










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.1 San José Feb. 2014
Patterns at Multi-Spatial Scales on Tropical Island Stream Insect Assemblages (Gorgona Island Natural National Park, Colombia, Tropical Eastern Pacific)
Abstract
Tropical Eastern Pacific island streams (TEPis) differ from other neotropical streams in their rainy climate, mixed sedimentary-volcanic geology and faunal composition. Yet, their relationships between environmental characteristics and stream biota remain unexplored. We analyzed the environmental subject at three spatial scales using a fully nested sampling design (6 streams, 2 reaches within each stream, 2 habitats within each reach, and 4 replicates per habitat) on Gorgona Island (Colombia). Sampling was carried out in two months with contrasting rainfall during early 2009. We studied the spatial variation of assemblage composition and density along with 27 independent variables within two contrasting rainfall conditions. Five stream-scale variables, two reach-scale variables, and five habitat-scale variables were selected using a Canonical Correspondence Analysis (CCA). A partial CCA showed that the total variance explained was 13.98%, while stream- and habitat-scale variables explained the highest proportion of the variance (5.74 and 5.01%, respectively). Dissolved oxygen (as affected by rainfall), high-density use zone (a management category), and sedimentary geology were the best descriptors of insect assemblages. The two latter descriptors affected fine-scale variables such as total benthic organic matter and gravel substratum, respectively. A Nested ANOVA showed significant differences in total density and richness among streams and habitats, and significant differences between the two sampling months regardless of the spatial scale. The evenness showed a significant stream- and habitat-dependent temporal variability. These results suggested that rainfall regime in Gorgona Island might be a driver of insect assemblage dynamics mediated by water chemistry and substratum properties. Spatial assemblage variability here is greater within habitats (among samples), and a minor fraction occurs at habitat- and stream-scales, while no longitudinal pattern was observed probably due to the short courses. Temporal variability should be further studied relative to rainfall and discharge regimes. Rev. Biol. Trop. 62 (Suppl. 1): 65-83. Epub 2014 February 01.
Key words: protected areas, benthos, biodiversity, stream, Neotropics.
Resumen
Las quebradas insulares del Pacífico Tropical Oriental difieren de otras quebradas neotropicales en términos de regímenes de lluvia, geología sedimentaria-volcánica, y composición faunística; pero la relación entre características ambientales y la biota de estas quebradas insulares permanece inexplorada. En este estudio, conducido en Isla Gorgona, se analizó el efecto restrictivo de 27 variables ambientales sobre la distribución, composición y densidad de los insectos acuáticos en tres escalas espaciales; se consideraron además diferencias en las intensidades de la lluvia entre los muestreos. Los muestreos se realizaron en 2009, en dos meses con intensidades de lluvia contrastantes; se siguió un diseño muestral anidado (6 quebradas, 2 tramos en cada quebrada, 2 hábitats en cada tramo, y 4 réplicas por hábitat). En un ACC fueron seleccionadas 5 variables operando a la escala-quebrada, 2 a la escala-tamo y 5 a la escala-hábitat. Un ACC parcial mostró que la varianza total explicada fue de 13.98%; las variables de las escalas hábitat y quebrada explicaron la mayor proporción (5.74 y 5.01%, respectivamente). Los mejores descriptores del “ensamblaje” fueron, oxígeno disuelto (influenciado por la lluvia), zona de alta densidad de uso y geología sedimentaria. Las dos últimas afectaron variables de escalas menores tales como, materia orgánica béntica y sustrato de gravas. Un ANOVA anidado presentó diferencias significativas de densidad total y de riqueza entre quebradas y hábitats, así como entre meses. La equidad mostró una variabilidad temporal significante dependiente de la quebrada y del hábitat. Se sugiere que el régimen de lluvia podría ser el conductor principal de las dinámicas, pues influye en la química del agua y en las propiedades de los sustratos. La variabilidad espacial de la composición y la estructura es mayor dentro de los hábitats (entre las muestras), y en menor proporción en las escalas quebrada y hábitat. No se definió un patrón longitudinal debido probablemente a la corta longitud de las quebradas. La variabilidad temporal debe ser estudiada a fondo en función de los regímenes de lluvia y de descarga.
Palabras clave: áreas protegidas, bentos, biodiversidad, corriente, Neotrópico.
A major goal of stream ecology is to identify and to explain non-random patterns of species composition in natural communities. This topic has been widely studied in continental landscapes but scarcely in islands, particularly within the humid tropics. Aquatic insects have proven to be useful for studying spatial patterns of community organization in continental streams. Spatial patterns have been explored using the hierarchical model of fluvial landscape (Frissell, Liss, Warren & Hurley, 1986), consisting of a serial nestedness from the watershed to the microhabitat. The regional scale of observation has been incorporated by recent studies (e.g. Corkum, 1991; Sandin & Johnson, 2004). By employing this hierarchical model, researchers have identified the important abiotic variables, ecological mechanisms, and ecosystem processes that correlate with the distribution of taxa, and with the structure and composition of insect assemblages at specific scales of observation.
The controlling variables operating at particular spatial and temporal scales have been called landscape filters (sensu Poff, 1997), similarly to a series of hypothetical sieves constraining the colonization of local sites by the regional pool of species. These filters are defined as habitat features or ecological and environmental attributes that influence the probability of occurrence of individual species as members of a local assemblage. Since both the “Hierarchical Fluvial Landscape” model and the “Landscape Filter” concept were proposed from temperate zone settings, most of the empirical field tests have been conducted in high latitudes. Consequently, as with many other hypotheses regarding the structure and function of stream communities (Jackson & Sweeney, 1995), the applicability of these models to tropical streams remain untested, and therefore, many filters remain unnoticed in tropical streams. Recently, Boulton et al. (2008) summarized the general differences between temperate and tropical streams, highlighting that temperature and precipitation regimes are probably the most distinctive features. Apart from these two differences, since geology, climate, and biomes are as diverse within the tropics as within the temperate zone, most hypotheses in stream ecology could be straight-forward tested in tropical settings.
In contrast to the temperate-tropical comparison, greater dissimilarities are observed between continental and island streams. In the specific case of tropical islands, due to the filter effect of the oceanic separation, there is a biogeographic uniqueness in many groups enhanced by the geological complexity not found on their continental counterparts (Mueller-Dombois, 2002). For instance, Smith, Covich & Brasher (2003) indicated that streams on tropical islands exhibit relatively short, straight, and steep channels, and their watersheds are smaller and narrower compared to continental ones, and Covich, Crowl & Scatena (2003) suggested that island streams tend to be more driven by discrete meteorological events (e.g. cyclones) rather than by mean conditions due to their maritime climates. Since meso-scale climatic phenomena such as tropical storms and hurricanes are relatively unpredictable, studying stream insect assemblages in islands is useful to understanding how biodiversity is a reflection of habitat properties, and how adapted and resilient are the different taxonomic groups to natural disturbance regimes (Smith et al., 2003).
There are a few studies regarding filters restricting the richness and abundance of aquatic insects on tropical islands. However, it has been found that island origin, isolation, size, and age, as well as ocean currents and climate change, are important influences at a regional scale (e.g. March, Benstead, Pringle & Luckymis, 2003; Craig, 2003; Benbow, McIntosh, Burky & Way, 2005; Covich, 2006; Benstead, March, Pringle, Ewel & Short, 2009). At the basin scale, stream order and invasion by introduced species are major filters (Englund, 1999). At stream and reach scales, geology, land use, rainfall, erosion, dams, roads, predation, and competition are the major filters (Polhemus, 1995; March, Pringle, Townsend & Wilson, 2002; Craig, 2003; Greathouse & Pringle, 2005). Streambed roughness and water velocity are the main filters at the microhabitat scale (e.g. Hendrich & Yang, 1999). Nonetheless, this information has been obtained from multiple studies, each conducted at a single spatial scale. The only study employing observations at multiple spatial scales on insect diversity in tropical islands was carried out in an Eastern Pacific island (i.e.Coiba) (Boyero & Bailey, 2001, Boyero & Bosch, 2004). Although community composition and structure were greatly variable at the sampling-plot scale, biotic and abiotic filters were not measured in this study. Therefore, island streams within the Tropical Eastern Pacific remain largely unexplored (Coiba in Panama: Boyero & Bailey, 2001; Boyero & Bosch, 2004; Gorgona in Colombia: Gómez-Aguirre, Longo-Sánchez & Blanco, 2009) but they may exhibit climatic and geologic differences relative to areas more intensively studied in the Caribbean, thus providing new templets for testing ecological hypotheses.
We conducted this study in Gorgona Island, a natural national park located off the Pacific coast of Colombia, using a fully nested sampling design consisting of replicates (sampling quadrats), habitats (pools and riffles), and reaches (coastal plain and montane) within six streams. In this study we asked: (1) ¿Are the variables determining insect assemblage density linked in a hierarchical fashion? We propose that environmental filters in tropical islands follow a top-down pathway, from the island (regional scale) to the microhabitat scale (local scale). Therefore, we predict that the describing variables for the stream scale are geology, riparian land-use, and water chemistry; in the reach scale, these variables affect channel slope, streamflow, and riparian cover that, in turn, affect habitat-scale variables such as substrate types, depth, water temperature, and concentrations of organic and inorganic matter. (2) ¿How much of the variance in insect assemblage density is explained by variables operating in each of the three spatial scales studied (stream, reach, and habitat)? If driving processes operate in a top-down pathway, we consider that the variance explained would decrease sequentially from coarse to fine scales. Finally, we preliminary propose that the above patters are largely affected by changes in rainfall, acting at a regional scale (i.e. island), and therefore separated the analyses from two surveys conducted under contrasting rainfall conditions. To test these hypotheses, the objectives of this work were: (a) to assess the environmental variables operating at each spatial scale (stream, reach, habitat) that might constrain the assemblage composition and distribution; (b) to partition the total density variation into the influencing environmental variables; (c) to examine the spatial (slope, stream, reach, habitat) and temporal variation of assemblage structure.
Materials and methods
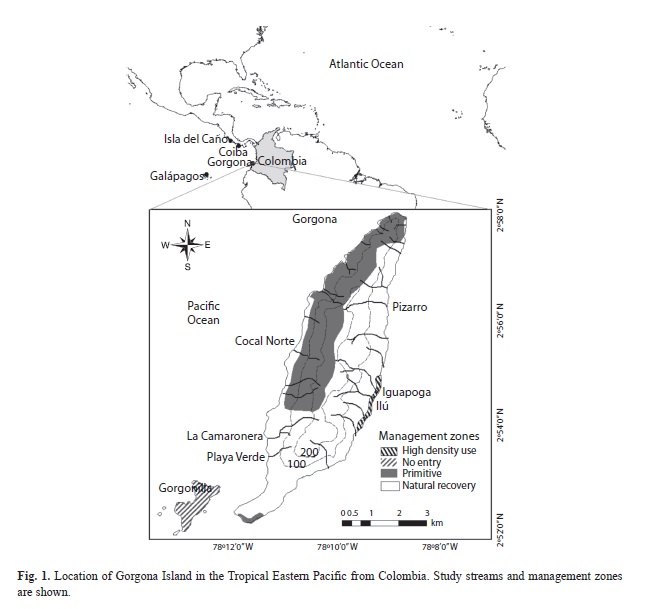
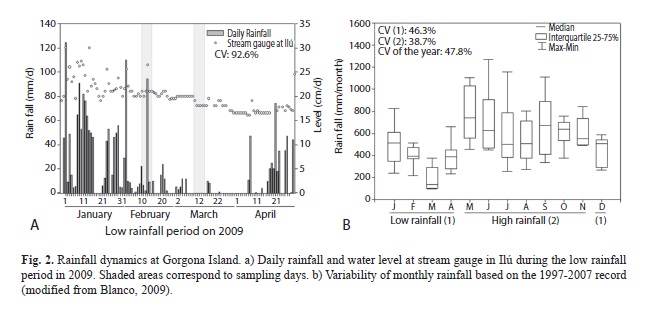
Study area. Gorgona is a small island (16km2) of volcanic origin, located at the edge of the continental shelf at 54km off the Pacific coast of Colombia (Guapi municipality; 2°47’-3°06’ N, 78°06’-78°18’ W; Fig. 1). Gorgona was a penitentiary island between 1960 and 1982, but it was declared as a National Natural Park in 1984. The relief is mountainous with a narrow width (< 3km) and a maximum elevation of 338 masl. The climate is superhumid tropical maritime due to the high air temperature and humidity (annual means 26°C and 90%; Rangel, 1995). This is controlled principally by the Inter Tropical Convergence Zone (ITCZ) and El Niño Southern Oscillation (ENSO). The annual rainfall regime is monomodal (max=1500mm/month; min=100), with peaks between May and November. A less rainy season occurs between December and April (Fig. 2b; Blanco, 2009). Mean total annual rainfall is significantly higher and more variable within and among years at Gorgona (6 661mm/year) than at the continent nearby (4 897mm/year; Blanco, 2009).
Sampling design. Sampling was conducted at 2 reaches along 6 streams (Ilú, Iguapoga, Pizarro, Playa Verde, La Camaronera and Cocal Norte) in February and March 2009. We conducted an explicit temporal analysis, considering two set of samples as independent statistical blocks because their differences in rainfall could have significant implications on insect assemblages (as evidenced by Gómez-Aguirre et al., 2009). Total rainfall was 377.7mm in February and 55.7mm in March, and the coefficient of variation for the entire low rainfall period (January-April) in 2009 was 92.6% (Fig. 2a), a significant value compared to the 47.8% of intra-annual variation (Fig. 2b).
Stream reaches were located in three different management zones: 1) primary native forest, 2) high-density use (the area where administrative and lodging buildings are located), and 3) natural recovery (the area where the prison was located and abandoned facilities remain) (Table 2, Fig. 1), established by the authority. Ilú, Iguapoga, and Pizarro, located in the Eastern slope, are disturbed streams by the presence of small dams for hydropower generation, and plantations of fruit trees. Playa Verde, La Camaronera, and Cocal Norte, located in the Western slope, do not show signs of human disturbances, and they were seemly unaffected during the penitentiary operation due to isolation.
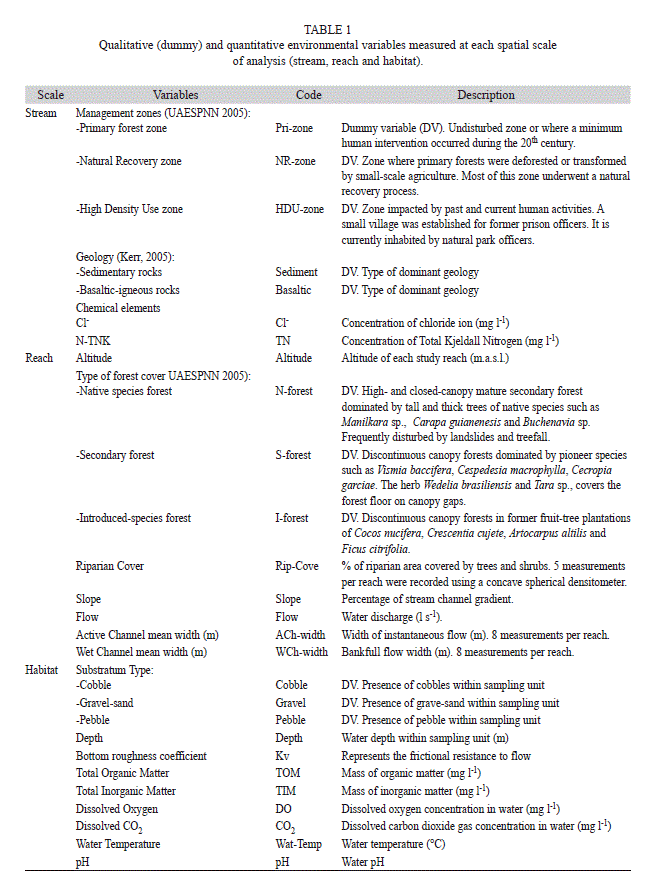
Five nested spatial scales were included in the study: island slope, stream (nested within slope), reach (nested within stream), habitat (nested within reach), and sample (sampling quadrat nested within habitat). In each slope (Western and Eastern) three streams were selected according to their accessibility. The two reaches were located in mid-elevations (montane segments) and lowlands (range: 70-10masl, maximum elevation 200masl), each of 50m-length. Two habitats were selected, a riffle and a pool (with two replicates for each one), and two Surber samples –with a 0.09m2 frame and 0.5mm mesh– were taken within each habitat replicate. The organisms were separated from substrates in the field and preserved in vials with 90% ethanol and then transported to the laboratory. Using standard methods and instruments, twenty seven environmental variables were measured at stream, reach, and habitat scales and, therefore, all analyses focused on these scales. Qualitative and quantitative environmental variables were measured in the field or obtained from the literature for each spatial scale. Classes of the qualitative environmental variables were transformed into dummy binary variables (Table 1).
Insect identifications. Insects were identified to genus or morphotype (or subfamily level for Chironomidae, Coleoptera, and Lepidoptera) using the following literature: Roldán (1988), Wiggins (1996), Aristizábal-García (2002), Pérez-Gutiérrez (2003), and Domínguez & Fernández (2009). Many taxa were confirmed by experts, and reference collections were archived at the Limnology laboratory at the Universidad de Antioquia (Colombia).
Statistical analysis. Taxa were classified as dominant, abundant, frequent, and rare. Natural-logarithm (ln) transformed abundance of taxa was plotted against frequency and the resulting plot was divided into four quadrants according to the intersection of the average ln-abundance and 50% of frequency (Magurran, 2004). The organisms in the upper left quadrant were considered dominant, those of the lower quadrant in the same side as rare; and those in the upper and lower right quadrants as frequent and occasional, respectively (not shown). To detect the existence of significant autocorrelation among the 27 environmental variables considered (Tables 1, 2), we conducted a Spearman correlation analysis. Active width, wet width, and dissolved carbon dioxide, all significantly correlated with streamflow (r≥0.60, α<0.05), were removed from further analyses.
Using a Detrended Correspondence Analysis (DCA) (Hill & Gauch, 1980) on insect density through the sampling units and periods, it was established that a direct gradient analysis (Canonical Correspondence Analysis, CCA) would be the most robust ordination method between the environmental variables and assemblage density. Additionally, a partial CCA (pCCA) (Borcard, Legendre & Drapeau, 1992) was carried out to evaluate the relationships among the explanatory variables at the three spatial scales (stream, reach, and habitat) and the assemblage density. To decide whether to use a linear (RDA) or a unimodal (CCA) ordination method, the gradient length was measured in DCA using a detrending by segments and non-linear rescaling of axes. DCA of insect density had a gradient length of 3.37 standard deviation units in the first axis, indicating that a unimodal model would best fit to the data (Lepŝ & Ŝmilauer, 2003).
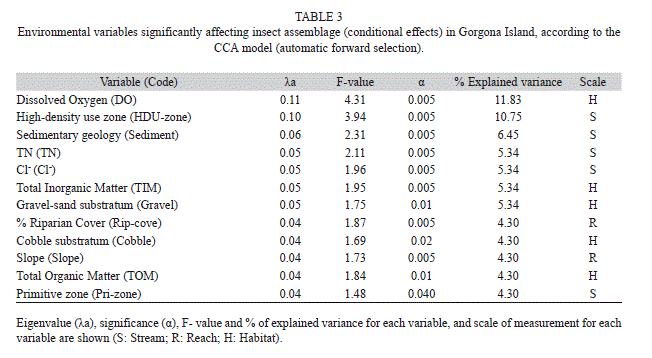
In CCA, the environmental dataset was centered and standardized prior to analysis, except for the categorical variables. The species density data were log10 (x+1) transformed, while rare species were down-weighted. Environmental variables used in CCA and pCCA were automatically forward-selected and tested with Monte Carlo permutation test with 199 unrestricted permutations. Five stream-scale variables, two reach-scale variables, and five habitat-scales variables were selected from the resultant suite of 24 variables (Table 3). These variables were significant (α-value<0.05) with inflation factors <20. Only taxa with the highest weight (the most frequent ones) were named in the CCA plot.
Using pCCA, we decomposed total variation of the biological response variables into: (a) the non-shared variation for a specific set of explanatory variables, (b) shared variation by all the measured variables, and (c) random error. To reveal the total variance explained, we used a CCA that included all variables sampled in stream, reach, and habitat scales (S+R+H), and no covariates were used. The variance unexplained was determined by the difference between the total variance (100%) and the variance explained by these levels [i.e. 100-(S+R+H)]. Several pCCA were calculated to establish the variance explained by each scale alone (S, R, and H), using the remaining scales as covariates (e.g. S-R-H, with R and H as covariate). Finally, several pCCA were carried out to calculate the shared effects by selected scales (e.g. S+R-H, with H as covariate). All ordination analyses were performed using CANOCO for Windows v. 4.5 (Ter Braak & Ŝmilauer, 2002). All taxa were used in the statistical analyses, because the omission of rare taxa may reduce the ability to describe important assemblage gradients (Cao, Williams & Williams, 1998).
To test for the nestedness of variance components on the insect assemblage density, richness, and Pielou’s evenness index (each as independent response variables), we used a nested ANOVA model. In addition, to control for any effect of time (i.e. rainfall) (fixed factor named here “month”), it was crossed with each one of the spatial scales (e.g. month*stream). In the analysis, “sample” was the error term, thus the significance of variation could not be calculated at the microhabitat scale. Additionally, we obtained the percentage of variance explained at each spatial scale for each of the estimates. This analysis was performed in MINITAB v. 14.
Results
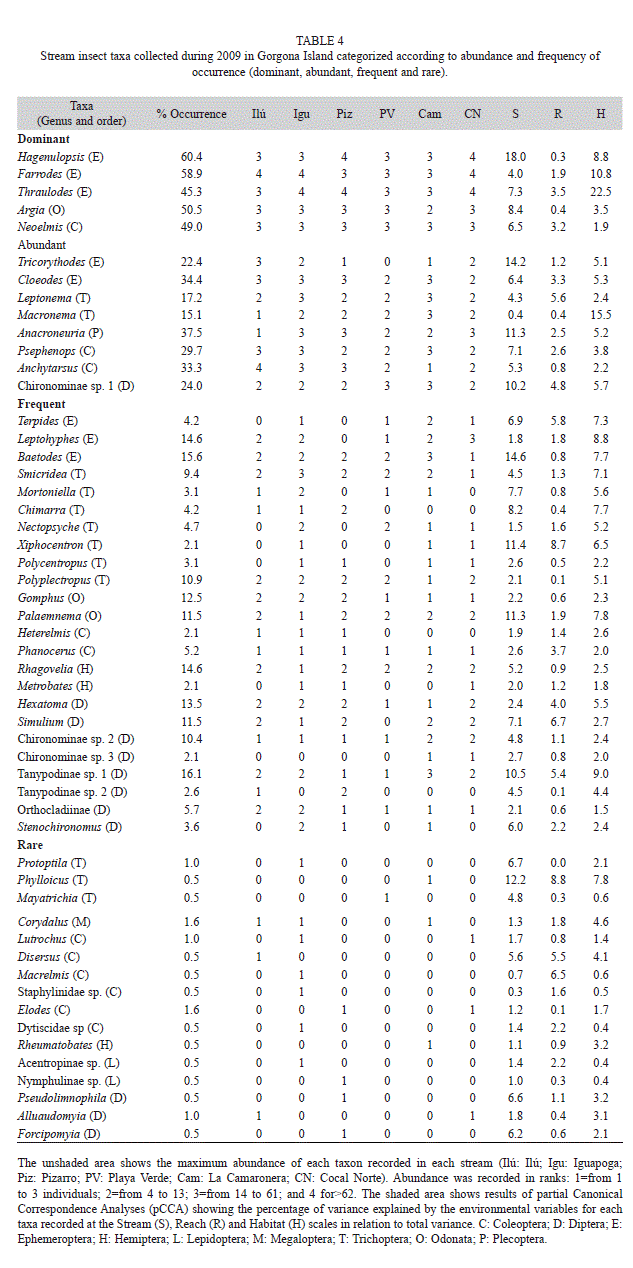
Insect assemblage. 1 168 individuals were identified in February and 1 877 in March, corresponding to 48 and 45 genera, respectively. During both periods, the orders with the highest taxon richness were Diptera (12), Trichoptera (11), Coleoptera (11), and Ephemeroptera (8), and the less rich orders were Hemiptera (3), Lepidoptera (2), Plecoptera (1), and Megaloptera (1) (Table 4). Iguapoga stream had the highest taxa richness (42) and Playa Verde stream had the lowest (28). The rest of the streams had intermediate values of richness (range: 33-36). The highest densities occurred in Cocal Norte (144.1ind. m-2), Iguapoga (126.6), and Pizarro (117.6), while the lowest occurred in Playa Verde (56.2). Intermediate values were found in Ilú and La Camaronera (102.3 and 98.8ind. m-2, respectively).
Out of 53 genera, there were 5 dominant genera, mainly represented by Ephemeroptera. There were 8 abundant taxa including Ephemeroptera, Trichoptera, Coleoptera, and Plecoptera, 24 frequent taxa (mostly Trichoptera and Diptera), and 16 rare taxa (mostly Coleoptera) (Table 4).
Multivariate relationships. In the CCA, the total variance (inertia) for the insect density matrix was 4.97. Out of this variance, 0.93 (18.61%) was explained by all 24 environmental variables. Twelve out of 24 variables explained 0.67 (13.48%) of total variance. The proportion of variance explained by each environmental variable was calculated by dividing the eigenvalue (λa) of each one (Table 3) by 0.93. Thus, the best predicting variables of the variation in the insect dataset were located at stream and habitat scales (32.18 and 31.11% in total, respectively). These variables were DO, high-density use zone, sedimentary geology, and total nitrogen (TN) and chloride ion (Cl-) concentrations. Reach-scale variables only explained 8.6% of total variance (Table 3).
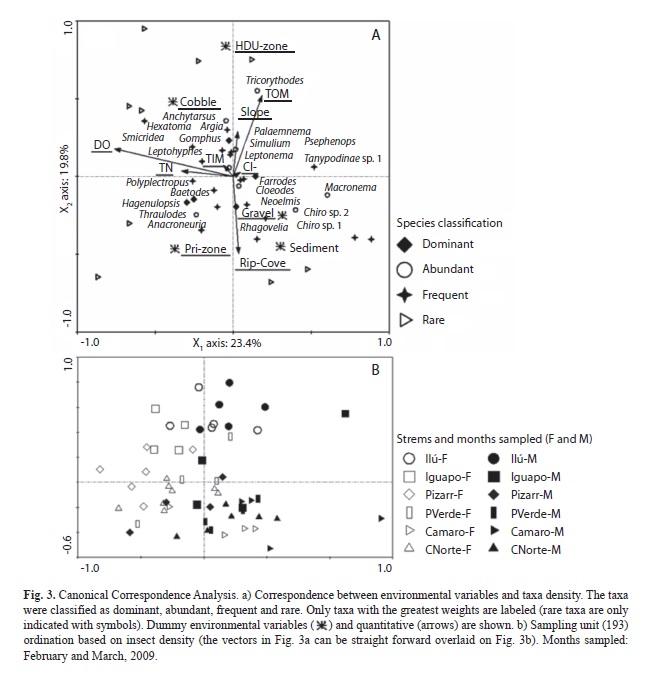
In the CCA, dissolved oxygen was the main environmental variable correlated with density along X1 axis. The X2 axis was tightly correlated with management zone categories and riparian cover. Streams in the High Density Use (HDU)-zone (Ilú and Iguapoga –disturbed streams–) were located on the positive side of X2 (Fig. 3b). Riparian cover was best correlated with streams at the negative side of the X2, regardless of stream location within either Primary (Pri) or Natural Recovery (NR) zones. The only stream in Pri-zone (Cocal Norte –undisturbed) was located on this end. Samples from streams in NR-zone (Playa Verde, La Camaronera –both undisturbed– and Pizarro –disturbed–) also tended to be located on this end but a significant correlation was not observed. On the contrary, Playa Verde and La Camaronera were strongly correlated with the sedimentary geology. Total organic matter and substrate type also were important variables along this axis. In general, X1 was represented in Fig. 3a by the environmental variables measured at habitat scale and X2 by the stream-scale variables. The values for physical and chemical variables are shown in Table 2.
The dominant taxa Farrodes, Thraulodes, and Hagenulopsis were correlated with X1, while Argia and Neoelmis with X2. The abundant taxa Tricorythodes, Cloeodes, Macronema and Anacroneuria were related with X1, and Leptonema, Anchytarsus and Chironomidae sp 3 with X2. Among frequent taxa, Tanypodinae sp 1, Chironomidae sp 1, Smicridea, Polyplectropus were correlated with X1, and Hexatoma and Simulium with X2. Among rare taxa, Elodes, Acentropinae sp, and Forcipomyia were correlated with X1, while Disersus and Macrelmis were associated with X2 (Fig. 3a). The rest of the taxa in the Fig. 3a were associated with the third and fourth axes (not plotted).
Relative importance of stream, reach, and habitat scales (variance partitioning). In the CCA, environmental variables explained 13.98% of insect density variation. The pure effect of stream-scale and habitat-scale variables explained a similar amount of the total variance (5.74 and 5.01%, respectively), and the pure effect of reach-scale variables explained the least (2.19%). The amount of shared variance (SRH) (either two-way or three-way interactions) individually accounted for less than 1% of variation.
Considering that not all taxa were equally well explained by the same set of environmental variables, we used the fit for taxa and the percentage of variance fit (≥6.5) by each set of explanatory variables as measurements to find out which taxa exhibited a great percent of explained variation (Table 4). Out of the 53 taxa, 30% was mostly explained by stream-scale variables. In turn, 9% of total taxa were mostly related to habitat-scale variables. Four percent of taxa were equally explained by stream- and reach-scale variables or stream- and habitat-scale variables. Only Macrelmis was explained by reach-scale variables (Table 4).
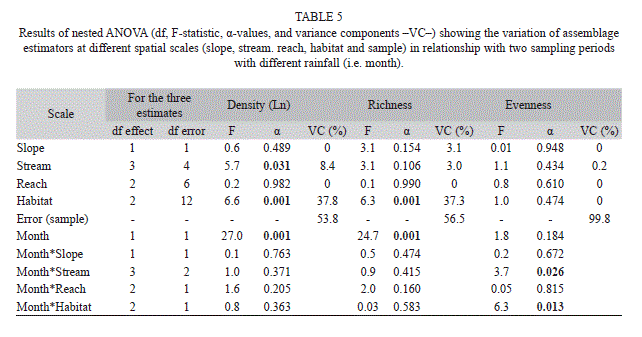
Nestedness in insect assemblage parameters and interaction with time. Total density exhibited a significant stream and habitat component of variance, however the greatest contribution was due to the latter (Table 5). The habitat component of variance was also significant for total richness. However, the error contributed to most of the variability in both variables. Total density and richness presented significant differences between the two sampling periods (nested ANOVA: main temporal effect α<0.001). However there was no significant interaction between sampling period and any spatial scale measured (Table 5). Temporal variation on total density and richness could be mediated by changes in the concentrations of DO, TN, and Cl- as shown in the X1 axis of CCA (Fig. 3b). In Fig. 3b, open symbols represented samples from streams during the high rainfall month exhibiting higher DO and TN concentrations, while filled symbols corresponded to samples from the low rainfall month, exhibiting opposite conditions. No spatial or temporal pattern was significant at any scale for Pielou’s evenness, yet a significant interaction between month and stream, and month and habitat was observed.
Discussion
Insularity on insect assemblage. The relatively high richness of aquatic insects in Gorgona Island seems to be explained by its continental origin, its proximity to the continent, and its relatively heterogeneous fluvial landscape. Therefore, we considered it a suitable ecosystem for colonization and establishment of insects. This statement is supported by the fact that out of the 191 genera reported for the entire Colombian Pacific Province (Zamora, 2002) we found 53 genera (9 orders) in Gorgona (Table 4). Furthermore, richness in Gorgona lies within the range (47-56 genera) reported for streams in the neighbouring continental ecoregions (García, Cantera, Zúñiga & Montoya, 2008; Mosquera, Palacios & Soto, 2008; Ramos, Zúñiga & Armbrecht, 2008). Our results are consistent with those found by Gómez-Aguirre (2009) in a previous survey of 9 of Gorgona’s streams during 2008 (51 genera were reported).
In addition, richness in Gorgona Island streams was one of the highest compared to other islands in the Tropical Eastern Pacific, regardless the degree of human intervention (e.g. Coiba at Panamá and Isla del Caño at Costa Rica, Gutiérrez-Fonseca, Ramírez, Umaña & Springer, 2013). While 31 families and 53 genera were recorded in Gorgona, 18 families and 27 genera were recorded in Isla del Caño (Springer, 2004), and 40 families (7 orders, no genera-level reports) in Coiba (Boyero & Bailey, 2001). In more remote oceanic islands in the mid Pacific, such as Kosrae in Micronesia, the richness is significantly lower (3 orders and 3 families: March et al., 2003; Benstead et al., 2009) than in continental islands in the Tropical Eastern Pacific.
The richest and most abundant families found in Gorgona were similar to those found in other neotropical islands (Coiba: Boyero & Bailey, 2001; Isla del Caño: Springer, 2004; Puerto Rico: Covich & McDowell, 1996, Ramírez & Hernández-Cruz, 2004). The common families in these islands were Baetidae, Leptophlebiidae, Elmidae, and Chironomidae. In Puerto Rico and Gorgona, Hydropsychidae was also abundant, while Leptohyphidae and Psephenidae were the taxa more abundant in Coiba and Gorgona.
As in other islands near continents, insect orders absent from oceanic islands were found in Gorgona Island. For instance, the stoneflies were represented by Perlidae in Gorgona, in Del Caño, and Coiba. Additionally, one species of Megaloptera (Corydalidae) was collected in Del Caño and Gorgona. Despite the small effect of insularity on Gorgona’s stream insects, the low abundance in all groups is in accordance with that found at Coiba and other tropical islands (e.g. Haynes, 1987).
In spite of the relatively high richness observed in Gorgona Island, some of the orders were represented by fewer taxa than their continental counterparts, similarly as reported by Hughes (2005) for Macaronesian archipelagos in the North Atlantic. At both geographic areas, it was found that most families contain a single genus with a few species (or even a single species such as in Macaronesia). In Gorgona Island, we found that at Playa Verde stream, for example, out of a total of 17 families, 13 were represented by a single genus. In continental streams of Neotropical Region, only Perlidae (Plecoptera) is commonly represented by a single genus (Anacroneuria)(Zúñiga, 2004).
The low richness of insect assemblages on certain islands is a consequence of the difficulties inherent in the colonization processes. Regionally, the biogeographical factors that influence potential settlers are the ability to disperse over oceanic barriers, the distance to the mainland, and the prevailing wind direction. At the island scale, the relevant variables are the age and size, the availability of running water, the hydrological regime and the types and quality of habitats (Covich, 2006).
Insect distribution across insular landscapes (spatial variance partitioning). Partial CCA indicated that the proportion of variance explained by the environmental variables measured at the three spatial scales (i.e. stream, reach, and habitat) was low (13.98%). This result could indicate that of the most species arriving to the island are randomly distributed at any stream or reach. Nevertheless, some generalities were identified within each scale.
Although Gorgona is a small island, it presents a landscape mosaic among streams, defined by forest types, geology, geomorphology, and water chemistry, this last variable was influenced by rainfall (Fig. 3a, b). These coarse-scale variables are among the most important controls of aquatic insect assemblage structure and composition in continental streams (e.g. Minshall 1984, Statzner, Gore & Resh, 1988, Richards, Haro, Johnson & Host, 1997). Hence, in pCCA the stream-scale variables explained the highest variance in the assemblage density (5.74%) and in several dominant, abundant, frequent, and rare taxa (Table 4). The sedimentary geology in La Camaronera and Playa Verde promote distinctive assemblages from the rest of the streams, which are predominantly underlined by basaltic rocks. The sedimentary geology may partially explain the high concentrations of TN and Cl- in Playa Verde and La Camaronera. Gómez-Aguirre (2009) also found a correlation between sedimentary geology and water pH. In addition, we consider that some stream-scale filters are strongly linked to current human uses. For instance, at the High Density Use zone there are many introduced plant species that currently dominate canopies and subcanopies in riparian forests in Iguapoga and Ilú streams. There also are water intakes and small dams for aqueduct and hydropower generation in Ilú and Pizarro streams.
The habitat was the second scale explaining a greater variance in the assemblage density (5.01%). We observed that type of substrate (pebbles in particular) is an important filter, which in turn is affected by geology. Therefore, as it has been recently highlighted in continental streams (Boyero & Bosch 2004, Principe, Raffaini, Gualdoni, Oberto & Corigliano, 2007, Duan, Wang & Tian, 2008), in Gorgona Island the substrate may also play a decisive role in structuring insect assemblages. Bearing in mind the dominance of pebbles and gravels in Playa Verde stream, one might explain the absence of Tricorythodes, and the lower abundance of Leptohyphes, Heterelmis, and Simulium relative to cobble-dominated streams. The high concentration of TN (4.67mg l-1) seemed particularly important in Playa Verde stream. Other variables also important at the habitat scale island-wide are the concentrations of DO, of TIM and TOM. These variables were not considered filters, but variables of influence or modulators –i.e. environmental variables that affect the density but not the presence of taxa in any moment (e.g. rainfall), hence they are not filters–, because despite temporal variations, their concentrations were within optimal ranges for the presence of fauna (Table 2).
Groups of variables at stream and habitat scales showed an important interaction (0.64%). This result is consistent with patterns found in continental streams, where it was established that flow, substrate, and water quality (habitat-scale variables) are influenced by landuse and geology –stream scale variables– (e.g. Vannote, Minshall, Cummins, Sedell & Cushing, 1980; Allan, 2004). Similarly to our study, in the continents, regional and local (e.g. land use and physical properties) variables explained a greater part of the among-site variance in insect assemblage (e.g. Minshall 1984, Li et al., 2001, Parsons, Thoms & Norris, 2004). TOM concentration and riparian cover were lower in the HDU-zone relative to Pri- and NR- zones, thus highlight the importance of past land use in Gorgona Island. This relationship has been previously reported in the literature for streams elsewhere (Allan, 2004).
The lowest percentage of variance explained (2.19%) by the reach-scale was probably due to the short segments and the low stream orders exhibited by most drainages. Despite the clear difference in slope between mountain and coastal plain reaches, the insects do not seem to discriminate between them because the habitats offered by reaches are probably equally suitable for insect colonization (e.g. Greathouse & Pringle, 2005). It must be considered that Gorgona Island streams have relatively short lengths (529±170m), low stream orders (2nd-3rd), shallow (<0.35m), and narrow (active channel width: ≤3.44m). Hence, the differences between reaches are not so remarkable, and therefore the slope was considered an influential variable (not a filter) as well as riparian coverage (%) that was lower at Ilú, Iguapoga, and Pizarro (Table 2).
Despite the significant correlations obtained in this research between environmental variables and insect density in Gorgona Island, further studies should test the importance of inter-specific interactions such as predation and bioturbation by shrimp and fish, playing an important role in many insular and coastal streams in the Neotropics (e.g. Flowers & Pringle, 1995; March et al., 2002; Ramírez & Hernández-Cruz, 2004). These processes may further explain part of the high percentage of non-explained variability in our study (86.02%). Due to the importance of interspecific interactions, Wiley, Kohler & Seelbach (1997) emphasized the relevance of considering the combination of landscape and biological variables. They found that the explained variance of Salmo trutta Linnaeus density was greater when landscape variables were combined with biotic variables [e.g. competition] (56.92%), while less variation was explained when these variables were analyzed separately (30.2 and 12.5%, respectively).
Temporal variability and rainfall. Despite of the limited temporal coverage of the study due to the prohibited costs to conduct year-round samplings, it was clear that rainfall conditions were contrasting enough between February and March to hypothesize that hydrologic disturbance may control biotic and abiotic dynamics in streams. Various studies in the tropics highlight the direct influence of rainfall and spates on assemblage’s structure (e.g. Dudgeon, 1993; Flecker & Feifarek, 1994; Ramírez & Pringle, 2006). In Gorgona Island, we found that insect density and richness was reduced during the rainy period, regardless of the spatial scale (Table 5), meaning that a reduction of density and richness during heavy rain events occur island-wide at all spatial scales in the same proportion. The effect of rainfall is mediated by specific variables operating at specific spatial scales, therefore, a greatest effect was noticed in assemblage composition. The influence of rainfall on density seems to be mediated by alterations on DO, TN, Cl-, TIM, and substratum. When these variables were included in the model, streamflow and rainfall were left out. Due to the large number of rare species, eveness does not seem to be highly variable across spatial scales. However, differences on stream and habitat characteristics (scales) may promote species-specific responses to heavy rainfall thus altering evenness. This result is consistent with the findings in the multivariate analysis, in which the largest proportion of explained variance of the density is given by the environmental variables operating at stream and habitat scales. Therefore, monitoring programs of these estimators of species diversity should be focused on these scales.
Although Gorgona has a monomodal rainfall regime with higher precipitation between May and November, the annual average rainfall is higher than that recorded in the continental coast of Colombia, being the intra-annual variation greater than the inter-annual (Blanco, 2009). Despite of the marked seasonality in mean rainfall and stream water level, there is a great variability within months (daily or weekly time scale), even during the low precipitation period. Therefore, we hypothesize that Gorgona Island stream hydrology can be regarded as aseasonal according to the criteria established by Sanford, Paaby, Luvall & Phillips (1994), despite of differences in mean rainfall between months shown in Fig. 2b. This might be similar to the observed in other wet tropical forests, such as La Selva in Costa Rica, where large amounts of rainfall can be expected at any time of the year [even during low rainfall (drier season)]. Furthermore, in Gorgona Island, changes in the stream channel geometry are frequent due to tree fall and bank collapse, which alter the habitats, flow and water chemistry and create barriers to the movement of organisms as a consequence of hydrological disturbances. Therefore, the assemblage might exhibit an aseasonal or more stable structure or an event-driven variability. Studies in other tropical regions (Wolda, 1978; Wolda & Flowers, 1985; Yule, 1996; Dudgeon, 1999, Jacobsen, Cressa, Mathooko & Dudgeon, 2008) report on the lack of relationship between insect assemblage structure and seasonal rainfall.
In summary, regarding the hypothesis, we did not observe a fully hierarchical structure or nestedness because of the following: 1) a slight proportion of the variance in insect assemblage composition, density and richness was explained by the environmental variables (filters) measured at the stream scale, 2) the variability at the reach scale was negligible, and 3) the largest proportion of the variance was observed among samples (substrate patch-scale or within habitat-scale). Of the slight portion explained, variables operating at stream and habitat scales explained the largest part of the variability. Longitudinal patterns were not significant, probably reflecting the homogenization due to the short segments of the streams, and the similarities of the instream conditions (excluding channel slope). The temporal differences observed between the two study months suggested that intra and inter-annual responses of insect assemblages should be studied with regard to rainfall and streamflow variability.
Acknowledgments
This research was funded by the GIS-BIO Project (code 551) “Gorgona Island Stream Bioassessment”, supported by the Comité para el Desarrollo de la Investigación (CODI) of Universidad de Antioquia to Juan F. Blanco, and a doctoral fellowship to Magnolia Longo from the same University. We thank to Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales de Colombia (UAESPNN-Gorgona) for issuing the research permit DTSO-G-03/08. We thank to A. Gómez, J.C. Arias, E. Parra, H. Montaño, J. Bonilla, Teo, and Alonso for assistance in the field. Thanks are also due to Armando, Goyo, Colacho, and Guasa for logistical support and sharing their knowledge about the island. Laboratory space and equipment at Universidad de Antioquia was provided by the Limnology Group (LimnoBasE-Biotamar). The following people provided taxonomic assistance: F. Muñoz, J. Posada (Trichoptera), E. Domínguez, C. Molineri, C. Nieto (Ephemeroptera), V. Manzo (Elmidae), F. Romero (Lepidoptera), S. Mazzuconi (Hemiptera) P. Torres, M. Michat, A. Bachmman (Coleoptera), E. Tejerina, A. Paggi (Chironomidae). Special thanks to J.J. Ramírez, N. Bonada, and E. Hernández for contributed with statistical analysis and commentaries, and thanks to L. Ortíz. Special thanks to the reviewers for their contributions. ELICE Publication No. 15.
References
Allan, J. D. (2004). Landscapes and riverscapes: The influence of land use on stream ecosystems. Annual Review of Ecology, Evolution, and Systematics, 35: 257-284. [ Links ]
Aristizábal-García, H. (2002). Los hemípteros de la película superficial del agua en Colombia. Parte 1: Familia Gerridae. Editorial Guadalupe LTDA: Bogotá, Colombia. [ Links ]
Benbow, M. E., McIntosh, M. D., Burky A. J. & Way C. M. (2005). The influence of stream flow reduction on the energetics of endemic Hawaiian torrenticolous aquatic insects, Telmatogeton Schiner andProcanace Hendel. Journal of Insect Conservation, 9: 175-185. [ Links ]
Benstead, J. P., March, J. G., Pringle, C. M., Ewel, K. C. & Short, J. W. (2009). Biodiversity and ecosystem function in species-poor communities: community structure and leaf litter breakdown in a Pacific island stream. Journal of the North American Benthological Society, 28: 454-465. [ Links ]
Blanco, J. F. (2009). The hydroclimatology of Gorgona Island: seasonal and ENSO-related patterns. Actualidades Biológicas, 31: 111-121. [ Links ]
Borcard, D., Legendre, P. & Drapeau, P. (1992). Partialling out the spatial component of ecological variation. Ecology, 73: 1045-1055. [ Links ]
Boulton, A. J., Boyero, L., Covich, A .P., Dobson, M., Lake, S. & Pearson, R. (2008). Are tropical streams ecologically different from temperate streams?. In D. Dudgeon (Ed.), Tropical Stream Ecology (pp. 257-284). Elsevier Science, London. [ Links ]
Boyero, L. & Bailey, R. C. (2001). Organization of macroinvertebrate communities at a hierarchy of spatial scales in a tropical stream. Hidrobiologia, 464: 219-225. [ Links ]
Boyero, L. & Bosch, J. (2004). Multiscale spatial variation of stone recolonization by macroinvertebrates in a Costa Rican stream. Journal of Tropical Ecology, 20: 85-95. [ Links ]
Cao, Y., Williams, D. D. & Williams, N. E. (1998). How important are rare species in aquatic community ecology and bioassessment? Association for the Sciences of Limnology and Oceanography, 43: 1403-1409. [ Links ]
Corkum, L. D. (1991). Spatial patterns of macroinvertebrate distributions along rivers in eastern deciduous forest and grassland biomes Journal of the North American Benthological Society, 10: 358-371. [ Links ]
Covich, A. P. & McDowell, W. H. (1996). The stream community. In D. P. Reagan & R. B. Waide (Eds.), The food web of a tropical rain forest (pp. 434-459). University of Chicago Press, Chicago. [ Links ]
Covich, A. P., Crowl, T. A. & Scatena, F. N. (2003). Effects of extreme low flows on freshwater shrimps in a perennial tropical stream. Freshwater Biology, 48: 1199-1206. [ Links ]
Covich, A. P. (2006). Dispersal-limited biodiversity of tropical insular streams. Polish Journal of Ecology, 54: 523-547. [ Links ]
Craig, D. A. (2003). Geomorphology, development of running water habitats, and evolution of black flies on Polynesian Islands. BioScience, 53: 1079-1093. [ Links ]
Domínguez, E. & Fernández, H. (2009). Macroinvertebrados bentónicos sudamericanos: sistemática y biología. Fundación Miguel Lillo: Argentina. [ Links ]
Duan, X., Wang, Z. & Tian, S. (2008). Effect of streambed substrate on macroinvertebrate biodiversity. Frontiers of Environmental Science & Engineering, 2: 122-128. [ Links ]
Dudgeon, D. (1993). The effects of spate-induced disturbance, predation and environmental complexity on macroinvertebrates in a tropical stream. Freshwater Biology, 30: 189-197. [ Links ]
Dudgeon, D. (1999). The population dynamics of three species of Calamoceratidae (Trichoptera) in a tropical forest stream. In H. Malicky & P. Chantaramongkol (Eds.), Proceedings of the 9th International Symposium on Trichoptera (pp. 83-91). University of Chiang Mai. [ Links ]
Englund, R. A. (1999). The impacts of introduced Poeciliid fish and Odonata on the endemic Megalagrion (Odonata) damselflies of Oahu Island, Hawaii. Journal of Insect Conservation, 3: 225-243. [ Links ]
Flecker, A. S. & Feifarek, B. P. (1994). Disturbance and the temporal variability of insect assemblages in two Andean streams. Freshwater Biology, 31: 131-142. [ Links ]
Flowers, R. W. & Pringle, C. M. (1995). Yearly fluctuations in the mayfly community of a tropical stream draining lowland pasture in Costa Rica. In L. D. Corkum & J. H. Ciborowski (Eds.), Current Directions in Research on Ephemeroptera (pp. 131-150). Canadian Scholars’ Press, Toronto. [ Links ]
Frissell, C. A., Liss, W. J., Warren, C. E. & Hurley, M. D. (1986). A hierarchical framework for stream habitat classification: viewing streams in a watershed context. Journal of Environmental Management,10: 199-214. [ Links ]
García, J. F., Cantera, J., Zúñiga, M. & Montoya, J. (2008). Estructura y diversidad de las comunidades de macroinvertebrados acuáticos en la cuenca baja del río Dagua (Andén Pacífico Vallecaucano-Colombia). Revista de Ciencias de la Universidad del Valle, 13: 27-48. [ Links ]
Gómez-Aguirre, A. M. (2009). Variación de la estructura y composición de los ensamblajes de macroinvertebrados bénticos en la jerarquía espacial de los sistemas lóticos del Parque Nacional Natural Gorgona (Undergraduate Dissertation). Universidad de Antioquia, Colombia. [ Links ]
Gómez-Aguirre, A. M., Longo-Sánchez, M. & Blanco, J. F. (2009). Macroinvertebrate assemblages in Gorgona Island streams: spatial patterns during two contrasting hydrologic periods. Actualidades Biológicas, 31: 161-178. [ Links ]
Greathouse, E. A. & Pringle, C. M. (2005). Does the river continuum concept apply on a tropical island? Longitudinal variation in a Puerto Rican stream. Canadian Journal of Fisheries and Aquatic Sciences, 63: 134-152. [ Links ]
Gutiérrez-Fonseca, P. E., Ramírez, A., Umaña, G. & Springer, M. (2013). Macroinvertebrados dulceacuícolas de la Isla del Coco, Costa Rica: especies y comparación con otras islas del Pacífico Tropical Oriental. Revista de Biología Tropical, 61: 657-668. [ Links ]
Haynes, A. (1987). Species richness, abundance and biomass of benthic invertebrates in a lowland tropical stream on the island of Viti Levu, Fiji. Archiv für Hydrobiologie, 110: 451-459. [ Links ]
Hendrich, L. & Yang, C. M. (1999). A contribution to the knowledge of the water beetle fauna of Pulau Tioman, peninsular Malaysia (Coleoptera: Noteridae, Dytiscidae, Hydrophilidae, Hydraenidae, Scirtidae, Limnichidae). Raffles Bulletin of Zoology, 6: 253-262. [ Links ]
Hill, M.O. & Gauch, H.G. (1980). Detrended correspondence analysis, an improvised ordination technique. Vegetation, 42: 47-58. [ Links ]
Hughes, S. J. (2005). Application of the Water Framework Directive to Macaronesian freshwater systems. Biology and Environment, 105: 185-193. [ Links ]
Jackson, J. K. & Sweeney, B. W. (1995). Egg and larval development times for 35 species of tropical stream insects from Costa Rica. Journal of the North American Benthological Society, 14: 115-130. [ Links ]
Jacobsen, D., Cressa, C., Mathooko, J. M. & Dudgeon, D. (2008). Macroinvertebrates: Composition, life histories and production. In D. Dudgeon (Ed.), Tropical Stream Ecology (pp. 65-105). Elsevier Science, London. [ Links ]
Kerr, A. C. (2005). La Isla de Gorgona, Colombia: A petrological enigma? Lithos, 84: 77-101. [ Links ]
Lepŝ, J. & Ŝmilauer, P. (2003). Multivariate analysis of ecological data using CANOCO. Cambridge University Press. Cambridge, UK. [ Links ]
Li, J., Herlihy, A., Gerth, W., Kaufmann, P., Gregory, S., Urquhart S. &. Larsen D. P. (2001). Variability in stream macroinvertebrates at multiple spatial scales. Freshwater Biology, 46: 87-97. [ Links ]
Magurran, A. E. (2004). Measuring biological diversity. Blackwell: Oxford. [ Links ]
March, J. G., Pringle, C. M., Townsend, M .J. & Wilson, A. I. (2002). Effects of freshwater shrimp assemblages on benthic communities along an altitudinal gradient of a tropical island stream. Freshwater Biology, 47: 377-390. [ Links ]
March, J. G., Benstead, J. P., Pringle, C. M. & Luckymis, M. (2003). Benthic community structure and invertebrate drift in a Pacific island stream, Kosrae, Micronesia. Biotropica, 35: 125-130. [ Links ]
Minshall, G.W. (1984). Aquatic insect-substratum relationships. In V. H. Resh & D. M. Rosenberg (Eds.), The Ecology of Aquatic Insects (pp. 358-400). Praeger Scientific, New York. [ Links ]
Mosquera, R. D., Palacios, M. L. & Soto, A. (2008). Bioindicación de la calidad del agua del río Cali usando macroinvertebrados acuáticos. In Asociación Colombiana de Limnología (Ed.). Resúmenes VIII Seminario Colombiano de Limnología y I Reunión Internacional de Cuencas Bajas y Zonas Estuarinas (pp.117). Colombia. [ Links ]
Mueller-Dombois, D. (2002). Forest vegetation across the Pacific: a biogeographically complex region with many analogous habitats. Plant Ecology, 163:155-176. [ Links ]
Parsons, M., Thoms, M. C. & Norris, R. H. (2004). Using hierarchy to select scales of measurement in multiscale studies of stream macroinvertebrate assemblages. Journal of the North American Benthological Society, 23: 157-170. [ Links ]
Pérez-Gutiérrez, L. A. (2003). Estudio biotaxonómico de los odonatos (Insecta: Odonata Fabricius, 1793) del distrito de Santa Marta (Magdalena-Colombia) (Undergraduate dissertation), Universidad del Magdalena, Colombia. [ Links ]
Poff, N. L. (1997). Landscape filters and species traits: Toward mechanistic understanding and prediction in stream ecology. Journal of the North American Benthological Society. 16: 391-409. [ Links ]
Polhemus, D .A. (1995). New Heteroptera and Odonata (Insecta) records and range extensions in the Hawaiian Islands. Bishop Museum Occasional Papers, 42: 42-43. [ Links ]
Principe, R. E., Raffaini, G. B., Gualdoni, C. M., Oberto A. M. & Corigliano M. C. (2007). Do hydraulic units define macroinvertebrate assemblages in mountain streams of central Argentina? Limnologica, 37: 323-336. [ Links ]
Rangel, O. 1995. Islas de Gorgona y Gorgonilla. In J. I. Rangel (Ed.), Diversidad Biótica ICN-UNAL-INDERENA (pp. 145-154). Colombia. [ Links ]
Ramírez, A. & Hernández-Cruz, L. R. (2004). Aquatic insect assemblages in shrimp-dominated tropical streams, Puerto Rico. Biotropica, 36: 259-266. [ Links ]
Ramírez, A. & Pringle, C. M. (2006). Temporal and spatial patterns in stream physicochemistry and insect assemblages in tropical lowland streams. Journal of the North American Benthological Society, 25:108-125. [ Links ]
Ramos, B. C., Zúñiga, M. & Armbrecht, I. (2008). Impacto de efluentes de minas de carbón sobre los macroinvertebrados. In Asociación Colombiana de Limnología (Eds.), Resúmenes VIII Seminario Colombiano de Limnología y I Reunión Internacional de Cuencas Bajas y Zonas Estuarinas (pp. 121). Colombia. [ Links ]
Richards, C., Haro, R. J., Johnson L. B. & Host G. E. (1997). Catchment and reach-scale properties as indicators of macroinvertebrate species traits. Freshwater Biology, 37: 219-230. [ Links ]
Roldán, G. (1988). Guía para el estudio de los macroinvertebrados acuáticos del Departamento de Antioquia. Fondo FEN-COLCIENCIAS: Colombia. [ Links ]
Sandin, L. & Johnson, R. K. (2004). Local, landscape and regional factors structuring benthic macroinvertebrate assemblages in Swedish streams. Landscape Ecology, 19: 501-514. [ Links ]
Sanford, R. L., Paaby, P., Luvall, J. C. & Phillips, E. (1994). Climate, geomorphology, and aquatic systems. In La Selva: L.A. McDade, K.S. Bawa, H.A. Hespenheide & G.S. Hartshorn (Eds.), Ecology and natural history of a Neotropical rainforest (pp. 19-33). University of Chicago Press, Chicago. [ Links ]
Smith, G. C., Covich, A. P. & Brasher, A. M. D. (2003). An ecological perspective on the biodiversity of tropical island streams. BioScience, 53: 1048-1051. [ Links ]
Springer M. (2004). Primer listado de insectos acuáticos de la isla del Caño, Costa Rica. Brenesia 62: 97-98. [ Links ]
Statzner, B., Gore, J. A. & Resh, V. H. (1988). Hydraulic stream ecology: Observed patterns and potential applications. Journal of the North American Benthological Society, 7: 307-360. [ Links ]
Ter Braak, C. J. F. & Šmilauer, T. (2002). CANOCO reference manual and user’s guide to CANOCO for windows: Software for canonical community ordination (version 4.5). Microcomputer Power: New York. [ Links ]
UAESPNN. (2005). Plan de Manejo Parque Nacional Natural Gorgona. Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales de Colombia (Ed.). Dirección Territorial Suroccidental, SUT 021105, Colombia. [ Links ]
Vannote, R. L., Minshall, G. W., Cummins, D. W., Sedell, J. R. & Cushing, C. E. (1980). The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences, 37: 130-137. [ Links ]
Wiggins, G. B. (1996). Larvae of the North American caddisfly genera (Trichoptera) (2nd edition). University of Toronto Press, Canada. [ Links ]
Wiley, M. J., Kohler, S. L. & Seelbach, P. W. (1997). Reconciling landscape and local views of aquatic communities: lessons from Michigan trout streams. Freshwater Biology, 37: 133-148. [ Links ]
Wolda, H. (1978). Seasonal fluctuations in rainfall, food and abundance of tropical insects. Journal of Animal Ecology, 47: 369-381. [ Links ]
Wolda, H. & Flowers, R. W. (1985). Seasonality and diversity of mayfly adults (Ephemeroptera) in a nonseasonal environment. Biotropica, 17: 330-335. [ Links ]
Yule, C. M. (1996). Spatial distribution of the invertebrate fauna of an aseasonal tropical stream on Bougainville Island, Papua New Guinea. Archiv für Hydrobiologie, 137: 227-249. [ Links ]
Zamora, H. (2002). Análisis biogeográfico de los Macroinvertebrados Acuáticos Epicontinentales (MAE) en el departamento del Cauca, Colombia. Revista de la Asociación Colombiana de Ciencias Biológicas,14: 37-51. [ Links ]
Zúñiga, M. 2004. Biodiversidad, distribución y ecología del orden Plecoptera (Insecta) en Colombia: potencial en bioindicación de calidad de agua. In Asociación Colombiana de Limnología (Eds.), Resúmenes VI Seminario Colombiano de Limnología y I reunión Internacional sobre Embalses Neotropicales (pp. 17-21). Universidad Pontificia Bolivariana, Colombia. [ Links ]
Aristizábal-García, H. (2002). Los hemípteros de la película superficial del agua en Colombia. Parte 1: Familia Gerridae. Editorial Guadalupe LTDA: Bogotá, Colombia. [ Links ]
Benbow, M. E., McIntosh, M. D., Burky A. J. & Way C. M. (2005). The influence of stream flow reduction on the energetics of endemic Hawaiian torrenticolous aquatic insects, Telmatogeton Schiner andProcanace Hendel. Journal of Insect Conservation, 9: 175-185. [ Links ]
Benstead, J. P., March, J. G., Pringle, C. M., Ewel, K. C. & Short, J. W. (2009). Biodiversity and ecosystem function in species-poor communities: community structure and leaf litter breakdown in a Pacific island stream. Journal of the North American Benthological Society, 28: 454-465. [ Links ]
Blanco, J. F. (2009). The hydroclimatology of Gorgona Island: seasonal and ENSO-related patterns. Actualidades Biológicas, 31: 111-121. [ Links ]
Borcard, D., Legendre, P. & Drapeau, P. (1992). Partialling out the spatial component of ecological variation. Ecology, 73: 1045-1055. [ Links ]
Boulton, A. J., Boyero, L., Covich, A .P., Dobson, M., Lake, S. & Pearson, R. (2008). Are tropical streams ecologically different from temperate streams?. In D. Dudgeon (Ed.), Tropical Stream Ecology (pp. 257-284). Elsevier Science, London. [ Links ]
Boyero, L. & Bailey, R. C. (2001). Organization of macroinvertebrate communities at a hierarchy of spatial scales in a tropical stream. Hidrobiologia, 464: 219-225. [ Links ]
Boyero, L. & Bosch, J. (2004). Multiscale spatial variation of stone recolonization by macroinvertebrates in a Costa Rican stream. Journal of Tropical Ecology, 20: 85-95. [ Links ]
Cao, Y., Williams, D. D. & Williams, N. E. (1998). How important are rare species in aquatic community ecology and bioassessment? Association for the Sciences of Limnology and Oceanography, 43: 1403-1409. [ Links ]
Corkum, L. D. (1991). Spatial patterns of macroinvertebrate distributions along rivers in eastern deciduous forest and grassland biomes Journal of the North American Benthological Society, 10: 358-371. [ Links ]
Covich, A. P. & McDowell, W. H. (1996). The stream community. In D. P. Reagan & R. B. Waide (Eds.), The food web of a tropical rain forest (pp. 434-459). University of Chicago Press, Chicago. [ Links ]
Covich, A. P., Crowl, T. A. & Scatena, F. N. (2003). Effects of extreme low flows on freshwater shrimps in a perennial tropical stream. Freshwater Biology, 48: 1199-1206. [ Links ]
Covich, A. P. (2006). Dispersal-limited biodiversity of tropical insular streams. Polish Journal of Ecology, 54: 523-547. [ Links ]
Craig, D. A. (2003). Geomorphology, development of running water habitats, and evolution of black flies on Polynesian Islands. BioScience, 53: 1079-1093. [ Links ]
Domínguez, E. & Fernández, H. (2009). Macroinvertebrados bentónicos sudamericanos: sistemática y biología. Fundación Miguel Lillo: Argentina. [ Links ]
Duan, X., Wang, Z. & Tian, S. (2008). Effect of streambed substrate on macroinvertebrate biodiversity. Frontiers of Environmental Science & Engineering, 2: 122-128. [ Links ]
Dudgeon, D. (1993). The effects of spate-induced disturbance, predation and environmental complexity on macroinvertebrates in a tropical stream. Freshwater Biology, 30: 189-197. [ Links ]
Dudgeon, D. (1999). The population dynamics of three species of Calamoceratidae (Trichoptera) in a tropical forest stream. In H. Malicky & P. Chantaramongkol (Eds.), Proceedings of the 9th International Symposium on Trichoptera (pp. 83-91). University of Chiang Mai. [ Links ]
Englund, R. A. (1999). The impacts of introduced Poeciliid fish and Odonata on the endemic Megalagrion (Odonata) damselflies of Oahu Island, Hawaii. Journal of Insect Conservation, 3: 225-243. [ Links ]
Flecker, A. S. & Feifarek, B. P. (1994). Disturbance and the temporal variability of insect assemblages in two Andean streams. Freshwater Biology, 31: 131-142. [ Links ]
Flowers, R. W. & Pringle, C. M. (1995). Yearly fluctuations in the mayfly community of a tropical stream draining lowland pasture in Costa Rica. In L. D. Corkum & J. H. Ciborowski (Eds.), Current Directions in Research on Ephemeroptera (pp. 131-150). Canadian Scholars’ Press, Toronto. [ Links ]
Frissell, C. A., Liss, W. J., Warren, C. E. & Hurley, M. D. (1986). A hierarchical framework for stream habitat classification: viewing streams in a watershed context. Journal of Environmental Management,10: 199-214. [ Links ]
García, J. F., Cantera, J., Zúñiga, M. & Montoya, J. (2008). Estructura y diversidad de las comunidades de macroinvertebrados acuáticos en la cuenca baja del río Dagua (Andén Pacífico Vallecaucano-Colombia). Revista de Ciencias de la Universidad del Valle, 13: 27-48. [ Links ]
Gómez-Aguirre, A. M. (2009). Variación de la estructura y composición de los ensamblajes de macroinvertebrados bénticos en la jerarquía espacial de los sistemas lóticos del Parque Nacional Natural Gorgona (Undergraduate Dissertation). Universidad de Antioquia, Colombia. [ Links ]
Gómez-Aguirre, A. M., Longo-Sánchez, M. & Blanco, J. F. (2009). Macroinvertebrate assemblages in Gorgona Island streams: spatial patterns during two contrasting hydrologic periods. Actualidades Biológicas, 31: 161-178. [ Links ]
Greathouse, E. A. & Pringle, C. M. (2005). Does the river continuum concept apply on a tropical island? Longitudinal variation in a Puerto Rican stream. Canadian Journal of Fisheries and Aquatic Sciences, 63: 134-152. [ Links ]
Gutiérrez-Fonseca, P. E., Ramírez, A., Umaña, G. & Springer, M. (2013). Macroinvertebrados dulceacuícolas de la Isla del Coco, Costa Rica: especies y comparación con otras islas del Pacífico Tropical Oriental. Revista de Biología Tropical, 61: 657-668. [ Links ]
Haynes, A. (1987). Species richness, abundance and biomass of benthic invertebrates in a lowland tropical stream on the island of Viti Levu, Fiji. Archiv für Hydrobiologie, 110: 451-459. [ Links ]
Hendrich, L. & Yang, C. M. (1999). A contribution to the knowledge of the water beetle fauna of Pulau Tioman, peninsular Malaysia (Coleoptera: Noteridae, Dytiscidae, Hydrophilidae, Hydraenidae, Scirtidae, Limnichidae). Raffles Bulletin of Zoology, 6: 253-262. [ Links ]
Hill, M.O. & Gauch, H.G. (1980). Detrended correspondence analysis, an improvised ordination technique. Vegetation, 42: 47-58. [ Links ]
Hughes, S. J. (2005). Application of the Water Framework Directive to Macaronesian freshwater systems. Biology and Environment, 105: 185-193. [ Links ]
Jackson, J. K. & Sweeney, B. W. (1995). Egg and larval development times for 35 species of tropical stream insects from Costa Rica. Journal of the North American Benthological Society, 14: 115-130. [ Links ]
Jacobsen, D., Cressa, C., Mathooko, J. M. & Dudgeon, D. (2008). Macroinvertebrates: Composition, life histories and production. In D. Dudgeon (Ed.), Tropical Stream Ecology (pp. 65-105). Elsevier Science, London. [ Links ]
Kerr, A. C. (2005). La Isla de Gorgona, Colombia: A petrological enigma? Lithos, 84: 77-101. [ Links ]
Lepŝ, J. & Ŝmilauer, P. (2003). Multivariate analysis of ecological data using CANOCO. Cambridge University Press. Cambridge, UK. [ Links ]
Li, J., Herlihy, A., Gerth, W., Kaufmann, P., Gregory, S., Urquhart S. &. Larsen D. P. (2001). Variability in stream macroinvertebrates at multiple spatial scales. Freshwater Biology, 46: 87-97. [ Links ]
Magurran, A. E. (2004). Measuring biological diversity. Blackwell: Oxford. [ Links ]
March, J. G., Pringle, C. M., Townsend, M .J. & Wilson, A. I. (2002). Effects of freshwater shrimp assemblages on benthic communities along an altitudinal gradient of a tropical island stream. Freshwater Biology, 47: 377-390. [ Links ]
March, J. G., Benstead, J. P., Pringle, C. M. & Luckymis, M. (2003). Benthic community structure and invertebrate drift in a Pacific island stream, Kosrae, Micronesia. Biotropica, 35: 125-130. [ Links ]
Minshall, G.W. (1984). Aquatic insect-substratum relationships. In V. H. Resh & D. M. Rosenberg (Eds.), The Ecology of Aquatic Insects (pp. 358-400). Praeger Scientific, New York. [ Links ]
Mosquera, R. D., Palacios, M. L. & Soto, A. (2008). Bioindicación de la calidad del agua del río Cali usando macroinvertebrados acuáticos. In Asociación Colombiana de Limnología (Ed.). Resúmenes VIII Seminario Colombiano de Limnología y I Reunión Internacional de Cuencas Bajas y Zonas Estuarinas (pp.117). Colombia. [ Links ]
Mueller-Dombois, D. (2002). Forest vegetation across the Pacific: a biogeographically complex region with many analogous habitats. Plant Ecology, 163:155-176. [ Links ]
Parsons, M., Thoms, M. C. & Norris, R. H. (2004). Using hierarchy to select scales of measurement in multiscale studies of stream macroinvertebrate assemblages. Journal of the North American Benthological Society, 23: 157-170. [ Links ]
Pérez-Gutiérrez, L. A. (2003). Estudio biotaxonómico de los odonatos (Insecta: Odonata Fabricius, 1793) del distrito de Santa Marta (Magdalena-Colombia) (Undergraduate dissertation), Universidad del Magdalena, Colombia. [ Links ]
Poff, N. L. (1997). Landscape filters and species traits: Toward mechanistic understanding and prediction in stream ecology. Journal of the North American Benthological Society. 16: 391-409. [ Links ]
Polhemus, D .A. (1995). New Heteroptera and Odonata (Insecta) records and range extensions in the Hawaiian Islands. Bishop Museum Occasional Papers, 42: 42-43. [ Links ]
Principe, R. E., Raffaini, G. B., Gualdoni, C. M., Oberto A. M. & Corigliano M. C. (2007). Do hydraulic units define macroinvertebrate assemblages in mountain streams of central Argentina? Limnologica, 37: 323-336. [ Links ]
Rangel, O. 1995. Islas de Gorgona y Gorgonilla. In J. I. Rangel (Ed.), Diversidad Biótica ICN-UNAL-INDERENA (pp. 145-154). Colombia. [ Links ]
Ramírez, A. & Hernández-Cruz, L. R. (2004). Aquatic insect assemblages in shrimp-dominated tropical streams, Puerto Rico. Biotropica, 36: 259-266. [ Links ]
Ramírez, A. & Pringle, C. M. (2006). Temporal and spatial patterns in stream physicochemistry and insect assemblages in tropical lowland streams. Journal of the North American Benthological Society, 25:108-125. [ Links ]
Ramos, B. C., Zúñiga, M. & Armbrecht, I. (2008). Impacto de efluentes de minas de carbón sobre los macroinvertebrados. In Asociación Colombiana de Limnología (Eds.), Resúmenes VIII Seminario Colombiano de Limnología y I Reunión Internacional de Cuencas Bajas y Zonas Estuarinas (pp. 121). Colombia. [ Links ]
Richards, C., Haro, R. J., Johnson L. B. & Host G. E. (1997). Catchment and reach-scale properties as indicators of macroinvertebrate species traits. Freshwater Biology, 37: 219-230. [ Links ]
Roldán, G. (1988). Guía para el estudio de los macroinvertebrados acuáticos del Departamento de Antioquia. Fondo FEN-COLCIENCIAS: Colombia. [ Links ]
Sandin, L. & Johnson, R. K. (2004). Local, landscape and regional factors structuring benthic macroinvertebrate assemblages in Swedish streams. Landscape Ecology, 19: 501-514. [ Links ]
Sanford, R. L., Paaby, P., Luvall, J. C. & Phillips, E. (1994). Climate, geomorphology, and aquatic systems. In La Selva: L.A. McDade, K.S. Bawa, H.A. Hespenheide & G.S. Hartshorn (Eds.), Ecology and natural history of a Neotropical rainforest (pp. 19-33). University of Chicago Press, Chicago. [ Links ]
Smith, G. C., Covich, A. P. & Brasher, A. M. D. (2003). An ecological perspective on the biodiversity of tropical island streams. BioScience, 53: 1048-1051. [ Links ]
Springer M. (2004). Primer listado de insectos acuáticos de la isla del Caño, Costa Rica. Brenesia 62: 97-98. [ Links ]
Statzner, B., Gore, J. A. & Resh, V. H. (1988). Hydraulic stream ecology: Observed patterns and potential applications. Journal of the North American Benthological Society, 7: 307-360. [ Links ]
Ter Braak, C. J. F. & Šmilauer, T. (2002). CANOCO reference manual and user’s guide to CANOCO for windows: Software for canonical community ordination (version 4.5). Microcomputer Power: New York. [ Links ]
UAESPNN. (2005). Plan de Manejo Parque Nacional Natural Gorgona. Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales de Colombia (Ed.). Dirección Territorial Suroccidental, SUT 021105, Colombia. [ Links ]
Vannote, R. L., Minshall, G. W., Cummins, D. W., Sedell, J. R. & Cushing, C. E. (1980). The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences, 37: 130-137. [ Links ]
Wiggins, G. B. (1996). Larvae of the North American caddisfly genera (Trichoptera) (2nd edition). University of Toronto Press, Canada. [ Links ]
Wiley, M. J., Kohler, S. L. & Seelbach, P. W. (1997). Reconciling landscape and local views of aquatic communities: lessons from Michigan trout streams. Freshwater Biology, 37: 133-148. [ Links ]
Wolda, H. (1978). Seasonal fluctuations in rainfall, food and abundance of tropical insects. Journal of Animal Ecology, 47: 369-381. [ Links ]
Wolda, H. & Flowers, R. W. (1985). Seasonality and diversity of mayfly adults (Ephemeroptera) in a nonseasonal environment. Biotropica, 17: 330-335. [ Links ]
Yule, C. M. (1996). Spatial distribution of the invertebrate fauna of an aseasonal tropical stream on Bougainville Island, Papua New Guinea. Archiv für Hydrobiologie, 137: 227-249. [ Links ]
Zamora, H. (2002). Análisis biogeográfico de los Macroinvertebrados Acuáticos Epicontinentales (MAE) en el departamento del Cauca, Colombia. Revista de la Asociación Colombiana de Ciencias Biológicas,14: 37-51. [ Links ]
Zúñiga, M. 2004. Biodiversidad, distribución y ecología del orden Plecoptera (Insecta) en Colombia: potencial en bioindicación de calidad de agua. In Asociación Colombiana de Limnología (Eds.), Resúmenes VI Seminario Colombiano de Limnología y I reunión Internacional sobre Embalses Neotropicales (pp. 17-21). Universidad Pontificia Bolivariana, Colombia. [ Links ]
1. Instituto de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Medellín, Colombia; mc_longo@hotmail.com
2. Instituto de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Medellín, Colombia; blanco@matematicas.udea.edu.co
Recibido 18-X-2013. Corregido 20-XI-2013. Aceptado 19-XII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}