










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.1 San José Feb. 2014
Tidepool fish assemblages of Gorgona Island, Colombian Pacific coast: a local and regional comparison
Abstract
The ecology of tidepool fishes has been extensively investigated worldwide over the past 40 years. As in many other ecological fields, studies in temperate zones outnumber studies carried out in tropical areas. Here, we document the short-term spatio-temporal variation of a tidepool fish assemblage from the Tropical Eastern Pacific (Gorgona Island, Colombia), compare our data with previous surveys made in 1993 and provide the first latitudinal comparison of tidepool fish assemblages in the Eastern Pacific coast (43°N to 36°S). During April-July 2006, monthly samplings were carried in 18 tidepools located at three sites this Island. A relatively species-rich tidepool fish assemblage (53 species) was found at Gorgona Island. Labrisomidae, Gobiidae and Gobiesocidae were the dominant resident components of this assemblage, whereas Pomacentridae, Muraenidae and Labridae constituted the transient component. Although the same dominant species were observed in surveys from 1993 and 2006, suggesting a high persistence of this assemblage over time, clear differences in the relative abundance of some species were also detected. These differences were partly the result of using different sampling methodologies (rotenone vs clove oil). Comparisons with other studies performed in tropical and temperate areas of the Eastern Pacific coast indicate a clear latitudinal pattern in the composition of these assemblages that correspond to well-documented biogeographic subdivisions of the marine fauna in this region. Further investigation of the ecological role that tidepool fishes play in rocky intertidal and subtidal areas of Gorgona Island and the whole Tropical Eastern Pacific region will benefit our understanding of the functioning of these important coastal ecosystems. Rev. Biol. Trop. 62 (Suppl. 1): 373-390. Epub 2014 February 01.
Key words: tidepool icthyofauna, community structure, zoogeography, Colombia, Eastern Pacific Ocean.
Resumen
A nivel mundial, la ecología de peces de charcos intermareales ha sido ampliamente investigada durante los últimos 40 años, especialmente en zonas templadas. En este trabajo, se documenta la variación espacio-temporal en corta escala de un ensamblaje de peces de charcos intermareales en el Pacífico Oriental Tropical (Isla Gorgona, Colombia), y se comparan nuestros resultados, con los resultados obtenidos durante muestreos realizados en la misma localidad en 1993. Además se realiza la primera comparación latitudinal de estos ensamblajes para la costa oriental del Océano Pacífico (43°N a 36°S). Durante abril a junio de 2006, se realizaron muestreos mensuales en 18 charcos intermareales distribuidos en tres localidades de Isla Gorgona, estableciendo un ensamblaje de peces de charcos intermareales con una alta riqueza de especies (53 especies). Labrisomidae, Gobiidae and Gobiesocidae fueron los componentes residentes dominantes en el ensamblaje, mientras que Pomacentridae, Muraenidae and Labridae fueron los componentes transitorios. Aunque el registro de las mismas especies dominantes en 1993 y 2006 sustenta la idea de una alta persistencia temporal de este ensamblaje, se detectaron diferencias significativas en la abundancia relativa de las especies. Estas diferencias podrían ser el resultado de las diferentes técnicas de captura utilizadas durante 1993 y 2006 (rotenona vs aceite de clavo). Al comparar los estudios sobre peces intermareales realizados en la zona tropical y zona templada de la costa oriental del Océano Pacífico, se identificó un claro patrón latitudinal en la composición de los ensamblajes, que es consecuente con la ampliamente documentada subdivisión biogeográfica de la fauna marina de esta región. Para comprender el funcionamiento de los ecosistemas costeros del Pacífico Oriental tropical, es necesario que futuros esfuerzos de investigación se encaminen a conocer el papel ecológico de los peces intermareales en esta región.
Palabras clave: ictiofauna intermareal, estructura de comunidad, zoogeografía, Colombia, Océano Pacífico Oriental.
Rocky shorelines constitute a predominant coastal seascape of the world, harbouring often very diverse biological communities. Organisms living in these zones are subject to environmental changes induced by tidal and day-night cycles (Little & Kitching, 1996; Palmer, 2000). To cope with these changing, often stressing, conditions, sessile and mobile organisms respond differently. Fishes, for example, can stay in intertidal zones when favourable conditions are present (i.e. at high tide) returning to subtidal habitats at low tide. A subset of coastal fishes can be specially adapted to remain in the intertidal zone even at low tide in tidepools (Gibson, 2003). Such difference in behaviour highlights the important role that fish can play in transporting energy within intertidal and between subtidal habitats.
Tidepools are common features within intertidal rocky shores, providing the opportunity to a large number of organisms to escape from harsh conditions on emerged rocks (Metaxas & Scheibling, 1993; Martins, Hawkins, Thompson & Jenkins, 2007). Tidepools are often considered as nursery grounds for fish and are also used as small isolated systems where organisms and their environmental conditions can be easily manipulated to advance the ecological understanding of these marine systems (Nielsen, 2001).
Patterns of spatio-temporal variation in tidepool fish assemblages have been studied for the last 40 years worldwide (see reviews in Gibson & Yoshiyama, 1999; Almada & Faria, 2004). These studies have been concentrated in temperate areas, while information from tropical seas is still scarce (Horn, Martin & Chotkowski, 1999). Several authors (e.g. Thomson & Lehner, 1976; Gibson, 1988; 1992; 1993; Gibson & Yoshiyama, 1999) have classified tidepool fishes into three main categories which vary in terminology among studies: (1) Resident or primary residents, are commonly small and cryptic species that spend all their life cycle within tidepools; (2) Transient or secondary residents, are those that spend only parts of their life cycle within tidepools and then migrate to subtidal habitats as adults; and (3) Accidental or casual visitors are typical subtidal species that can enter intertidal zones to forage and occasionally get trapped in tidepools at ebb tide. Although this classification is useful to distinguish major components of these assemblages, the lack of homogeneity in the criteria used to distinguish between categories prevents detailed comparisons among studies.
Although the Tropical Eastern Pacific regional fauna is not as diverse as the one in the Indo-West Pacific region, it presents a high level of endemism due to its isolation from other biogeographical regions (Glynn & Ault, 2000; Robertson & Allen, 2002). The near-shore fish fauna in the region is relatively well known mainly on coral reefs. In recent years, the great importance that rocky reefs may have for reef fish communities within the region has been pointed out (Robertson, 1998, Dominici-Arosemena & Wolff, 2006; Benfield, Baxter, Guzman & Mair, 2008). In this context, tidepool fishes on rocky shores of the region represent a subset of reef fish communities that has not been properly assessed.
This study aims to (1) document the short-term spatio-temporal patterns of tidepool fish assemblages found at Gorgona Island in the Colombian Pacific Ocean, (2) compare our results with data collected in the 1990’s (García & Zapata, unpublished data) at the same localities investigated here; and (3) compare the intertidal fish assemblage composition and organization found at Gorgona Island to that found at other localities in a wider area (Colombia and Eastern Pacific Ocean). By doing this, a regional synthesis of current knowledge on Eastern Pacific tidepool fish assemblages is provided.
Materials and methods
Study area: In the Tropical Eastern Pacific (TEP), the most isolated tropical marine biogeographic region of the world (Robertson, Grove & McCosker, 2004), rocky shore seascapes are predominant besides two large sandy and muddy coastline portions along the south eastern Gulf of California (350km) and the expanse from southern Mexico to El Salvador (1000km; Mora & Robertson, 2005; Robertson & Cramer, 2009). Other near-shore ecosystems such as coral reefs are uncommon, occupying <25km2 (Glynn & Ault, 2000). This holds true for the Colombian Pacific coast, with ca. 636 km of its shoreline dominated by rocky shores of different origin (López-Victoria et al., 2004). Rocky shores of volcanic origin are located in the northern part of the coast and in Gorgona and Malpelo Islands, whereas tertiary sedimentary rocky shores are located in the central (Buenaventura and Bahía Málaga) and southern parts (Istmo de Pichidó and Gallo Island) of the Colombian coast.
Gorgona is a continental island located in the southern part of the Colombian Pacific coast (2º58’10” N - 78º11’05” W). Marine habitats on the island include sandy beaches, coral reefs and rocky shores. In 2006, we sampled three different rocky intertidal areas on the Island: Ventana, Barra and Camaronera. The first two sites were located in the south and southeast portions of the Island and are separated by a <50m-long sandy beach. These two sites presented ca. 200m of a exposed rocky platform at low tide The third site, Camaronera, is located on the southwestern side of the Island, separated from the other two sites by a 1km-long sandy beach. The topographic characteristics of this site vary considerable compared to those of Ventana and Barra. Camaronera is characterized by gravel and sandy substrates. Subtidal habitats near the three sites include mixed sandy and rocky bottoms surrounded by small pocilloporid coral patches (Zapata, 2001a).
Sampling: Six tidepools located between the mid and low intertidal zone at each of the three sites described above were randomly selected during a preliminary field trip in March 2006. Repeated samplings of these pools were made during four months (April-July 2006) using a clove-oil solution (eugenol). This anaesthetic is used commonly for collecting coral reef and tidepool fishes (Griffiths, 2000; Ackerman & Bellwood, 2002; Robertson & Smith-Vaniz, 2008). The solution was prepared in a 1:8 clove oil to ethanol proportion and sprayed directly into the tidepools, particularly in small crevices and under boulders where fish may hide. The quantity of anaesthetic employed was variable depending on tidepool size. The reaction on fish species was also variable with the most vulnerable species showing erratic movements after ca. 5 minutes of clove oil application. Using small dip nets all fish within tidepools were captured searching under rocks and crevices when necessary. All fishes were then identified to species with field taxonomic guides adapted from Robertson & Allen (2002), counted and afterwards returned to adjacent tidepools where they rapidly recovered from the anaesthetic effect.
Prior to fish collections, length, width and depth of tidepools were measured to calculate sampled areas and volumes. Length was considered as the distance between the most distant edges of a pool and the width as the distance perpendicular to the midpoint length axis. Depth was calculated by averaging 10-15 random measures to the nearest 0.5 cm in different sections of the pools. With these data, an estimation of fish density (fish m-2 and fish m-3) was subsequently obtained.
Fish collections in 1993 were made using the ichthyocide rotenone (García & Zapata, unpublished data), which is regarded as a very effective sampling technique (Ackerman & Bellwood, 2002; Robertson & Smith-Vaniz, 2008). Collections were made monthly from January to July (except February) at the same sites where sampling was carried out in 2006 (i.e. Ventana, Barra and Camaronera). An average of seven tidepools per site for each month were sampled with rotenone during low tide. All fish were removed from the selected tidepools and subsequently preserved in 10% formaldehyde. In the laboratory fishes were transferred to 70% ethanol solution and identified using appropriate taxonomic keys for the area (e.g., Allen & Robertson, 1994).
The classification of species according to its occurrence in time was based on Gibson & Yoshiyama (1999). Resident species were those with an exclusive presence in tidepools. Transient species were those present only as juveniles in tidepools and also seen commonly in subtidal areas either as juveniles or adults. Accidental visitor species were those inhabiting subtidal habitats that occasionally got trapped in tidepools.
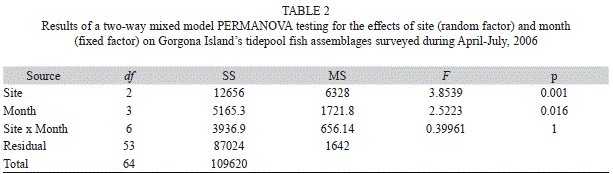
Data Analyses: Data taken in 2006 were used to test for differences in tidepool fish composition among sampling sites and months. For this purpose a 2-way permutational analysis of variance (PERMANOVA, Anderson, 2001) was performed. The analysis was conducted treating Site (three levels) as a random factor and Month (four levels) as a fixed factor. Samples among months were assumed to be independent due to the demonstrated rapid recovery of tidepool fish assemblages after perturbation (Willis & Roberts, 1996). The PERMDIST routine (an analog of the univariate Levene’s test) was used to check if results from the PERMANOVA test were an artifact of difference in dispersion among groups (Anderson, 2006). To explore the multivariate patterns shown in the PERMANOVA test, constrained and unconstrained ordinations were used (non-metric multidimensional scaling or nMDS and canonical analysis of principal coordinates or CAP). Species responsible for differences along the CAP axes were determined as those species’ abundances having Spearman correlations >0.4 with any of the canonical discriminant axes (Anderson & Willis, 2003). All analyses were performed on square-root transformed density data (fish m-2) based on the Bray-Curtis similarity measure.
Due to the different sampling procedures employed in 1993 (rotenone, García & Zapata, unpublished data) and 2006 (clove oil), data from both periods could not be directly compared. However, in an attempt to evaluate: (1) the possible effect of the sampling method and, (2) the persistence of these assemblages over time, we used an unconstrained ordination technique (Principal coordinates analysis-PCO) on species relative abundance data for the three sites that were sampled in 1993 and 2006. Data were left untransformed and the Bray-Curtis similarity measure was employed to generate the similarity matrix upon which the PCO was performed. PCO is very flexible as it is the non-metric multi-dimensional scaling (nMDS) ordination technique, because it can be used on any resemblance matrix. However, PCO differs from nMDS in that PCO projects points in space onto axes according to the chosen resemblance measure, whereas nMDS is focused in preserving the rank order of similarities in a defined number of dimensions (Anderson, Gorley & Clarke, 2008).
Tidepool fish assemblages along the Eastern Pacific Ocean were compared based on 17 sampling locations spanning the West coast of USA at 43°N to the southern part of Chile at 36°S. The data were obtained from 10 studies and one unpublished data set from Colombia (Castellanos-Galindo, periods). All these studies had as a main aim the sampling of the entire tidepool ichthyofauna at each location. Data from these studies were organized and analyzed at the species and family level. Classification analyses were then performed using the Bray-Curtis similarity measure on presence/absence data (Sørensen similarity measure). PERMANOVA+for PRIMER software was used to conduct all analyses (Anderson et al., 2008).
Results
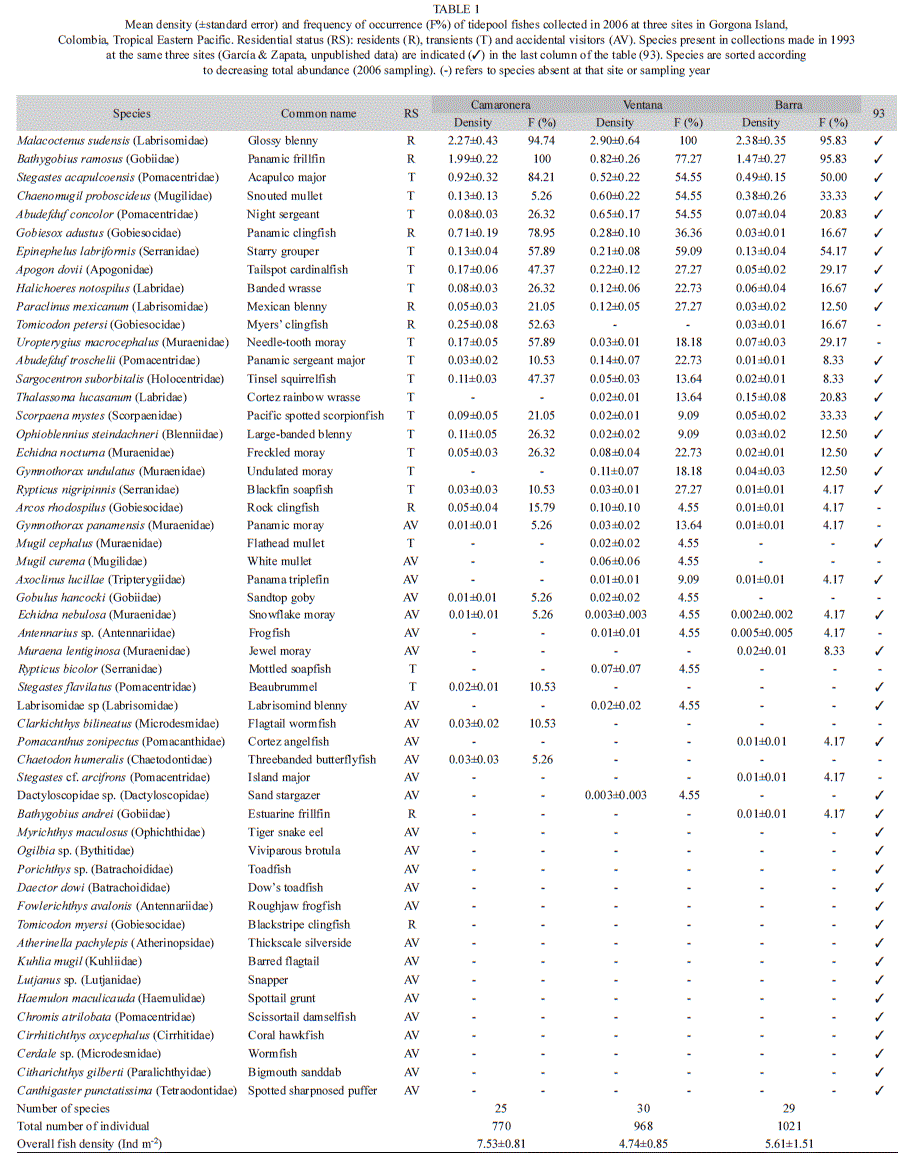
Small-scale spatio-temporal patterns. The tidepool ichthyofauna of Gorgona Island during April-July 2006 comprised 38 species grouped in 18 families. Three taxa, Malacoctenus sudensis (Labrisomidae), Bathygobius ramosus (Gobiidae) and Stegastes acapulcoensis (Pomacentridae), accounted for 67% of the total abundance. The most speciose families were Muraenidae (six species) and Pomacentridae (five species). The abundance of Labrisomidae, Gobiidae, Pomacentridae, Mugilidae and Gobiesocidae accounted for >85% of the total abundance (Table 1). Twenty one percent of the species were classified as residents, 47% as transients, and 32% as accidental visitors. The relative abundance of these same groups, however, was greater for residents (65%) than for transient species (34%) with accidental visitors representing a small fraction of the assemblage (1%).
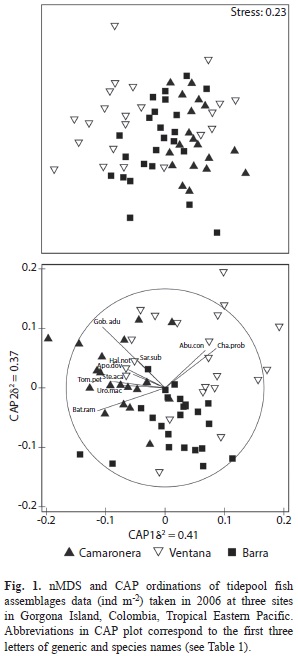
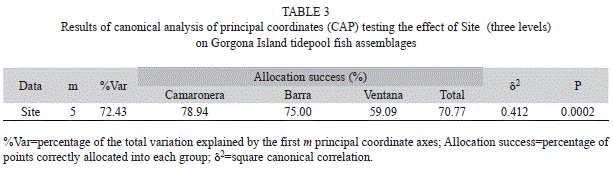
The PERMAVOVA test showed significant differences in fish assemblages among sites and among sampling dates (Table 2). The variation in multivariate dispersion, however, was significant for the site factor (PERMDISP, F=7.11, p=0.022), indicating that caution should be taken when interpreting the PERMANOVA results. Non-metric multi-dimensional scale (nMDS) and CAP ordinations showed different results with no clear aggregation patterns among samples in the nMDS (high stress value =0.23) and a relatively well-defined grouping of samples along the CAP axis 1 in the CAP ordination (Fig. 1). In the CAP ordination, samples from Camaronera were generally located to the left of the CAP axis 1, whereas samples from Ventana and Barra were distributed to the right (Fig. 1). As expected, the leave-one-out allocation success from the CAP analysis was higher at Camaronera and lower at Ventana, implying that assemblages from Ventana were harder to predict than the ones from Camaronera or Barra (Table 3). This is also evident in the PERMDISP results, in which the largest dispersion of assemblages was observed in Ventana samples (average Bray-Curtis distance-to-centroid =41%).
Ten species were found to be strongly correlated with CAP axis 1, thus differentiating assemblages from the three sites. Abudefduf concolor (r=0.41) and Chaenomugil proboscideus (r=0.52) were species strongly correlated with CAP axis 1 and assemblages at Ventana. The mean density (ind m-2±SD) of these species at Ventana was the highest amongst sites (A. concolor =0.65±0.17 and C. proboscideus =0.60±0.22). Similarly, species with strong correlation with CAP axis 1 (Camaronera) were Gobiesox adustus (r=-0.63), Bathygobius ramosus (r=-0.68), Tomicodon petersi (r=-0.55), Uropterygius macrocephalus (r=-0.43) and Sargocentron suborbitalis. (r=-0.27). All these species presented the highest densities at Camaronera (Table 1).
In 1993, 9 395 fish were collected during the six-month study period. Forty-one species grouped in 26 families comprised the tidepool fish assemblage. Five species accounted for ca. 65% of the total fish abundance (M. sudensis-26.7%, S. acapulcoensis-13.5%, A. concolor-9.7, B. ramosus-8.2% and Abudefduf troschelii-7.6%). This indicates a higher evenness in the assemblage from 1993 compared to the one sampled in 2006, when the same relative abundance was reached by only three species. Despite this difference, four of the five most important species during both sampling periods were the same. The dominant families in 1993, in decreasing order, were Labrisomidae, Pomacentridae, Gobiidae, Labridae and Mugilidae. These five families accounted for 80% of the total abundance. Finally, the relative abundance of the residential categories in 1993 was different compared to sampling in 2006. Relative abundance of transient species reached ca. 60%, whereas residents did not reach 40% of the total abundance. This was due to the considerably fewer number of individuals captured of the resident species B. ramosus in 1993. In contrast, three pomacentrids (S. acapulcoensis, A. concolor and A. troschelii) and one labrid (Halichoeres notospilus), all transient species, were consistently more abundant during sampling in 1993 compared to 2006.
Twenty-seven species were shared between both sampling periods. Fifteen species were found only in 1993 when sampling with rotenone, whereas 11 species were found only in 2006 with clove oil sampling (Table 1). The PCO ordination showed separation between both sampling periods (1993 and 2006), with most samples collected in 2006 distributed to the left axis of the PCO axis 1 (Fig. 2). Similarly, most samples collected in 1993, independent of site, were grouped to the right of the PCO axis 1. The species responsible for differences among sampling times (correlations >0.4 with PCO axis 1) were B. ramosus (r=-0.88), T. petersi (r=-0.67), Arcos rodosphilus (r=-0.50), M. sudensis (r=-0.49) and U. macrocephaulus (r=-0.44, all correlated with samples from 2006); and S. acapulcoensis (r=0.71), Rypticus nigripinnis (r=0.72), H. notospilus (r=0.60), Porichtys sp. (r=0.55), A. troschelii (r=0.52), S. suborbitalis (r=0.51) and Apogon dovii (r=0.49, all correlated with samples from 1993).
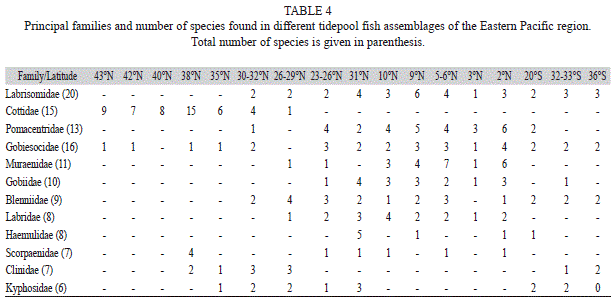
Zoogeographical analysis: Based on all the available data, the tidepool ichthyofauna in the Eastern Pacific Ocean (43°N-36°S) contained at least 214 species grouped in 52 families. Perciformes represented 70% of the families and 63% of species present in this region. Scorpaeniformes and Anguilliformes were the next two most species-rich fish orders (13% and 6%, respectively). Within the Perciformes, the suborder Blennioidei had the highest species representation (44), with some of these species being especially abundant and dominant in assemblages from the Panamic province (sensu, Robertson & Cramer, 2009) and also in assemblages of Chile. The suborder Labroidei (28 species) was also especially dominant in the tropical portion of the region with some of the species contributing importantly to the transient residential category. Pomacentridae and Labridae were the most important labroid families in the tropics, whereas Embiotocidae was found from 26° to 38°N. In the northern portion of the Eastern Pacific (>26°N), Scorpaeniformes became the dominant group, being Cottidae a very diverse family in tidepools of the area. In terms of abundance, assemblages in this area were almost exclusively dominated by Cottidae with some species of Stichaeidae or Clinidae having a relative importance in some areas. In the southern part of the region (<20°S), apart from the Blennioidei, a Percoidei family (Kyphosidae) became very important in terms of abundance within these assemblages. At the southernmost part of this area (36°S), the suborder Gobiesocoidei was very abundant together with the Blennioidei (Table 4).
When comparing tidepool fish assemblages along the latitudinal gradient three consistent groups were observed. Assemblages with tropical affinities were grouped coinciding with the Tropical Eastern Pacific region boundaries. This group includes studies carried out in the southernmost part of the west coast of the Gulf of California (23°-26°N; Ruiz-Campos et al. (2010)), a study carried out inside the Gulf of California (Cortez Province; Thomson & Lehner (1976)) and studies carried out in the Panamic province (Costa Rica and Colombia; Weaver (1970), Castellanos-Galindo, Giraldo & Rubio (2005) and this study). A second group was defined by assemblages of the northern part of the Eastern Pacific coast (35°-43°N; Yoshiyama, Sassaman & Lea (1986)), which are part of the California and Oregon provinces (Fig. 3). Finally, the tidepool fish assemblage from the warm-temperate Peru-Chilean Province is clearly distinguished from the rest (Fig. 3).
Discussion
Species richness, composition and residential status: The tidepool fish assemblage found at Gorgona Island was relatively diverse compared to assemblages at other localities in the Colombian Pacific coast or the rest of the Tropical Eastern Pacific Ocean. These results combined with information from other localities in the Colombian Pacific indicate a dominance of Perciform families represented mainly by the sub-orders Labroidei, Blennioidei and Gobioidei in this part of the TEP. At Gorgona Island, a considerably high number of species was found, with M. sudensis (Labrisomidae), S. acapulcoensis (Pomacentridae) and B. ramosus (Gobiidae) being the most abundant species. Resident species at Gorgona belonged to the same families found at another locality of the Colombian Pacific with an estuarine environment (Bahía Málaga; Castellanos-Galindo et al., 2005), but the total number of species and the number of species per family was higher in Gorgona than at Bahía Málaga (14 species). Additional examination of tidepool fish assemblages at two other localities in the northern Colombian Pacific (Ensenada de Utría and Cabo Marzo), where rocky shores are the predominant seascape, show that these assemblages are similarly diverse to those encountered at Gorgona Island (ca. 40 species; Castellanos-Galindo, unpublished data). Labrisomidae, Gobiidae, Pomacentridae, Gobiesocidae, Muraenidae and Labridae dominate the assemblage composition at these three localities.
The great differences in species richness between Gorgona Island and Bahía Málaga can be explained by a combination of factors, which include the different sampling effort (time, number of pools and number of sites) and differences in the topographic characteristics of the tidepools sampled. However, the most important difference may be the presence of a richer subtidal fish community in the marine environment of Gorgona Island compared to the poorer estuarine subtidal fish community in Bahía Málaga. The presence of larger areas of rocky and coral reefs in Gorgona in comparison to Bahía Málaga, may also explain the higher number of transient and accidental visitors found in tidepools of Gorgona. A comparison with results of visual censuses of coral reef fishes at Gorgona indicates that at least 20 coral reef fish species are commonly found in tidepools (Zapata & Morales, 1997; Zapata, 2001b). A very similar result was found by Weaver (1970) in the Pacific coast of Costa Rica where the assemblage structure of the tidepool ichthyofauna differed between three localities with different subtidal characteristics (estuarine, coral reefs). Playas del Coco, adjacent to coral reefs, had a higher species richness (28) than the other localities.
When comparing the composition of residents there is concordance at the family level between studies in Costa Rica and Colombia; however, Gorgona presents a higher number of species within each of these families. Differences can also be observed in the dominance (relative abundance) of fish species. Whereas in Bahía Málaga, Colombia, one single species (Bathygobius ramosus) comprises >50% of the total tidepool fish assemblage, approximately this same percentage is shared by the three principal species in Gorgona Island (M. sudensis, S. acapulcoensis, B. ramosus). Nonetheless, when comparing the five most abundant species at each locality, the same three species dominate.
In terms of relative abundance, the residential categories differed between Gorgona Island and Bahía Málaga, even though the number of resident species in the latter locality is higher. Griffiths (2003) found a very similar pattern in temperate Australia as in Bahía Málaga, where permanent resident species dominate over transients. Conversely, Thomson and Lehner (1976) found a pattern in the Gulf of California more similar to the one observed at Gorgona Island, where the percentage of transient species was higher. Mahon & Mahon (1994) in the Caribbean Sea, documented a dominance of true residents over partial ones (transients species sensu Gibson & Yoshiyama (1999)); however, many juvenile Anguilliform species were classified as true residents by these authors, whereas Gibson & Yoshiyama (1999) consider them as transient species. If anguilliforms are pooled with the rest of partial residents, both categories (residents and transients) reach almost the same percentages. Therefore, it is reasonable to expect that tidepool fish assemblages will be more likely to contain a higher number of transient species when adjacent subtidal ichthyofaunas are diverse (i.e., near coral reefs). Conversely, when tidepools occur near species-poor subtidal habitats, resident species will be more likely to dominate tidepool fish assemblages.
The use of intertidal habitats by fishes at high tide has rarely been examined in tropical areas. However, Castellanos-Galindo, Krumme & Willis (2010) examined this aspect in rocky shores of the northern Colombian Pacific, and found that 73 fish species used these intertidal habitats at high tide. Common species can be identified between those observed at high tide and those remaining in tidepools at low tide, especially for members of the families Pomacentridae and Labridae. However, a large proportion of cryptic resident species found in tidepools, were not recorded by Castellanos-Galindo et al. (2010), due to the methodology employed in that study (i.e., underwater visual censuses). Combining both sources of information (tidepool sampling and intertidal visual censuses at high tide) it could be estimated that ca. 100 fish species use intertidal habitats in Colombian Pacific rocky shores. This highlights the importance that these habitats have for shore fishes and also suggest the existence of important links between subtidal and intertidal areas that are regulated by tides and the availability of intertidal prey sources for fishes.
Patterns of temporal variation: Temporal variation in tidepool fish assemblages has been evaluated at several scales from day/night to periods of 10 years. In a month or year-round scale, variation in community structure has been detected in the majority of studies (review in Gibson & Yoshiyama, 1999, but see also Davis, 2000a; Griffiths, 2003; Arakaki & Tokeshi, 2006). However, other surveys, which include some in tropical regions, have found that such variability is not present (temperate: Prochazka, 1996; tropical: Chang, Lee, Lee & Chen, 1973; Mok & Wen, 1985; Castellanos-Galindo et al., 2005). Temporal variations often have been attributed to the strong changing seasonal conditions observed year-round in temperate latitudes, where most of these studies have been carried out. No short-term variation was observed during the four months of this study. Although the statistical tests suggested differences between sampling months, these were driven by the high variance associated to each of the samples. The leave-one-out allocation success procedure for the Month factor was low (40%). This indicates difficulties in differentiating samples coming from a specific month mainly because of large multivariate dispersion within months. These results are in contradiction with most of the studies carried out in temperate regions, but not with the studies carried out in the tropics. The absence of strong seasonal changes in the Colombian Pacific compared to temperate areas may explain these contrasting results. The relative stability of environmental conditions in these coastal biotopes may allow these assemblages to maintain their structure and function through short temporal scales. It will be necessary to examine if variation in community structure occurs during the rainy season in Gorgona Island (May-December) to extend our conclusions to larger temporal scales.
Finally, studies at greater temporal scales (years or decades), have taken place mainly in localities of the West coast of the USA. Similarities in assemblage structure are very consistent there over the sampling periods indicating that these assemblages are persistent over large temporal scales (Grossman, 1982; Yoshiyama, 1981; Yoshiyama et al., 1986, see review in Almada & Faria, 2004). The results obtained from a comparison of samplings in 1993 and 2006 in Gorgona Island support the results obtained in temperate areas. The dominant species were the same between sampling periods. The number of shared species (27 out of 53) between both periods can reflect that some degree of unpredictability in the composition of transient and accidental visitors exist, but that the core members of the fish assemblage are persistent over time. The variation in the proportions found is very likely due to the use of different sampling techniques. In 1993, rotenone was used. This substance is highly effective, but has been shown to have differing effects on fish species. Weaver (1970) mentioned that B. ramosus (Gobiidae) was highly resistant to rotenone when poisoning tidepools of Pacific Costa Rica. This may be the reason why the relative abundance of this species changed between 1993 and 2006 sampling at Gorgona Island (8% and 22%, respectively). Likewise, Weaver (1970) identified species of Pomacentridae as highly susceptible to rotenone. The relative abundance of the species of this family was considerably higher in 1993 than in 2006 at Gorgona Island.
Another important conclusion derived from the low variability in the assemblage structure over time is that this assemblage is able to recover in a short period (<1 month in this case), after severe perturbations such as those caused by sampling (i.e. defaunation). It has been suggested previously that re-colonization may occur from individuals of adjacent pools migrating to defaunated pools within few days (Willis & Roberts, 1996). Finally, phenomena occurring at large temporal scales, such as the El Niño-Southern Oscillation (ENSO), may affect community structure (Davis, 2000a). This is an aspect that has been poorly examined in tidepool fish assemblages. The TEP offers great potential to improve our understanding of the effects of ENSO on intertidal rocky shore communities because this region is greatly affected by this phenomenon.
Patterns of spatial variation: Differences in habitat complexity often lead to changes in assemblage structure (Mahon & Mahon, 1994; Griffiths, Davis & West, 2006; Macieira & Joyeux, 2011). This may be the case for the inter-site differences observed at Gorgona Island. Griffiths et al. (2006) showed that the availability of rocks and macro-algae significantly affected the species composition and abundance of fishes visiting tidepools in Australia. Davis (2000b) reached a similar conclusion after she manipulated tidepool complexity in California. At Gorgona Island, the different topographic characteristics of the sites where tidepools were sampled may be partially responsible for the differences in assemblage composition. Mean fish density (especially of benthic species; e.g. G. adustus, B. ramosus, U. macrocephalus, T. petersi) was considerably higher at Camaronera than at the other two sites. At Camaronera, shallower tidepools composed of gravel and sand might offer more microhabitats to (benthic) fishes than tidepools at Ventana and Barra where tidepools had relatively low microhabitat complexity. In contrast, schooling species and those inhabiting the water column (e.g. C. proboscideus, A. troschelii) were more characteristic of Ventana than of Camaronera. Further studies should directly test the effect of complexity on fish assemblage composition at these sites.
Latitudinal patterns in the Eastern Pacific. The taxonomic composition of tidepool fish assemblages found in the Eastern Pacific is in agreement with results of previous analyses by Chotkowski, Buth & Prochazka (1999) and Prochazka, Chotkowski & Buth. (1999). The number of tidepool fish species found in the Eastern Pacific corresponds to ca. 30% the total number of species documented by Chotkowski et al. (1999). This percentage is likely to be an over-estimation considering that there is not an appropriate documentation of tidepool fish faunas in several tropical diverse areas of the Indo-West Pacific region.
The relatively small area that is available for colonization (especially in rocky shore areas) is the main restriction that fishes that reside in intertidal areas may face. This constraint is probably responsible for the limited range of body plans, sizes and taxonomic groups that are observed in tidepool fishes (Chotkowski et al., 1999). The Blennioidei and Labroidei suborders accounted for 21% and 12% of the total number of 702 species found in the revision made by Chotkowski et al. (1999). In the present comparison of tidepool fishes in the Tropical Eastern Pacific, these same suborders accounted for 29% and 19% of the total number of tidepool fish species (Blennioidei and Labroidei). The Neotropics are a major center of diversification for members of the Blennioidei with a high degree of endemism (Hastings, 2009). Three of the six families that comprise this suborder (Labrisomidae, Chaenopsidae and Dactyloscopidae) are endemic or nearly endemic to the New World. Most species within this suborder inhabit shallow coastal waters and have small body sizes. These two characteristics are likely responsible for the successful invasion and residency of this group in rocky intertidal areas (i.e. tidepools) in the Neotropics (especially the Tropical Eastern Pacific). Contrary to the Blennioidei, the Labroidei are transient components of the tidepool ichthyofauna of the Tropical Eastern Pacific region. Labridae and Pomacentridae are species-rich families in rocky reef areas, where access to intertidal habitats (including tidepools) offers additional food sources and/or refuge from predation to young or small individuals of these species.
At regional scales, few attempts have been made to document tidepool fish assemblage differences, but recently Boyle & Horn (2006) made comparisons at different levels of central Chile and California tidepool fish assemblages in the Eastern Pacific finding no common species and only two shared families (Gobiesocidae and Clinidae). The present regional comparison of tidepool ichthyofaunas between the temperate and tropical regions of the Eastern Pacific Ocean shows taxonomic differences even at the family level in line with the observations of Prochazka et al. (1999). In the northern hemisphere at 37ºN, Yoshiyama (1981) found a dominance of species (more than 50% of all species) by members of the families Cottidae (Scorpaeniformes) and Stichaeidae (Perciformes); these families are totally absent from tropical southern latitudes. The most speciose families of the tropical regions are Pomacentridae, Muraenidae, Gobiidae and Labrisomidae. It is noteworthy that anguilliform species (mainly Muraenidae) are present only in the tropical portion of the Eastern Pacific (Table 4). This was also noted by Mahon & Mahon (1994) in the Caribbean where they found a similar number of species from this order.
In the southern part of the Eastern Pacific at 32-33ºS, Muñoz & Ojeda (1997) identified a predominance of species of the families Labrisomidae and Gobiesocidae. Nevertheless, the species dominating these assemblages belonged to Blenniidae (Scartichthys viridis), Tripterygiidae (Tripterygion chilensis) and Kyphosidae (Girella laevifrons). The presence of these families in intertidal habitats of the Tropical Eastern Pacific is marginal and often composed of transients and accidental visitors (i.e. Ophioblennius steindachneri and Axoclinus lucillae). Although dispersed, tidepool fish studies in the Tropical Eastern Pacific region (and the Colombian Pacific coast) have been slowly increasing, filling in a gap acknowledged by previous authors (Prochazka et al., 1999). Further investigation at discrete regions of Central America and the southern limit of the TEP (i.e., Ecuador and Peru), will help to corroborate the patterns drawn in this initial zoogeographic comparison.
In summary, tidepool fish assemblages found at Gorgona Island are likely among the most species-rich of this type of assemblages in the TEP region and the whole Eastern Pacific coast. The observed inter-site differences in assemblage structure may be directly related to differences in the topographic structure of tidepools found at each of the study sites. Rotenone and clove oil are both effective methods to survey tidepool fish assemblages, however, caution should be taken when comparing results due to differences in the susceptibility of fish species to each of these methods. These differences may be reflected in changes in the proportion of the dominant species. The regional patterns observed here reflect commonly defined biogeographical patterns observed for the shore fish fauna of the Eastern Pacific. A distinct species composition was observed for Chilean and northern USA west coast localities, whereas similarities could be observed for localities in the TEP including the Cortez province. Tidepool fish assemblages in some areas of the TEP are as species-rich as in the same habitats in the Caribbean region. The later region is considered to have greater fish species richness in certain shallow water habitats (e.g. coral reefs; Floeter et al., 2008). Therefore, other factors operating at local scales may be important in determining the number of fish species in these habitats.
The dynamics of tidepool fish assemblages at high tide and the possible connections to the subtidal environments are practically unknown in the region and should be addressed in the future to fully understand the importance of these particular assemblages in the whole coastal seascape continuum. Finally, tidepool fish assemblages at Gorgona Island and in the whole TEP constitute excellent systems for testing the effects of climatic variability intrinsic to this region, providing understanding that is urgently needed in an era of accelerated climatic changes.
Acknowledgments
Sampling at Gorgona Island in 2006 was possible thanks to the logistic and financial support of Animal Ecology Research Group at Universidad del Valle, Scientific Research Station Henry von Prahl at National Natural Park Gorgona, Colciencias-Univalle Research Project RC-243-2004/1106-09-17092 to A. Giraldo and F.A. Zapata, and Colciencias “Young Researcher Program” grant to G. Castellanos-Galindo (p-20005-0028). Tide Pool sampling was granted by UAESPNN under research permit DTSO-gr-003/2006 to A. Giraldo. Information on tidepool fish assemblages of Gorgona in 1993 was extracted from samples collected by H.F. García for his unpublished B.Sc. thesis supervised by F.A. Zapata and deposited at Universidad del Valle. We are indebt to E. Escarria, E. Velasco, D. Ramirez and J. Benitez for their dedication during sampling activities. P. Hastings, J. McCosker and J. Van Tassell provided useful literature. English proofreading by P. Tuda and comments on early versions of this manuscript by U. Krumme, M. Wolff and I. Freytag and two anonymous reviewers are deeply appreciated.
References
Ackerman, J. L. & Bellwood, D. R. (2002). Comparative efficiency of clove oil vs. rotenone for sampling tropical reef fish assemblages. Journal of Fish Biology, 60: 893-901. [ Links ]
Allen, G. R. & Robertson, D. R. (1994). Fishes of the tropical eastern Pacific. University of Hawaii Press, Honolulu, Hawaii. [ Links ]
Almada, V. C. & Faria, C. (2004). Temporal variation of rocky intertidal resident fish assemblages-patterns and possible mechanisms with a note on sampling protocols. Reviews in Fish Biology and Fisheries, 14: 239-250. [ Links ]
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Australian Journal of Ecology, 26: 32-46. [ Links ]
Anderson, M. J. (2006). Distance-based tests for homogeneity of multivariate dispersions. Biometrics, 62: 245-253. [ Links ]
Anderson, M. J. & Willis, T. J. (2003). Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology, 84: 511-525. [ Links ]
Anderson, M. J., Gorley, R. N. & Clarke, K. R. (2008). PERMANOVA+for PRIMER: Guide to software and statistical methods. PRIMER-E:Plymouth, United Kingdom. [ Links ]
Arakaki, S. & Tokeshi, M. (2006). Short-term dynamics of tidepool fish community: diel and seasonal variation. Environmental Biology of Fishes, 76: 221-235. [ Links ]
Benfield, S., Baxter, L., Guzman, H. M. & Mair, J. M. (2008). A comparison of coral reef and coral community fish assemblages in Pacific Panama and environmental factors governing their structure. Journal of the Marine Biological Association of the United Kingdom, 88: 1331-1341. [ Links ]
Berrios, V. L. & Vargas, M. E. (2000). Estructura del ensamble de peces intermareales de la costa rocosa del norte de Chile. Revista de Biología Marina y Oceanografía, 35:73-81. [ Links ]
Boyle, K. S. & Horn, M. H. (2006). Comparison of feeding guild structure and ecomorphology of intertidal fish assemblages from central California and central Chile. Marine Ecology Progress Series, 319: 65-84. [ Links ]
Briggs, J. C. & Bowen, B. W. (2012). A realignment of marine biogeographic provinces with particular reference to fish distribution. Journal of Biogeography, 39: 12-30. [ Links ]
Castellanos-Galindo, G. A., Giraldo, A. & Rubio, E. A. (2005). Community structure of an assemblage of tidepool fishes on a Tropical Eastern Pacific rocky shore, Colombia. Journal of Fish Biology, 67: 392-408. [ Links ]
Castellanos-Galindo, G. A., Krumme, U. & Willis, T. J. (2010). Tidal influences on fish distributions on tropical eastern Pacific rocky shores (Colombia). Marine Ecology Progress Series, 416: 241-254. [ Links ]
Chang, K. H., Lee, S. C., Lee, J. C. & Chen, C. P. (1973). Ecological study on some intertidal fishes of Taiwan. Bulletin Institute of Zoology Academia Sinica, 12: 45-50. [ Links ]
Chotkowski, M. A., Buth, D. G. & Prochazka, K. (1999). Systematics of Intertidal fishes. In M. H. Horn, K. L. M. Martin & M. A. Chotkowski (Eds.), Intertidal Fishes: Life in two worlds (pp. 297-331). Academic Press, London, United Kingdom. [ Links ]
Davis, J. L. D. (2000a). Changes in a tidepool fish assemblage on two scales of environmental variation: Seasonal and El Niño Southern Oscillation. Limnology and Oceanography, 45: 1368-1379. [ Links ]
Davis, J. L. D. (2000b). Spatial and seasonal patterns of habitat partitioning in a guild of southern California tidepool fishes. Marine Ecology Progress Series, 196: 253-268. [ Links ]
Dominici-Arosemena A. & Wolff, M. (2006). Reef fish community structure in the Tropical eastern Pacific (Panamá): living on a relatively stable rocky reef environment. Helgoland Marine Research, 60: 287-305. [ Links ]
Floeter S. R., Rocha, L. A., Robertson, D. R., Joyeux, J. C., Smith-Vaniz, W. F., Wirtz, P., Edwards, A. J., Barreiros, J. P., Ferreira, C. E. L., Gasparini, J. L., Brito, A., Falcón, J. M., Bowen, B., & Bernardi, G. (2008). Atlantic reef fish biogeography and evolution. Journal of Biogeography, 35: 22-47. [ Links ]
Gibson, R. N. (1988). Patterns of movement in intertidal fishes. In G. Chelazzi & M. Vannini (Eds.), Behavioural Adaptation to Intertidal Life (pp. 55-63). Plenum Press, New York, USA. [ Links ]
Gibson, R. N. (1992). Tidally synchronized bahaviuor in marine fishes. In M. A. Ali (Ed.), Rhythms in Fishes (pp. 55-63). Plenum Press, New York. USA. [ Links ]
Gibson, R. N. (1993). Intertidal teleosts: life in a fluctuating environment,. In T. J. Pitcher (Ed.), Behaviour of Teleost Fishes (pp. 513-536). Chapman & Hall, London, United Kingdom. [ Links ]
Gibson, R. N. (2003). Go with the flow: tidal migration in marine animals. Hydrobiologia 503: 153-161. [ Links ]
Gibson, R. N. & Yoshiyama, R. M. (1999). Intertidal fish communities. In M. H. Horn, K. L. M. Martin & M. A. Chotkowski (Eds.), Intertidal Fishes: Life in two worlds (pp. 264-296) Academic Press, London, United Kingdom. [ Links ]
Glynn, P. W. & Ault, J. S. (2000). A biogeographic analysis and review of the far eastern Pacific coral reef region. Coral Reefs, 19: 1-23. [ Links ]
Griffiths, S. P. (2000). The use of clove oil as an anaesthetic and method for sampling intertidal rockpool fishes. Journal of Fish Biology, 57: 1453-1464. [ Links ]
Griffiths, S. P. (2003). Rockpool ichthyofaunas of temperate Australia: species composition, residency and biogeographic patterns. Estuarine, Coastal and Shelf Science, 58: 173-186. [ Links ]
Griffiths, S. P., Davis, A. R. & West, R. J. (2006). Role of habitat complexity in structuring temperate rockpool ichthyofaunas. Marine Ecology Progress Series, 313: 227-239. [ Links ]
Grossman, G. D. (1982). Dynamics and organization of a rocky intertidal fish assemblage: the persistence and resilience of taxocene structure. American Naturalist, 119: 611-637. [ Links ]
Hastings, P. A. (2009). Biogeography of New World blennies. In R. Patzner, E. Gonçalves, P. Hastings & B. Kapoor (Eds.), The Biology of Blennies (pp. 95-118). Science Publishers, Enfield, New Hampshire, United Kingdom. [ Links ]
Horn, M. H., Martin, K. L. M. & Chotkowski, M. A. (1999). Introduction. In M. H. Horn, K. L. M. Martin & M. A. Chotkowski (Eds.), Intertidal Fishes: Life in two worlds (pp.1-6). Academic Press, London, United Kingdom. [ Links ]
Little, C. & Kitching, J. A. (1996). The biology of rocky shores. Oxford University Press, Oxford, United Kingdom. [ Links ]
López-Victoria, M., Cantera, J. R., Díaz, J. M., Rozo, D. M., Posada, B. O. & Osorno, A. (2004). Estado de los litorales rocosos en Colombia: acantilados y playas rocosas. In INVEMAR (Ed.), Informe del estado de los ambientes marinos y costeros en Colombia: año 2003 (pp. 171-182). INVEMAR, Santa Marta, Colombia.
Macieira, R. M. & Joyeux, J. C. (2011). Distribution patterns of tidepool fishes on a tropical flat reef. Fishery Bulletin, 109: 305-315. [ Links ]
Mahon, R. & Mahon, S. D. (1994). Structure and resilience of a tidepool fish assemblage at Barbados. Environmental Biology of Fishes, 41: 171-190. [ Links ]
Martins, G. M., Hawkins, S. J., Thompson, R. C. & Jenkins, S. R. (2007). Community structure and functioning in intertidal rock pools: effects of pool size and shore height at different successional stages. Marine Ecology Progress Series, 329: 43-57. [ Links ]
Metaxas, A. & Scheibling, R. E. (1993). Community structure and organization of tidepools. Marine Ecology Progress Series, 98: 187-198. [ Links ]
Mok, H. & Wen, P. (1985). Intertidal fish community ecology on Lu Tao Island (Green Island), Taiwan. Journal of the Taiwan Museum, 38: 81-118. [ Links ]
Mora, C. & Robertson, D. R. (2005). Factors shaping the range-size frequency distribution of the endemic fish fauna of the Tropical Eastern Pacific. Journal of Biogeography, 32: 277-286. [ Links ]
Muñoz, A. A. & Ojeda, F. P. (1997). Feeding guild structure of a rocky intertidal fish assemblage in Central Chile. Environmental Biology of Fishes, 49: 471-479. [ Links ]
Nielsen, K. J. (2001). Bottom-up and Top-down forces in tide pools: test of a food chain model in an intertidal community. Ecological Monographs, 71: 187-217. [ Links ]
Palmer, J. D. (2000). The clocks controlling the tide-associated rhythms of intertidal animals. BioEssays, 22: 32-37. [ Links ]
Prochazka, K. (1996). Seasonal patterns in a temperate intertidal fish community on the west coast of South Africa. Environmental Biology of Fishes, 45: 133-140. [ Links ]
Prochazka, K., Chotkowski, M. A. & Buth, D. G. (1999). Biogeography of intertidal Fishes. In M. H. Horn, K. L. M. Martin & M. A. Chotkowski (Eds.), Intertidal Fishes: Life in two worlds (pp. 332-355). Academic Press, London, United Kingdom. [ Links ]
Quijada, P. A. & Caceres, C. W. (2000). Patrones de abundancia, composición trófica y distribución especial del ensamble de peces intermareales de la zona centro-sur de Chile. Revista Chilena de Historia Natural, 73: 739-747. [ Links ]
Robertson, D. R. (1998). Do coral-reef fish faunas have a distinctive taxonomic structure? Coral Reefs, 17:179-186. [ Links ]
Robertson, D. R. & Allen, G. (2002). Shorefishes of the Tropical Eastern Pacific: an information system. CD-ROM. Smithsonian Tropical Research Institute, Balboa, Panamá [ Links ].
Robertson, D. R., Grove, J. S. & McCosker, J. E. (2004). Tropical Transpacific shorefishes. Pacific Science, 58: 507-565. [ Links ]
Robertson, D. R. & Smith-Vaniz, W. F. (2008). Rotenone: an essential but demonized tool for assessing marine fish diversity. BioScience, 58: 165-170. [ Links ]
Robertson, D. R. & Cramer, K. L. (2009). Shore fishes and biogeographic subdivisions of the tropical eastern Pacific. Marine Ecology Progress Series, 380: 1-17. [ Links ]
Ruiz-Campos, G., González-Guzmán, S., Ramírez-Valdéz, A., González-Acosta, A. F., Castro-Aguirre, J. L. & De La Cruz-Agüero, J. (2010). Composition, density and biogeographic affinities of the rocky intertidal fishes on the western coast of the Baja California peninsula, Mexico. California Cooperative Oceanic Fisheries Investigations, 51: 210-220. [ Links ]
Thomson, D. A. & Lehner, C. E. (1976). Resilience of a rocky intertidal fish community in a physically unstable environment. Journal of Experimental Marine Biology and Ecology, 22: 1-29. [ Links ]
Weaver, P. L. (1970). Species diversity and ecology of tidepool fishes in three Pacific coastal areas of Costa Rica. Revista de Biología Tropical, 17: 165-185. [ Links ]
Willis, T. J. & Roberts, C. D. (1996). Recolonisation and recruitment of fishes to intertidal rockpools at Wellington, New Zealand. Environmental Biology of Fishes, 47: 329-343. [ Links ]
Yoshiyama, R. M. (1981). Distribution and abundance patterns of rocky intertidal fishes in Central California. Environmental Biology of Fishes, 6: 315-332. [ Links ]
Yoshiyama, R. M., Sassaman, C. & Lea, R. N. (1986). Rocky intertidal fish communities of California: temporal and spatial variation. Environmental Biology of Fishes, 17: 23-40. [ Links ]
Zapata, F. A. & Morales, Y. (1997). Spatial and temporal patterns of fish diversity in a coral reef at Gorgona Island. Proceedings of the 8th International Coral Reef Symposium, 1: 1029-1034. [ Links ]
Zapata, F. A. (2001a). Formaciones coralinas de Isla Gorgona. In L. M. Barrios & M. Lopez-Victoria (Eds.), Gorgona Marina: contribución al conocimiento de una isla única (pp. 27-50). INVEMAR, Serie Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
Zapata, F. A. (2001b). Peces Marinos de Gorgona: Genenralidades. In L. M. Barrios & M. Lopez-Victoria (Eds.), Gorgona Marina: contribución al conocimiento de una isla única (pp. 107-109). INVEMAR, Serie Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
Allen, G. R. & Robertson, D. R. (1994). Fishes of the tropical eastern Pacific. University of Hawaii Press, Honolulu, Hawaii. [ Links ]
Almada, V. C. & Faria, C. (2004). Temporal variation of rocky intertidal resident fish assemblages-patterns and possible mechanisms with a note on sampling protocols. Reviews in Fish Biology and Fisheries, 14: 239-250. [ Links ]
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Australian Journal of Ecology, 26: 32-46. [ Links ]
Anderson, M. J. (2006). Distance-based tests for homogeneity of multivariate dispersions. Biometrics, 62: 245-253. [ Links ]
Anderson, M. J. & Willis, T. J. (2003). Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology, 84: 511-525. [ Links ]
Anderson, M. J., Gorley, R. N. & Clarke, K. R. (2008). PERMANOVA+for PRIMER: Guide to software and statistical methods. PRIMER-E:Plymouth, United Kingdom. [ Links ]
Arakaki, S. & Tokeshi, M. (2006). Short-term dynamics of tidepool fish community: diel and seasonal variation. Environmental Biology of Fishes, 76: 221-235. [ Links ]
Benfield, S., Baxter, L., Guzman, H. M. & Mair, J. M. (2008). A comparison of coral reef and coral community fish assemblages in Pacific Panama and environmental factors governing their structure. Journal of the Marine Biological Association of the United Kingdom, 88: 1331-1341. [ Links ]
Berrios, V. L. & Vargas, M. E. (2000). Estructura del ensamble de peces intermareales de la costa rocosa del norte de Chile. Revista de Biología Marina y Oceanografía, 35:73-81. [ Links ]
Boyle, K. S. & Horn, M. H. (2006). Comparison of feeding guild structure and ecomorphology of intertidal fish assemblages from central California and central Chile. Marine Ecology Progress Series, 319: 65-84. [ Links ]
Briggs, J. C. & Bowen, B. W. (2012). A realignment of marine biogeographic provinces with particular reference to fish distribution. Journal of Biogeography, 39: 12-30. [ Links ]
Castellanos-Galindo, G. A., Giraldo, A. & Rubio, E. A. (2005). Community structure of an assemblage of tidepool fishes on a Tropical Eastern Pacific rocky shore, Colombia. Journal of Fish Biology, 67: 392-408. [ Links ]
Castellanos-Galindo, G. A., Krumme, U. & Willis, T. J. (2010). Tidal influences on fish distributions on tropical eastern Pacific rocky shores (Colombia). Marine Ecology Progress Series, 416: 241-254. [ Links ]
Chang, K. H., Lee, S. C., Lee, J. C. & Chen, C. P. (1973). Ecological study on some intertidal fishes of Taiwan. Bulletin Institute of Zoology Academia Sinica, 12: 45-50. [ Links ]
Chotkowski, M. A., Buth, D. G. & Prochazka, K. (1999). Systematics of Intertidal fishes. In M. H. Horn, K. L. M. Martin & M. A. Chotkowski (Eds.), Intertidal Fishes: Life in two worlds (pp. 297-331). Academic Press, London, United Kingdom. [ Links ]
Davis, J. L. D. (2000a). Changes in a tidepool fish assemblage on two scales of environmental variation: Seasonal and El Niño Southern Oscillation. Limnology and Oceanography, 45: 1368-1379. [ Links ]
Davis, J. L. D. (2000b). Spatial and seasonal patterns of habitat partitioning in a guild of southern California tidepool fishes. Marine Ecology Progress Series, 196: 253-268. [ Links ]
Dominici-Arosemena A. & Wolff, M. (2006). Reef fish community structure in the Tropical eastern Pacific (Panamá): living on a relatively stable rocky reef environment. Helgoland Marine Research, 60: 287-305. [ Links ]
Floeter S. R., Rocha, L. A., Robertson, D. R., Joyeux, J. C., Smith-Vaniz, W. F., Wirtz, P., Edwards, A. J., Barreiros, J. P., Ferreira, C. E. L., Gasparini, J. L., Brito, A., Falcón, J. M., Bowen, B., & Bernardi, G. (2008). Atlantic reef fish biogeography and evolution. Journal of Biogeography, 35: 22-47. [ Links ]
Gibson, R. N. (1988). Patterns of movement in intertidal fishes. In G. Chelazzi & M. Vannini (Eds.), Behavioural Adaptation to Intertidal Life (pp. 55-63). Plenum Press, New York, USA. [ Links ]
Gibson, R. N. (1992). Tidally synchronized bahaviuor in marine fishes. In M. A. Ali (Ed.), Rhythms in Fishes (pp. 55-63). Plenum Press, New York. USA. [ Links ]
Gibson, R. N. (1993). Intertidal teleosts: life in a fluctuating environment,. In T. J. Pitcher (Ed.), Behaviour of Teleost Fishes (pp. 513-536). Chapman & Hall, London, United Kingdom. [ Links ]
Gibson, R. N. (2003). Go with the flow: tidal migration in marine animals. Hydrobiologia 503: 153-161. [ Links ]
Gibson, R. N. & Yoshiyama, R. M. (1999). Intertidal fish communities. In M. H. Horn, K. L. M. Martin & M. A. Chotkowski (Eds.), Intertidal Fishes: Life in two worlds (pp. 264-296) Academic Press, London, United Kingdom. [ Links ]
Glynn, P. W. & Ault, J. S. (2000). A biogeographic analysis and review of the far eastern Pacific coral reef region. Coral Reefs, 19: 1-23. [ Links ]
Griffiths, S. P. (2000). The use of clove oil as an anaesthetic and method for sampling intertidal rockpool fishes. Journal of Fish Biology, 57: 1453-1464. [ Links ]
Griffiths, S. P. (2003). Rockpool ichthyofaunas of temperate Australia: species composition, residency and biogeographic patterns. Estuarine, Coastal and Shelf Science, 58: 173-186. [ Links ]
Griffiths, S. P., Davis, A. R. & West, R. J. (2006). Role of habitat complexity in structuring temperate rockpool ichthyofaunas. Marine Ecology Progress Series, 313: 227-239. [ Links ]
Grossman, G. D. (1982). Dynamics and organization of a rocky intertidal fish assemblage: the persistence and resilience of taxocene structure. American Naturalist, 119: 611-637. [ Links ]
Hastings, P. A. (2009). Biogeography of New World blennies. In R. Patzner, E. Gonçalves, P. Hastings & B. Kapoor (Eds.), The Biology of Blennies (pp. 95-118). Science Publishers, Enfield, New Hampshire, United Kingdom. [ Links ]
Horn, M. H., Martin, K. L. M. & Chotkowski, M. A. (1999). Introduction. In M. H. Horn, K. L. M. Martin & M. A. Chotkowski (Eds.), Intertidal Fishes: Life in two worlds (pp.1-6). Academic Press, London, United Kingdom. [ Links ]
Little, C. & Kitching, J. A. (1996). The biology of rocky shores. Oxford University Press, Oxford, United Kingdom. [ Links ]
López-Victoria, M., Cantera, J. R., Díaz, J. M., Rozo, D. M., Posada, B. O. & Osorno, A. (2004). Estado de los litorales rocosos en Colombia: acantilados y playas rocosas. In INVEMAR (Ed.), Informe del estado de los ambientes marinos y costeros en Colombia: año 2003 (pp. 171-182). INVEMAR, Santa Marta, Colombia.
Macieira, R. M. & Joyeux, J. C. (2011). Distribution patterns of tidepool fishes on a tropical flat reef. Fishery Bulletin, 109: 305-315. [ Links ]
Mahon, R. & Mahon, S. D. (1994). Structure and resilience of a tidepool fish assemblage at Barbados. Environmental Biology of Fishes, 41: 171-190. [ Links ]
Martins, G. M., Hawkins, S. J., Thompson, R. C. & Jenkins, S. R. (2007). Community structure and functioning in intertidal rock pools: effects of pool size and shore height at different successional stages. Marine Ecology Progress Series, 329: 43-57. [ Links ]
Metaxas, A. & Scheibling, R. E. (1993). Community structure and organization of tidepools. Marine Ecology Progress Series, 98: 187-198. [ Links ]
Mok, H. & Wen, P. (1985). Intertidal fish community ecology on Lu Tao Island (Green Island), Taiwan. Journal of the Taiwan Museum, 38: 81-118. [ Links ]
Mora, C. & Robertson, D. R. (2005). Factors shaping the range-size frequency distribution of the endemic fish fauna of the Tropical Eastern Pacific. Journal of Biogeography, 32: 277-286. [ Links ]
Muñoz, A. A. & Ojeda, F. P. (1997). Feeding guild structure of a rocky intertidal fish assemblage in Central Chile. Environmental Biology of Fishes, 49: 471-479. [ Links ]
Nielsen, K. J. (2001). Bottom-up and Top-down forces in tide pools: test of a food chain model in an intertidal community. Ecological Monographs, 71: 187-217. [ Links ]
Palmer, J. D. (2000). The clocks controlling the tide-associated rhythms of intertidal animals. BioEssays, 22: 32-37. [ Links ]
Prochazka, K. (1996). Seasonal patterns in a temperate intertidal fish community on the west coast of South Africa. Environmental Biology of Fishes, 45: 133-140. [ Links ]
Prochazka, K., Chotkowski, M. A. & Buth, D. G. (1999). Biogeography of intertidal Fishes. In M. H. Horn, K. L. M. Martin & M. A. Chotkowski (Eds.), Intertidal Fishes: Life in two worlds (pp. 332-355). Academic Press, London, United Kingdom. [ Links ]
Quijada, P. A. & Caceres, C. W. (2000). Patrones de abundancia, composición trófica y distribución especial del ensamble de peces intermareales de la zona centro-sur de Chile. Revista Chilena de Historia Natural, 73: 739-747. [ Links ]
Robertson, D. R. (1998). Do coral-reef fish faunas have a distinctive taxonomic structure? Coral Reefs, 17:179-186. [ Links ]
Robertson, D. R. & Allen, G. (2002). Shorefishes of the Tropical Eastern Pacific: an information system. CD-ROM. Smithsonian Tropical Research Institute, Balboa, Panamá [ Links ].
Robertson, D. R., Grove, J. S. & McCosker, J. E. (2004). Tropical Transpacific shorefishes. Pacific Science, 58: 507-565. [ Links ]
Robertson, D. R. & Smith-Vaniz, W. F. (2008). Rotenone: an essential but demonized tool for assessing marine fish diversity. BioScience, 58: 165-170. [ Links ]
Robertson, D. R. & Cramer, K. L. (2009). Shore fishes and biogeographic subdivisions of the tropical eastern Pacific. Marine Ecology Progress Series, 380: 1-17. [ Links ]
Ruiz-Campos, G., González-Guzmán, S., Ramírez-Valdéz, A., González-Acosta, A. F., Castro-Aguirre, J. L. & De La Cruz-Agüero, J. (2010). Composition, density and biogeographic affinities of the rocky intertidal fishes on the western coast of the Baja California peninsula, Mexico. California Cooperative Oceanic Fisheries Investigations, 51: 210-220. [ Links ]
Thomson, D. A. & Lehner, C. E. (1976). Resilience of a rocky intertidal fish community in a physically unstable environment. Journal of Experimental Marine Biology and Ecology, 22: 1-29. [ Links ]
Weaver, P. L. (1970). Species diversity and ecology of tidepool fishes in three Pacific coastal areas of Costa Rica. Revista de Biología Tropical, 17: 165-185. [ Links ]
Willis, T. J. & Roberts, C. D. (1996). Recolonisation and recruitment of fishes to intertidal rockpools at Wellington, New Zealand. Environmental Biology of Fishes, 47: 329-343. [ Links ]
Yoshiyama, R. M. (1981). Distribution and abundance patterns of rocky intertidal fishes in Central California. Environmental Biology of Fishes, 6: 315-332. [ Links ]
Yoshiyama, R. M., Sassaman, C. & Lea, R. N. (1986). Rocky intertidal fish communities of California: temporal and spatial variation. Environmental Biology of Fishes, 17: 23-40. [ Links ]
Zapata, F. A. & Morales, Y. (1997). Spatial and temporal patterns of fish diversity in a coral reef at Gorgona Island. Proceedings of the 8th International Coral Reef Symposium, 1: 1029-1034. [ Links ]
Zapata, F. A. (2001a). Formaciones coralinas de Isla Gorgona. In L. M. Barrios & M. Lopez-Victoria (Eds.), Gorgona Marina: contribución al conocimiento de una isla única (pp. 27-50). INVEMAR, Serie Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
Zapata, F. A. (2001b). Peces Marinos de Gorgona: Genenralidades. In L. M. Barrios & M. Lopez-Victoria (Eds.), Gorgona Marina: contribución al conocimiento de una isla única (pp. 107-109). INVEMAR, Serie Publicaciones Especiales No. 7, Santa Marta, Colombia. [ Links ]
1. Leibniz Center for Tropical Marine Ecology (ZMT). Fahrenheitstr. 6, 28359 Bremen, Germany; gustavoa80@yahoo.com. Grupo de Investigación en Ecología Animal, Departamento de Biología, Universidad del Valle. A.A. 25360, Cali, Colombia. Centre of Excellence in Marine Sciences (CEMarin), Cra. 2 No. 11-68, Rodadero, Santa Marta, Colombia.
2. Grupo de Investigación en Ecología Animal, Departamento de Biología, Universidad del Valle. A.A. 25360, Cali, Colombia; agiraldo@correounivalle.edu.co; ecologia@univalle.edu.co
3. Grupo de Investigación en Ecología de Arrecifes Coralinos, Departamento de Biología, Universidad del Valle. A.A. 25360, Cali, Colombia; fernando.zapata@correounivalle.edu.co
Recibido 18-X-2013. Corregido 20-XI-2013. Aceptado 19-XII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}