










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.1 San José Feb. 2014
Estructura de la comunidad y distribución vertical de la macrofauna de fondos blandos en isla Gorgona, Pacífico Colombiano
Community structure and vertical distribution of the soft bottom macrofauna on Isla Gorgona, Colombian Pacific
Community structure and vertical distribution of the soft bottom macrofauna on Isla Gorgona, Colombian Pacific
Abstract
Soft bottoms are important in marine ecosystems but their ecology have been less studied in the tropics than in temperate areas. This article describes the temporal, spatial, and vertical distribution of the community structure of subtidal soft bottom macrofauna on Isla Gorgona, Colombian Pacific. Samples were collected in March and June 2011 by corers at 10m depth at five points around the island. Seventy-three species were recorded; polychaetes (55%) and crustaceans (30%) were the dominant groups in number of individuals. The community structure assessed by classification and ordination analysis did not change temporarily, but it shows differences between sampling points, due to the structure of the substrate. The highest abundances of macrofauna were recorded at points with dominance of fine particles (Farallones). However, another point with a similar substrate (Planchón) showed low abundance, apparently due to bioturbation exerted by the fish Heteroconger sp. The presence of Heteroconger sp. in El Planchón could also affect the vertical distribution of the macrofauna, where their relative abundance was homogeneous across the analyzed layers (1-2-3-4-5-10cm). In the other four sites, the abundance of the macrofauna was mainly limited to the first two centimeters of the sediment. Playa Blanca and Remanso showed substrates composed mainly of gravel, recording at the last sampling point the lowest richness, diversity and abundance. Rev. Biol. Trop. 62 (Suppl. 1): 169-188. Epub 2014 February 01.
Key words: macrofauna, community structure, soft bottom, eastern tropical Pacific, Colombia, Isla Gorgona.
Resumen
Se evaluó la variación temporal, espacial y vertical de la estructura comunitaria de la macrofauna de fondos blandos submareales en isla Gorgona, Pacífico colombiano. Las muestras fueron colectadas en marzo y junio de 2011 mediante descorazonadores a 10m de profundidad en cinco puntos alrededor de la isla. Se registraron un total de 73 especies; poliquetos (55%) y crustáceos (30%) fueron los grupos dominantes en número de individuos. La estructura comunitaria evaluada mediante análisis de clasificación y ordenación, no varió temporalmente, pero si presentó diferencias entre puntos de muestreo, al parecer relacionada con la estructura del sedimento. Las mayores abundancias de la macrofauna se registraron en puntos con dominancia de partículas finas (Farallones), pero en otro punto con sustrato similar (Planchón), la abundancia fue baja, al parecer por la bioperturbación ejercida por el pez Heteroconger sp. La presencia de Heteroconger sp. en el Planchón posiblemente afectó la distribución vertical de la macrofauna, ya que en este último punto la abundancia relativa fue homogénea entre las capas analizadas (1-2-3-4-5-10cm). En los otros cuatro sitios, la abundancia de la macrofauna estuvo limitada principalmente a los primeros dos centímetros del sedimento. En Playa Blanca y Remanso el sustrato fue principalmente gravoso, registrándose en el último punto la menor riqueza, diversidad y abundancia.
Palabras clave: macrofauna, estructura de la comunidad, fondos blandos, Pacífico Oriental Tropical, Colombia, isla Gorgona.
Los fondos blandos submareales son claves para el funcionamiento de los ecosistemas marinos. El conocimiento que se tiene sobre este ecosistema proviene de investigaciones realizadas principalmente en zonas templadas, mientras que en zonas tropicales aspectos descriptivos, como la composición de especies, estructurales como los valores de abundancia e incluso las relaciones funcionales son poco conocidas, particularmente en el Pacífico Oriental Tropical (POT) (Zajac, 2008; Mair et al., 2009). Este tipo de información es necesaria para comprender los procesos ecológicos que estructuran una comunidad, además proporcionan elementos para el manejo ambiental y se utilizan para la selección de áreas prioritarias para la conservación (McBreen, Wilson, Mackie & Aonghusa, 2008).
En términos generales, los trabajos desarrollados en fondos blandos submareales evidencian una asociación entre el tipo de sustrato o el tamaño de partícula y la estructura de la comunidad bentónica (Guzmán-Alvis, Lattig & Ruiz, 2006; Zajac, 2008). Esta relación puede condicionar un patrón definido en la distribución de las especies (Guzmán-Alvis & Carrasco, 2005), el cual es principalmente de tipo agregado (Schlacher et al., 1998; Underwood & Chapman, 2005; Hyland et al., 2006). La relación intrínseca entre los organismos bentónicos y el tipo de sustrato se establece principalmente por la oferta de alimento. Esta relación se da en especial para las especies alimentadoras de depósito (Lohrer, Chiaroni, Hewitt & Thrush, 2008; Mair et al., 2009), grupo trófico común en los fondos blandos marinos del trópico en donde generalmente dominan poliquetos y anfípodos (Maurer & Vargas, 1984; Bone & Klein, 2000; López, Cladera, San Martín, Laborda & Aguado, 2002; Lucero, Cantera & Romero, 2006; Mair et al., 2009).
Los estudios de los fondos blandos en Colombia se han desarrollado principalmente en el Caribe (e.g. Guzmán-Alvis & Díaz, 1993; Guzmán-Alvis, Solano, Córdoba-Tejada & López-Rodríguez, 2001; Guzmán-Alvis & Carrasco, 2005; Cortés, 2010), mientras que para el Pacífico estos se reducen en su mayoría a los realizados en las bahías de Buenaventura y Málaga, así como en la desembocadura del río San Juan (e.g. Cantera & Restrepo, 1995; Lucero et al., 2006; Herrera, 2011). Específicamente para isla Gorgona, el estudio realizado por Solano, Cortés & Ruíz (2001), fue el primero en describir las variaciones espacio-temporales de la estructura de la macrofauna de los fondos blandos submareales y su relación con algunas condiciones ambientales durante 1999. Estos autores encontraron que la estructura de la comunidad de la macrofauna no varió entre profundidades o zonas de la isla, pero presentó una tendencia entre épocas climáticas, debido a mayores valores en la abundancia y diversidad durante la época seca. En este sentido, Lohrer et al. (2008), destacan que el transporte de partículas en suspensión durante la época de lluvias afecta negativamente a las especies alimentadoras de depósito (Lohrer et al., 2008), incrementándose este efecto cerca a la línea de costa (Hyland et al., 2006). Posteriormente, Cortés, Solano & Ruíz-López (2012) utilizando otras técnicas de análisis multivariado sobre el mismo conjunto de datos de Solano et al. (2001), identificaron diferencias espaciales en la estructura de la comunidad de la macrofauna de los fondos blandos en isla Gorgona, las cuales relacionaron con cambios en la abundancia de las familias y composición del sedimento entre las zonas evaluadas. Estos autores también reportaron diferencias en la estructura de la comunidad entre los periodos enero-junio y agosto-diciembre a partir de un análisis temporal por zonas.
Isla Gorgona es un Parque Nacional Natural (PNN) de Colombia que simultáneamente es parte integral del Corredor Marino de Conservación y Uso sostenible (CMAR) que abarca diferentes zonas del POT. Estas declaratorias son debidas a su biodiversidad marina y a los ecosistemas estratégicos que presenta, especialmente los arrecifes coralinos, que han sido estudiados y monitoreados durante los últimos 14 años. No obstante, la información sobre la biología y los procesos ecológicos desarrollados en los demás ambientes submareales en general, y suelos blandos en particular son escasos. Por lo tanto, el objetivo del presente estudio fue evaluar la distribución espacial, temporal y vertical de la estructura de la comunidad de la macrofauna de los fondos blandos en isla Gorgona, información que fortalecerá las estrategias de manejo y la implementación de acciones de conservación hacia uno de los valores objeto de conservación de este Parque Nacional Natural, como lo son los fondos blandos submareales.
Materiales y métodos
Área de estudio: Isla Gorgona (2º58’10” N - 78º11’05” W) es el área insular más extensa (13.3km2) en el Pacífico colombiano, ubicada en el sector sur a 30km del continente. Es una isla de origen volcánico, que presenta diferentes unidades geomorfológicas asociadas a la línea de costa, entre las que se destacan planos arenosos en el sector sur y oriental, playas coralinas en el sector oriental y costas rocosas con bloques y cantos rodados en el sector norte y occidental (Llinás, Pinto, Peña & Caro, 1990; Giraldo, 2012). Oceanográficamente se identifican dos periodos contrastantes, un periodo de agua fría y salina entre enero y marzo, y un periodo de agua cálida y de bajas salinidades entre abril y diciembre, los cuales están asociados con el ciclo anual de desplazamiento de la zona de convergencia intertropical sobre el Pacífico colombiano (Giraldo et al., 2008). Estos periodos oceanográficos están estrechamente asociados con las condiciones hidroclimatológicas en esta localidad. En términos generales el clima en Gorgona es de tipo superhúmedo tropical marítimo, con temperatura del aire por arriba de 26°C y porcentaje de humedad relativa cercana al 90%. El tipo de precipitación es unimodal con registros entre 180mm a 400mm mensuales entre enero y marzo, y entre 550mm a 750mm entre abril y diciembre (Rangel & Rudas, 1990; Díaz, Pinzón, Perdomo, Barrios & López-Victoria, 2001; Blanco, 2009).
Procedimiento de muestreo: Las muestras de sedimento fueron recolectadas en marzo (época seca) y junio (época de lluvias) de 2011, en cinco puntos en la zona nororiental, oriental y suroccidental de isla Gorgona (Remanso, Planchón, Azufrada, Playa Blanca y Farallones), y fueron extraídas mediante buceo autónomo a aproximadamente 10m de profundidad teniendo como referencia el nivel de marea mínima. En cada punto se recolectaron tres (marzo) o cinco (junio) muestras de sedimento separadas entre sí de tres a cinco metros, utilizando durante marzo descorazonadores (corers) de acrílico de 10cm de diámetro por 60cm de largo (área total por sitio 0.024m2), y durante junio descorazonadores de policarbonato de 9.5cm de diámetro por 50cm de largo (área total por sitio 0.028m2). En cada punto los descorazonadores fueron introducidos 12cm aproximadamente en el sedimento, y posteriormente en tierra se procedió a dividir los primeros 10cm del núcleo de sedimento en seis capas, las primeras cinco de 1cm de grosor y la última de 5cm de grosor.
Una vez obtenidas las capas del sedimento estas fueron preservadas en formol buferizado al 4%, al cual se le adicionó Rosa de Bengala con el fin de teñir los invertebrados y facilitar así su separación. Después de 24h se procedió a separar los invertebrados, tamizando cada una de las capas con un tamiz de 500µm. Los individuos recolectados fueron preservados en alcohol al 70% hasta su posterior análisis en laboratorio. De los cinco descorazonadores recolectados durante junio en cada sitio, uno fue congelado (-20°C) para realizar posteriormente el análisis de la estructura del sedimento: granulometría, carbonatos y materia orgánica.
Por problemas logísticos durante marzo en el punto Farallones no fue posible colectar todas las muestras, razón por la cual los datos de esta zona para este periodo no se incluyeron en el análisis de datos. Así mismo, por problemas logísticos en la Azufrada en marzo y en Playa Blanca en junio, uno de los descorazonadores no fue divido por capas. En el Remanso cada descorazonador fue procesado como una unidad debido al tipo de sustrato dominante, por lo tanto en este sector no fue posible evaluar la distribución vertical de la macrofauna. Considerando que la variación en el tamaño del área de cada unidad de muestreo entre marzo y junio fue de 0.001m2, se asumió para el análisis de la información que el cambio de tipo de descorazonador entre periodos no afectó las probabilidades de captura de los organismos en los diferentes sitios de muestreo.
Análisis de laboratorio: La identificación se realizó hasta el menor nivel taxonómico posible utilizando claves especializadas en los diferentes grupos taxonómicos (poliquetos, anfípodos, tanaidáceos, decápodos y moluscos) para la región del POT (Keen, 1971; Fauchald, 1977; Cantera et al., 1979; Barnard & Karaman, 1991a; 1991b; Rouse & Pleijel, 2001; Heard et al., 2003; Lazarus, Cobo, Herrera & Satizabal, 2011).
Para la realización de los análisis sedimentológicos todas las muestras fueron secadas en un horno eléctrico a 80°C durante 36h. Para establecer la granulomentría, se tomaron 50g de sedimento de cada una de las capas y se pasaron por una batería compuesta por tamices para gravas gruesas (4.000mm), gravas (2.000mm), arena gruesa (0.850mm), arena media (0.425mm), arena fina (0.250mm), arena muy fina (0.120mm), limos (0.060mm) y arcillas (0.010mm), y posteriormente, cada una de las fracciones se pesó en una balanza analítica (Bale & Kenny, 2005).
Para el análisis de la materia orgánica, se tomaron 10g de sedimento de cada una de las capas y se sometió a combustión en una mufla a 500°C durante 5h, obteniendo el contenido de materia orgánica por diferencia de peso. Para el análisis de carbonatos, se sometieron 20g de sedimento de cada una de las capa a degradación por HCl (5ml, 0.5M), obteniéndose el porcentaje de carbonatos presente por diferencia de peso (Climent & García, 2012).
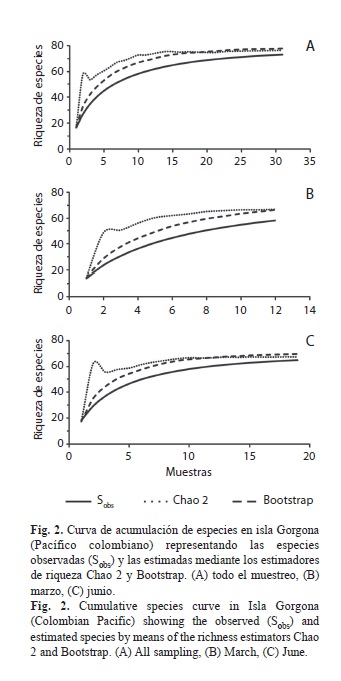
Análisis de datos: Por limitaciones logísticas y presupuestales en este trabajo no fue posible cubrir el área mínima que tradicionalmente es sugerida para cada unidad de muestreo (0.1m2) cuando se realizan estudios de macrofauna de fondos blandos submareales (Eleftheriou & Moore, 2005). Por lo tanto, se evaluó la representatividad del muestreo realizado mediante los estimadores de riqueza Chao 2, el cual se basa en la distribución de las especies entre las muestras considerando aquellas especies que están presentes en una o dos muestras, y Bootstrap el cual considera la proporción de muestras que contienen cada especie (Colwell & Coddington, 1994). Ambos estimadores proporcionan información sobre la riqueza de especies esperada en la zona de estudio, a partir de la información del esfuerzo de muestreo realizado. El cálculo de estos estimadores se realizó utilizando el programa EstimateS 9.1.0 (Colwell, 2013).
La estructura de la comunidad de la macrofauna se evaluó a nivel de familia mediante análisis univariados y multivariados considerando cada núcleo de sedimento como una unidad de muestreo independiente. La similitud en la estructura de la comunidad entre los sitios de muestreo y entre las épocas climáticas se evaluó mediante el índice de similitud de Bray-Curtis, incluyendo únicamente las familias que presentaron una abundancia mayor o igual al 4% en alguno de los puntos de muestreo. A partir de la matriz de similitud obtenida se realizó un análisis de clasificación (cluster) mediante el método de enlace por grupos promediados, y la validez entre las uniones se evaluó mediante la rutina SIMPROF con el programa PRIMER 6.1.13 (Clarke & Warwick, 2001). Después de realizada la rutina SIMPROF se seleccionó en el dendograma un nivel de corte del 50%, toda vez que algunos de los grupos formados mediante el SIMPROF pueden definirse a un nivel muy fino de detalle y no necesariamente presentar importancia biológica (Clarke, Somerfield & Gorley, 2008). Así mismo, se realizó el análisis no métrico multidimensional (nMDS), con el objetivo de representar gráficamente las relaciones entre la estructura de la comunidad. Con el fin de disminuir el efecto de las especies dominantes, los datos de abundancia fueron previamente transformados mediante la raíz cuarta (Clarke & Warwick, 2001).
Los grupos obtenidos en el análisis de clasificación se describieron mediante el análisis SIMPER, con el cual se evaluó cuáles fueron las familias que contribuyeron a la diferenciación de cada grupo, análisis que fue realizado con el programa PRIMER 6.1.13 (Clarke & Warwick, 2001). Se evaluó mediante un ANOVA de una vía sí la abundancia obtenida para los conglomerados presentó diferencias significativas, teniendo en cuenta que se cumplieron con los supuestos de normalidad y homogeneidad de varianzas. Cuando se encontraron diferencias significativas se realizó una prueba a posteriori de Tukey para tamaños de muestra desiguales con el programa STATISTICA 7.0. La distribución vertical de la macrofauna de los fondos blandos se evaluó de forma descriptiva considerando las 10 especies más abundantes durante los dos muestreos.
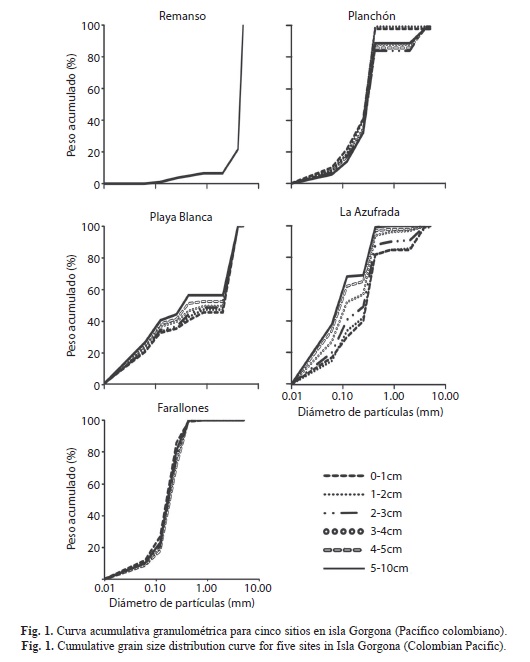
Para el análisis de los sedimentos se desarrollaron curvas granulométricas acumulativas a partir del porcentaje de sedimento que pasó por cada tamiz.
Resultados
Estructura del sedimento: Se identificaron diferencias en la estructura del sedimento entre los sitios de estudio. En el Remanso y Playa Blanca predominaron los sustratos gruesos, con predominio de gravas (14%) y gravas gruesas (78%) en el Remanso (1-10cm del sedimento analizados como una sola unidad), y predominio de gravas (51%) en todas las capas en Playa Blanca, posiblemente por la presencia de fragmentos de coral, que se mezclaron en una baja proporción con sedimentos finos como arenas y limos (Fig. 1). En los otros tres sitios, el principal componente del sedimento en todas las capas fueron las arenas finas (Planchón: 55%, Azufrada: 37%) y muy finas (Farallones: 57%), identificándose en el Planchón dos capas, una superior (0-2cm) con deposición de arenas finas y muy finas, y una inferior (hasta 10cm) con una mezcla de arenas finas, muy finas y un porcentaje bajo de gravas (Fig. 1). Por el contrario, en la Azufrada se encontró una disminución en las gravas y un aumento en limos y arcillas desde la superficie hacia las capas inferiores del sedimento (Fig. 1).
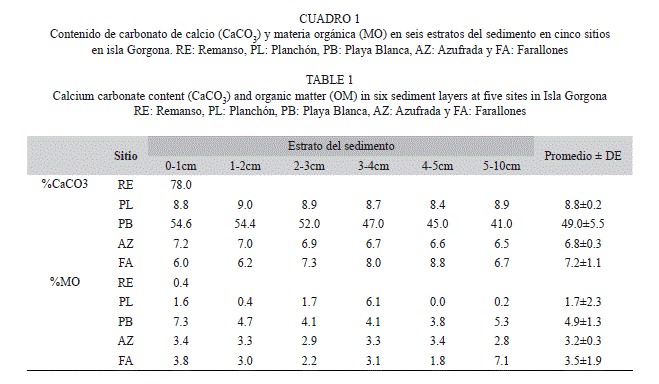
Las diferencias en la estructura del sedimento entre los sitios de muestreo estuvieron acompañadas de variaciones en la presencia de estructuras biológicas de carbonato de calcio (CaCO3) y contenido de materia orgánica (MO). De esta forma, el Remanso y Playa Blanca presentaron altas concentraciones de CaCO3, como resultado de la presencia de fragmentos de coral en los dos sitios, así como de rocas con algas calcáreas en el Remanso (Cuadro 1). En contraste, la Azufrada, el Planchón y Farallones se caracterizaron por presentar concentraciones bajas de CaCO3, debido a la degradación de pequeños fragmentos erosionados de coral (Azufrada), espículas de equinodermos (Azufrada y Planchón), y conchas degradadas de moluscos y bivalvos (los tres sitios) (Cuadro 1). En general, la concentración del CaCO3 presentó poca variación entre las capas del sedimento.
El contenido de materia orgánica (MO) varió entre los sitios de muestreo, presentando concentraciones bajas en el Remanso y Planchón, y concentraciones intermedias en la Azufrada, Farallones y Playa Blanca (Cuadro 1). Verticalmente, la distribución de la MO fue variable en el Planchón y en Farallones, registrándose las mayores concentraciones en el cuarto centímetro y última capa del sedimento, respectivamente (Cuadro 1). En contraste, en Playa Blanca y la Azufrada, la distribución vertical de la MO fue homogénea entre las diferentes capas (Cuadro 1). Igualmente, es importante destacar que sólo en el Planchón se detectó material vegetal fragmentado a lo largo de la columna de sedimento.
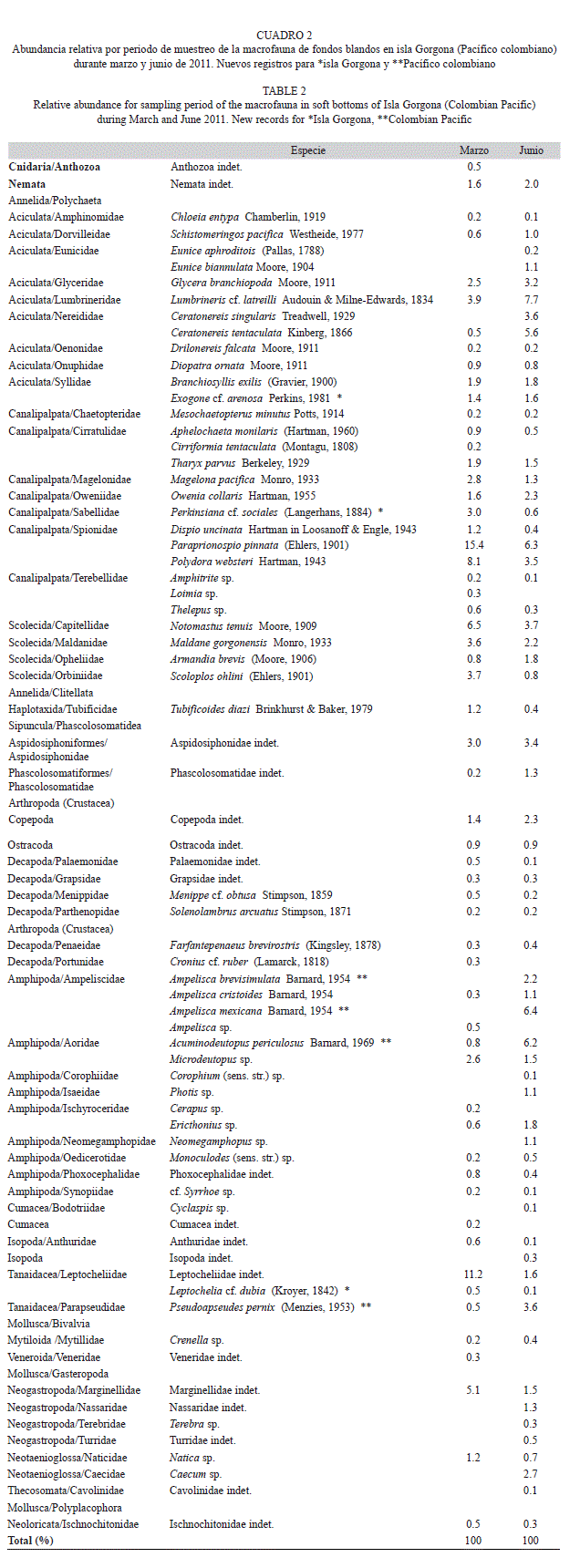
Estructura de la comunidad: Durante el periodo de estudio se recolectaron 2 230 individuos (654 en marzo y 1 576 en junio) pertenecientes a 73 especies en 58 familias. De acuerdo a los estimadores de riqueza Chao 2 y Bootstrap, la representatividad del muestreo considerando los dos periodos de estudio fue entre el 94% (Bootstrap) y el 96% (Chao 2). La representatividad del muestreo en junio fue del 94% (Bootstrap) y 96% (Chao 2), mientras que para el mes de marzo fue del 87% según ambos estimadores (Fig. 2)
La riqueza de especies de la macrofauna de fondos blandos en isla Gorgona fue mayor durante junio (65 especies), donde los poliquetos (27 especies en 20 familias, 53% de los individuos) y los crustáceos (25 especies en 20 familias, 32% de los individuos) fueron los grupos dominantes en términos del número de especies y de individuos (Cuadro 2). Aunque durante marzo la riqueza de especies fue menor (58 especies), también los poliquetos (26 especies en 18 familias, 63% de los individuos) y crustáceos (22 especies en 18 familias, 23% de los individuos) fueron los grupos dominantes (Cuadro 2). Los moluscos, que generalmente son un componente importante de los fondos blandos submareales, presentaron una contribución menor durante los dos periodos evaluados en el presente estudio (marzo: cinco especies en cinco familias, 7% de los individuos; junio: nueve especies en nueve familias, 8% de los individuos). Las especies de poliquetos, Exogone cf. arenosa, Mesochaetopterus minutus, Perkinsiana sociales y Notomastus tenuis, así como los crustáceos, Ampelisca brevisimulata, Ampelisca mexicana, Acuminodeutopus periculosus, Leptochelia cf. dubia y Pseudoapseudes pernix, constituyen nuevos registros para isla Gorgona.
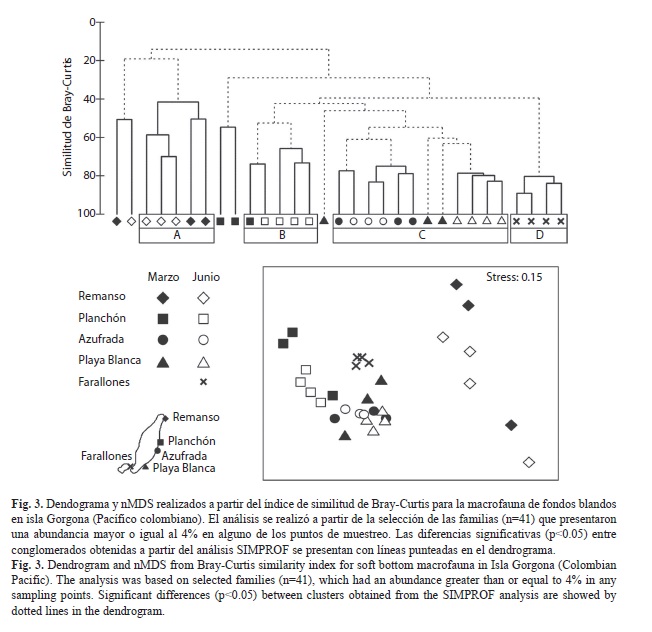
La estructura de la comunidad de la macrofauna de los fondos blandos en isla Gorgona varió significativamente entre los sitios de muestreo pero no entre las épocas climáticas (SIMPROF, p<0.05), ya que las muestras se agruparon por zonas independientemente del periodo de muestreo (Fig. 3). De los nueve conglomerados obtenidos mediante el análisis de clasificación, sólo se consideraron cuatro para análisis posteriores, correspondientes a aquellos grupos que presentaron estructura multivariada (SIMPROF, p<0.05), y que adicionalmente, en los casos donde el nivel de división obtenido mediante el SIMPROF fue muy fino, presentaron un nivel de similitud mayor o igual al 50%. Este resultado fue consecuente con el obtenido mediante el análisis de ordenación, ya que las muestras de los grupos A (Remanso), B (Planchón) y D (Farallones) se separaron claramente en el análisis de ordenación, mientras que las muestras de las zonas arrecifales (grupo C) se presentaron como un solo conglomerado en este diagrama (Fig. 3).
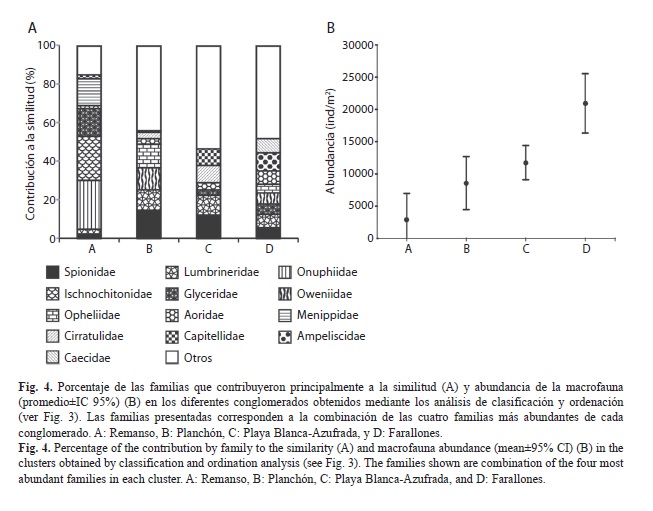
De los cuatro conglomerados analizados, se encontró que el grupo A estuvo conformado por dos muestras obtenidas en marzo y tres obtenidas en junio en el Remanso (Fig. 3). Este grupo presentó la mayor disimilitud con respecto a los otros tres grupos obtenidos, en términos de la composición de las familias y su abundancia (SIMPER, A vs. B: 88%, A vs. C: 81%, A vs. D: 81%). En este sentido, el grupo A se caracterizó por presentar el menor número de familias (17) y los menores valores de abundancia, así como porque las familias que contribuyeron principalmente a su similitud fueron los poliquetos Onuphidae (26%) y Glyceridae (14%), los poliplacóforos Ischnochitonidae (23%) y los decápodos Menippidae (14%) (Fig. 4).
El grupo B estuvo conformado por una muestra obtenida en marzo y cuatro muestras obtenidas en junio en el Planchón (Fig. 3), y presentó un porcentaje de disimilitud del 57% con el grupo C y del 61% con el grupo D. Este grupo se caracterizó por presentar valores intermedios de riqueza de familias (24) y abundancia, y porque cuatro familias de poliquetos, Spionidae (14%), Opheliidae (12%), Oweniidae (11%) y Lumbrineridae (11%) fueron las que contribuyeron principalmente a su similitud (Fig. 4). Por su parte, el grupo C estuvo conformado por una combinación de muestras obtenidas en ambas zonas arrecifales (Azufrada: tres en marzo y tres en junio, Playa Blanca: dos en marzo y cuatro en junio) (Fig. 3), y presentó un porcentaje de disimilitud del 61% con el grupo D. Este grupo presentó la mayor riqueza de familias (28) y valores intermedios de abundancia, y las familias de poliquetos Spionidae (12%), Lumbrineridae (10%), Cirratulidae (9%) y Capitellidae (9%) fueron las que contribuyeron principalmente a su similitud (Fig. 4).
El último de los conglomerados (grupo D) estuvo conformado por las cuatro muestras obtenidas en junio en Farallones (Fig. 3). Este grupo se caracterizó por presentar valores intermedios de riqueza de familias (22), los mayores valores de abundancia, así como porque dos familias de anfípodos, Ampeliscidae (9%) y Aoridae (7%), una de gasterópodos, Caecidae (7%), y una de poliquetos, Lumbrineridae (7%), fueron las que contribuyeron principalmente a su similitud (Fig. 4). La abundancia presentó valores mayores en el grupo D (Farallones) con respecto a los otros tres grupos analizados (F=12.94, p<0.01), y también fue mayor en el grupo C (zonas arrecifales) que en el grupo A (Remanso) (F=12.94, p=0.02).
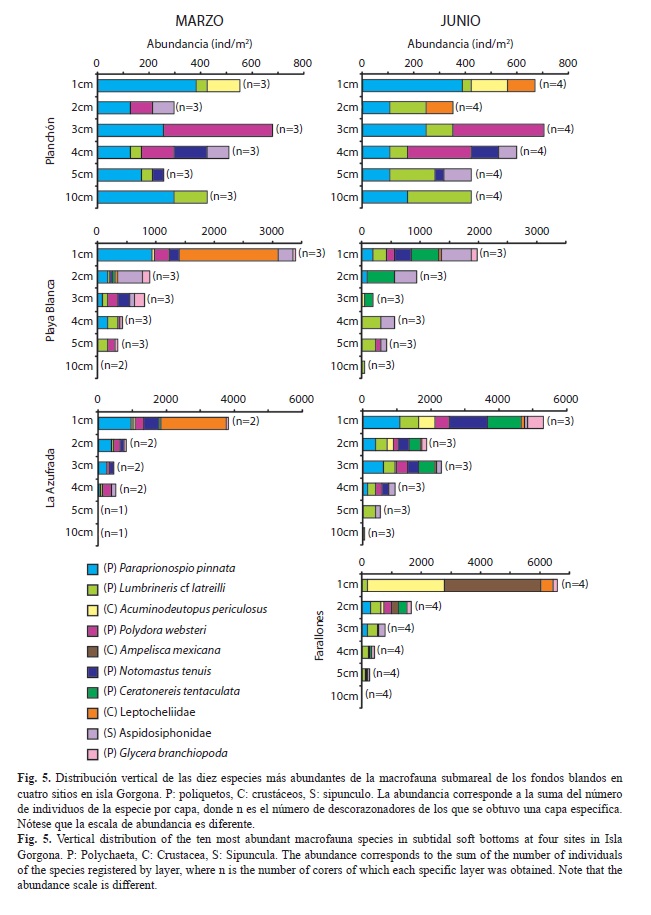
Distribución vertical: La distribución vertical de la macrofauna en los fondos blandos de la Azufrada, Playa Blanca y Farallones en isla Gorgona presentó, tanto en marzo como en junio, más del 70% de la abundancia en los primeros tres centímetros del sedimento, como resultado de la mayor abundancia de poliquetos y crustáceos en estas capas (Fig. 5). Durante marzo, en Playa Blanca y la Azufrada el tanaidáceo Leptocheliidae (50% de la abundancia en ambos sitios) y el poliqueto Paraprionospio pinnata (25% y 28%, respectivamente) fueron las especies dominantes en el primer centímetro de sedimento (Fig. 5). En esta misma capa durante junio, la abundancia estuvo repartida principalmente entre el sipuncúlido Aspidosiphonidae (26%) y los poliquetos Ceratonereis tentaculata (24%) y Notomastus tenuis (14%) en Playa Blanca, mientras que en la Azufrada, la abundancia estuvo repartida entre los poliquetos N. tenuis (21%), P. pinnata (20%) y C. tentaculata (19%) (Fig. 5). Después del cuarto centímetro de sedimento tanto en Playa Blanca como en la Azufrada la abundancia de la macrofauna disminuyó a menos del 10% de la abundancia total. En estas últimas capas, la abundancia estuvo representada principalmente por poliquetos, destacándose que la especie Lumbrineris cf. latreilli presentó la mayor abundancia relativa o incluso fue la única especie que estuvo presente en el quinto y del quinto al décimo centímetro del sedimento (Fig. 5). En Farallones durante junio, se encontró que el 85% de la abundancia de la macrofauna se presentó en los primeros dos centímetros del sedimento, la cual estuvo representada principalmente por los anfípodos Ampelisca mexicana (42%) y Acuminodeutopus periculosus (33%) (Fig. 5).
Por el contrario, en el Planchón (marzo y junio), la distribución vertical de la macrofauna fue homogénea entre las diferentes capas, siendo este en el único sitio donde más del 40% de la abundancia se presentó después del cuarto centímetro de sedimento (Fig. 5). En esta localidad, el poliqueto P. pinnata fue la especie más abundante (50% en marzo y 42% en junio), la cual, aunque estuvo presente en todas las capas analizadas, presentó las mayores abundancias en el primer centímetro del sedimento. Por su parte, el poliqueto Polydora websteri se registró sólo entre el segundo y cuarto centímetro del sedimento, presentando las mayores abundancias en el tercer centímetro, mientras que L. cf. latreilli aunque se registró en capas superficiales, presentó las mayores abundancias del quinto al décimo centímetro de sedimento (Fig. 5).
Discusión
Estructura de la comunidad: La comunidad de macroinvertebrados de fondos blandos submareales en isla Gorgona estuvo representada principalmente por poliquetos (40% de las especies, 55% del número de individuos) y crustáceos (40% de las especies, 30% del número de individuos). Este patrón es consistente con lo descrito por Solano et al., (2001) para la isla, quienes registraron a los poliquetos como el grupo macrobentónico más representativo en familias y morfotipos de especies, con baja presencia de otros grupos. Igualmente, la dominancia en número de individuos de poliquetos y crustáceos en los fondos blandos submareales ha sido reportada para otras localidades tropicales, como el Archipiélago Las Perlas (Golfo de Panamá) (poliquetos: 74%, crustáceos: 14%) (Mair et al., 2009), el Golfo de Nicoya (poliquetos: 68%, crustáceos: 21%) (Maurer & Vargas, 1984) y el Golfo Dulce en Costa Rica (poliquetos: 89%, crustáceos: 7%) (León-Morales & Vargas, 1998), el Golfo Triste en Venezuela (Bone & Klein, 2000) y el Golfo de Salamanca en el Caribe colombiano (Cortés, 2010), aunque contrario a lo reportado por Guevara-Fletcher, Cantera, Mejía-Ladino & Cortés (2011) para un estuario tectónico con fondo rocoso en el Pacífico colombiano (Bahía Málaga), donde moluscos y crustáceos dominaron la comunidad. A pesar de que no se cubrió el área mínima de muestreo sugerida en la literatura científica para el estudio de fauna presente en fondos blandos submareales, los resultados obtenidos fueron consecuentes con lo reportado previamente para isla Gorgona e incluso para otras localidades tropicales. En este sentido, se destaca la representatividad obtenida para el muestreo, registrándose más del 85% de las especies esperadas.
Los organismos bentónicos submareales presentan un papel clave en los ecosistemas marinos ya que son los encargados de reciclar las partículas y fragmentos de materia orgánica que se deposita en el sedimento (Mann, 2000). La dependencia directa de estos organismos a la disponibilidad de materia orgánica externa, permiten identificar el modelo trófico dominante como de cascada-arriba o bottom-up, ya que la fuente de energía que alimenta el sistema llega en forma de pulsos temporales, modulando las interacciones tróficas y delimitando la estructura de la comunidad (Turner & Millward, 2002; Giménez, Dimitriadis, Carranza, Borthagaray & Rodríguez, 2006; Gillet, Mouloud, Durou & Deutsch, 2008). Las especies registradas para isla Gorgona en el presente estudio se pueden clasificar en seis grupos tróficos: carnívoros, filtradores, depositarios no selectivos, depositarios selectivos, herbívoros y omnívoros (grupos establecidos a partir de López & Levinton, 1987; Macdonald et al., 2010). Las especies depositarias en conjunto conforman el grupo más rico y abundante, seguido por los carnívoros. Este resultado concuerda con lo esperado, ya que la posible llegada de material orgánico particulado por pulsos, favorece el desarrollo poblacional de especies que responden rápidamente a estos, y sugieren una ventaja de usar directamente el sedimento como fuente de alimento (Alongi, 1989; Elías, Vallarinot, Scagliolat & Isla, 2004). Adicionalmente, la presencia de especies carnívoras sugiere depredación de grupos taxonómicos como microcrustáceos, nematodos, o estados juveniles de macroinvertebrados.
La estructura de la comunidad de la macrofauna de los fondos blandos submareales en isla Gorgona durante el presente estudio varió entre los sitios de muestreo pero no entre las épocas climáticas. Este resultado contrasta con lo reportado por Solano et al. (2001) para la isla, quienes encontraron variaciones a nivel estacional pero no a nivel espacial, asociadas a mayores abundancias de la macrofauna durante los primeros meses del año (época seca), coincidiendo a su vez con la época de mayor productividad en la columna de agua. No obstante, en un análisis posterior a los datos utilizados por estos autores, Cortés et al. (2012) encontraron que la estructura de la comunidad de la macrofauna varió entre los sitios de muestreo, concordando con lo registrado en el presente estudio. Adicionalmente, estos autores realizaron un análisis de clasificación y ordenación a partir de los grupos espaciales obtenidos (Punta Brava-Yundigua-La Máncora vs. Remanso), encontrando que la estructura de la comunidad fue diferente entre meses, con un primer grupo formado por los meses de enero a junio, y un segundo grupo conformado por los meses de agosto a diciembre. De acuerdo con estos autores, las variaciones temporales en la estructura de la comunidad se debieron a la disminución de cienos y materia orgánica en el sedimento (oferta de alimento) durante el periodo agosto-diciembre. La ausencia de variación estacional en la estructura de la comunidad de la macrofauna durante el presente estudio, sugiere que las variaciones estacionales en las condiciones ambientales de la columna de agua (e.g. temperatura, salinidad, oxígeno disuelto, fitoplancton, zooplancton) posiblemente no sean el principal factor que modula las variaciones en la composición y abundancia de la macrofauna en los fondos blandos de isla Gorgona. Por lo tanto, se hace necesario evaluar el efecto de factores puntuales como la composición del sedimento o el contenido de materia orgánica y carbonatos sobre esta comunidad.
Las variaciones temporales en la abundancia relativa de la macrofauna de los fondos blandos submareales también pueden estar moduladas por procesos biológicos como eventos reproductivos, mayor disponibilidad de alimento, depredación, competencia, u otro tipo de interacciones (Woodin, 1976; Wilson, 1991). En este sentido, la presencia de hembras ovadas durante la época seca del tanaidáceo Leptocheliidae en Playa Blanca y la Azufrada, así como el aumento de la abundancia de los poliquetos nereididos Ceratonereis tentaculata y Ceratonereis singularis en Playa Blanca durante la época de lluvias, sugieren que eventos reproductivos fueron el factor que influenció principalmente el cambio temporal en la abundancia de estas especies. Por el contrario, la alta dominancia del poliqueto Paraprionospio pinnata en este estudio, sugiere que los organismos de esta especie presentan adaptaciones que les permiten explotar fuentes repentinas de alimento y ser competitivamente superiores, lo cual al parecer se relaciona con su forma de alimentación (especie depositaria superficial) y sus altas tasas de ingestión. Esto podría explicar la alta dominancia que ha sido registrada para esta especie en otras localidades tropicales (e.g. Delgado-Blas, 2001) y templadas (e.g. Muñoz & Salamanca, 2001).
La variación espacial en la estructura de la comunidad de la macrofauna de fondos blandos registrada para isla Gorgona en esta investigación sugiere que las características del sedimento en cada uno de los sitios de muestreo posee una identidad espacial particular que determina el tipo y el número de organismos que la habitan. En otras localidades del Pacífico colombiano (e.g. Cantera et al., 1979; Herrera, 2002; Lucero et al., 2006; Guevara-Fletcher et al., 2011; Herrera 2011), y en diferentes ambientes tropicales (e.g. López et al., 2002; Guzmán-Alvis et al., 2006; Mair et al., 2009; Cortés 2010), la composición del sedimento ha sido identificado como el principal factor que modula la estructura de las comunidades de los fondos blandos submareales. En este sentido, la estructura del sedimento en Farallones, la Azufrada y el Planchón, presentaron una predominancia de partículas finas, lo cual es característico de ambientes con poca energía en el movimiento de aguas permitiendo su decantación. Esto podría explicar las mayores abundancias registradas en Farallones y las zonas arrecifales, con comunidades compuestas principalmente por organismos que se alimentan de partículas en suspensión y alimentadores depositarios (e.g. Ampeliscidae y Spionidae, respectivamente). Este resultado coincide con lo reportado por Mair et al. (2009) para el Archipiélago Las Perlas (Golfo de Panamá), zona con características similares a isla Gorgona (e.g. presencia de arrecifes coralinos y rocosos), donde altos valores de diversidad y abundancia se asociaron con sedimentos arenosos con presencia de conchas.
La principal característica que permitió que Farallones se presentara como una comunidad con una estructura particular y significativamente diferente a la de los otros sitios de muestreo, fue la alta abundancia de crustáceos (64%), siendo las especies más representativas Ampelisca mexicana (Ampeliscidae) y Acuminodeutopus periculosus (Aoridae). Aunque no se tiene conocimiento sobre la biología de estas especies, en general las especies de la familia Ampeliscidae y de la familia Aoridae, se caracterizan por construir tubos utilizando partículas del sedimento, así como por alimentarse de agregados de detritos y de material en suspensión (Barnard & Karaman, 1991a). De acuerdo con Woodin (1976) y Wilson (1991), cuando el crecimiento y abundancia de organismos depositarios (e.g. Paraprionospio pinnata - Spionidae) es limitado como en el caso de Farallones, es posible que se favorezca el crecimiento e incremento en abundancia de organismos filtradores (e.g. A. mexicana y A. periculosus).
El sector de Farallones es una zona donde no hay una aparente fuente directa de materia orgánica desde sistemas aledaños, a diferencia de la Azufrada y Playa Blanca que se encuentran en una zona de alta productividad (arrecifes coralinos) y del Planchón que recibe grandes cantidades de materia orgánica provenientes de la desembocadura de riachuelos, por lo que los organismos bentónicos tienen en la columna de agua su principal fuente de alimento. Este resultado es consecuente con lo reportado por Mair et al. (2009) para la zona central del Archipiélago Las Perlas, donde las mayores abundancias de crustáceos (familia Ampeliscidae) en los fondos blandos, estuvieron asociadas con sedimentos que presentaron una mezcla de partículas como arena, lodo y conchas. Otro elemento a considerar en el sector de Farallones, es la posible baja presión por depredación, ya que a diferencia de la Azufrada y Playa Blanca (arrecifes coralinos), el Planchón (praderas de anguilas) y el Remanso (sustrato rocoso) en donde se observan altas densidades de peces, en Farallones las densidades de peces son bajas posiblemente por la homogeneidad del sustrato (gran extensión de fondos arenosos). De acuerdo con Maurer & Vargas (1984), la presión por depredación por peces puede disminuir la diversidad y abundancia de la macrofauna de fondos blandos submareales, como lo reportan para la zona estuarina del Golfo de Nicoya (Costa Rica).
En contraste, en los otros cuatro sitios de muestreo los poliquetos fueron el grupo que contribuyó principalmente a la abundancia (72-75%). No obstante, las variaciones en la composición de las familias y la abundancia relativa permitieron que se formaran comunidades con estructuras características, excepto para las zonas arrecifales (Azufrada y Playa Blanca) que se presentaron como un solo conglomerado. En estas zonas al parecer fue más importante en estructurar la comunidad el tipo de sistema que la estructura del sedimento en el punto particular de muestreo, ya que la estructura del sedimento en la Azufrada fue principalmente fino con bajas concentraciones de carbonato de calcio y sin presencia de fragmentos de coral, mientras que en Playa Blanca el sedimento presentó un alto contenido de gravas, fragmentos de coral y altas concentraciones de carbonato de calcio. Esto posiblemente se debió a que las muestras en la Azufrada fueron recolectadas en una zona del arrecife que presenta una depresión en forma de cráter donde el crecimiento coralino es limitado (Zapata, 2001). No obstante, al estar inmerso el punto de muestreo dentro del arrecife, la composición de la macrofauna posiblemente estuvo influenciada por este.
Aunque en el Planchón al igual que en Farallones el tipo de sedimento estuvo compuesto principalmente por arenas finas a muy finas, la abundancia de la macrofauna submareal fue baja. Es posible que la presencia de peces del género Heteroconger sp. (Congridae) estén ejerciendo un fuerte efecto de bioperturbación, ya que estos con sus movimientos ondulatorios desestabilizan el sedimento al salir y volver a enterrarse. La bioperturbación es la alteración de la estructura del sedimento debida a la actividad de los organismos (e.g. al enterrarse o alimentarse), lo cual puede ocasionar la oxigenación de capas inferiores del sedimento, así como cambios en la distribución vertical del tamaño del grano del mismo (Wilson, 1991; Reise, 2002). No obstante, el efecto de bioperturbación provocado por algunos peces y decápodos, puede ejercer un efecto negativo sobre la abundancia de organismos suspensívoros y constructores de tubos por sofocación (e.g. los poliquetos Spionidae) (Wilson, 1991), lo cual posiblemente esté ocurriendo en el Planchón.
Por su parte, en el Remanso debido al tipo de sustrato (>80% gravas gruesas, bajas concentraciones de materia orgánica <0.5%), la comunidad de la macrofauna presentó las mayores diferencias con respecto a los otros cuatro sitios evaluados. En esta zona, la riqueza de familias y la abundancia fueron significativamente menores, donde la alta porosidad del sustrato favoreció la presencia de especies reptantes. Las especies más abundantes registradas en este sitio fueron tanto fauna que habita al interior del sedimento, como por ejemplo los poliquetos Onuphidae que habitan en tubos y Eunicidae que pueden excavar sustratos de carbonato de calcio (Fauchald, 1977), como fauna que habita asociada a sustratos duros como rocas y cascajo de coral (e.g. el quitón Ischnochitonidae). Al igual que lo registrado en el presente estudio, Cortés et al. (2012) encontraron que el Remanso fue el sector que presentó la mayor diferencia en la estructura de la comunidad con respecto a los otros sectores evaluados. No obstante, estos autores registraron el mayor número de familias y abundancia en el Remanso, lo cual atribuyeron a la mayor presencia de gránulos y heterogeneidad del sustrato.
Distribución vertical: La distribución vertical de la fauna en los fondos blandos submareales se encuentra en general limitada por las condiciones químicas y biológicas del sedimento, principalmente disponibilidad de oxígeno y alimento, los cuales usualmente se presentan en mayores concentraciones en los primeros centímetros del sedimento (Giere, 2009). Por este motivo, la mayor abundancia de la macrofuana en los fondos blandos submareales tanto en ambientes tropicales (e.g. Cortés, 2010) como templados (e.g. Hines & Comtois, 1985; Smith & Kukert, 1996; Rosenberg et al., 2001) se presenta en las capas superficiales del sedimento (0-5cm), como fue registrado en Playa Blanca, Azufrada y Farallones en isla Gorgona. Así mismo, la distribución vertical de los poliquetos y crustáceos encontrada para estas localidades coincide con lo reportado en otros estudios, donde los poliquetos aunque tienden a presentar su mayor abundancia en la capa superficial del sedimento, pueden presentar bajo condiciones particulares una mayor amplitud en la distribución vertical a la presentada por los crustáceos, en los cuales la distribución vertical se limita en general a la capa superficial (0-1cm) (e.g. Hines & Comtois, 1985; Cortés, 2010). Esto parece estar relacionado también con la forma de alimentación, ya que los crustáceos registrados en este estudio son principalmente filtradores y por lo tanto requieren estar en contacto directo con la columna de agua, mientras que los poliquetos más abundantes fueron alimentadores de depósito de superficie y subsuperficie.
En contraste, las abundancias relativas mayores hacia capas más profundas del sedimento pueden ser ocasionadas por el estilo de vida particular de algún organismo que esté produciendo un cambio en las características verticales del sedimento (bioperturbación) (Wilson, 1991). Es posible que la presencia del pez Heteroconger sp. en los fondos blandos submareales en el sector del Planchón, esté facilitando la oxigenación, así como la distribución del alimento hacia capas más profundas del sedimento. Esto es consecuente con los resultados obtenidos para la estructura del sedimento en este sitio, pues sólo en el Planchón se registró una alta abundancia relativa de materia orgánica en el cuarto centímetro del sedimento, y sólo en este sitio se registraron fragmentos vegetales a lo largo de todas las capas del sedimento. Posiblemente, estas características particulares les permitió a los poliquetos Paraprionospio pinnata, Lumbrineris cf. latreilli y Polydora websteri habitar en capas más profundas. Por lo tanto en el Planchón, la presencia del pez Heteroconger sp. podría estar favoreciendo una distribución más amplia de la macrofauna a lo largo del sedimento, aunque en conjunto con la presencia de detritus vegetal podría estar afectando su abundancia (Alongi & Christoffersen, 1992; Wilson, 1991).
Es importante que futuros estudios que evalúen los factores que modulan la estructura de la comunidad de los fondos blandos submareales en isla Gorgona, consideren de acuerdo a lo propuesto por Alongi (1989) para el estudio de fondos blandos en regiones tropicales, el efecto de las descargas de agua dulce, el acoplamiento bento-pelágico, la química del sedimento (e.g. reciclaje de nutrientes), tolerancias fisiológicas a temperatura y salinidad, producción secundaria, así como estudios tróficos entre otros. Igualmente, como lo reportaron Mair et al. (2009) para el Archipiélago Las Perlas (Golfo de Panamá), la importancia de los estudios de la estructura de la macrofauna de fondos blandos submareales y su relación con las características físico-químicas y biológicas de los sedimentos, permiten entender los factores que modulan la distribución, diversidad, abundancia y tipos de comunidad de estos sistemas, información que es indispensable para cualquier Área Marina Protegida como el PNN Gorgona, y que contribuye con el desarrollo e implementación de estrategias apropiadas de manejo que asegurarán una protección adecuada de cada hábitat o comunidad representativa.
En conclusión, la estructura de la comunidad de la macrofauna de los fondos blandos submareales en isla Gorgona estuvo conformada principalmente por poliquetos y crustáceos, y presentó variaciones a nivel espacial pero no entre épocas climáticas. Esta variación al parecer es producto de las diferencias en la estructura del sedimento entre los diferentes sectores de la isla. Igualmente, la distribución vertical de la macrofauna estuvo limitada principalmente hacia las capas superficiales del sedimento en Farallones, Playa Blanca y Azufrada, mientras que en el Planchón la distribución fue amplia a lo largo de la columna del sedimento (1-10cm), al parecer por la bioperturbación ejercida por el pez Heteroconger sp.
Agradecimientos
A Selene Escobar, Héctor Chirimía, Luis Felipe Piñeros, Natalia Uribe, Katherine Mejía y Andrés Cuellar por su colaboración durante la toma de muestras. A los funcionarios del PNN Gorgona por su apoyo logístico durante los muestreos. A Ángela Guzmán (Universidad Nacional de Colombia) por el préstamo de los colectores para la toma de muestras durante marzo de 2011. A Luz Ángela López de Mesa por su colaboración en la identificación de los moluscos. A la estación científica Henry von Prahl del PNN Gorgona por facilitar el espacio requerido para el procesamiento inicial de las muestras. A la Universidad ICESI por facilitar el laboratorio para el análisis de carbonatos. Dos evaluadores anónimos contribuyeron con sus sugerencias a mejorar el presente documento. La UAESPNN permitió el desarrollo de este proyecto en el PNN Gorgona bajo el permiso de investigación PIBD-DTSO-011-10. Este trabajo es un producto del proyecto de investigación “Evaluación del estado actual de los objetos de conservación faunísticos en Isla Gorgona: una aproximación holística a la valoración ecológica del PNN Gorgona” y fue financiado por Conservación Internacional Colombia, Fondo para la Acción Ambiental y la Niñez, Universidad del Valle y Fundación Squalus.
Referencias
Alongi, D. M. (1989). Ecology of tropical soft-bottom benthos: a review with emphasis on emerging concepts. Revista de Biología Tropical, 37: 85-100. [ Links ]
Alongi, D. M. & Christoffersen, P. (1992). Benthic infauna and organism-sediment relations in a shallow, tropical coastal area: influence of outwelled mangrove detritus and physical disturbance. Marine Ecology Progress Series, 81: 229-245. [ Links ]
Bale, A. J. & Kenny, A. J. (2005). Sediment analysis and seabed characterization. In A. Eleftheriou & A. McIntyre (Eds.), Methods for the study of marine benthos (pp. 43-86). Wiley-Blackwell. London, United Kingdom. [ Links ]
Barnard, J. L. & Karaman, G. S. (1991a). The Families and genera of marine gammaridean Amphipoda (except marine gammaroids). Part 1. Records of the Australian Museum, 13: 1-417. [ Links ]
Barnard, J. L. & Karaman, G. S. (1991b). The Families and genera of marine gammaridean Amphipoda (except marine gammaroids). Part 2. Records of the Australian Museum, 13: 419-866. [ Links ]
Blanco, J. F. (2009). The hydroclimatology of Gorgona Island: seasonal and ENSO-related patterns. Actualidades Biológicas, 31: 111-121. [ Links ]
Bone, D. & Klein, E. (2000). Temporal variations in a tropical soft-bottom community, Venezuela. Journal of Coastal Research, 16: 278-286. [ Links ]
Cantera, J. R., Rubio, E. A., Borrero, F. J., Contreras, R., Zapata, F. & Buttkus, E. (1979). Taxonomía y distribución de los moluscos litorales de la Isla Gorgona, Colombia. In H. Von Prahl, M. Grögl & F. Guhl. (Eds.), Gorgona (p. 141-168). Universidad de Los Andes. Bogotá, Colombia. [ Links ]
Cantera, J. R. & Restrepo, J. D. (1995). (Eds.). Delta del río San Juan, Bahías de Málaga y Buenaventura, Pacífico Colombiano. Tomo II. Universidad EAFIT-Universidad del Valle. Medellín, Colombia. [ Links ]
Clarke, K. R & Warwick, R. M. (2001). Change in marine communities: an approach to statistical analysis and interpretation. Primer-E. Plymouth, United Kingdom. [ Links ]
Clarke, K. R., Somerfield, P. J. & Gorley, R. N. (2008). Testing of null hypotheses in exploratory community analyses: similarity profiles and biota-environment linkage. Journal of Experimental Marine Biology and Ecology, 366: 56-69. [ Links ]
Climent, M. T. & García, B. (2012). Cálculos estequiométricos: reacción entre carbonato de calcio y ácido clorhídrico. Retrieved from http://roderic.uv.es/handle/10550/25937. [ Links ]
Colwell, R. K. & Coddington, J. A. (1994). Estimating terrestrial biodiversity through extrapolation. Transactions of the Royal Society Biological Sciences, 345: 101-118. [ Links ]
Colwell, R. K. (2013). EstimateS: Statistical estimation of species richness and shared species from samples. Version 9.1.0. User’s Guide and application. Retrieved from http://purl.oclc.org/estimates. [ Links ]
Cortés, F. A. (2010). Estratificación vertical y distribución horizontal de la comunidad macroinfaunal sublitoral en los sedimentos marinos (Tesis de Maestría). Universidad Nacional de Colombia, Colombia. [ Links ]
Cortés, F. A., Solano, O. D. & Ruíz-López, J. A. (2012). Variación espacio-temporal de la fauna macrobentónica asociada a fondos blandos y su relación con factores ambientales en el Parque Nacional Natural Gorgona, Pacífico colombiano. Boletín de Investigaciones Marinas y Costeras, 41: 323-353. [ Links ]
Delgado-Blas, V. H. (2001). Distribución espacial y temporal de poliquetos (Polychaeta) bénticos de la plataforma continental de Tamaulipas, Golfo de México. Revista de Biología Tropical, 49: 141-147. [ Links ]
Díaz, J. M., Pinzón, J. H., Perdomo, A. M, Barrios, L. M. & López-Victoria, M. (2001). Generalidades. In L. M. Barrios & M. L. Victoria (Eds.). Gorgona marina: Contribución al conocimiento de una isla única (pp. 17-26.). INVEMAR, Serie de Publicaciones especiales N° 7. Santa Marta, Colombia. [ Links ]
Eleftheriou, A. & Moore, D. C. (2005). Macrofauna techniques. In A. Eleftheriou & A. McIntyre (Eds.), Methods for the study of marine benthos (pp. 160-228). Wiley-Blackwell. London, United Kingdom. [ Links ]
Elías, R., Vallarinot., E. A., Scagliolat, M. & Isla, F. I. (2004). Macrobenthic distribution patterns at a sewage disposal site in the inner shelf off Mar del Plata (SW Atlantic). Journal of Coastal Research, 20: 1176-1182. [ Links ]
Fauchald, K. (1977). Polychaetes from intertidal areas in Panama, with a review of previous shallow-water records. Smithson. Contributions to Zoology, 221: 1-81. [ Links ]
Giere, O. (2009). Meiobenthology: the microscopic motile fauna of aquatic sediments. Springer. USA. [ Links ]
Gillet, P., Mouloud, M., Durou, C. & Deutsch, B. (2008). Response of Nereis diversicolor population (Polychaeta, Nereididae) to the pollution impact Authie and Seine estuaries (France). Estuarine, Coastal and Shelf Science, 76: 201-210. [ Links ]
Giménez, L., Dimitriadis, C., Carranza, A., Borthagaray, A. I. & Rodríguez, M. (2006). Unravelling the complex structure of a benthic community: A multiscale-multianalytical approach to an estuarine sandflat. Estuarine, Coastal and Shelf Science, 68: 462-472. [ Links ]
Giraldo, A., Rodríguez-Rubio, E. & Zapata, F. (2008). Condiciones oceanográficas en isla Gorgona, Pacífico oriental tropical de Colombia. Latin American Journal of Aquatic Research, 36: 121-128. [ Links ]
Giraldo, A. (2012). Geomorfología e hidroclimatología de Isla Gorgona. In A. Giraldo & B. Valencia (Eds.). Isla Gorgona: paraíso de biodiversidad y ciencia (pp. 17-26). Universidad del Valle. Cali, Colombia. [ Links ]
Guevara-Fletcher, C. E., Cantera, J. R., Mejía-Ladino, L. M. & Cortés, F. A. (2011). Benthic macrofauna associated with submerged bottoms of a tectonic estuary in Tropical Eastern Pacific. Journal of Marine Biology. doi:10.1155/2011/193759. [ Links ]
Guzmán-Alvis, A. & Díaz, J. M. (1993). Distribución espacial de la taxocenosis Annelida-Mollusca en la plataforma continental del Golfo de Salamanca, Caribe colombiano. Boletín de Investigaciones Marinas y Costeras, 22: 45-59. [ Links ]
Guzmán-Alvis, A., Solano, O. D., Córdoba-Tejada, M. E. & López-Rodríguez, A. C. (2001). Comunidad macroinfaunal de fondos someros tropicales en el Caribe colombiano. Boletín de Investigaciones Marinas y Costeras,. 30: 39-65. [ Links ]
Guzmán-Alvis, A. & Carrasco, F. (2005). Taxonomic aggregation and redundancy in a tropical macroinfaunal assemblage of the southern Caribbean in the detection of temporal patterns. Scientia Marina, 69: 133-141. [ Links ]
Guzmán-Alvis, A., Lattig, P. & Ruiz, J. A. (2006). Spatial and temporal characterization of soft bottom polychaetes in a shallow tropical bay (Colombian caribbean). Boletín de Investigaciones Marinas y Costeras, 35: 19-36. [ Links ]
Heard, R. W., Hansknecht, T. & Larsen, K. (2003). An illustrated identification guide to Florida Tanaidacea (Crustacea: Peracarida) occurring in depths of less than 200m. Florida Department of Environmental Protection. Annual Report, WM828. [ Links ]
Herrera, L. (2002). Temporal dynamic of the macroinvertebrates communities associated with the mangrove and sand ecosystems at Pianguita (Colombian pacific) from October 1999 to March 2001. Investigaciones Marinas, 30: 146-148. [ Links ]
Herrera, L. (2011). Aspectos de la biodiversidad y ecología de los anélidos poliquetos del Pacífico colombiano (Tesis doctoral). Universidad del Valle, Cali, Colombia. [ Links ]
Hines, A. H. & Comtois, K. L. (1985). Vertical distribution of infauna in sediments of a subestuary of central Chesapeake Bay. Estuaries, 8: 296-304. [ Links ]
Hyland, J., Cooksey, C., Balthis, L., Fulton, M., Bearden, D., McFall, G. & Kendall, M. (2006). The soft-bottom macrobenthos of Gray Reef National Marine Sanctuary and nearby shelf waters of the coast of Georgia, USA. Journal of Experimental Marine Biology and Ecology, 330: 307-326. [ Links ]
Keen A. M. (1971). Seashells of tropical west America: marine mollusks from Baja California to Perú. Stanford University Press. Stanford, USA. [ Links ]
Lazarus, J. F., Cobo, A. M., Herrera, L. & Satizabal, C. A. (2011). Crustacea. In J. R. Cantera (Ed.), Guía para la identificación de estadios de vida vulnerable de organismos marinos de Bahía Málaga (pp. 58-89). Editorial de la Universidad del Valle. Cali, Colombia. [ Links ]
León-Morales, R. & Vargas, J. A. (1998). Macroinfauna of a tropical fjord-like embayment, Golfo Dulce, Costa Rica. Revista de Biología Tropical, 46: 81-90. [ Links ]
Llinás, R., Pinto, J., Peña, F. & Caro, F. (1990). Geología. In J. Aguirre. & O. Rangel (Eds.). Biota y ecosistemas de Gorgona (pp. 55-65). Fondo FEN. Bogotá, Colombia. [ Links ]
Lohrer, A. M., Chiaroni, L. D., Hewitt, J. E. & Thrush, S. F. (2008). Biogenic disturbance determines invasion success in a subtidal soft-sediment system. Ecology, 89: 1299-1307. [ Links ]
López, G. R. & Levinton, J. S. (1987). Ecology of deposit-feeding animals in marine sediments. The Quarterly Review of Biology, 63: 235-260. [ Links ]
López, E., Cladera., P., San Martín, G., Laborda, A. & Aguado, M. T. (2002). Polychaetes assemblages inhabiting intertidal soft bottoms associated with mangroves systems in Coiba National Park (East Pacific, Panama). Wetlands Ecology and Management, 10: 233-242. [ Links ]
Lucero, C. H., Cantera, J. R. & Romero, I. C. (2006). Variability of macrobenthic assemblages under abnormal climatic conditions in a small scale tropical estuary. Estuarine, Coastal and Shelf Science, 68: 17-26. [ Links ]
Macdonald, T. A., Burd, B. J., Macdonald, V. I. & Van Roodselaar, A. (2010). Taxonomic and feeding guild classification for the marine benthic macroinvertebrates of the Strait of Georgia, British Columbia. Canadian Technical Reports of Fisheries and Aquatic Sciences, 2874: 63. [ Links ]
Mair, J. M., Cunningham, S. L., Sibaja-Corderom, J. A., Guzmán, H. M., Arroyo, M. F., Merino D. & Vargas, R. (2009). Mapping benthic faunal communities in the shallow and deep sediments of Las Perlas Archipelago, Pacific Panama. Marine Pollution Bulletin, 58: 385-373. [ Links ]
Mann, H. (2000). Ecology of coastal waters: with implications for management. Wiley-Blackwell. Oxford, England. [ Links ]
Maurer, D. & Vargas, J. A. (1984). Diversity of softbottom benthos in a tropical estuary: Gulf of Nicoya, Costa Rica. Marine Biology, 81: 97-106. [ Links ]
McBreen, F., Wilson, J. G., Mackie, A. S. Y. & Aonghusa, C. N. (2008). Seabed mapping in the southern Irish Sea: predicting benthic biological communities based on sediment characteristics. Hydrobiología, 606: 93-103. [ Links ]
Muñoz, P. & Salamanca, M. (2001). Flujo de plomo particulado a los sedimentos marinos y su incorporación en Paraprionospio pinnata (Polychaeta: Spionidae) en Bahía Concepción (36° Lat. S), Chile. Gayana (Concepción), 65: 155-166. [ Links ]
Rangel, O. & Rudas, A. (1990). Macroclima de Gorgona y de la región costera aledaña. In J. Aguirre & O. Rangel (Eds.). Biota y ecosistemas de Gorgona (p. 13-41). Fondo FEN. Bogotá, Colombia. [ Links ]
Reise, K. (2002). Sediment mediated species interactions in coastal waters. Journal of Sea Research, 48: 127-141. [ Links ]
Rosenberg, R., Nilsson, H. C. & Diaz, R. J. (2001). Response of Benthic Fauna and Changing Sediment Redox Profiles over a Hypoxic Gradient. Estuarine, Coastal and Shelf Science, 53: 343-350. [ Links ]
Rouse, G. W. & Pleijel, F. (2001). Polychaetes. Oxford University Press. Oxford, United Kingdom. [ Links ]
Schlacher, T.A., Newell, P., Clavier, J., Schlacher-Hoenlinger, M. A., Chevillon, C. & Britton, J. (1998). Soft-sediment benthic community structure in a coral reef lagoon-the prominence of spatial heterogeneity and ‘spot endemism’. Marine Ecology Progress Series, 174: 159-174. [ Links ]
Smith, C. R. & Kukert, H. (1996). Macrobenthic community structure, secondary production, and rates of bioturbation and sedimentation at the Kiine’ohe Bay Lagoon Floor. Pacific Science, 50: 211-229. [ Links ]
Solano, O. D., Cortés, F. A. & Ruíz, J. A. (2001). Ambientes y comunidades de fondos blandos. In L. M. Barrios. & M. López-Victoria (Eds.), Gorgona marina: Contribución al conocimiento de una isla única (pp. 65-78.). INVEMAR, Serie de Publicaciones especiales N° 7. Santa Marta, Colombia. [ Links ]
Turner, A. & Millward, G. E. (2002) Suspended Particles: their role in estuarine biogeochemical cycles. Estuarine, Coastal and Shelf Science, 55: 857-883. [ Links ]
Underwood, A. J. & Chapman, M. G. (2005). Design and analysis in benthic surveys. In A. Eleftheriou & A. McIntyre (Eds.), Methods for the study of marine benthos (pp. 1-42). Wiley-Blackwell. London, United Kingdom. [ Links ]
Wilson, W. H. (1991). Competition and predation in marine soft-sediment communities. Annual Review of Ecology, Evolution, and Systematics, 21: 221-241. [ Links ]
Woodin, S. A. (1976). Adult-larval interactions in dense infaunal assemblages: patterns of abundance. Journal of Marine Research, 34: 25-41. [ Links ]
Zajac, R. N. (2008). Challenges in marine, soft-sediment benthoscape ecology. Landscape Ecology, 23: 7-18. [ Links ]
Zapata, F. A. (2001). Formaciones coralinas de Isla Gorgona. In L. M. Barrios & M. López-Victoria (Eds.), Gorgona marina: Contribución al conocimiento de una isla única (pp. 27-40). INVEMAR, Serie de Publicaciones especiales N° 7. Santa Marta, Colombia. [ Links ]
Alongi, D. M. & Christoffersen, P. (1992). Benthic infauna and organism-sediment relations in a shallow, tropical coastal area: influence of outwelled mangrove detritus and physical disturbance. Marine Ecology Progress Series, 81: 229-245. [ Links ]
Bale, A. J. & Kenny, A. J. (2005). Sediment analysis and seabed characterization. In A. Eleftheriou & A. McIntyre (Eds.), Methods for the study of marine benthos (pp. 43-86). Wiley-Blackwell. London, United Kingdom. [ Links ]
Barnard, J. L. & Karaman, G. S. (1991a). The Families and genera of marine gammaridean Amphipoda (except marine gammaroids). Part 1. Records of the Australian Museum, 13: 1-417. [ Links ]
Barnard, J. L. & Karaman, G. S. (1991b). The Families and genera of marine gammaridean Amphipoda (except marine gammaroids). Part 2. Records of the Australian Museum, 13: 419-866. [ Links ]
Blanco, J. F. (2009). The hydroclimatology of Gorgona Island: seasonal and ENSO-related patterns. Actualidades Biológicas, 31: 111-121. [ Links ]
Bone, D. & Klein, E. (2000). Temporal variations in a tropical soft-bottom community, Venezuela. Journal of Coastal Research, 16: 278-286. [ Links ]
Cantera, J. R., Rubio, E. A., Borrero, F. J., Contreras, R., Zapata, F. & Buttkus, E. (1979). Taxonomía y distribución de los moluscos litorales de la Isla Gorgona, Colombia. In H. Von Prahl, M. Grögl & F. Guhl. (Eds.), Gorgona (p. 141-168). Universidad de Los Andes. Bogotá, Colombia. [ Links ]
Cantera, J. R. & Restrepo, J. D. (1995). (Eds.). Delta del río San Juan, Bahías de Málaga y Buenaventura, Pacífico Colombiano. Tomo II. Universidad EAFIT-Universidad del Valle. Medellín, Colombia. [ Links ]
Clarke, K. R & Warwick, R. M. (2001). Change in marine communities: an approach to statistical analysis and interpretation. Primer-E. Plymouth, United Kingdom. [ Links ]
Clarke, K. R., Somerfield, P. J. & Gorley, R. N. (2008). Testing of null hypotheses in exploratory community analyses: similarity profiles and biota-environment linkage. Journal of Experimental Marine Biology and Ecology, 366: 56-69. [ Links ]
Climent, M. T. & García, B. (2012). Cálculos estequiométricos: reacción entre carbonato de calcio y ácido clorhídrico. Retrieved from http://roderic.uv.es/handle/10550/25937. [ Links ]
Colwell, R. K. & Coddington, J. A. (1994). Estimating terrestrial biodiversity through extrapolation. Transactions of the Royal Society Biological Sciences, 345: 101-118. [ Links ]
Colwell, R. K. (2013). EstimateS: Statistical estimation of species richness and shared species from samples. Version 9.1.0. User’s Guide and application. Retrieved from http://purl.oclc.org/estimates. [ Links ]
Cortés, F. A. (2010). Estratificación vertical y distribución horizontal de la comunidad macroinfaunal sublitoral en los sedimentos marinos (Tesis de Maestría). Universidad Nacional de Colombia, Colombia. [ Links ]
Cortés, F. A., Solano, O. D. & Ruíz-López, J. A. (2012). Variación espacio-temporal de la fauna macrobentónica asociada a fondos blandos y su relación con factores ambientales en el Parque Nacional Natural Gorgona, Pacífico colombiano. Boletín de Investigaciones Marinas y Costeras, 41: 323-353. [ Links ]
Delgado-Blas, V. H. (2001). Distribución espacial y temporal de poliquetos (Polychaeta) bénticos de la plataforma continental de Tamaulipas, Golfo de México. Revista de Biología Tropical, 49: 141-147. [ Links ]
Díaz, J. M., Pinzón, J. H., Perdomo, A. M, Barrios, L. M. & López-Victoria, M. (2001). Generalidades. In L. M. Barrios & M. L. Victoria (Eds.). Gorgona marina: Contribución al conocimiento de una isla única (pp. 17-26.). INVEMAR, Serie de Publicaciones especiales N° 7. Santa Marta, Colombia. [ Links ]
Eleftheriou, A. & Moore, D. C. (2005). Macrofauna techniques. In A. Eleftheriou & A. McIntyre (Eds.), Methods for the study of marine benthos (pp. 160-228). Wiley-Blackwell. London, United Kingdom. [ Links ]
Elías, R., Vallarinot., E. A., Scagliolat, M. & Isla, F. I. (2004). Macrobenthic distribution patterns at a sewage disposal site in the inner shelf off Mar del Plata (SW Atlantic). Journal of Coastal Research, 20: 1176-1182. [ Links ]
Fauchald, K. (1977). Polychaetes from intertidal areas in Panama, with a review of previous shallow-water records. Smithson. Contributions to Zoology, 221: 1-81. [ Links ]
Giere, O. (2009). Meiobenthology: the microscopic motile fauna of aquatic sediments. Springer. USA. [ Links ]
Gillet, P., Mouloud, M., Durou, C. & Deutsch, B. (2008). Response of Nereis diversicolor population (Polychaeta, Nereididae) to the pollution impact Authie and Seine estuaries (France). Estuarine, Coastal and Shelf Science, 76: 201-210. [ Links ]
Giménez, L., Dimitriadis, C., Carranza, A., Borthagaray, A. I. & Rodríguez, M. (2006). Unravelling the complex structure of a benthic community: A multiscale-multianalytical approach to an estuarine sandflat. Estuarine, Coastal and Shelf Science, 68: 462-472. [ Links ]
Giraldo, A., Rodríguez-Rubio, E. & Zapata, F. (2008). Condiciones oceanográficas en isla Gorgona, Pacífico oriental tropical de Colombia. Latin American Journal of Aquatic Research, 36: 121-128. [ Links ]
Giraldo, A. (2012). Geomorfología e hidroclimatología de Isla Gorgona. In A. Giraldo & B. Valencia (Eds.). Isla Gorgona: paraíso de biodiversidad y ciencia (pp. 17-26). Universidad del Valle. Cali, Colombia. [ Links ]
Guevara-Fletcher, C. E., Cantera, J. R., Mejía-Ladino, L. M. & Cortés, F. A. (2011). Benthic macrofauna associated with submerged bottoms of a tectonic estuary in Tropical Eastern Pacific. Journal of Marine Biology. doi:10.1155/2011/193759. [ Links ]
Guzmán-Alvis, A. & Díaz, J. M. (1993). Distribución espacial de la taxocenosis Annelida-Mollusca en la plataforma continental del Golfo de Salamanca, Caribe colombiano. Boletín de Investigaciones Marinas y Costeras, 22: 45-59. [ Links ]
Guzmán-Alvis, A., Solano, O. D., Córdoba-Tejada, M. E. & López-Rodríguez, A. C. (2001). Comunidad macroinfaunal de fondos someros tropicales en el Caribe colombiano. Boletín de Investigaciones Marinas y Costeras,. 30: 39-65. [ Links ]
Guzmán-Alvis, A. & Carrasco, F. (2005). Taxonomic aggregation and redundancy in a tropical macroinfaunal assemblage of the southern Caribbean in the detection of temporal patterns. Scientia Marina, 69: 133-141. [ Links ]
Guzmán-Alvis, A., Lattig, P. & Ruiz, J. A. (2006). Spatial and temporal characterization of soft bottom polychaetes in a shallow tropical bay (Colombian caribbean). Boletín de Investigaciones Marinas y Costeras, 35: 19-36. [ Links ]
Heard, R. W., Hansknecht, T. & Larsen, K. (2003). An illustrated identification guide to Florida Tanaidacea (Crustacea: Peracarida) occurring in depths of less than 200m. Florida Department of Environmental Protection. Annual Report, WM828. [ Links ]
Herrera, L. (2002). Temporal dynamic of the macroinvertebrates communities associated with the mangrove and sand ecosystems at Pianguita (Colombian pacific) from October 1999 to March 2001. Investigaciones Marinas, 30: 146-148. [ Links ]
Herrera, L. (2011). Aspectos de la biodiversidad y ecología de los anélidos poliquetos del Pacífico colombiano (Tesis doctoral). Universidad del Valle, Cali, Colombia. [ Links ]
Hines, A. H. & Comtois, K. L. (1985). Vertical distribution of infauna in sediments of a subestuary of central Chesapeake Bay. Estuaries, 8: 296-304. [ Links ]
Hyland, J., Cooksey, C., Balthis, L., Fulton, M., Bearden, D., McFall, G. & Kendall, M. (2006). The soft-bottom macrobenthos of Gray Reef National Marine Sanctuary and nearby shelf waters of the coast of Georgia, USA. Journal of Experimental Marine Biology and Ecology, 330: 307-326. [ Links ]
Keen A. M. (1971). Seashells of tropical west America: marine mollusks from Baja California to Perú. Stanford University Press. Stanford, USA. [ Links ]
Lazarus, J. F., Cobo, A. M., Herrera, L. & Satizabal, C. A. (2011). Crustacea. In J. R. Cantera (Ed.), Guía para la identificación de estadios de vida vulnerable de organismos marinos de Bahía Málaga (pp. 58-89). Editorial de la Universidad del Valle. Cali, Colombia. [ Links ]
León-Morales, R. & Vargas, J. A. (1998). Macroinfauna of a tropical fjord-like embayment, Golfo Dulce, Costa Rica. Revista de Biología Tropical, 46: 81-90. [ Links ]
Llinás, R., Pinto, J., Peña, F. & Caro, F. (1990). Geología. In J. Aguirre. & O. Rangel (Eds.). Biota y ecosistemas de Gorgona (pp. 55-65). Fondo FEN. Bogotá, Colombia. [ Links ]
Lohrer, A. M., Chiaroni, L. D., Hewitt, J. E. & Thrush, S. F. (2008). Biogenic disturbance determines invasion success in a subtidal soft-sediment system. Ecology, 89: 1299-1307. [ Links ]
López, G. R. & Levinton, J. S. (1987). Ecology of deposit-feeding animals in marine sediments. The Quarterly Review of Biology, 63: 235-260. [ Links ]
López, E., Cladera., P., San Martín, G., Laborda, A. & Aguado, M. T. (2002). Polychaetes assemblages inhabiting intertidal soft bottoms associated with mangroves systems in Coiba National Park (East Pacific, Panama). Wetlands Ecology and Management, 10: 233-242. [ Links ]
Lucero, C. H., Cantera, J. R. & Romero, I. C. (2006). Variability of macrobenthic assemblages under abnormal climatic conditions in a small scale tropical estuary. Estuarine, Coastal and Shelf Science, 68: 17-26. [ Links ]
Macdonald, T. A., Burd, B. J., Macdonald, V. I. & Van Roodselaar, A. (2010). Taxonomic and feeding guild classification for the marine benthic macroinvertebrates of the Strait of Georgia, British Columbia. Canadian Technical Reports of Fisheries and Aquatic Sciences, 2874: 63. [ Links ]
Mair, J. M., Cunningham, S. L., Sibaja-Corderom, J. A., Guzmán, H. M., Arroyo, M. F., Merino D. & Vargas, R. (2009). Mapping benthic faunal communities in the shallow and deep sediments of Las Perlas Archipelago, Pacific Panama. Marine Pollution Bulletin, 58: 385-373. [ Links ]
Mann, H. (2000). Ecology of coastal waters: with implications for management. Wiley-Blackwell. Oxford, England. [ Links ]
Maurer, D. & Vargas, J. A. (1984). Diversity of softbottom benthos in a tropical estuary: Gulf of Nicoya, Costa Rica. Marine Biology, 81: 97-106. [ Links ]
McBreen, F., Wilson, J. G., Mackie, A. S. Y. & Aonghusa, C. N. (2008). Seabed mapping in the southern Irish Sea: predicting benthic biological communities based on sediment characteristics. Hydrobiología, 606: 93-103. [ Links ]
Muñoz, P. & Salamanca, M. (2001). Flujo de plomo particulado a los sedimentos marinos y su incorporación en Paraprionospio pinnata (Polychaeta: Spionidae) en Bahía Concepción (36° Lat. S), Chile. Gayana (Concepción), 65: 155-166. [ Links ]
Rangel, O. & Rudas, A. (1990). Macroclima de Gorgona y de la región costera aledaña. In J. Aguirre & O. Rangel (Eds.). Biota y ecosistemas de Gorgona (p. 13-41). Fondo FEN. Bogotá, Colombia. [ Links ]
Reise, K. (2002). Sediment mediated species interactions in coastal waters. Journal of Sea Research, 48: 127-141. [ Links ]
Rosenberg, R., Nilsson, H. C. & Diaz, R. J. (2001). Response of Benthic Fauna and Changing Sediment Redox Profiles over a Hypoxic Gradient. Estuarine, Coastal and Shelf Science, 53: 343-350. [ Links ]
Rouse, G. W. & Pleijel, F. (2001). Polychaetes. Oxford University Press. Oxford, United Kingdom. [ Links ]
Schlacher, T.A., Newell, P., Clavier, J., Schlacher-Hoenlinger, M. A., Chevillon, C. & Britton, J. (1998). Soft-sediment benthic community structure in a coral reef lagoon-the prominence of spatial heterogeneity and ‘spot endemism’. Marine Ecology Progress Series, 174: 159-174. [ Links ]
Smith, C. R. & Kukert, H. (1996). Macrobenthic community structure, secondary production, and rates of bioturbation and sedimentation at the Kiine’ohe Bay Lagoon Floor. Pacific Science, 50: 211-229. [ Links ]
Solano, O. D., Cortés, F. A. & Ruíz, J. A. (2001). Ambientes y comunidades de fondos blandos. In L. M. Barrios. & M. López-Victoria (Eds.), Gorgona marina: Contribución al conocimiento de una isla única (pp. 65-78.). INVEMAR, Serie de Publicaciones especiales N° 7. Santa Marta, Colombia. [ Links ]
Turner, A. & Millward, G. E. (2002) Suspended Particles: their role in estuarine biogeochemical cycles. Estuarine, Coastal and Shelf Science, 55: 857-883. [ Links ]
Underwood, A. J. & Chapman, M. G. (2005). Design and analysis in benthic surveys. In A. Eleftheriou & A. McIntyre (Eds.), Methods for the study of marine benthos (pp. 1-42). Wiley-Blackwell. London, United Kingdom. [ Links ]
Wilson, W. H. (1991). Competition and predation in marine soft-sediment communities. Annual Review of Ecology, Evolution, and Systematics, 21: 221-241. [ Links ]
Woodin, S. A. (1976). Adult-larval interactions in dense infaunal assemblages: patterns of abundance. Journal of Marine Research, 34: 25-41. [ Links ]
Zajac, R. N. (2008). Challenges in marine, soft-sediment benthoscape ecology. Landscape Ecology, 23: 7-18. [ Links ]
Zapata, F. A. (2001). Formaciones coralinas de Isla Gorgona. In L. M. Barrios & M. López-Victoria (Eds.), Gorgona marina: Contribución al conocimiento de una isla única (pp. 27-40). INVEMAR, Serie de Publicaciones especiales N° 7. Santa Marta, Colombia. [ Links ]
1. Departamento de Biología, Universidad del Valle, A.A. 25360 Cali, Colombia; bellival@univalle.edu.co
2. Ecomanglares, Grupo de Investigación en Ecología de Estuarios y Manglares, Departamento de Biología, Universidad del Valle, A.A. 25360 Cali, Colombia; leoheroz@yahoo.com
3. Departamento de Biología, Universidad del Valle, A.A. 25360 Cali, Colombia; alan.giraldo@correounivalle.edu.co
Recibido 18-X-2013. Corregido 20-XI-2013. Aceptado 19-XII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}