortalidad, pero también tiene un efecto indirecto al afectar el comportamiento alimentario y reproductivo de los individuos (Lima & Dill, 1990; Ylönen, 1994; Korpimaki & Krebs, 1996).
Por otra parte la presencia de plantas introducidas, como la palma africana (Elaeis guineensis), podrían estar dando un aporte constante de recursos alimentarios que permitiría un crecimiento constante de las poblaciones (Stevenson & Quiñones, 1993; Williams & Vaughan, 2001; Hemingway & Bynum, 2005). Este fenómeno ha sido registrado en otras especies de vertebrados como la Pava andina (Penelope perspicax) (Ríos, Londoño & Muñoz, 2005), y para C. capucinus curtus en isla Gorgona el patrón de agregación hacia el sector centro-oriental de la isla, en donde se encuentran presentes estas palmas, estaría sugiriendo este tipo de relación. Sin embargo, es algo que debe ser puesto a prueba en campo.
Considerando la viabilidad de la relación entre la palma africana y el mono cariblanco, es altamente probable que las diferencias en la densidad de esta especie entre el bosque primitivo y el bosque en recuperación natural, sea una consecuencia de la mayor disponibilidad espacial y temporal de recursos alimentarios en las zonas en recuperación en donde se encuentran plantas exóticas, como es el sector del Poblado. Cuando el hábitat ofrece una mayor cantidad de frutos, los grupos de C. capucinus recorren menores distancias ya que suplen rápidamente sus requerimientos energéticos, teniendo ámbitos hogareños más pequeños. Al estar más agregados, los grupos tenderán a ser de mayor tamaño, incrementando consecuentemente su densidad (Dumbar, 1988; Bicca-Marques, 2003).
En localidades continentales e insulares de Centro América, los grupos de C. capucinus están conformados principalmente por hembras adultas (30%-35%) (Robinson & Janson, 1987; Pruetz & Leasor, 2002), mientras que en la isla Gorgona los grupos estuvieron conformados principalmente por jóvenes (59%). Los machos de esta especie presentan una maduración sexual tardía con relación a las hembras (Robinson & Janson, 1987), razón por la cual los grupos tienden a estar dominados por hembras. Sin embargo, la dominancia de jóvenes en los grupos de isla Gorgona sugiere que la tasa de mortalidad natural de neonatos y jóvenes en esta localidad puede ser significativamente menor al esperado en ambientes continentales.
Perezoso de tres dedos de cuello marrón (Bradipus variegatus gorgon): De acuerdo con información presencial documentada por los funcionarios de Parques Nacionales y la estación científica Henry von Prahl del PNN Gorgona, durante el año 2005 se presentó una súbita mortandad de B. variegatus gorgon. Posterior a esta pandemia, Cuevas-Correa (2007) había reportado que la densidad de esta especie era de entre 8.8 y 10.0ind/km2 en esta localidad. Sin embargo, de acuerdo con los resultados que se reportan en el presente trabajo, la densidad poblacional de B. variegatus gorgon en esta localidad se ha reducido en un 70%.
La reducción de la población de B. variegatus gorgon en un poco más de cinco años, sumado a la baja tasa reproductiva de esta especie (Britton, 1941; Soares & Carneiro, 2002), pueden fácilmente conducir a esta población a una inviabilidad temporal por efecto de “cuello de botella” poblacional (Mills, 2013). Adicionalmente, la estructura sexual de la población podría incrementar dicho efecto. En poblaciones continentales de esta especie la proporción de sexos reportada es de 1:1 (Meritt, 1976; Montgomery & Sunquist, 1975; Acevedo-Qu & Sánchez, 2007), mientras que en la población de B. variegatus gorgon en isla Gorgona, la proporción de sexos es de 2:1. Aunque esta desproporción sexual puede ser una característica inherente a esta población, es altamente probable que bajo estas condiciones la tasa de encuentro entre parentales (macho-hembra) se reduzca significativamente, lo que afectaría drásticamente la tasa reproductiva de la población. Sin embargo, es importante mencionar que incluso la densidad estimada por Cuevas-Correa (2007) es comparativamente baja con relación a los reportes de densidad realizados en otras localidades (Cuadro 2), por lo que un efecto residual por la presión de uso sobre esta especie durante la época de la prisión, no puede ser descartado (Alberico, 1987; Valencia et al., 2012).
En términos generales, las poblaciones de B. variegatus se ajustan adecuadamente a hábitats simplificados (Anderson, Rowcliffe & Cowlishaw, 2007) como los de ambientes en recuperación, ya que sus recursos alimentarios tienden a distribuirse de manera uniforme (Chiarello, 1999). Además, la presencia de cursos de agua dulce es quizás una de las características del hábitat que más influyen en su patrón de distribución espacial (Losada, 2007). En este sentido, las condiciones ambientales en isla Gorgona y las estrategias de manejo que han sido implementadas para el ambiente continental en esta área protegida (UAESPNN, 1998), podrían reducir la variación de la densidad B. variegatus gorgon entre la zona primitiva y la zona de recuperación natural terrestre.
Recomendaciones generales: La estimación de la densidad de individuos en poblaciones naturales es altamente variable, ya que dependen del tipo de bosque en el que se realiza el monitoreo, el método de censo que se utilice y el estimador de densidad seleccionado (Taube, Vie, Fournier, Gentry & Duplantier, 1999), por lo tanto las comparaciones y conclusiones que se pueden realizar con base a este estudio presentan limitaciones. Sin embargo, este trabajo es el primer esfuerzo sistemático para evaluar la densidad poblacional de C. capucinus curtus y B. variegatus gorgon utilizando métodos estandarizados de estimación poblacional, lo cual sirve como línea básica para evaluar tendencias en el futuro. De igual manera estos resultados plantean nuevas preguntas de investigación que deben ser atendidas para fortalecer las medidas de manejo implementadas en esta área natural protegida. Por ejemplo evaluar el impacto de una población grande de C. capucinus curtus sobre sus presas potenciales, que incluso pueden llegar a ser especies endémicas de esta localidad como el Largato Azul (Anolis gorgon); así como evaluar si el estado poblacional de B. variegatus gorgon está asociado con la salud de los individuos (veterinario) o con la salud de la población (genético). De igual manera, los esfuerzos que se generen para la conservación de estos taxa deben tener como antesala verificar la validez de la condición de “subespecie endémica”. Para responder a estas preguntas es necesario establecer un programa de monitoreo de las poblaciones, que no solo genere información sobre el tamaño, sino que permita recopilar información específica sobre la historia natural de estos taxones, complementadas con métodos actuales de investigación taxonómica, como las herramientas moleculares.
Agradecimientos
A Andrés Quintero Ángel, Rodrigo Lozano Osorio, Julio Herrera, José Omar Ortiz, Fabio Sarria, David Velásquez, Juan Pablo Ospina por su colaboración durante las jornadas de campo. A los funcionarios del PNN Gorgona, en especial a los funcionarios de la Estación Científica Henry von Prahl, Luis Fernando Payan y Héctor Chirimía por su decidido apoyo para el desarrollo de esta investigación. De igual manera agradecemos a Oscar Murillo, Vladimir Rojas y Néstor Roncancio por sus valiosos aportes y recomendaciones en la versión definitiva del documento. Este trabajo fue cofinanciado por la Universidad del Valle, Conservación Internacional Colombia, el Fondo Para la Acción Ambiental y la Niñez, la Fundación SQUALUS y el PNN Gorgona, y estuvo amparado por el permiso de investigación PIBD DTSO 011-10.
Referencias
Abreu, C., Siqueira, R. & Moura, J. (2001). Nota sobre uma amostra populacional de preguicas Bradypus variegatus Schnz, 1825, na lha de Itamaracá, Pernambuco, Brasil. Revista Nordestina de Biología, 15: 31-41. [ Links ]
Acevedo-Qu, J. F. & Sánchez, D. P. (2007). Abundancia y preferencia de hábitat de Bradypus variegatus y Choloepus hoffmanni, durante la época seca en Arboletes, Antioqui (Trabajo de Grado). Universidad de Antioquia, Medellín, Colombia. [ Links ]
Aguirre, J. & Rangel, O. (1990). Biota y ecosistemas de Gorgona. Fondo FEN, Bogotá, Colombia. [ Links ]
Alberico, M. S. (1986). Los mamíferos. In H. von Prahl & M. Alberico (Eds.), Isla de Gorgona (pp. 191-209). Biblioteca Textos Universitarios, Banco Popular, Bogotá, Colombia. [ Links ]
Alberico, M. S. (1987). Los mamíferos. In J. I. Borrero (Ed.), Gorgona (pp. 69-78). Fundación Mejor Ambiente, Cali, Colombia. [ Links ]
Anderson, J., Rowcliffe, M. & Cowlishaw, G. (2007). Does the matrix matter? A forest primate in a complex agricultural landscape. Biological Conservation, 135: 212-222. [ Links ]
Bangs, O. (1905). The vertebrata of Gorgona Island. Bulletin of the Museum of Comparative Zoology at Harvard, 46: 89-91. [ Links ]
Barrios, L. M. & López-Victoria, M. (eds.) 2001. Gorgona marina: contribución al conocimiento de una isla única. INVEMAR, Serie Publicaciones Especiales, No. 7, Santa Marta, Colombia. [ Links ]
Bibby, C. J., Burguess, N. D., Hill, D. A. & Mustoe, S. (2000). Bird census techniques. Academic Press, London, United Kingdom. [ Links ]
Bicca-Marques, J. C. (2003). How do howler monkeys cope with habitat fragmentation?. In L. K. Marsh (Ed.), Primates in Fragment: ecology and conservation (pp. 283-308). Springer, London, United Kingdom. [ Links ]
Blanco, J. F. (2009). The hydroclimatology of Gorgona Island: seasonal and ENSO-related patterns. Actualidades Biológicas, 31: 111-121. [ Links ]
Boback, S. M. (2003). Body size evolution in snakes: evidence from island populations. COPEIA, 2003: 81-94. [ Links ]
Britton, S. W. (1941). Form and function in the sloth. The Quarterly Review of Biology, 16: 13-34. [ Links ]
Buckland, S. T., Anderson, D. R., Burnham, K. P. & Laake, J. L. (1993). Distance sampling. Estimating abundance of biological populations. Chapman & Hall, London, United Kingdom. [ Links ]
Buckland, S. T., Anderson, D. R., Burnham, K. P., Laake, J. L., Borchers, D. L. & Thomas, L. (2001). Introduction to distance sampling: Estimating abundance of biological populations. Oxford University Press. Oxford, United Kingdom. [ Links ]
Carvajal-Nieto, P., Giraldo, A. & Plese, T. (2013). Densidad poblacional y algunos aspectos de uso de hábitat del perezoso de tres dedos (Bradypus variegatus) en un ambiente insular del chocó biogeográfico colombiano. Boletín Científico Museo de Historia Natural, 17(1):101-110. [ Links ]
Case, T. J. (1978). A general explanation for insular body size trends in terrestrial vertebrates. Ecology, 59: 1-18. [ Links ]
Chamorro, C. (1990). Suelos. In J. Aguirre & J. O. Rangel (Eds.), Biota y ecosistemas de Gorgona (pp. 65-72). Fondo FEN, Bogotá, Colombia. [ Links ]
Chiarello, A. G. (1999). Effects of fragmentation of the Atlantic forest on mammal communities in south-eastern Brazil. Biological Conservation, 89: 71-82. [ Links ]
Clark, C. J., Poulsen, J. R. & Parker, V. T. (2001). The role of arboreal seed dispersal groups on the seed rain of a lowland tropical forest. Biotropica, 33: 606-620. [ Links ]
Cuevas-Correa, D. (2007). Estimación de la densidad poblacional, patrón de distribución espacial y preferencia de hábitat del perezoso de tres dedos Bradypus variegatus gorgon y consideraciones para su conservación en el Parque Nacional Natural Gorgona (Tesis de Grado). Pontificia Universidad Javeriana, Bogotá, Colombia. [ Links ]
De Robledo, C. (1997). Gorgona Isla Prisión. Crónicas. Pijao Editores, Santa Fe de Bogotá, Colombia. [ Links ]
Defler, T. R. & Pintor, D. (1985). Censuring primates by transect in a forest of known primate density. International Journal of Primatology, 6: 243-259. [ Links ]
Defler, T. R. (2004). Primates of Colombia. Conservation International, Washington D.C., USA. [ Links ]
Díaz, M., López-Victoria, M. & Barrios, L. M. (2001a). Introducción. In L. M. Barrios & M. López-Victoria (Eds.), Gorgona Marina: Contribución al reconocimiento de una isla única (pp. 13-16). INVEMAR, Serie Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
Díaz, M., Pinzón, J. H., Perdomo, A. M., Barrios, L. B. & López-Victoria, M. (2001b). Generalidades. In L. M. Barrios & M. López-Victoria (Eds.), Gorgona Marina: Contribución al reconocimiento de una isla única (pp. 17-26). INVEMAR, Serie Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
Dumbar, R. I. (1988). Primate social systems. Chapman & Hall, London, United Kingdom. [ Links ]
Estrada, A. & Fleming, T. H. (1986). Frugivores and seed dispersal. Junk Publishers, Dordrecht, South Holland, Netherlands. [ Links ]
Fedigan, L. M., Fedigan, L. & Chapman, C. (1985). A census of Alouatta palliata and Cebus capucinus monkeys in Santa Rosa Nacional Park, Costa Rica. Brenesia, 23: 309-322. [ Links ]
Freese, C. H. (1976). Cebus capucinus (mono cara blanca, white-faced capuchin). In D. H. Janzen (Ed.), Costa Rican natural history (pp. 458-460). University of Chicago Press, Chicago, United States. [ Links ]
George, T. L. (1987). Greater land bird densities on island vs. mainland: Relation to nest predation level. Ecology, 68: 1393-1400. [ Links ]
Giraldo, A. & Valencia, B. (2012) (Eds.) Isla Gorgona: Paraíso de Biodiversidad y Ciencia. Programa Editorial de la Universidad Del Valle, Cali, Colombia. [ Links ]
Hammer O, Harper, D. & Ryan, P. (2001). PAST: paleontological statistics software for education and data analysis. Palaeontología Electrónica, 4: 1-9. [ Links ]
Haysen, V. (2009). Bradypus variegatus (Pilosa: Bradypodidae). Mammalian Species, 42: 19-32. [ Links ]
Hemingway, C. A. & Bynum, N. (2005). The influence of seasonality on primate diet and ranging. In D. Brockman & C. P. van Schaik (Eds.), Seasonality in primates: studies of living and extinct human and Non-human primates (pp. 57-104). Cambridge University Press, Cambridge, United Kingdom. [ Links ]
Howe, H. F. & Smallwood, J. (1982). Ecology of Seed Dispersal. Annual Review of Ecology, Evolution and Systematics, 13: 201-228. [ Links ]
Korpimaki, E. & Krebs, C. J. (1996). Predation and population cycles on small mammals. Bioscience, 46: 754-764. [ Links ]
Krebs, J. (1995). Two paradigms of population regulation. Wildlife Research, 22: 1-10. [ Links ]
Lack, D. L. (1976). Island biology: Illustrated by the land birds of Jamaica. Blackwell Scientific, Oxford, United Kingdom. [ Links ]
Lima, S. L. & Dill, L. M. (1990). Behavioral decisions made under the risk of predation: a review and prospectus. Canadian Journal of Zoology, 68: 619-640. [ Links ]
Lomolino, M. V. (1986). Mammalian community structure on islands: Immigration, extinction and interactive effects. Biological Journal of the Linnean Society, 28: 1-21. [ Links ]
Losada, T. (2007). Conocimiento local y uso sobre los perezosos por la comunidad indígena Jusy Monilla Amena, Amazonas, Colombia (Tesis de pregrado). Pontificia Javeriana, Bogotá, Colombia. [ Links ]
MacArthur, R. H. & Wilson, E. O. (1967). The theory of island biogeography. Princeton University Press, Princeton, New Jersey, USA. [ Links ]
Meritt, D. A. Jr. (1976). Sex ratios of Hoffmann’s Sloth, Choloepus hoffmanni Petters and Three-toed Sloth, Bradypus infuscatus Wagler in Panama. American Midland Naturalist, 96: 472-473. [ Links ]
Mills, L. S. (2013). Conservation of wildlife population: Demography genetics and management. Wiley-Blackwell, Oxford, United Kingdom. [ Links ]
Montgomery, G. G. & Sunquist, M. E. (1975). Impact of sloths on Neotropical energy flow and nutrient cycling. In E. Medina & F. Golly (Eds.), Trends Tropical Ecology; Ecological Studies IV (pp, 69-98). Springer Verlag, New York, USA. [ Links ]
Nilsson, S. G., Bjorkman, C., Forslund, P. & Hoglund, J. (1985). Egg predation in forest bird communities on islands and mainland. Oecología, 66: 511-515. [ Links ]
Oppenheimer, J. R. (1990). Cebus capucinus: ámbito doméstico, dinámica de población y relaciones interespecíficas. In E. G. Leigh, A. S. Rand & D. M. Windsor (Eds.), Ecología de un bosque tropical: ciclos estacionales y cambios a largo plazo (pp. 337-356). Smithsonian Institution Press, Washington D.C., USA. [ Links ]
Pruetz, L. & Leasor, J. (2002). Survey of three primate species in forest fragments at La Suerte Biological Field Station, Costa Rica. Neotropical Primates, 10: 4-8. [ Links ]
Rangel, J. O. & Rudas, A. (1990). Macroclima de Gorgona y de la región costera aledaña. In J. Aguirre & J. O. Rangel (Eds.), Biota y ecosistemas de Gorgona (pp. 13-41). Fondo FEN, Bogotá, Colombia. [ Links ]
Rangel, J. O. & Rivera-Díaz, O. (2004). Diversidad y riqueza de espermatófitos en el Chocó biogeográfico. In J. O. Rangel (Ed.), Colombia Diversidad Biótica IV: El Chocó Biogeográfico/Costa Pacífica (pp. 83-104). Instituto de Ciencias Naturales, Bogotá, Colombia. [ Links ]
Ríos, M. M., Londoño, G. A. & Muñoz, M. C. (2005). Densidad poblacional e historia natural de la Pava Negra (Aburria aburri) en los Andes centrales de Colombia. Ornitología Neotropical, 16: 205-217. [ Links ]
Robinson, J. G. & Janson, C. H. (1987). Capuchins, squirrel monkeys and atelines: socioecological convergence with old world primates. In B. Smuts, D. Cheney, R. Seyfarth, R. Wrangham & T. Struhsaker (Eds.), Primate Societies (pp. 69-82). The University of Chicago press, Chicago, USA. [ Links ]
Roncancio, N., & Gómez-Posada, C. (2009). Sympatric Alouatta seniculus and cebus capucinus in an Andean forest fragment in Colombia: a survey of population density. Neotropical Primates, 16: 51-56. [ Links ]
Soares, C. A. & Carneiro, R. S. (2002). Social behavior between mothers and young of sloths Bradypus variegatus Schinz, 1825 (Xenarthra: Bradypodidae). Brazilian Journal of Biology, 62: 249-252. [ Links ]
Stevenson, P. & Quiñones, M. (1993). Vertical stratification of four new world primates at Tinigua national park, Colombia La Macarena, Colombia. Field Studies of New World Monkeys, 8: 11-18. [ Links ]
Taube, E., Vie. J. C., Fournier, P., Gentry, C. & Duplantier, J. M. (1999). Distribution of two sympatric species of sloths (Choloepus didactylus and Bradypus tridactylus) along the sinnamary river, French Guiana. Biotropica, 31: 686-691. [ Links ]
Thomas, O. (1926). On mammals from Gorgona island with description of a new sloth. Annals and Magazine of Natural History, 9: 309-311. [ Links ]
Thomas, L., Buckland, S. T., Rexstad, E. A., Laake, J. L., Strindberg, S., Hedley, S. L., Bishop, J. R. B., Marques, T. A. & Burnham, K. P. (2010). Distance software: design and analysis of distance sampling surveys for estimating population size. Journal of Applied Ecology, 47: 5-14. doi: 10.1111/j.1365-2664.2009.01737.x [ Links ]
Tilman, D. (1994). Competition and biodiversity in spatially structured habitats. Ecology, 75: 2-16. [ Links ]
Torres, E. (1986). Historia de Gorgona. In H. von Prahl & M. Alberico (Eds.), Isla de Gorgona (pp. 9-17). Biblioteca Textos Universitarios, Banco Popular, Bogotá, Colombia. [ Links ]
UAESPNN. (2010). El sistema de Parques Nacionales Naturales de Colombia. Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales, Ministerio del Medio Ambiente. Editorial Nomos, Bogotá, Colombia. [ Links ]
Valencia, A., Villaquiran, D., Castro, F., Murillo, O., Bedoya, J., Carvajal, P., Astorquiza, M., Garcés-Restrepo, M. F., Yusti, A. P., Velandia, J., Sánchez, M. & Giraldo, A. (2012). Vertebrados del ambiente terrestre del Parque Nacional Natural Gorgona. In A. Giraldo & B. Valencia (Eds.), Isla Gorgona: Paraíso de Biodiversidad y Ciencia (pp. 193-221). Programa Editorial de la Universidad Del Valle, Cali, Colombia. [ Links ]
von Prahl, H., Guhl, F. & Grogl, M. (1979). Gorgona. Grupo Futura Editorial: Universidad de los Andes, Bogotá, Colombia. [ Links ]
Williams, H. E. & Vaughan, C. (2001). White-faced monkey (Cebus capucinus) ecology and management in neotropical agricultural landscapes during the dry season. Revista de Biología Tropical, 49(3-4): 1199-1206. [ Links ]
Williamson, M. (1981). Island populations. Oxford University Press, Oxford, United Kingdom. [ Links ]
Ylönen, H. (1994). Vole cycles and antidepredatory behavior. Trends in Ecology & Evolution, 9: 426-430. [ Links ]
Zar, J. H. (1999). Biostatistical Analysis. Prentice Hall, New Jersey, USA. [ Links ]
Acevedo-Qu, J. F. & Sánchez, D. P. (2007). Abundancia y preferencia de hábitat de Bradypus variegatus y Choloepus hoffmanni, durante la época seca en Arboletes, Antioqui (Trabajo de Grado). Universidad de Antioquia, Medellín, Colombia. [ Links ]
Aguirre, J. & Rangel, O. (1990). Biota y ecosistemas de Gorgona. Fondo FEN, Bogotá, Colombia. [ Links ]
Alberico, M. S. (1986). Los mamíferos. In H. von Prahl & M. Alberico (Eds.), Isla de Gorgona (pp. 191-209). Biblioteca Textos Universitarios, Banco Popular, Bogotá, Colombia. [ Links ]
Alberico, M. S. (1987). Los mamíferos. In J. I. Borrero (Ed.), Gorgona (pp. 69-78). Fundación Mejor Ambiente, Cali, Colombia. [ Links ]
Anderson, J., Rowcliffe, M. & Cowlishaw, G. (2007). Does the matrix matter? A forest primate in a complex agricultural landscape. Biological Conservation, 135: 212-222. [ Links ]
Bangs, O. (1905). The vertebrata of Gorgona Island. Bulletin of the Museum of Comparative Zoology at Harvard, 46: 89-91. [ Links ]
Barrios, L. M. & López-Victoria, M. (eds.) 2001. Gorgona marina: contribución al conocimiento de una isla única. INVEMAR, Serie Publicaciones Especiales, No. 7, Santa Marta, Colombia. [ Links ]
Bibby, C. J., Burguess, N. D., Hill, D. A. & Mustoe, S. (2000). Bird census techniques. Academic Press, London, United Kingdom. [ Links ]
Bicca-Marques, J. C. (2003). How do howler monkeys cope with habitat fragmentation?. In L. K. Marsh (Ed.), Primates in Fragment: ecology and conservation (pp. 283-308). Springer, London, United Kingdom. [ Links ]
Blanco, J. F. (2009). The hydroclimatology of Gorgona Island: seasonal and ENSO-related patterns. Actualidades Biológicas, 31: 111-121. [ Links ]
Boback, S. M. (2003). Body size evolution in snakes: evidence from island populations. COPEIA, 2003: 81-94. [ Links ]
Britton, S. W. (1941). Form and function in the sloth. The Quarterly Review of Biology, 16: 13-34. [ Links ]
Buckland, S. T., Anderson, D. R., Burnham, K. P. & Laake, J. L. (1993). Distance sampling. Estimating abundance of biological populations. Chapman & Hall, London, United Kingdom. [ Links ]
Buckland, S. T., Anderson, D. R., Burnham, K. P., Laake, J. L., Borchers, D. L. & Thomas, L. (2001). Introduction to distance sampling: Estimating abundance of biological populations. Oxford University Press. Oxford, United Kingdom. [ Links ]
Carvajal-Nieto, P., Giraldo, A. & Plese, T. (2013). Densidad poblacional y algunos aspectos de uso de hábitat del perezoso de tres dedos (Bradypus variegatus) en un ambiente insular del chocó biogeográfico colombiano. Boletín Científico Museo de Historia Natural, 17(1):101-110. [ Links ]
Case, T. J. (1978). A general explanation for insular body size trends in terrestrial vertebrates. Ecology, 59: 1-18. [ Links ]
Chamorro, C. (1990). Suelos. In J. Aguirre & J. O. Rangel (Eds.), Biota y ecosistemas de Gorgona (pp. 65-72). Fondo FEN, Bogotá, Colombia. [ Links ]
Chiarello, A. G. (1999). Effects of fragmentation of the Atlantic forest on mammal communities in south-eastern Brazil. Biological Conservation, 89: 71-82. [ Links ]
Clark, C. J., Poulsen, J. R. & Parker, V. T. (2001). The role of arboreal seed dispersal groups on the seed rain of a lowland tropical forest. Biotropica, 33: 606-620. [ Links ]
Cuevas-Correa, D. (2007). Estimación de la densidad poblacional, patrón de distribución espacial y preferencia de hábitat del perezoso de tres dedos Bradypus variegatus gorgon y consideraciones para su conservación en el Parque Nacional Natural Gorgona (Tesis de Grado). Pontificia Universidad Javeriana, Bogotá, Colombia. [ Links ]
De Robledo, C. (1997). Gorgona Isla Prisión. Crónicas. Pijao Editores, Santa Fe de Bogotá, Colombia. [ Links ]
Defler, T. R. & Pintor, D. (1985). Censuring primates by transect in a forest of known primate density. International Journal of Primatology, 6: 243-259. [ Links ]
Defler, T. R. (2004). Primates of Colombia. Conservation International, Washington D.C., USA. [ Links ]
Díaz, M., López-Victoria, M. & Barrios, L. M. (2001a). Introducción. In L. M. Barrios & M. López-Victoria (Eds.), Gorgona Marina: Contribución al reconocimiento de una isla única (pp. 13-16). INVEMAR, Serie Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
Díaz, M., Pinzón, J. H., Perdomo, A. M., Barrios, L. B. & López-Victoria, M. (2001b). Generalidades. In L. M. Barrios & M. López-Victoria (Eds.), Gorgona Marina: Contribución al reconocimiento de una isla única (pp. 17-26). INVEMAR, Serie Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
Dumbar, R. I. (1988). Primate social systems. Chapman & Hall, London, United Kingdom. [ Links ]
Estrada, A. & Fleming, T. H. (1986). Frugivores and seed dispersal. Junk Publishers, Dordrecht, South Holland, Netherlands. [ Links ]
Fedigan, L. M., Fedigan, L. & Chapman, C. (1985). A census of Alouatta palliata and Cebus capucinus monkeys in Santa Rosa Nacional Park, Costa Rica. Brenesia, 23: 309-322. [ Links ]
Freese, C. H. (1976). Cebus capucinus (mono cara blanca, white-faced capuchin). In D. H. Janzen (Ed.), Costa Rican natural history (pp. 458-460). University of Chicago Press, Chicago, United States. [ Links ]
George, T. L. (1987). Greater land bird densities on island vs. mainland: Relation to nest predation level. Ecology, 68: 1393-1400. [ Links ]
Giraldo, A. & Valencia, B. (2012) (Eds.) Isla Gorgona: Paraíso de Biodiversidad y Ciencia. Programa Editorial de la Universidad Del Valle, Cali, Colombia. [ Links ]
Hammer O, Harper, D. & Ryan, P. (2001). PAST: paleontological statistics software for education and data analysis. Palaeontología Electrónica, 4: 1-9. [ Links ]
Haysen, V. (2009). Bradypus variegatus (Pilosa: Bradypodidae). Mammalian Species, 42: 19-32. [ Links ]
Hemingway, C. A. & Bynum, N. (2005). The influence of seasonality on primate diet and ranging. In D. Brockman & C. P. van Schaik (Eds.), Seasonality in primates: studies of living and extinct human and Non-human primates (pp. 57-104). Cambridge University Press, Cambridge, United Kingdom. [ Links ]
Howe, H. F. & Smallwood, J. (1982). Ecology of Seed Dispersal. Annual Review of Ecology, Evolution and Systematics, 13: 201-228. [ Links ]
Korpimaki, E. & Krebs, C. J. (1996). Predation and population cycles on small mammals. Bioscience, 46: 754-764. [ Links ]
Krebs, J. (1995). Two paradigms of population regulation. Wildlife Research, 22: 1-10. [ Links ]
Lack, D. L. (1976). Island biology: Illustrated by the land birds of Jamaica. Blackwell Scientific, Oxford, United Kingdom. [ Links ]
Lima, S. L. & Dill, L. M. (1990). Behavioral decisions made under the risk of predation: a review and prospectus. Canadian Journal of Zoology, 68: 619-640. [ Links ]
Lomolino, M. V. (1986). Mammalian community structure on islands: Immigration, extinction and interactive effects. Biological Journal of the Linnean Society, 28: 1-21. [ Links ]
Losada, T. (2007). Conocimiento local y uso sobre los perezosos por la comunidad indígena Jusy Monilla Amena, Amazonas, Colombia (Tesis de pregrado). Pontificia Javeriana, Bogotá, Colombia. [ Links ]
MacArthur, R. H. & Wilson, E. O. (1967). The theory of island biogeography. Princeton University Press, Princeton, New Jersey, USA. [ Links ]
Meritt, D. A. Jr. (1976). Sex ratios of Hoffmann’s Sloth, Choloepus hoffmanni Petters and Three-toed Sloth, Bradypus infuscatus Wagler in Panama. American Midland Naturalist, 96: 472-473. [ Links ]
Mills, L. S. (2013). Conservation of wildlife population: Demography genetics and management. Wiley-Blackwell, Oxford, United Kingdom. [ Links ]
Montgomery, G. G. & Sunquist, M. E. (1975). Impact of sloths on Neotropical energy flow and nutrient cycling. In E. Medina & F. Golly (Eds.), Trends Tropical Ecology; Ecological Studies IV (pp, 69-98). Springer Verlag, New York, USA. [ Links ]
Nilsson, S. G., Bjorkman, C., Forslund, P. & Hoglund, J. (1985). Egg predation in forest bird communities on islands and mainland. Oecología, 66: 511-515. [ Links ]
Oppenheimer, J. R. (1990). Cebus capucinus: ámbito doméstico, dinámica de población y relaciones interespecíficas. In E. G. Leigh, A. S. Rand & D. M. Windsor (Eds.), Ecología de un bosque tropical: ciclos estacionales y cambios a largo plazo (pp. 337-356). Smithsonian Institution Press, Washington D.C., USA. [ Links ]
Pruetz, L. & Leasor, J. (2002). Survey of three primate species in forest fragments at La Suerte Biological Field Station, Costa Rica. Neotropical Primates, 10: 4-8. [ Links ]
Rangel, J. O. & Rudas, A. (1990). Macroclima de Gorgona y de la región costera aledaña. In J. Aguirre & J. O. Rangel (Eds.), Biota y ecosistemas de Gorgona (pp. 13-41). Fondo FEN, Bogotá, Colombia. [ Links ]
Rangel, J. O. & Rivera-Díaz, O. (2004). Diversidad y riqueza de espermatófitos en el Chocó biogeográfico. In J. O. Rangel (Ed.), Colombia Diversidad Biótica IV: El Chocó Biogeográfico/Costa Pacífica (pp. 83-104). Instituto de Ciencias Naturales, Bogotá, Colombia. [ Links ]
Ríos, M. M., Londoño, G. A. & Muñoz, M. C. (2005). Densidad poblacional e historia natural de la Pava Negra (Aburria aburri) en los Andes centrales de Colombia. Ornitología Neotropical, 16: 205-217. [ Links ]
Robinson, J. G. & Janson, C. H. (1987). Capuchins, squirrel monkeys and atelines: socioecological convergence with old world primates. In B. Smuts, D. Cheney, R. Seyfarth, R. Wrangham & T. Struhsaker (Eds.), Primate Societies (pp. 69-82). The University of Chicago press, Chicago, USA. [ Links ]
Roncancio, N., & Gómez-Posada, C. (2009). Sympatric Alouatta seniculus and cebus capucinus in an Andean forest fragment in Colombia: a survey of population density. Neotropical Primates, 16: 51-56. [ Links ]
Soares, C. A. & Carneiro, R. S. (2002). Social behavior between mothers and young of sloths Bradypus variegatus Schinz, 1825 (Xenarthra: Bradypodidae). Brazilian Journal of Biology, 62: 249-252. [ Links ]
Stevenson, P. & Quiñones, M. (1993). Vertical stratification of four new world primates at Tinigua national park, Colombia La Macarena, Colombia. Field Studies of New World Monkeys, 8: 11-18. [ Links ]
Taube, E., Vie. J. C., Fournier, P., Gentry, C. & Duplantier, J. M. (1999). Distribution of two sympatric species of sloths (Choloepus didactylus and Bradypus tridactylus) along the sinnamary river, French Guiana. Biotropica, 31: 686-691. [ Links ]
Thomas, O. (1926). On mammals from Gorgona island with description of a new sloth. Annals and Magazine of Natural History, 9: 309-311. [ Links ]
Thomas, L., Buckland, S. T., Rexstad, E. A., Laake, J. L., Strindberg, S., Hedley, S. L., Bishop, J. R. B., Marques, T. A. & Burnham, K. P. (2010). Distance software: design and analysis of distance sampling surveys for estimating population size. Journal of Applied Ecology, 47: 5-14. doi: 10.1111/j.1365-2664.2009.01737.x [ Links ]
Tilman, D. (1994). Competition and biodiversity in spatially structured habitats. Ecology, 75: 2-16. [ Links ]
Torres, E. (1986). Historia de Gorgona. In H. von Prahl & M. Alberico (Eds.), Isla de Gorgona (pp. 9-17). Biblioteca Textos Universitarios, Banco Popular, Bogotá, Colombia. [ Links ]
UAESPNN. (2010). El sistema de Parques Nacionales Naturales de Colombia. Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales, Ministerio del Medio Ambiente. Editorial Nomos, Bogotá, Colombia. [ Links ]
Valencia, A., Villaquiran, D., Castro, F., Murillo, O., Bedoya, J., Carvajal, P., Astorquiza, M., Garcés-Restrepo, M. F., Yusti, A. P., Velandia, J., Sánchez, M. & Giraldo, A. (2012). Vertebrados del ambiente terrestre del Parque Nacional Natural Gorgona. In A. Giraldo & B. Valencia (Eds.), Isla Gorgona: Paraíso de Biodiversidad y Ciencia (pp. 193-221). Programa Editorial de la Universidad Del Valle, Cali, Colombia. [ Links ]
von Prahl, H., Guhl, F. & Grogl, M. (1979). Gorgona. Grupo Futura Editorial: Universidad de los Andes, Bogotá, Colombia. [ Links ]
Williams, H. E. & Vaughan, C. (2001). White-faced monkey (Cebus capucinus) ecology and management in neotropical agricultural landscapes during the dry season. Revista de Biología Tropical, 49(3-4): 1199-1206. [ Links ]
Williamson, M. (1981). Island populations. Oxford University Press, Oxford, United Kingdom. [ Links ]
Ylönen, H. (1994). Vole cycles and antidepredatory behavior. Trends in Ecology & Evolution, 9: 426-430. [ Links ]
Zar, J. H. (1999). Biostatistical Analysis. Prentice Hall, New Jersey, USA. [ Links ]
1. Universidad del Valle, Facultad de Ciencias Naturales y Exactas, Departamento de Biología. Calle 13 No 100-00, Cali, Colombia. ecologia@univalle.edu.co
2. Universidad de Nariño, Departamento de Biología; milenaastorquiza@gmail.com
Recibido 18-X-2013. Corregido 20-XI-2013. Aceptado 19-XII-2013.


{kind=link}
{kind=link}
{kind=link}