










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.1 San José Feb. 2014
Octocoral densities and mortalities in Gorgona Island, Colombia, Tropical Eastern Pacific
Juan A. Sánchez1*, Néstor E. Ardila2*, Julio Andrade3*, Luisa F. Dueñas4*, Raúl Navas5* & Diana Ballesteros6*
Abstract
Owing to the decrease of reef organisms in general, it has become essential to study populations that are prone to marine diseases, with the purpose of developing accurate survivorship predictions and in turn alarm on triggers and drivers of disease outbreaks. In this study, we quantified the octocorals of Gorgona island, Tropical Eastern Pacific (Colombia), during 2007 and 2009 documenting a mass mortality occurred during 2008. We recorded 16 octocoral species with densities that ranged between 2 and 30 colonies m-2. Most abundant octocorals were Leptogorgia alba and Pacifigorgia spp. (Gorgoniidae: Octocorallia). During 2009 we noticed a mass mortality involving Pacifigorgia irene, P. adamsi, P. rubicunda and P. eximia, with a reduction of 70% of the colonies between 12 and 20 m in water depth. Around 5% of seafans during 2007 had an epizootic disease similar to aspergillosis, which seems the cause of the mass octocoral mortality. This disease outbreak observed in Gorgona island, and other nearby areas of the Colombian Pacific during 2007-2010, corresponded to extended periods of anomalous elevated seawater surface temperatures and thermal anomalies during the upwelling season of 2008. Constant monitoring of seawater temperatures and octocoral populations are urgently needed in this area to understand the nature of this new disease outbreak. Rev. Biol. Trop. 62 (Suppl. 1): 209-219. Epub 2014 February 01.
Key words: Octocorals, aspergillosis, Pacifigorgia, Tropical Eastern Pacific, rising seawater temperatures, marine diseases, mass mortality, Gorgona Island, Colombia.
Resumen
Debido a la disminución de los organismos de los arrecifes en general, es fundamental conocer las poblaciones de organismos propensos a enfermedades marinas, con el propósito de desarrollar predicciones precisas de supervivencia y a su vez dar la alarma sobre los motivos y las causas de los brotes de las enfermedades. En este estudio, hemos cuantificado los octocorales de la Isla Gorgona, Pacífico Oriental Tropical (Colombia), durante 2007 y 2009, y documentamos una mortalidad masiva que se produjo durante el año 2008. Se registraron 16 especies de octocorales con densidades que oscilan entre 2 y 30 colonias m-2. Los octocorales más abundantes fueron Leptogorgia alba y Pacifigorgia spp. (Gorgoniidae: Octocorallia). Durante el año 2009 nos dimos cuenta de una mortalidad masiva que afectaba a Pacifigorgia irene, P. adamsi, P. rubicunda y P. eximia, con una reducción del 70% de las colonias entre 12 y 20 m de profundidad. Alrededor del 5% de los abanicos de mar durante el año 2007 tenían una epizootia similar a la aspergilosis, que parece ser la causa de la mortalidad masiva de octocorales. Este brote de la enfermedad observada en la Isla Gorgona, y otras zonas cercanas del Pacífico colombiano durante el período 2007-2010, corresponden a períodos prolongados de altas temperaturas superficiales del mar y de anomalías térmicas durante la temporada de surgencia durante 2008. Se necesita con urgencia un monitoreo constante de las temperaturas marinas y de las poblaciones de octocorales en esta área para comprender la naturaleza de este nuevo brote de enfermedad marina.
Palabras clave: Octocorales, aspergilosis, Pacifigorgia, Pacífico Oriental Tropical, aumento de las temperaturas del agua de mar, enfermedades marinas, mortalidad masiva, Isla Gorgona, Colombia.
Emerging and re-emerging marine diseases have increased in the last four decades (Ward & Lafferty, 2004; Bruno et al., 2007). Infectious coral diseases are now recognized as important modifying factors in natural systems, being involved in changes of reproduction and growth rates, community structure, and host genetic diversity (Sutherland, Porter & Torres, 2004). Coral diseases could be related with current environmental conditions, which include increasing seawater temperatures, changes in nutrient availability, pollution, and invasive species (Harvell, Altizer, Cattadori, Harrington & Weil, 2009; Sokolow, 2009). To comprehend the emerging appearance of diseases worldwide, which seem related with anthropogenic stress on ecosystems, complete information on host range is urgently needed (Harvell et al., 2004). A clear understanding also needs host range information in conjunction with evidence on the environmental drivers leading to disease (Harvell et al., 2007). Unfortunately, many coastal regions of the world are poorly, or not at all, monitored (Spalding et al., 2007), which prevent us for having a complete and simultaneous picture on marine diseases emergence.
The major threat of emerging marine diseases is when they lead to mass mortalities. Disease outbreaks in coral reefs can encompass the demise of entire populations from local scales up to >90% of the species (Lessios, Robertson & Cubit, 1984; Bruno et al., 2007; Sánchez et al., 2010). The best documented case of a mass mortality in octocorals is the seafan Gorgonia ventalina in the Caribbean during the 1980s (Garzon-Ferreira & Zea, 1992; Smith, Ives, Nagelkerken & Ritchie, 1996). Octocoral mass mortalities have been documented worldwide and are related to predator outbreaks (Schärer & Nemeth, 2010), catastrophic high seawater temperatures (Cerrano et al., 2000; Lasker, 2005) and fungal diseases (Geiser, Taylor, Ritchie & Smith, 1998b), among others. The aftermath of a mass mortality usually involves detrimental effects on octocoral population demography (Linares, Coma, & Zabala, 2008; Bruno, Ellner, Vu, & Harvell, 2011). This study documents the first octocoral mass mortality in Gorgona Island, Tropical Eastern Pacific (TEP).
The TEP includes a number of endemisms in oceanic islands (e.g., Cocos, Malpelo, and Galapagos) and remains greatly unexplored and under monitored. Unfortunately, it is known that over 10% of its fauna has elevated extinction risk (Polidoro et al., 2012). Despite the highly diverse and endemic octocoral TEP fauna, with genera such as Pacifigorgia that includes 35 species (Guzman & Breedy, 2012), no information on octocoral conservation status is hitherto available. Detailed octocoral abundance and density data is available for Malpelo Island, where Pacifigorgia seafans can reach densities up to 20col m-2 between 5 and 20m deep (Sánchez, Gómez, Escobar & Dueñas, 2011). Despite its great isolation and overall conservation state, between 2009 and 2010, up to 66% of colonies were found dead in some rocky reefs around Malpelo. In addition, a fungal disease similar to aspergillosis, commonly affecting Gorgonia in the Caribbean, was recently identified infecting several octocorals specially the seafans Pacifigorgia irene and P. eximia [Barrero-Canosa et al. (2013)]. That finding demonstrates the presence of Aspergillus sydowii in the tissue of diseased Pacifigorgia seafans, which comprise the signature of the aspergillosis epizootic (Toledo-Hernández, Zuluaga-Montero, Bones-González, Sabat & Bayman, 2008), nearly 30 years after the appearance of the outbreak in the Caribbean. To better understand the extend of the emergence of this new disease outbreak, this study provides (1) updated information on octocoral densities and species abundance in 2007 for Gorgona island, Colombia, and (2) the report of a mass octocoral mortality that occurred between 2007-2009.
Materials and methods
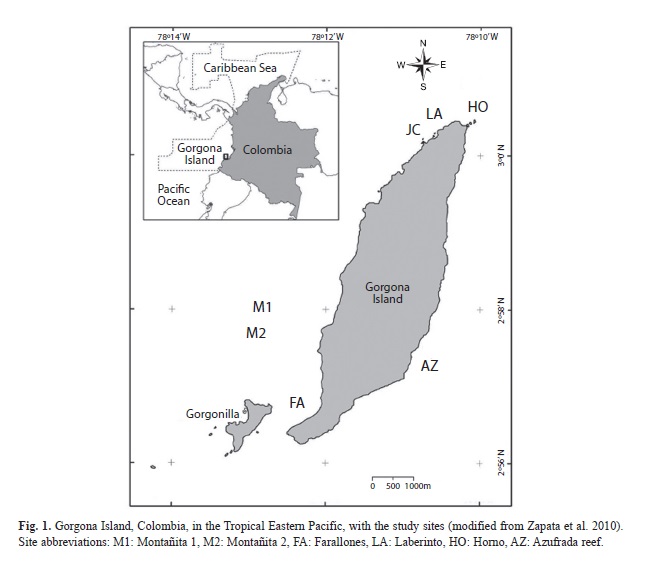
The study took place on the windward infralittorals of Gorgona islands, TEP, Colombia (Fig. 1). The scleractinian corals of Gorgona island have been studied in detail, given that the leeward side of the island includes one of the few well-developed coral reefs of the TEP (Zapata, Rodríguez-Ramírez, Caro-Zambrano & Garzón-Ferreira, 2010). In contrast, octocorals in Gorgona island have not been studied except for a few records of species in a taxonomic account of the Colombian Pacific (Prahl, Escobar & Molina, 1986). Octocorals occur in high densities from the intertidal zone down to approximately 30m at the windward side of Gorgona island. Hard substrate correspond to rocky cliffs and outcrops of igneous origin (Kerr, 2005). Octocorals (Octocorallia: Cnidaria) from the genera Pacifigorgia, Leptogorgia (Gorgoniidae) and Muricea (Plexauridae) are the most conspicuous invertebrates in this environment. In addition, the rocky substrate presents high abundances of sea urchins and bivalves, and high cover of coralline algae, filamentous algae, the azooxanthellate coral Tubastrea coccinea, a great variety of small invertebrates (anemones, hydroids, tunicates and zoanthids), and the occasional presence of sponges and hermatypic corals (Pocillopora spp., Porites spp., Psammocora stellata and Pavona spp.).
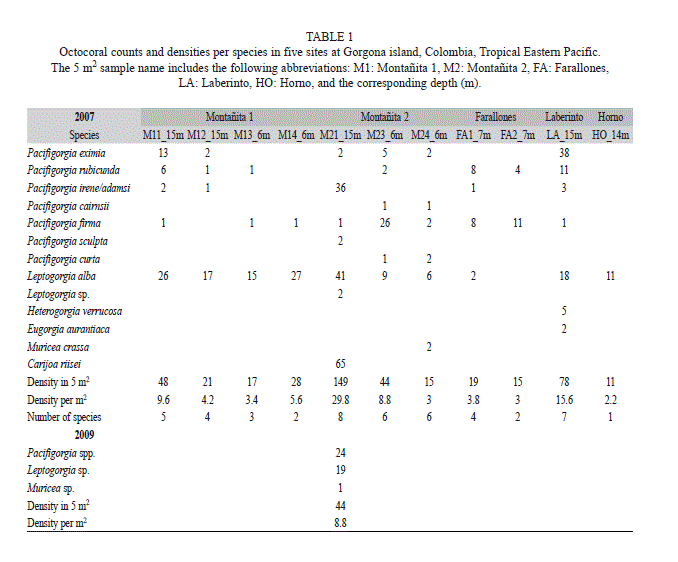
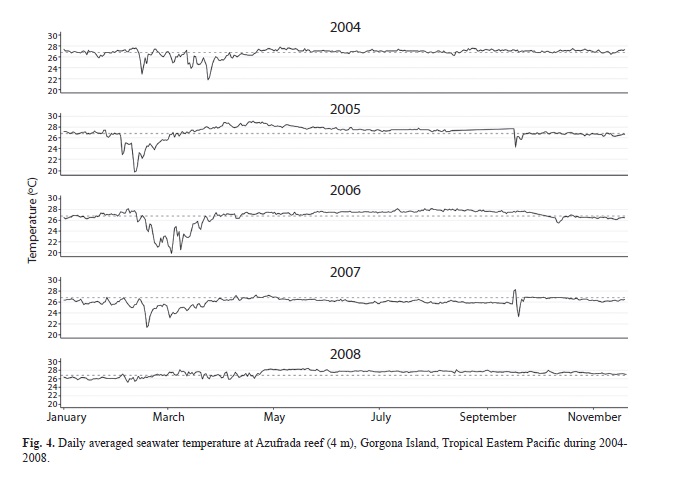
The first octocoral survey at Gorgona Island was conducted during September 2007 on most shallow hard-bottom habitats (6 and 20m) around the island (Fig. 1, Table 1). We completed a sampling effort of 11 dives with more than 9 hours of SCUBA. Octocoral densities were recorded on 50m transects, in which every 5m colony density was counted on two quadrants of 0.25m2 on each side of transect (Etnoyer, Wirshing, & Sánchez, 2010). For each morphospecies observed (e.g., including differences in colonial form and color) detailed digital photographs were taken, and small tissue samples (<3cm2) were maintained in seawater in sterile bags and then were preserved in absolute ethanol and dry for later sclerites analysis. We surveyed again some of the sampled sites (M2, FA and HO) during November 2009. At Montañita 2 (M2) we clearly noticed a mass octocoral mortality and conducted a 50m-transect in the same fashion and locality corresponding to the 15m site (Table 1). In addition, we cemented steel rods at the ends of two parallel 50 m transects (15m and 12m) and tagged a number of octocoral colonies for further monitoring. Digital photographs were taken underwater for each octocoral sample, most quadrants, and the general condition of the octocoral population during both 2007 and 2009. During 2011 we had a short visit to Montañita 2 to check the transect area (Fig. 3E-F). In addition, we included daily underwater temperatures measured between 2004-2008, which could determine the environmental cue triggering the disease. We placed an underwater temperature logger (Hobo, onset) at the Pocillopora spp. reef (Azufrada, AZ: Fig. 1) at 4 m of water depth.
Results
The octocoral survey included 16 species recorded in the transects with a greater dominance of Pacifigorgia spp. (9 species) and Leptogorgia spp. (two species, mostly Leptogorgia alba), and other genera represented by a single species Eugorgia auriantiaca, Muricea crassa, Heterogorgia verrucosa and Carijoa riisei. The latter is an invasive species from the Caribbean, which presented a high-density mat (~13 colonies m-2) in Montañita 2 (Table 1). Although, C. riisei was also found in other locations (HO and LA) but with lower abundances, it was not recorded in transects (see details in Sánchez and Ballesteros, 2014 this volume). Octocorals densities ranged between 2 and 30col m-2. Laberinto and Montañita (M1 and M2) sites had the higher species richness and density (Table 1). Most abundant octocorals were Leptogorgia alba, Pacifigorgia eximia, P. rubicunda, P. firma and P. irene/adamsi, the latter two species were nearly impossible to tell apart underwater consequently we pooled the counts from both species.
In general, octocorals preferred the windward rocky cliffs and outcrops at the Western side of Gorgona Island, where transects were carried out (Fig. 1). Only Leptogorgia alba was occasionally observed near the coral reefs and rubble/sand areas of the Eastern side of the island. During 2007, a noteworthy result was that about 5% of Pacifigorgia spp. had noticeable halos of an epizootic disease and/or dead tissue (Fig. 2 and 3 A-D). During 2009, in Montañita 2 we witnessed the aftermath of a seafan mass mortality, most likely Pacifigorgia irene/adamsi, P. rubicunda and P. eximia, with a reduction in octocoral density from 30 col m-2 in 2007 down to less than 9 col m-2 in 2009, a reduction of 70% of the colonies (Table 1). The mortality was particularly obvious below 12m and the numerous remains of seafan axial skeletons, not observed during 2007, were a proof of the mass mortality (Fig. 2). Local divers noticed the diseased corals and recalled the mass mortality probably during August-September 2008 (J. Orlando López pers. comm.). Other sites of the island (M1 and HO) had also dead octocoral skeletons suggesting that the mass mortality included most of the octocorals communities in Gorgona Island.
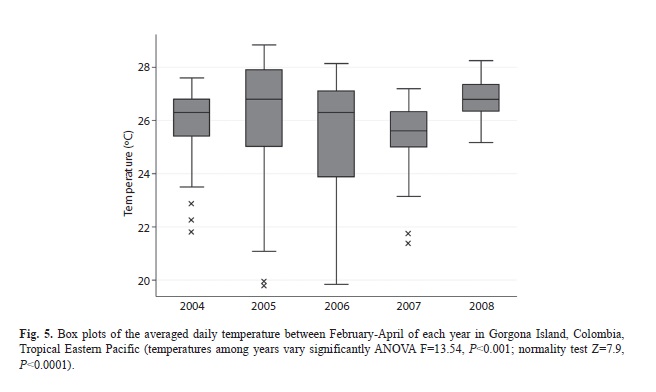
The general behavior of seawater temperatures, between October 2003 and November 2008 averaged 26.78ºC (19-20, min-max), and showed a seasonal cold period with high temperature amplitude during January and May (Fig. 4). Although no significant differences were found among years, 2008 was clearly the year with narrower seawater temperature amplitude and the higher minimum values during the cold period (Fig. 5).
Discussion
Gorgona Island, during 2007, sustained dense and diverse octocoral communities totaling 16 species. Rocky habitats between 6-20m included up to seven coexisting species at high densities (2-30col m-2). The maximum densities were higher than most Caribbean octocoral communities (Sánchez et al., 2005; Etnoyer et al., 2010), whereas the number of species per site is slightly lower in Gorgona Island. A notable difference with Caribbean octocoral communities is that TEP assemblages do not coexist with branching reef-building corals, e.g., Pocillopora spp., which form dense coral-dominated areas in Gorgona Island (Zapata et al., 2010). Similar octocoral densities have been observed in the oceanic island of Malpelo in a community dominated by only two seafan species together with L. alba (Sánchez et al., 2011). Likewise, this same community structure is also observed in the Pacific coast of Colombia (Cabo Corrientes, Choco) but with higher species diversity than the two islands (unpublished data). This suggests that octocoral diversity in this region follows an “island biogeography” type of pattern.
Our 2007 survey showed, for the first time, alarming signs of an epizootic disease in colonies from at least four species (Fig. 3). Seafans exhibited loss of tissue in a fashion resembling the Caribbean seafan fungal disease aspergillosis (Nagelkerken et al., 1997; Geiser et al., 1998a; Smith, Harvell & Kim, 1998; Alker, Smith & Kim, 2001; Smith & Weil, 2004). Similarly, P. eximia and P. irene were surveyed during 2009 for fungal pathogens in the Pacific coast of Colombia (Choco area) and the pathogenic agent of aspergillosis, Aspergillus sydowi, was isolated in diseased colonies (Barrero-Canosa et al. 2013). A similar epizootic disease was observed in Malpelo Island, a pristine oceanic rock escarpment, that affected up to 66% of colonies. During November 2009 and February 2010, this disease lead to local mass mortalities in some areas with a population reduction between 1 and 21% in most sites (Sánchez et al. 2011). Most sites in the Colombian TEP showed an overall recovery of the diseased during 2011 and one of the affected seafans, P. irene, shows remarkable regeneration of surviving colonies (Fig. 3E-F).
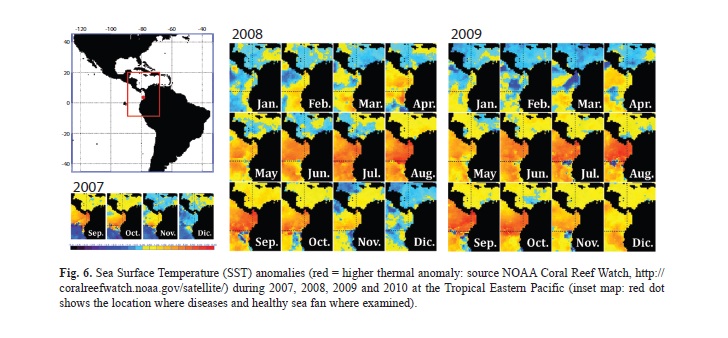
During 2008, the year when the mass mortality most likely occurred, seawater temperatures in Gorgona Island were warmer than the preceding years. In addition, from 2007 to 2010, Sea Surface Temperatures (SSTs) have been considered anomalous (Fig. 6). The Colombian Pacific basin involves a complex interaction of six wind masses with the Intertropical Convergence zone, which produces a constant upwelling area with strong winds and vertical water flows from January to April (Díaz, Málikov & Villegas, 2008). Our temperature logger retrieved data clearly showing the upwelling period (Figs. 4-5). Interestingly, it has been recently suggested that climate change can particularly affect upwelling zones at the tropics. Increased water vapor, the main greenhouse gas, intensifies thermal low-pressure cells over coastal landmasses leading to abnormal wind stress in upwelling regions (Bakun, Field, Redondo-Rodriguez & Weeks, 2010). It is possible that this upwelling region is also subjected to changes in wind stress and SSTs during the cold season, which could explain the observed pattern during 2008. The octocoral mass mortality at Gorgona Island is another case in a long list where thermal stress is related to coral disease outbreaks and mortality (Bruno et al., 2007).
During 2007-2010, mass and partial mortalities occurred at Gorgona and Malpelo Islands, as well as the Pacific coast of Colombia. These years correspond to an extended period of anomalous elevated SSTs (Fig. 6). According to Alker et al. (2001), aspergillosis appearance could be related to rising seawater temperatures. In the Caribbean, anomalous high temperatures reduce the anti-fungal activity of Gorgonia ventalina (Gorgoniidae), which interferes with the coral’s defense mechanisms leading to an easy disease spread (Ward, Kim & Harvell, 2006; Mydlarz, Holthouse, Peters & Harvell, 2008). The disease susceptibility is clearly restricted to a reduced number of species and until now no report on aspergillosis-like mass mortalities for the TEP area has been published. The Pacifigorgia seafan species affected in Gorgona Island and other rocky reefs in the Colombian TEP, could be an earlier warning on the devastating effects of emerging diseases exacerbated by global warming conditions. The Colombian TEP seems to be a new marine diseases hotspot. Constant monitoring of seawater temperatures and octocoral demography and health is urgently recommended in order to clarify the triggers and drivers of disease outbreaks as well as to understand the resistance and resilience of these new aspergillosis hosts.
Acknowledgments
This study was funded by the Universidad de los Andes, Colombia (Interfacultades, Vicerrectoria de Investigaciones, and Facultad de Ciencias), National Geographic Society-Waitt grants, COLCIENCIAS (grant No. 1204-521-29002) and two expeditions (2007 and 2009) organized by the PNN-Gorgona (Nancy Murillo, Ximena Zorrilla, Eduardo Sandoval and all the park staff) and Fundación Yubarta (Lilian Florez) as well as a short visit to Gorgona Island during 2011 supported by Fundación Malpelo (Sandra Bessudo, Germán Soler and Stan Teillaud). We appreciate the help from colleagues and students from BIOMMAR for assistant in the field and laboratory specially Nelson Manrique, Johanna Parra, Edwin Gomez, Martha Cárdenas, Laura Romero, Dairo Escobar and Juliana Sintura. Johanna Vega (SIMAC-INVEMAR) helped us tremendously during the field sampling in 2009. Odalisca Breedy and Hector Guzman kindly revised and corrected the identifications of the octocorals collected in this study. We recognize the participation and support from local communities and to the crew of vessel Maria Patricia.
References
Alker, A. P., Smith, G. W. & Kim, K. (2001). Characterization of Aspergillus sydowii (Thom et Church), a fungal pathogen of Caribbean sea fan corals. Hydrobiología, 460: 105-111. [ Links ]
Bakun, A., Field, D. B., Redondo-Rodriguez, A. N. A. & Weeks, S. J. (2010). Greenhouse gas, upwelling-favorable winds, and the future of coastal ocean upwelling ecosystems. Global Change Biology, 16: 1213-1228. [ Links ]
Barrero-Canosa J., L. Dueñas & J.A. Sánchez (2013) Isolation of Potential Fungal Pathogens in gorgonian corals at the Tropical Eastern Pacific. Coral Reefs 32: 35-41. [ Links ]
Bruno, J. F., Selig, E. R., Casey, K. S., Page, C. A., Willis, B. L., Harvell, C. D., Sweatman, H. & Melendy, A. M.. (2007). Thermal Stress and Coral Cover as Drivers of Coral Disease Outbreaks. PLoS Biol, 5:e124. [ Links ]
Bruno, J. F., Ellner, S. P., Vu, I. & Harvell, C. D. (2011). Impacts of Aspergillosis on sea fan coral demography: modeling a moving target. Ecological Monographs, 81: 123-139. [ Links ]
Cerrano, C., Bavestrello, G., Bianchi, C. N., Cattaneo-Vietti, R., Bava, S., Morganti, C., Morri, C., Picco, P., Sara, G., Schiaparelli, S., Siccardi, A., & Sponga, F. (2000). A catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (North-western Mediterranean), summer 1999. Ecology Letters, 3: 284-293. [ Links ]
Díaz, D., Málikov, I., & Villegas, N. (2008). Características de las zonas de surgencia de la cuenca del Pacífico colombiano y su relación con la zona de convergencia intertropical. Boletín Científico. Centro de Investigaciones Oceanográficas e Hidrográficas del Caribe, 26: 59-71. [ Links ]
Etnoyer, P. J., Wirshing, H. H. & Sánchez, J. A. (2010). Rapid Assessment of Octocoral Diversity and Habitat on Saba Bank, Netherlands Antilles. PLoS One, 5:e10668. [ Links ]
Garzon-Ferreira, J. & Zea, S. (1992). A Mass Mortality of Gorgonia ventalina (Cnidaria, Gorgoniidae) in the Santa Marta Area, Caribbean Coast of Colombia. Bulletin of Marine Science, 50: 522-526. [ Links ]
Geiser, D. M., Taylor, J. W., Ritchie, K. B. & Smith, G. W. (1998a). Cause of sea fan death in the West Indies. Nature, 394: 137-138. [ Links ]
Geiser, D. M., Taylor, J. W., Ritchie, K. B., & Smith, G. W. (1998b). Cause of sea fan death in the West Indies. Nature, 394: 137-138. [ Links ]
Guzman, H. M. & Breedy, O. (2012). Pacifigorgia marviva (Anthozoa: Octocorallia) a new species from Coiba National Park, Pacific Panama. Journal of the Marine Biological Association of the United Kingdom, 92: 693-698. [ Links ]
Harvell, D., Aronson, R., Baron, N., Connell, J., Dobson, A., Ellner, S., Gerber, L., Kim, K., Kuris, A., McCallum, H., Lafferty, K., McKay, B., Porter, J., Pascual, M., Smith, G., Sutherland, K. & Ward, J. (2004). The rising tide of ocean diseases: unsolved problems and research priorities. Frontiers in Ecology and the Environment, 2: 375-382. [ Links ]
Harvell, D., Jordan-Dahlgren, E., Merkel, S., Rosenberg, E., Raymundo, L., Smith, G., Weil, E., Willis, B. & Coral, G. E. F. (2007). Coral Disease, Environmental Drivers, and the Balance between Coral and Microbial Associates. Oceanography, 20: 172-195. [ Links ]
Harvell, D., Altizer, S., Cattadori, I. M., Harrington, L. & Weil, E. (2009). Climate change and wildlife diseases: When does the host matter the most? Ecology, 90: 912-920. [ Links ]
Kerr, A. C. (2005). La Isla de Gorgona, Colombia: A petrological enigma? Lithos, 84: 77-101. [ Links ]
Lasker, H. R. (2005). Gorgonian mortality during a thermal event in the Bahamas. Bulletin Marine Science, 76: 155-162. [ Links ]
Lessios, H. A., Robertson, D. R. & Cubit, J. D. (1984). Spread of Diadema mass mortality through the Caribbean. Science, 226: 335-337. [ Links ]
Linares, C., Coma, R., & Zabala, M. (2008). Effects of a mass mortality event on gorgonian reproduction. Coral Reefs, 27: 27-34. [ Links ]
Mydlarz, L. D., Holthouse, S. F., Peters, E. C. & Harvell, C. D. (2008). Cellular responses in sea fan corals: granular amoebocytes react to pathogen and climate stressors. PLoS One, 3:e1811. [ Links ]
Nagelkerken, I., Buchan, K., Smith, G. W., Bonair, K., Bush P., Garzon-Ferreira, J., Botero, L., Gayle, P., Harvell, C. D., Heberer, C., Kim, K., Petrovic, C., Pors, L., & Yoshioka, P. (1997). Widespread disease in Caribbean sea fans. II: Patterns of infection and tissue loss. Marine Ecology Progress Series, 160: 255-263. [ Links ]
Prahl, H. V., Escobar, D. & Molina, G. (1986). Octocorales (Octocorallia: Gorgonüdae y Plexauridae) de aguas someras del Pacífico Colombiano. Revista de Biología Tropical, 34(1): 13-33. [ Links ]
Polidoro, B. A., Brooks, T., Carpenter, K. E., Edgar, G. J., Henderson, S., Sanciangco, J. & Robertson, D. R. (2012). Patterns of extinction risk and threat for marine vertebrates and habitat-forming species in the Tropical Eastern Pacific. Marine Ecology Progress Series, 448: 93-104. [ Links ]
Sánchez, J. A., Pizarro, V., Acosta, A. R. S., Catillo, P. A., Herron, P., Martínez, J. C., Montoya, P. & Orozco, C. (2005). Evaluating coral reef benthic communities in remote Caribbean atolls (Quitasueño, Serrana, and Roncador banks) to recommend marine-protected areas for the seaflower biosphere reserve. Atoll Research Bulletin. National museum of natural history, Washington, D.C. [ Links ]
Sánchez, J. A., Herrera, S., Navas-Camacho, R., Rodríguez-Ramírez, A., Herron, P., Pizarro, V., Acosta, A. R., Castillo, P. A., Montoya, P. & Orozco, C. (2010). White plague-like coral disease in remote reefs of the Western Caribbean. Revista de Biología Tropical, 58: 145-154. [ Links ]
Sánchez, J. A., Gómez, C. E., Escobar, D., & Dueñas, L. F. (2011). Diversidad, abundancia y amenazas de los octocorales de la isla Malpelo, Pacífico Oriental Tropical, Colombia. Boletín de Investigaciones Marinas y Costeras, 40: 139-154. [ Links ]
Sánchez J.A. & D. Ballesteros (2014) The invasive snowflake coral (Carijoa riisei) in the Tropical Eastern Pacific, Colombia. Revista de Biología Tropical 62 (Suppl. 1): 197-207. [ Links ]
Schärer, M. & Nemeth, M. (2010). Mass mortality of gorgonians due to a Cyphoma gibbosum (Linnaeus) population outbreak at Mona Island, Puerto Rico. Coral Reefs, 29: 533-533. [ Links ]
Smith, G. W., Ives, L. D., Nagelkerken, I. A. & Ritchie, K. B. (1996). Caribbean sea-fan mortalities. Nature, 383: 487-487. [ Links ]
Smith, G. W., Harvell, C. D. & Kim, K. (1998). Response of sea fans to infection with Aspergillus sp. (Fungi). Revista de Biología Tropical, 46: 205-208. [ Links ]
Smith, G. W. & Weil, E. (2004). Aspergillosis of Gorgonians. In E. Rosenberg & Y. Loya (Eds.), Coral Health and Disease (pp. 279-287). New York, USA. [ Links ]
Sokolow, S. H. (2009). Effects of a changing climate on the dynamics of coral infectious disease: a review of the evidence. Diseases of Aquatic Organisms, 87:5-18. [ Links ]
Spalding, M. D., Fox, H. E., Allen, G. R., Davidson, N., Ferdaña, Z. A., Finlayson, A. X., Halpern, B. S., Jorge, M. A., Lombana, A. L., Lourie, S. A., Martin, K. D., McManus, E., Molnar, J., Recchia, C. A., & Robertson, J. (2007). Marine Ecoregions of the World: A Bioregionalization of Coastal and Shelf Areas. BioScience, 57: 573-583. [ Links ]
Sutherland, K. P., Porter, J. W. & Torres, C. (2004). Disease and immunity in Caribbean and Indo-Pacific zooxanthellate corals. Marine Ecology-Progress Series, 266:273-302. [ Links ]
Toledo-Hernández, C., Zuluaga-Montero, A., Bones-González, A., Sabat, A. M. & Bayman, P. (2008). Fungi in healthy and diseased sea fans (Gorgonia ventalina): Is Aspergillus sydowii always the pathogen? Coral Reefs, 27: 707-714. [ Links ]
Ward, J. R. & Lafferty, K. D. (2004). The Elusive Baseline of Marine Disease: Are Diseases in Ocean Ecosystems Increasing? PLoS Biol, 2:e120. [ Links ]
Ward, J. R., Kim, K. & Harvell, C. D. (2006). Temperature affects coral disease resistance and pathogen growth. Marine Ecology-Progress Series, 329: 115-121. [ Links ]
Zapata, F. A., Rodríguez-Ramírez, A., Caro-Zambrano, C. & Garzón-Ferreira, J. (2010). Mid-term coral-algal dynamics and conservation status of a Gorgona Island (Tropical Eastern Pacific) coral reef. Revista de Biología Tropical, 58: 81-94. [ Links ]
1. Departamento de Ciencias Biológicas-Facultad de Ciencias, Laboratorio de Biología Molecular Marina-BIOMMAR, Universidad de los Andes, Bogotá, Colombia; juansanc@uniandes.edu.co
Bakun, A., Field, D. B., Redondo-Rodriguez, A. N. A. & Weeks, S. J. (2010). Greenhouse gas, upwelling-favorable winds, and the future of coastal ocean upwelling ecosystems. Global Change Biology, 16: 1213-1228. [ Links ]
Barrero-Canosa J., L. Dueñas & J.A. Sánchez (2013) Isolation of Potential Fungal Pathogens in gorgonian corals at the Tropical Eastern Pacific. Coral Reefs 32: 35-41. [ Links ]
Bruno, J. F., Selig, E. R., Casey, K. S., Page, C. A., Willis, B. L., Harvell, C. D., Sweatman, H. & Melendy, A. M.. (2007). Thermal Stress and Coral Cover as Drivers of Coral Disease Outbreaks. PLoS Biol, 5:e124. [ Links ]
Bruno, J. F., Ellner, S. P., Vu, I. & Harvell, C. D. (2011). Impacts of Aspergillosis on sea fan coral demography: modeling a moving target. Ecological Monographs, 81: 123-139. [ Links ]
Cerrano, C., Bavestrello, G., Bianchi, C. N., Cattaneo-Vietti, R., Bava, S., Morganti, C., Morri, C., Picco, P., Sara, G., Schiaparelli, S., Siccardi, A., & Sponga, F. (2000). A catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (North-western Mediterranean), summer 1999. Ecology Letters, 3: 284-293. [ Links ]
Díaz, D., Málikov, I., & Villegas, N. (2008). Características de las zonas de surgencia de la cuenca del Pacífico colombiano y su relación con la zona de convergencia intertropical. Boletín Científico. Centro de Investigaciones Oceanográficas e Hidrográficas del Caribe, 26: 59-71. [ Links ]
Etnoyer, P. J., Wirshing, H. H. & Sánchez, J. A. (2010). Rapid Assessment of Octocoral Diversity and Habitat on Saba Bank, Netherlands Antilles. PLoS One, 5:e10668. [ Links ]
Garzon-Ferreira, J. & Zea, S. (1992). A Mass Mortality of Gorgonia ventalina (Cnidaria, Gorgoniidae) in the Santa Marta Area, Caribbean Coast of Colombia. Bulletin of Marine Science, 50: 522-526. [ Links ]
Geiser, D. M., Taylor, J. W., Ritchie, K. B. & Smith, G. W. (1998a). Cause of sea fan death in the West Indies. Nature, 394: 137-138. [ Links ]
Geiser, D. M., Taylor, J. W., Ritchie, K. B., & Smith, G. W. (1998b). Cause of sea fan death in the West Indies. Nature, 394: 137-138. [ Links ]
Guzman, H. M. & Breedy, O. (2012). Pacifigorgia marviva (Anthozoa: Octocorallia) a new species from Coiba National Park, Pacific Panama. Journal of the Marine Biological Association of the United Kingdom, 92: 693-698. [ Links ]
Harvell, D., Aronson, R., Baron, N., Connell, J., Dobson, A., Ellner, S., Gerber, L., Kim, K., Kuris, A., McCallum, H., Lafferty, K., McKay, B., Porter, J., Pascual, M., Smith, G., Sutherland, K. & Ward, J. (2004). The rising tide of ocean diseases: unsolved problems and research priorities. Frontiers in Ecology and the Environment, 2: 375-382. [ Links ]
Harvell, D., Jordan-Dahlgren, E., Merkel, S., Rosenberg, E., Raymundo, L., Smith, G., Weil, E., Willis, B. & Coral, G. E. F. (2007). Coral Disease, Environmental Drivers, and the Balance between Coral and Microbial Associates. Oceanography, 20: 172-195. [ Links ]
Harvell, D., Altizer, S., Cattadori, I. M., Harrington, L. & Weil, E. (2009). Climate change and wildlife diseases: When does the host matter the most? Ecology, 90: 912-920. [ Links ]
Kerr, A. C. (2005). La Isla de Gorgona, Colombia: A petrological enigma? Lithos, 84: 77-101. [ Links ]
Lasker, H. R. (2005). Gorgonian mortality during a thermal event in the Bahamas. Bulletin Marine Science, 76: 155-162. [ Links ]
Lessios, H. A., Robertson, D. R. & Cubit, J. D. (1984). Spread of Diadema mass mortality through the Caribbean. Science, 226: 335-337. [ Links ]
Linares, C., Coma, R., & Zabala, M. (2008). Effects of a mass mortality event on gorgonian reproduction. Coral Reefs, 27: 27-34. [ Links ]
Mydlarz, L. D., Holthouse, S. F., Peters, E. C. & Harvell, C. D. (2008). Cellular responses in sea fan corals: granular amoebocytes react to pathogen and climate stressors. PLoS One, 3:e1811. [ Links ]
Nagelkerken, I., Buchan, K., Smith, G. W., Bonair, K., Bush P., Garzon-Ferreira, J., Botero, L., Gayle, P., Harvell, C. D., Heberer, C., Kim, K., Petrovic, C., Pors, L., & Yoshioka, P. (1997). Widespread disease in Caribbean sea fans. II: Patterns of infection and tissue loss. Marine Ecology Progress Series, 160: 255-263. [ Links ]
Prahl, H. V., Escobar, D. & Molina, G. (1986). Octocorales (Octocorallia: Gorgonüdae y Plexauridae) de aguas someras del Pacífico Colombiano. Revista de Biología Tropical, 34(1): 13-33. [ Links ]
Polidoro, B. A., Brooks, T., Carpenter, K. E., Edgar, G. J., Henderson, S., Sanciangco, J. & Robertson, D. R. (2012). Patterns of extinction risk and threat for marine vertebrates and habitat-forming species in the Tropical Eastern Pacific. Marine Ecology Progress Series, 448: 93-104. [ Links ]
Sánchez, J. A., Pizarro, V., Acosta, A. R. S., Catillo, P. A., Herron, P., Martínez, J. C., Montoya, P. & Orozco, C. (2005). Evaluating coral reef benthic communities in remote Caribbean atolls (Quitasueño, Serrana, and Roncador banks) to recommend marine-protected areas for the seaflower biosphere reserve. Atoll Research Bulletin. National museum of natural history, Washington, D.C. [ Links ]
Sánchez, J. A., Herrera, S., Navas-Camacho, R., Rodríguez-Ramírez, A., Herron, P., Pizarro, V., Acosta, A. R., Castillo, P. A., Montoya, P. & Orozco, C. (2010). White plague-like coral disease in remote reefs of the Western Caribbean. Revista de Biología Tropical, 58: 145-154. [ Links ]
Sánchez, J. A., Gómez, C. E., Escobar, D., & Dueñas, L. F. (2011). Diversidad, abundancia y amenazas de los octocorales de la isla Malpelo, Pacífico Oriental Tropical, Colombia. Boletín de Investigaciones Marinas y Costeras, 40: 139-154. [ Links ]
Sánchez J.A. & D. Ballesteros (2014) The invasive snowflake coral (Carijoa riisei) in the Tropical Eastern Pacific, Colombia. Revista de Biología Tropical 62 (Suppl. 1): 197-207. [ Links ]
Schärer, M. & Nemeth, M. (2010). Mass mortality of gorgonians due to a Cyphoma gibbosum (Linnaeus) population outbreak at Mona Island, Puerto Rico. Coral Reefs, 29: 533-533. [ Links ]
Smith, G. W., Ives, L. D., Nagelkerken, I. A. & Ritchie, K. B. (1996). Caribbean sea-fan mortalities. Nature, 383: 487-487. [ Links ]
Smith, G. W., Harvell, C. D. & Kim, K. (1998). Response of sea fans to infection with Aspergillus sp. (Fungi). Revista de Biología Tropical, 46: 205-208. [ Links ]
Smith, G. W. & Weil, E. (2004). Aspergillosis of Gorgonians. In E. Rosenberg & Y. Loya (Eds.), Coral Health and Disease (pp. 279-287). New York, USA. [ Links ]
Sokolow, S. H. (2009). Effects of a changing climate on the dynamics of coral infectious disease: a review of the evidence. Diseases of Aquatic Organisms, 87:5-18. [ Links ]
Spalding, M. D., Fox, H. E., Allen, G. R., Davidson, N., Ferdaña, Z. A., Finlayson, A. X., Halpern, B. S., Jorge, M. A., Lombana, A. L., Lourie, S. A., Martin, K. D., McManus, E., Molnar, J., Recchia, C. A., & Robertson, J. (2007). Marine Ecoregions of the World: A Bioregionalization of Coastal and Shelf Areas. BioScience, 57: 573-583. [ Links ]
Sutherland, K. P., Porter, J. W. & Torres, C. (2004). Disease and immunity in Caribbean and Indo-Pacific zooxanthellate corals. Marine Ecology-Progress Series, 266:273-302. [ Links ]
Toledo-Hernández, C., Zuluaga-Montero, A., Bones-González, A., Sabat, A. M. & Bayman, P. (2008). Fungi in healthy and diseased sea fans (Gorgonia ventalina): Is Aspergillus sydowii always the pathogen? Coral Reefs, 27: 707-714. [ Links ]
Ward, J. R. & Lafferty, K. D. (2004). The Elusive Baseline of Marine Disease: Are Diseases in Ocean Ecosystems Increasing? PLoS Biol, 2:e120. [ Links ]
Ward, J. R., Kim, K. & Harvell, C. D. (2006). Temperature affects coral disease resistance and pathogen growth. Marine Ecology-Progress Series, 329: 115-121. [ Links ]
Zapata, F. A., Rodríguez-Ramírez, A., Caro-Zambrano, C. & Garzón-Ferreira, J. (2010). Mid-term coral-algal dynamics and conservation status of a Gorgona Island (Tropical Eastern Pacific) coral reef. Revista de Biología Tropical, 58: 81-94. [ Links ]
1. Departamento de Ciencias Biológicas-Facultad de Ciencias, Laboratorio de Biología Molecular Marina-BIOMMAR, Universidad de los Andes, Bogotá, Colombia; juansanc@uniandes.edu.co
2. Departamento de Ciencias Biológicas-Facultad de Ciencias, Laboratorio de Biología Molecular Marina-BIOMMAR, Universidad de los Andes, Bogotá, Colombia; Ecomar Consultoria Ambiental, Bogotá, Colombia.
3. Departamento de Ciencias Biológicas-Facultad de Ciencias, Laboratorio de Biología Molecular Marina-BIOMMAR, Universidad de los Andes, Bogotá, Colombia; Ecoral, Medellín, Colombia.
4. Departamento de Ciencias Biológicas-Facultad de Ciencias, Laboratorio de Biología Molecular Marina-BIOMMAR, Universidad de los Andes, Bogotá, Colombia.
5. Programa de Biodiversidad y Ecosistemas Marinos-BEM, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta DTCH, Magdalena, Colombia.
6. Programa de Biodiversidad y Ecosistemas Marinos-BEM, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta DTCH, Magdalena, Colombia.
Recibido 18-X-2013. Corregido 20-XI-2013. Aceptado 19-XII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}