










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.1 San José Feb. 2014
A comparison of visual and collection-based methods for assessing community structure of coral reef fishes in the Tropical Eastern Pacific
Abstract
Gorgona Island, the major insular area in the Colombian Pacific Ocean, is characterized by a remarkably high biological and ecosystem diversity for this area of the world. Coral reefs are well developed and their fish communities have been described using conventional visual surveys. These methods, however, are known to be biased towards detecting larger and more mobile species, tending to ignore small and cryptobenthic species. The two main objectives of this study were to describe the assemblage structure of the cryptobenthic fish fauna and estimate the extent to which this fauna is underestimated by visual surveys.At the beginning and the end of the warm season, we compared the cryptobenthic fish assemblage recorded using visual surveys against the one recorded using “enclosed anesthetic/rotenone samples” on isolated coral colonies (N=54 beginning of warm season; N=17 end of warm season). The crypthobenthic fish fauna associated to coral colonies was characterized by small body sizes and was composed mainly by species of the families Antennaridae, Blennidae, Gobiidae, Labrisomidae, Muraenidae, Serranidae, Scorpaenidae and Syngnathidae. Conventional visual surveys underestimated overall species richness by 28-36% and number of individuals by 16-35%. Noteworthy, four species recorded during this study using “enclosed anesthetic/rotenone samples” were new records for Gorgona Island. Although both sampling methods can detect a largely overlapping group of species, the “enclosed anesthetic/rotenone samples” method was able to detect more individuals and species, including several species that visual surveys fail to detect. Although this study is the first effort to describe the cryptobenthic fish assemblage associated to coral reefs in the Tropical Eastern Pacific, our results suggest that these assemblages are an important component of the reef fish community in the region in terms of biodiversity and functional roles. Rev. Biol. Trop. 62 (Suppl. 1): 359-371. Epub 2014 February 01.
Key words: cryptobenthic fishes, coral reefs, sampling methods, visual surveys, enclosed anesthetic/rotenone samples.
Resumen
Métodos de muestreo convencionales (censos visuales) usados en el estudio de peces arrecifales han llevado a una constante subestimación de la riqueza de especies, ya que estos tienden a ignorar especies pequeñas y crípticas. En este estudio describimos la estructura del ensamblaje de peces criptobentónicos en Isla Gorgona, Colombia y estimamos el grado en que este ensamblaje es subestimado al usar censos visuales. Al comienzo y al final de la estación cálida comparamos la capacidad de detección de censos visuales vs “muestreos cerrados con anestésico/rotenona” (N=54 comienzo; N=17 final). La fauna criptobentónica se caracterizó por mantener individuos de tamaños corporales pequeños, pertenecientes principalmente a las familias Antennaridae, Blennidae, Gobiidae, Labrisomidae, Muraenidae, Serranidae, Scorpaenidae and Syngnathidae. Los censos visuales subestimaron la riqueza de especies en un 28-36% y el número de individuos en un 16-35%. Aunque un gran número de especies son detectadas usando ambos métodos, los “muestreos cerrados con anestésico/rotenona” detectan un mayor rango de especies. Este estudio, el primer esfuerzo para describir el ensamblaje de peces criptobentónicos asociados a arrecifes coralinos en Colombia, sugiere que la fauna criptobentónica puede ser un componente importante de la comunidad arrecifal en términos de biodiversidad y papel funcional.
Palabras clave: peces criptobentónicos, arrecifes de coral, métodos de muestreo, encuestas visuales, muestras de anestesia/rotenona.
Non-destructive sampling techniques, such as visual surveys, have been widely used in studies of coral reef fish ecology ever since their introduction by Brock (1954). However, these techniques have several sources of error due to fish mobility, observer experience and transect size (Thompson & Mapstone, 1997; Ackerman & Bellwood, 2000; Palacios & Zapata, 2011). This leads to a sampling bias toward larger and more mobile species, tending to underestimate small and cryptobenthic fishes. Although it has been suggested that consistent errors and biases in the visual estimates might be acceptable in comparative studies (Brock, 1982; Ackerman & Bellwood, 2000; Willis, 2001), underestimating the cryptobenthic component of reef fish communities might have a potentially larger impact on the understanding of their community assembly, trophic ecology and ultimately on their conservation (Mora, Titterson & Myers, 2008). As a response to this, the number of studies on the ecology of cryptobenthic fish communities is growing (Depczynski & Bellwood, 2003; Kovacic, Patzner & Schliewen, 2012; Harborne, Jelks, Smith-Vaniz & Rocha, 2012).
In comparison with the Caribbean and the Indo-Pacific regions, the number of studies describing reef fish communities in the Tropical Eastern Pacific (TEP) is relatively scarce (e.g., Pérez-España, Galván-Magaña & Abitia-Cárdenas, 1996; Zapata & Morales, 1997; Aburto-Oropeza & Balart, 2001; Espinoza & Salas, 2005; Alvarez-Filip, Reyes-Bonilla & Calderon-Aguilera, 2006; Dominici-Arosemena & Wolff, 2006; Benfield, Baxter, Guzman & Mair, 2008; Galván-Villa, Arreola-Robles, Ríos-Jara & Rodriguez-Zaragoza, 2010). Moreover, all of these studies have been based on visual surveys, which are known to lead to an underestimation of community richness and population densities (Brock, 1982, Bellwood & Alcala, 1988). Although there are several studies based on captures as sampling method, their goal was to generate inventories and preliminary zoogeographic analyses (Prahl, 1986; Rubio, 1990) or to study early life-history (Mora & Zapata, 2000; Lozano & Zapata, 2003). Therefore a complete study of the reef fish community is still lacking, which is key as a basis for ecological research, management and conservation plans.
This study is the first to quantitatively examine the cryptobenthic fish component of a coral reef community in the Tropical Eastern Pacific. We accomplished this by enclosing isolated coral colonies and thoroughly sampling their fish communities with an anesthetic/rotenone mixture. A comparison between traditional visual surveys against the “enclosed anesthetic/rotenone samples” method allowed us, besides studying the structure of the cryptobenthic fish assemblages, to estimate the extent to which visual surveys underestimate this component of the reef fish community. Our study ultimately contributes to gain more complete understanding of reef fish community structure in the region.
Materials and methods
Study area: Gorgona (2°59’ N -7 8°12’ W) is a continental island located 35km off the Pacific coast of Colombia. Here, coral reefs are small (~10 ha) and shallow (<10 m) and Despite of their modest development, coral reefs in Gorgona Island are the best developed reefs of the Colombian Pacific, comparable to other well developed reefs in the TEP (Glynn, Prahl & Guhl, 1982; Zapata, 2001).
Field work was carried out during September-October of 2007 and during April-May of 2008. These two sampling periods covered the end and the beginning of the yearly warm season. Temporal patterns of sea surface temperature at Gorgona Island are characterized by a cold and high salinity season between January and April (Giraldo, Rodríguez-Rubio & Zapata, 2008), followed by an increase in temperature and decrease in salinity in April (beginning of the warm season), followed by a gradual reversal in temperature and salinity conditions until December (Zapata, 2001; Giraldo et al., 2008). These temporal climatic patterns have been shown to have an effect on the coral reef fish community (Zapata & Morales, 1997) due to an increase in recruitment during warmer periods as it has been the case in other regions (Sale & Dybdahl, 1975; Talbot, Russel & Anderson, 1978; Bohnsack & Talbot, 1980; Sale & Douglas, 1984; Coles & Tarr, 1990).
The field surveys were carried out at six study sites (“La Azufrada 1”, “La Azufrada 2”, “Yundigua”, “El Muelle”, “El Arrecifito” and “Playa Blanca”) on the eastern part of Gorgona Island.
Collection: To examine differences in the description of community structure based on data obtained through different sampling methods, quantitative sampling of the fish community associated to isolated coral colonies was carried out using visual surveys and “enclosed anesthetic/rotenone samples”. Anesthetic clove oil and rotenone have been the most used and effective substances for sampling fishes (Robertson & Smith-Vaniz, 2008; Robertson & Smith-Vaniz, 2010). Therefore, here we combined both clove oil and rotenone in a single sampling method. In total 71 isolated coral colonies were sampled, 54 during 2007 and 17 during 2008. Out of the 54 colonies sampled during 2007, 28 colonies were branching corals (Pocillopora capitata, Pocillopora damicornis, Pocillopora eydoxi) and 26 were massive corals (Pavona clavus, Pavona varians, Pavona gigantea, Pavona chiriquiensis, Pavona frondifera, Gardineroseris planulata). Out of the 17 colonies sampled during 2008, six were massive corals (Gardineroseris planulata, Pavona gigantea, Pavona clavus) and 11 were branching corals (Pocillopora capitata). In total 20 colonies were sampled at “La Azufrada 1”, four at “La Azufrada 2”, 18 at “Yundigua”, 18 at “El Muelle”, eight at “El Arrecifito” and three at “Playa Blanca”.
All selected colonies had a total volume <1m3 (available volume per colony: 3 000-7 700cm3 for massive corals and 7000-15 000cm3 for branching corals) and were sufficiently isolated from other colonies to allow manipulation. The procedure consisted of two parts: first, we carried out a meticulous visual survey of the fish fauna during 5 minutes. Second, we enclosed the colony using a 1m3 cage (a PVC-plastic structure covered by plastic netting with lateral openings for manipulation) and added a Clove oil –rotenone– 95% ethanol solution (21%:9%:70% respectively). We allowed the solution to act during 5min and then we collected all individuals during 15-20min. Finally, all individuals were measured, weighed and identified following mainly Robertson & Allen (2006).
Data analysis: The cryptobenthic fish assemblage was described in terms of species richness and number of families and individuals for every period and sampling method. We examined differences between sampling methods in number of individuals for every species using paired t-tests.
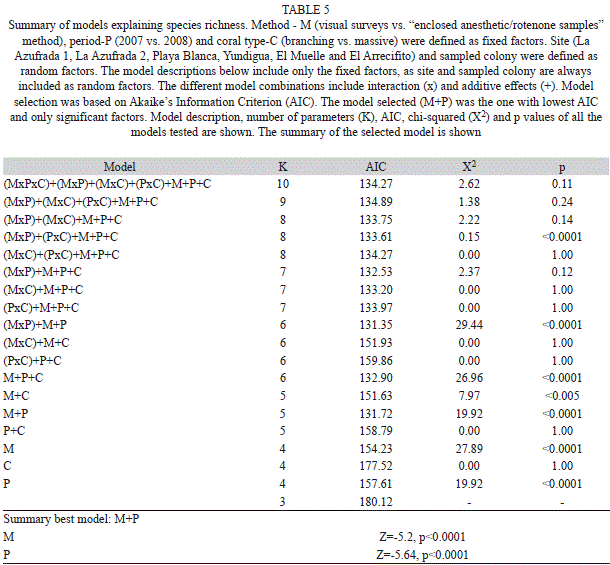
We used generalized linear mixed models with a Poisson error structure to explain species richness. The models included three fixed factors: sampling method (visual vs. capture), period (2007 vs. 2008), and coral type (branching vs. massive coral), and two random factors: study site (6 study sites) and colony. In total we compared 19 models. The most saturated model included method, period and coral type as fixed factors, all their possible interaction effects and the two random effects (site and colony nested within site). The simplest model was an empty model including only the two random effects. We used a step-wise procedure that removed factors until the most parsimonious (lowest Akaike’s Information Criterion (AIC) value) model was obtained (Johnson & Omland, 2004). The model selected was the one with only significant factors and lowest AIC unless a simpler model differed by <2 AIC points (Burnham & Anderson, 2002). The assumption that model residuals were Poisson distributed was met (X2=83.44, df=137, p=0.99). Although the available volume per colony (the space available for fish shelter) showed wide variation, this factor was not included in the analyses because (1) this variable was only estimated for the first sampling period, and (2) colony volume did not affect number of species (massive corals: r2=0.004, F(1.21)=0.08, p=0.77; branching corals: r2=0.094, F(1.25)= 2.6, p=0.12) and individuals (massive corals: r2=0.004, F(1,21)=0.09, p=0.77; branching corals: r2=0.05, F(1.25)=1.35, p=0.25) occurring in the coral colonies.
To study whether visual surveys and “enclosed anesthetic/rotenone samples” can detect a similar number of species, a simple correlation between species richness detected by visual surveys and species richness detected by “enclosed anesthetic/rotenone samples” was carried out. The same analysis was carried out for the total abundance detected by the two sampling methods. Chromis atrilobata was excluded from these analyses because its schooling behavior and high abundance made it act as an outlier. Square root transformation of the abundance data was performed in order to meet the assumption of normality of the model residuals (Shapiro Test). We also examined whether the slopes of the correlation lines differed from 1 and whether the intercepts differed from 0 to better assess the differences between the two methods.
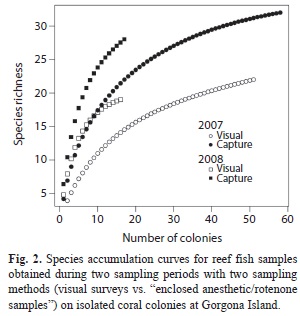
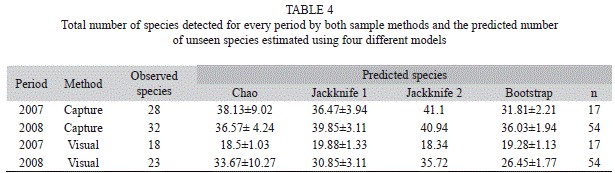
To examine how many species can be detected with different sampling methods when increasing sample size, we used species accumulation curves (SAC) using Kindt’s exact accumulator method (Oksanen et al., 2010). Additionally, we studied whether differences in sampling methods and periods affect the predictions of the total number of expected species for different species richness estimators: Chao, Jackknife 1, Jackknife 2 and Bootstrap (Oksanen et al., 2010).
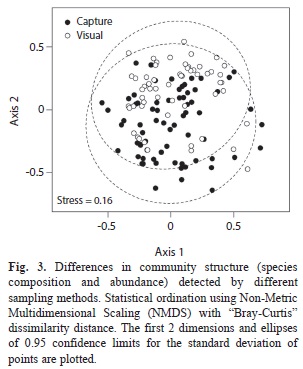
Finally, to study how community structure differs with the sampling method used, the sampling period and coral type, a Non-Metric Multidimensional Scaling (NMDS) was used (Quinn & Keough, 2002). The matrix of dissimilarities was calculated using the Bray-Curtis distance measure. The “goodness-of-fit” of the final configuration was assessed by a Kruskal’s stress (Quinn & Keough, 2002). To find the global solution we used 100 random starts, thus avoiding local optima. We used the lowest number of dimensions possible (4) for which final solutions were more stable and with an acceptable stress value (<0.2). To test whether the observed community structure in each coral colony changes with the sampling method used, a Permutational Multivariate Analysis of Variance (PERMANOVA) was applied (Anderson, 2001; Oksanen et al., 2010). The matrix of dissimilarities was calculated using the Bray-Curtis distance measure. The distance matrix was based on 1000 permutations of the data. Colony was set as a random factor. Three of the 71 colonies were excluded from the analysis because visually no individuals were detected, making the calculation of Bray-Curtis differences impossible. These colonies without observed species were massive corals, sampled in 2007, two of them from “La Azufrada 1” and one from “El Muelle”.
All analyses were performed using R-2.11.1 (R. Development Core Team, 2010) and the R packages: vegan (Oksanen et al., 2010), ecodist (Goslee & Urban, 2007), ellipse (Murdoch & Chow, 2007), lme4 (Bates, Maechler & Bolker, 2013) and plotrix (Lemon, 2006).
Results
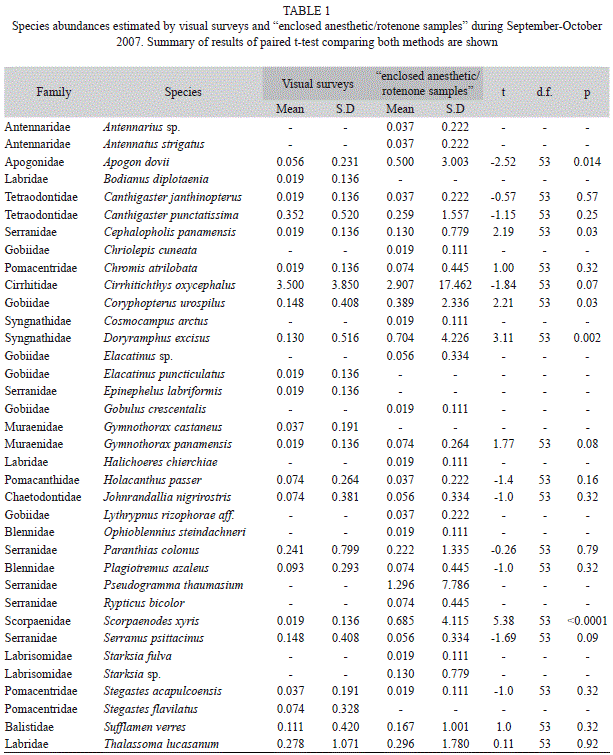
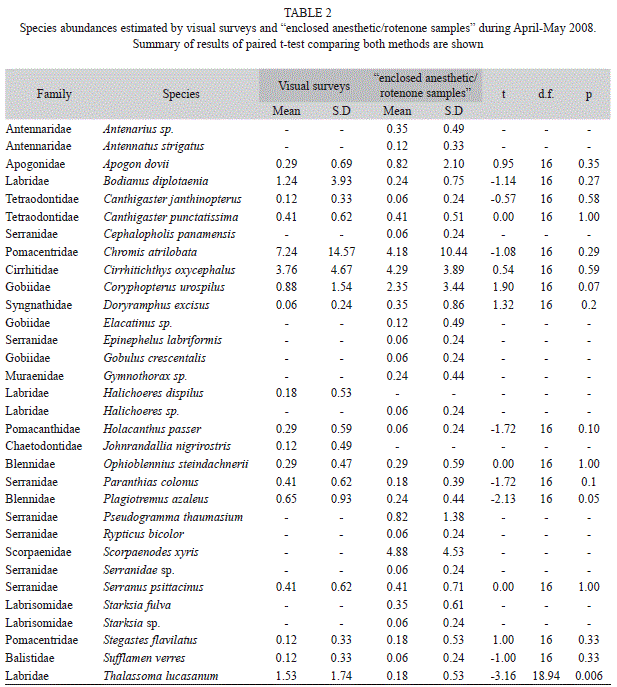
Visual surveys in general detected fewer species and total number of individuals than “enclosed anesthetic/rotenone samples”. In 2007, 365 individuals of 28 species were recorded using direct capture and 308 individuals of 18 species were recorded using visual surveys. Enclosed anesthetic/rotenone sampling detected more species and more individuals than visual surveys in 90.7% and 85.2% of colonies, respectively. At this time 13 species were only detected by “enclosed anesthetic/rotenone samples”, five were detected only by visual surveys and 18 species were recorded using both sampling methods; visual censuses significantly underestimated the abundance of five of the latter species (Table 1). During 2008, 457 individuals of 32 species were recorded using “enclosed anesthetic/rotenone samples” and 297 individuals of 23 species were recorded using visual surveys. Enclosed anesthetic/rotenone sampling detected more species and more individuals than visual surveys in 94.1% and 92.9% of colonies, respectively. During this period, 14 species were detected only by “enclosed anesthetic/rotenone samples”, two were detected only by visual surveys and 16 species were recorded by both sampling methods. Thalassoma lucasanum, showed higher abundance using visual surveys and Plagiotremus azaleus was marginally more abundant using visual surveys (Table 2).
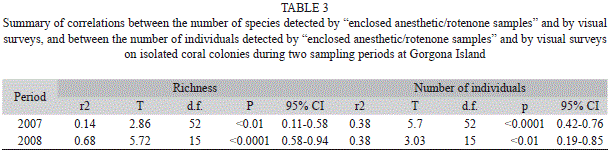
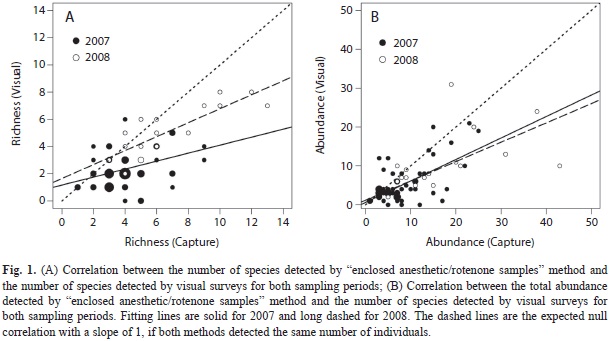
The number of species detected visually was positively correlated with the number of species detected by capture for both sampling periods (Table 3). However, visual surveys consistently underestimated the number of species; the slope of the relation was significantly lower than 1 for both sampling periods (Fig. 1A, Table 3). Similarly, although the total number of individuals detected visually was positively correlated with the total number of individuals detected by capture for both sampling periods (Fig. 1B, Table 3), the slope of the relation was significantly lower than 1.
In both periods, “enclosed anesthetic/rotenone samples” detected more species than visual surveys for the same sampling effort (Fig. 2). Models to estimate the number of expected species always predicted more species when analyzing the data obtained by “enclosed anesthetic/rotenone samples” than by visual surveys. These differences were 39.4 to 55.4% in 2007 and 8 to 16.5% in 2008 (Table 4).
The variability in species richness was primarily explained by period and sampling method, and secondarily by site and colony (Table 5). The average number of observed species was higher when using “enclosed anesthetic/rotenone samples” than when using visual surveys (4.79±0.03 vs. 3.04±0.03) and it was higher during 2008 than during 2007 (5.94±0.08 vs. 3.28±0.02).
Although the species composition detected by visual surveys and “enclosed anesthetic/rotenone samples” overlapped, “enclosed anesthetic/rotenone samples” detected a different component of the community y than visual surveys (Fig. 3). Patterns of community structure differed depending on the method used for sampling the community (PERMANOVA, R2=0.077, F(1, 137)=11.36, p=0.0009).
Discussion
The crypthobenthic fish fauna associated to isolated coral colonies at Gorgona Island was composed mainly by small species with secretive habits or cryptic coloration of the families Antennaridae, Blennidae, Gobiidae, Labrisomidae, Muraenidae, Serranidae, Scorpaenidae and Syngnathidae. Our results strongly suggest that conventional visual surveys underestimate both the species richness and abundance of this assemblage.
Even though our results show that the number of species and individuals detected by both techniques are positively correlated, analyses of these correlations show that visual surveys are not as effective as the “enclosed anesthetic/rotenone samples” method. Here, we showed that visual surveys detect less individuals and species than the “enclosed anesthetic/rotenone samples” method. Furthermore, we showed that the observed pattern of community structure depends on the type of sampling method used. Although both sampling techniques detect an overlapping group of species, each technique can also detect an exclusive group. However, the “enclosed anesthetic/rotenone samples” method can detect more species and more individuals. This has also been reported for reef fishes in the Bahamas when using rotenone collections vs. visual surveys (Smith & Tyler, 1972). Fishes whose home ranges are a little larger than the reef patch studied and that are not particularly cryptic can be equally recorded by both methods. Small and cryptic fishes are underestimated when using visual surveys and larger fishes with schooling behaviour that live in mid-water are usually underestimated using rotenone collections. This suggests that both sampling techniques are complementary in the sense that they are able to detect a different component of the community.
The percentage of underestimation reported in our study (28-36% of the species richness and between 16-35% of the number of individuals) is similar to the one reported at Orpheus Island in the Great Barrier Reef, Australia (37.5%, Ackerman & Bellwood, 2000) and at Kaneohe Bay, Oahu, Hawaii (34.6%, Brock, 1982). More extreme underestimations have been reported for habitat types different than coral reefs in the northern coast of New Zealand (82% richness and 73% number of individuals, Willis, 2001). The results from that study suggest a reduction in detection capability of visual censuses with an increase of habitat complexity. Therefore, how accurately visual censuses can estimate species richness will depend on how species-rich and habitat-complex the study area is.
Although samples based on captures can detect a higher number of species, a complete census is never obtained (Smith, 1973). For instance, in the Bahamas, Smith (1973) estimated that a single rotenone sampling can detect only 75% of the total number of species in a single reef patch. However, this estimation could have been biased in different ways. Smith (1973) sampled a single patch several times during 1 to 5 days and assumed that the cumulative fish fauna detected during this period was 100% of the community. However, sampled colonies were not enclosed, therefore, mobile fishes are able to escape the rotenone cloud before it has any effect and new individuals are able to colonize the patch after the rotenone cloud is diluted.
The larger number of species and individuals recorded in 2008 is consistent with the seasonal pattern of temporal variation in richness exhibited by the whole coral reef fish community at “La Azufrada” coral reef at Gorgona Island (Zapata & Morales, 1997). A peak of species richness was reported between April and June, months where the sea surface temperature is normally higher (28-30°C). Seasonality in species richness has also been reported for other localities both for natural and artificial reefs and has been associated with an increase in recruitment during warmer periods (Sale & Dybdahl, 1975; Talbot et al., 1978; Bohnsack & Talbot, 1980; Sale & Douglas, 1984; Coles & Tarr, 1990).
Our results show that the underestimation related to sampling method cannot be surmounted by using functions that estimate species richness by extrapolation. This gives the warning that predictions of total species richness when data are obtained by sampling methods with different detection capabilities should be used with caution. For 2007, the predicted number of species based on visual surveys is in the range of the number of species that were captured and only slightly lower that the number of species predicted based on “enclosed anesthetic/rotenone samples”. The estimators Jackknife 1 and Jackknife 2 were the best estimators for the warm season. A lack of convergence of the predicted number of species for 2008 can be caused by the small sample size (N=17) in comparison with 2007 (N=54). It has been shown that these estimators can be very sensitive to sample size (Herzog, Kessler & Cahill, 2002), as small sample size can inflate the number of singletons and doubletons recorded.
It has been estimated that current taxonomic inventories of fish faunas associated with reefs lack about 18% of the complete species inventory (Mora et al., 2008). Particularly species that attain small body sizes have been underestimated in the TEP, and possible undiscovered species might belong to this group of species (Zapata & Robertson, 2007). During this study we found four new records of species for Gorgona Island (Chriolepis cuneata, Gobulus crescentalis, Cosmocampus arctus, Canthigaster janthinoptera), three of which are secretive or cryptic and represent southward range extensions in the TEP (Alzate, Muñoz, Zapata & Giraldo, 2012). Therefore, our study highlights the importance of using clove oil or rotenone to effectively sample the cryptobenthic fish community on coral reefs, which allows a better description of the community structure and more accurate estimates of the distribution, abundance and diversity patterns of the whole reef fish community. However, sampling methods such as the one used in this study are increasingly disfavored, mainly due to difficulties in the obtention of permits for collecting specimens in natural reserves and because anesthetics like clove oil and poisons like rotenone have been unfairly demonized by managers (Robertson & Smith-Vaniz, 2008; 2010). This brings substantial flaws in description of diversity patterns development of conservation strategies because studies are generally based on incomplete and inaccurate inventories (Dubois, 2003; Mace, 2004; Morrison, Marcot & Mannan, 2006; Mora et al., 2008). As a consequence conservation actions can be inadequate, excluding habitats and specific taxa (Rojas, 1992; Dubois, 2003; Khuroo, Dar, Khan & Malik, 2007). Reef fish populations are highly resilient and can satisfactorily withstand natural disturbances (Robertson & Smith-Vaniz, 2008). After intermediate scale perturbations (e.g. hurricanes) populations recover quickly and even faster after small scale perturbations (e.g. captures, Smith, 1973; Robertson & Smith-Vaniz, 2008). Therefore, the use of clove oil or rotenone for adequately sampling reef fish communities should be reconsidered, as it might be useful as a complementary method to visual surveys when used with caution.
Our study represents an important step towards a complete understanding of the reef fish community structure at Gorgona Island in particular and the eastern Pacific in general. This type of study must be extended to other areas and habitat types in order to help complete taxonomic inventories. Once a community is fully known, further efforts can be directed to understand its trophic structure and assembly rules.
Acknowledgments
We thank J. C. Benítez, S. Cuevas and C. G. Muñoz for help during field work. We also thank F. van der Plas, D. R. Robertson and an anonymous reviewer for helpful comments on previous versions of the manuscript. The Unidad Administrativa Especial del Sistema Nacional de Parques Naturales (UAESPNN) provided permits for this research (DTSO-G-23 08/2007). This study was financially supported by grant No. 2063 from Fundación para la Promoción de la Investigación y la Tecnología del Banco de la República de Colombia to A. Giraldo.
References
Aburto-Oropeza, O. & Balart, E. F. (2001). Community structure of reef fish in several habitats of a rocky reef in the Gulf of California. Marine Ecology Progress Series, 22: 283-305. [ Links ]
Ackerman, J. & Bellwood, D. (2000). Reef fish assemblages: a re-evaluation using enclosed rotenone stations. Marine Ecology Progress Series, 206: 227-237. [ Links ]
Alvarez-Filip, L., Reyes-Bonilla, H. & Calderon-Aguilera, L. E. (2006). Community structure of fishes in Cabo Pulmo Reef, Gulf of California. Marine Ecology Progress Series, 27: 253-262. [ Links ]
Alzate, A., Muñoz, C. G., Zapata, F. A. & Giraldo, A. (2012). New records of cryptobenthic fishes in coral reef habitats of Gorgona Islands, Colombia, Tropical Eastern Pacific. Boletín de Investigaciones Marinas y Costeras, 41: 229-235. [ Links ]
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Australian Journal of Ecology, 26: 32-46. [ Links ]
Bates, D., Maechler, M. & Bolker, B. (2013). Lme4: linear mixed-effects models using S4 classes. R. package version 0.999999-2. [ Links ]
Bellwood, D. R. & Alcala, A. C. (1988). The effects of a minimum length specification on visual estimates of density and biomass of coral reef fishes. Coral Reefs, 7: 23-27. [ Links ]
Benfield, S., Baxter, L., Guzman, H. M. & Mair, J. M. (2008). A comparison of coral reef and coral communities fish assemblages in Pacific Panama and environmental factors governing their structure. Journal of the Marine Biological Association of the United Kingdom, 88: 1331-1341. [ Links ]
Bohnsack, J. A. & Talbot, F. H. (1980). Species packing by reef fishes on Australian and Caribbean reefs: an experimental approach. Bulletin of Marine Science, 30: 710-723. [ Links ]
Brock, V. E. (1954). A preliminary report on a method of estimating reef fish populations. Journal of Wildlife Management, 18: 297-308. [ Links ]
Brock. R. E. (1982). A critique of the visual census method for assessing coral reef fish populations. Bulletin of Marine Science, 32: 269-276. [ Links ]
Burnham, K. P. & Anderson, D. R. (2002). Model selection and multimodel inference: a practical information-theoretical approach. Springer-Verlag, New York, USA. [ Links ]
Coles, S. L. & Tarr, A. B. (1990). Reef fish assemblages in the western Arabian Gulf: a geographically isolated population in an extreme environment. Bulletin of Marine Science, 47: 696-720. [ Links ]
Depczynski, M. & Bellwood, D. R. (2003). The role of cryptobenthic reef fishes in coral reef trophodynamics. Marine Ecology Progress Series, 256: 183-191. [ Links ]
Dominici-Arosemena, A. & Wolff, M. (2006). Reef fish community structure in the Tropical Eastern Pacific (Panamá): living on a relatively stable rocky reef environment. Helgoland Journal of Marine Research, 60: 287-305. [ Links ]
Dubois, A. (2003). The relationships between taxonomy and conservation biology in the century of extinctions. Comptes Rendus Biologies, 326: s9-s21. [ Links ]
Espinoza, M. & Salas, E. (2005). Estructura de las comunidades de peces de arrecife en las Islas Catalinas y Playa Ocotal, Pacífico norte de Costa Rica. Revista de Biología Tropical, 53: 523-536. [ Links ]
Galván-Villa, C. M., Arreola-Robles, J. L., Ríos-Jara, E. & Rodriguez-Zaragoza, F. A. (2010). Ensamblajes de peces arrecifales y su relación con el habitat bentónico de la isla Isabel, Nmayarit, México. Revista de Biología Marina y Oceanografía, 45: 311-324. [ Links ]
Giraldo, A., Rodríguez-Rubio, E. & Zapata, F. A. (2008). Condiciones oceanográficas en Isla Gorgona, Pacífico Oriental Tropical de Colombia. Latin American Journal of Aquatic Research, 36: 121-128. [ Links ]
Glynn, P. W., Prahl, H. v. & Guhl, F. (1982). Coral reef of Gorgona Island, Colombia, with special reference to corallivores and the influence on community structural reef development. Anales del Instituto de Investigaciones Marinas de Punta de Betín, 12: 185-214. [ Links ]
Goslee, S. C. & Urban, D. L. (2007). The ecodist package for dissimilarity-based analysis of ecological data. Journal of Statistical Software, 22:1-19. [ Links ]
Harborne, A. R., Jelks, H. L., Smith-Vaniz, W. F. & Rocha, L. A. (2012). Abiotic and biotic controls of cryptobenthic fish assemblages across a Caribbean seascape. Coral Reefs, 31: 977-990. [ Links ]
Herzog, S. K., Kessler, M. & Cahill, T. M. (2002). Estimating species richness of tropical bird communities from rapid assessment data. Auk, 119: 749-769. [ Links ]
Johnson, J. B. & Omland, K. S. (2004). Model selection in ecology and evolution. Trends in Ecology & Evolution, 19: 101-108. [ Links ]
Khuroo, A. A., Dar, G. H., Khan, Z. S. & Malik, A. H. (2007). Exploring an inherent interface between taxonomy and biodiversity: current problems and future challenges. Journal for Nature Conservation, 15: 256-261. [ Links ]
Kovacic, M., Patzner, R. A. & Schliewen, U. (2012). A first quantitative assessment of the ecology of cryptobenthic fishes in the Mediterranean Sea. Marine Biology, 159: 2731-2742. [ Links ]
Lemon, J. (2006). Plotrix: a package in the red light district of R. R-News, 6: 8-12. [ Links ]
Lozano, S. & Zapata, F. A. (2003). Short-term temporal patterns of settlement and settlement strategies of coral reef fishes in the tropical eastern Pacific. Marine Biology, 142: 399-409. [ Links ]
Mace, G. M. (2004). The role of taxonomy in species conservation. Philosophical Transactions of the Royal Society Biological Sciences, 359: 711-719. [ Links ]
Mora, C. & Zapata, F. A. (2000). Effects of predatory site-attached fish on abundance and body size of early post-settled reef fishes from Gorgona Island, Colombia. Proceedings of the 9th International Coral Reef Symposium, 1: 475-480. [ Links ]
Mora, C., Titterson, D. P. & Myers, R. A. (2008). The completeness of taxonomic inventories for describing the global diversity and distribution of marine fishes. Philosophical Transactions of the Royal Society Biological Sciences, 275: 149-155. [ Links ]
Morrison, M. L., Marcot, B. G. & Mannan, R. W. (2006). Wildlife-habitat relationships: concepts and application. Island Press, Washington, D.C. [ Links ]
Murdoch, D & Chow, E. D. (2007). Ellipse: functions for drawing ellipses and ellipse-like confidence regions. R package version 0.3-5. [ Links ]
Oksanen, J., Blanchet, F. G., Kind, R., Legendre, P., O’Hara, R. B., Simpson, G. L., Solymos, P., Henry, M., Stevens, H. & Wagner, H. (2010). Vegan: community ecology package. R package version 1.17-3. [ Links ]
Palacios, M. M. & Zapata, F. A. (2011). Sesgos asociados a censos visuales de peces arrecifales en el Pacífico Oriental Tropical: implicaciones del uso de transectos sencillos o de tamaño mixto, de la experiencia de los buzos y de la estimación de la talla. Boletín de Investigaciones Marinas y Costeras, 40: 117-138. [ Links ]
Pérez-España, H., Galván-Magaña, F. & Abitia-Cárdenas, L. A. (1996). Variaciones temporales y espaciales en la estructura de la comunidad de peces de arrecifes rocosos del suroeste del golfo de California, México. Ciencias Marinas, 22: 273-294. [ Links ]
Prahl, H. (1986). Notas sobre la geología, climatología, régimen de mareas y oceanografía. In H. Prahl and M. Alberico (Eds.), Isla Gorgona (pp. 19-27.). Biblioteca Banco Popular, Bogotá, Colobia. [ Links ]
Quinn, G. P. & Keough, M. J. (2002). Experimental design and data analysis for biologist. Cambridge University Press, England. [ Links ]
R Development Core Team. (2010). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Retrieved from //www.R-project.org [ Links ]
Robertson, D. R. & Allen, G. R. (2006). Shorefishes of the tropical eastern Pacific: an information system, ver. 2. DVD-ROM. Smithsonian Tropical Research Institute, Balboa, Panamá [ Links ].
Robertson, D. R. & Smith-Vaniz, W. F. (2008). Rotenone: An essential but demonized tool for assessing marine fish diversity. Bioscience, 58:165-170. [ Links ]
Robertson, D. R. & Smith-Vaniz, W. F. (2010). Use of clove oil in collecting coral reef fishes for research. Marine Ecology Progress Series, 401: 295-302. [ Links ]
Rojas, M. (1992). The species problem and conservation: what are we protecting? Conservation Biology, 6: 170-178. [ Links ]
Rubio, E. A. (1990). Ictiofauna Indopacífica asociada a los corales de la Isla de Gorgona. Revista de Ciencias, 2: 97-105. [ Links ]
Sale, P. F. & Dybdahl, R. (1975). Determinants of community structure for coral reef fishes in an experimental habitat. Ecology, 56: 1343-1355. [ Links ]
Sale, P. F. & Douglas, W. A. (1984). Temporal variability in the community structure of fish on coral patch reefs and the relation of community structure to reef structure. Ecology, 65:409-422. [ Links ]
Smith, C. L. & Tyler, J. C. (1972). Space resource sharing in a coral reef community. Bulletin of Natural History Museum Los Angeles City, 14: 125-170. [ Links ]
Smith, C. L. (1973). Small rotenone stations: a tool for studying coral reef fish communities. American Museum Novitates, 2512: 1-21. [ Links ]
Talbot, F. H., Russel, B. C. & Anderson, G. R. V. (1978). Coral reef fish communities: unstable, high diversity systems? Ecological Monographs, 48: 425-440. [ Links ]
Thompson, A. A. & Mapstone, B. D. (1997). Observer effects and training in underwater visual surveys of reef fishes. Marine Ecology Progress Series, 154: 56-63. [ Links ]
Willis, T. J. (2001). Visual census methods underestimate density and diversity of cryptic reef fishes. Journal of Fish Biology, 59: 1408-1411. [ Links ]
Zapata, F. A. & Morales, Y. A. (1997). Spatial and temporal patterns of fish diversity in a coral reef at Gorgona Island, Colombia. Proceedings of the 8th International Coral Reef Symposium, 1: 1029-1034. [ Links ]
Zapata, F. A. (2001). Formaciones coralinas de isla Gorgona. In L. M. Barrios & M. López-Victoria (Eds.), Gorgona Marina: Contribución al conocimiento de una isla única (pp. 27-40). INVEMAR, Serie de Publicaciones Especiales 7, Santa Marta, Colombia. [ Links ]
Zapata, F. A. & Robertson, D. R. (2007). How many species of shore fishes are there in the Tropical Eastern Pacific? Journal of Biogeography, 34: 38-51. [ Links ]
Ackerman, J. & Bellwood, D. (2000). Reef fish assemblages: a re-evaluation using enclosed rotenone stations. Marine Ecology Progress Series, 206: 227-237. [ Links ]
Alvarez-Filip, L., Reyes-Bonilla, H. & Calderon-Aguilera, L. E. (2006). Community structure of fishes in Cabo Pulmo Reef, Gulf of California. Marine Ecology Progress Series, 27: 253-262. [ Links ]
Alzate, A., Muñoz, C. G., Zapata, F. A. & Giraldo, A. (2012). New records of cryptobenthic fishes in coral reef habitats of Gorgona Islands, Colombia, Tropical Eastern Pacific. Boletín de Investigaciones Marinas y Costeras, 41: 229-235. [ Links ]
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Australian Journal of Ecology, 26: 32-46. [ Links ]
Bates, D., Maechler, M. & Bolker, B. (2013). Lme4: linear mixed-effects models using S4 classes. R. package version 0.999999-2. [ Links ]
Bellwood, D. R. & Alcala, A. C. (1988). The effects of a minimum length specification on visual estimates of density and biomass of coral reef fishes. Coral Reefs, 7: 23-27. [ Links ]
Benfield, S., Baxter, L., Guzman, H. M. & Mair, J. M. (2008). A comparison of coral reef and coral communities fish assemblages in Pacific Panama and environmental factors governing their structure. Journal of the Marine Biological Association of the United Kingdom, 88: 1331-1341. [ Links ]
Bohnsack, J. A. & Talbot, F. H. (1980). Species packing by reef fishes on Australian and Caribbean reefs: an experimental approach. Bulletin of Marine Science, 30: 710-723. [ Links ]
Brock, V. E. (1954). A preliminary report on a method of estimating reef fish populations. Journal of Wildlife Management, 18: 297-308. [ Links ]
Brock. R. E. (1982). A critique of the visual census method for assessing coral reef fish populations. Bulletin of Marine Science, 32: 269-276. [ Links ]
Burnham, K. P. & Anderson, D. R. (2002). Model selection and multimodel inference: a practical information-theoretical approach. Springer-Verlag, New York, USA. [ Links ]
Coles, S. L. & Tarr, A. B. (1990). Reef fish assemblages in the western Arabian Gulf: a geographically isolated population in an extreme environment. Bulletin of Marine Science, 47: 696-720. [ Links ]
Depczynski, M. & Bellwood, D. R. (2003). The role of cryptobenthic reef fishes in coral reef trophodynamics. Marine Ecology Progress Series, 256: 183-191. [ Links ]
Dominici-Arosemena, A. & Wolff, M. (2006). Reef fish community structure in the Tropical Eastern Pacific (Panamá): living on a relatively stable rocky reef environment. Helgoland Journal of Marine Research, 60: 287-305. [ Links ]
Dubois, A. (2003). The relationships between taxonomy and conservation biology in the century of extinctions. Comptes Rendus Biologies, 326: s9-s21. [ Links ]
Espinoza, M. & Salas, E. (2005). Estructura de las comunidades de peces de arrecife en las Islas Catalinas y Playa Ocotal, Pacífico norte de Costa Rica. Revista de Biología Tropical, 53: 523-536. [ Links ]
Galván-Villa, C. M., Arreola-Robles, J. L., Ríos-Jara, E. & Rodriguez-Zaragoza, F. A. (2010). Ensamblajes de peces arrecifales y su relación con el habitat bentónico de la isla Isabel, Nmayarit, México. Revista de Biología Marina y Oceanografía, 45: 311-324. [ Links ]
Giraldo, A., Rodríguez-Rubio, E. & Zapata, F. A. (2008). Condiciones oceanográficas en Isla Gorgona, Pacífico Oriental Tropical de Colombia. Latin American Journal of Aquatic Research, 36: 121-128. [ Links ]
Glynn, P. W., Prahl, H. v. & Guhl, F. (1982). Coral reef of Gorgona Island, Colombia, with special reference to corallivores and the influence on community structural reef development. Anales del Instituto de Investigaciones Marinas de Punta de Betín, 12: 185-214. [ Links ]
Goslee, S. C. & Urban, D. L. (2007). The ecodist package for dissimilarity-based analysis of ecological data. Journal of Statistical Software, 22:1-19. [ Links ]
Harborne, A. R., Jelks, H. L., Smith-Vaniz, W. F. & Rocha, L. A. (2012). Abiotic and biotic controls of cryptobenthic fish assemblages across a Caribbean seascape. Coral Reefs, 31: 977-990. [ Links ]
Herzog, S. K., Kessler, M. & Cahill, T. M. (2002). Estimating species richness of tropical bird communities from rapid assessment data. Auk, 119: 749-769. [ Links ]
Johnson, J. B. & Omland, K. S. (2004). Model selection in ecology and evolution. Trends in Ecology & Evolution, 19: 101-108. [ Links ]
Khuroo, A. A., Dar, G. H., Khan, Z. S. & Malik, A. H. (2007). Exploring an inherent interface between taxonomy and biodiversity: current problems and future challenges. Journal for Nature Conservation, 15: 256-261. [ Links ]
Kovacic, M., Patzner, R. A. & Schliewen, U. (2012). A first quantitative assessment of the ecology of cryptobenthic fishes in the Mediterranean Sea. Marine Biology, 159: 2731-2742. [ Links ]
Lemon, J. (2006). Plotrix: a package in the red light district of R. R-News, 6: 8-12. [ Links ]
Lozano, S. & Zapata, F. A. (2003). Short-term temporal patterns of settlement and settlement strategies of coral reef fishes in the tropical eastern Pacific. Marine Biology, 142: 399-409. [ Links ]
Mace, G. M. (2004). The role of taxonomy in species conservation. Philosophical Transactions of the Royal Society Biological Sciences, 359: 711-719. [ Links ]
Mora, C. & Zapata, F. A. (2000). Effects of predatory site-attached fish on abundance and body size of early post-settled reef fishes from Gorgona Island, Colombia. Proceedings of the 9th International Coral Reef Symposium, 1: 475-480. [ Links ]
Mora, C., Titterson, D. P. & Myers, R. A. (2008). The completeness of taxonomic inventories for describing the global diversity and distribution of marine fishes. Philosophical Transactions of the Royal Society Biological Sciences, 275: 149-155. [ Links ]
Morrison, M. L., Marcot, B. G. & Mannan, R. W. (2006). Wildlife-habitat relationships: concepts and application. Island Press, Washington, D.C. [ Links ]
Murdoch, D & Chow, E. D. (2007). Ellipse: functions for drawing ellipses and ellipse-like confidence regions. R package version 0.3-5. [ Links ]
Oksanen, J., Blanchet, F. G., Kind, R., Legendre, P., O’Hara, R. B., Simpson, G. L., Solymos, P., Henry, M., Stevens, H. & Wagner, H. (2010). Vegan: community ecology package. R package version 1.17-3. [ Links ]
Palacios, M. M. & Zapata, F. A. (2011). Sesgos asociados a censos visuales de peces arrecifales en el Pacífico Oriental Tropical: implicaciones del uso de transectos sencillos o de tamaño mixto, de la experiencia de los buzos y de la estimación de la talla. Boletín de Investigaciones Marinas y Costeras, 40: 117-138. [ Links ]
Pérez-España, H., Galván-Magaña, F. & Abitia-Cárdenas, L. A. (1996). Variaciones temporales y espaciales en la estructura de la comunidad de peces de arrecifes rocosos del suroeste del golfo de California, México. Ciencias Marinas, 22: 273-294. [ Links ]
Prahl, H. (1986). Notas sobre la geología, climatología, régimen de mareas y oceanografía. In H. Prahl and M. Alberico (Eds.), Isla Gorgona (pp. 19-27.). Biblioteca Banco Popular, Bogotá, Colobia. [ Links ]
Quinn, G. P. & Keough, M. J. (2002). Experimental design and data analysis for biologist. Cambridge University Press, England. [ Links ]
R Development Core Team. (2010). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Retrieved from //www.R-project.org [ Links ]
Robertson, D. R. & Allen, G. R. (2006). Shorefishes of the tropical eastern Pacific: an information system, ver. 2. DVD-ROM. Smithsonian Tropical Research Institute, Balboa, Panamá [ Links ].
Robertson, D. R. & Smith-Vaniz, W. F. (2008). Rotenone: An essential but demonized tool for assessing marine fish diversity. Bioscience, 58:165-170. [ Links ]
Robertson, D. R. & Smith-Vaniz, W. F. (2010). Use of clove oil in collecting coral reef fishes for research. Marine Ecology Progress Series, 401: 295-302. [ Links ]
Rojas, M. (1992). The species problem and conservation: what are we protecting? Conservation Biology, 6: 170-178. [ Links ]
Rubio, E. A. (1990). Ictiofauna Indopacífica asociada a los corales de la Isla de Gorgona. Revista de Ciencias, 2: 97-105. [ Links ]
Sale, P. F. & Dybdahl, R. (1975). Determinants of community structure for coral reef fishes in an experimental habitat. Ecology, 56: 1343-1355. [ Links ]
Sale, P. F. & Douglas, W. A. (1984). Temporal variability in the community structure of fish on coral patch reefs and the relation of community structure to reef structure. Ecology, 65:409-422. [ Links ]
Smith, C. L. & Tyler, J. C. (1972). Space resource sharing in a coral reef community. Bulletin of Natural History Museum Los Angeles City, 14: 125-170. [ Links ]
Smith, C. L. (1973). Small rotenone stations: a tool for studying coral reef fish communities. American Museum Novitates, 2512: 1-21. [ Links ]
Talbot, F. H., Russel, B. C. & Anderson, G. R. V. (1978). Coral reef fish communities: unstable, high diversity systems? Ecological Monographs, 48: 425-440. [ Links ]
Thompson, A. A. & Mapstone, B. D. (1997). Observer effects and training in underwater visual surveys of reef fishes. Marine Ecology Progress Series, 154: 56-63. [ Links ]
Willis, T. J. (2001). Visual census methods underestimate density and diversity of cryptic reef fishes. Journal of Fish Biology, 59: 1408-1411. [ Links ]
Zapata, F. A. & Morales, Y. A. (1997). Spatial and temporal patterns of fish diversity in a coral reef at Gorgona Island, Colombia. Proceedings of the 8th International Coral Reef Symposium, 1: 1029-1034. [ Links ]
Zapata, F. A. (2001). Formaciones coralinas de isla Gorgona. In L. M. Barrios & M. López-Victoria (Eds.), Gorgona Marina: Contribución al conocimiento de una isla única (pp. 27-40). INVEMAR, Serie de Publicaciones Especiales 7, Santa Marta, Colombia. [ Links ]
Zapata, F. A. & Robertson, D. R. (2007). How many species of shore fishes are there in the Tropical Eastern Pacific? Journal of Biogeography, 34: 38-51. [ Links ]
1. Coral Reef Ecology Research Group, Department of Biology, Universidad del Valle, A.A. 25360, Cali, Colombia; fernando.zapata@correounivalle.edu.co, ecologia@univalle.edu.co
2. Community and Conservation Ecology, Centre for Ecological and Evolutionary Studies, University of Groningen, Groningen, the Netherlands.
3. Terrestrial Ecology Unit, Ghent University, Ghent, Belgium; adria.alzate@gmail.com
Recibido 18-X-2013. Corregido 20-XI-2013. Aceptado 19-XII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}