










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.4 San José Oct./Dec. 2014
Ethnic background and CYP2D6 genetic polymorphisms in Costa Ricans
Antecedentes étnicos y polimorfismo genético del CYP2D6 en los costarricenses
Abstract
Key words: CYP2D6,
Resumen
El Consorcio de la Red Iberoamericana de Farmacogenética CEIBA.FP ha demostrado que existen diferencias en cuanto a CYP2D6 en las poblaciones latinoamericanas. Sin embargo, hasta ahora, se sabe poco de este gen de importancia farmacogenética en la población de Costa Rica, la cual tiene una ancestría trihíbrida.
Los genotipos CYP2D6 fueron determinados por XL-PCR y PCR tiempo real. Se determinaron las variantes alélicas *2, *3, *4, *5, *6, *10, *17, *29, *35 y *41. Para la población total estudiada las frecuencia de PMs y UMs fueron respectivamente 6% y 6.5%. El porcentaje de individuos UMs fue mayor en la población mestiza que en la amerindia.
La frecuencia de UMs varió de 3.6 a 10.1% y la de PMs de 1.4 a 10.1% en los grupos costarricenses. Las frecuencias más altas de UMs (10.1%) y de PMs (10.2%) se encontraron respectivamente en las poblaciones mestiza y amerindia. En conclusión, las frecuencias de UMs y PMs de CYP2D6 varían ampliamente en las poblaciones mestiza, amerindia y afro-caribeña de Costa Rica. Investigaciones
futuras en la población de Costa Rica deberían orientarse a identificar nuevas variantes del CYP2D6 mediante métodos de secuenciación, así como a determinar el fenotipo de CYP2D6 con el objetivo de establecer la relación fenotipo-genotipo. Finalmente, es necesario realizar estudios adicionales que involucren marcadores genéticos de ancestría en la población costarricense.
Palabras clave: CYP2D6, Costa Rica, amerindios, afrocaribeños, mestizos, poblaciones, metabolizadores lentos, metabolizadores ultra-rápidos.
CYP2D6 is involved in the metabolism of widely used drugs, such as antidepressants, antipsychotics, antihypertensives, analgesics, and beta-blockers (Ingelman-Sundberg, 2005).
The CYP2D6 gene, located on chromosome 22q13.1, is highly polymorphic, with alleles causing absent, reduced, normal and increased catalytic activity (CYP Alleles Nomenclature Database).
Inter-ethnic differences in such cytochrome P450 polymorphism might be partially responsible for the variations in drug disposition among populations. During the 16th century, people from the Iberian Peninsula arrived to different places of
A high frequency of Ultra-rapid Metabolizers (UMs) has been previously described in Spanish population (Llerena, Dorado, & Peñas-Lledó, 2009; Peñas-Lledó et al., 2012).
Given the
In Central American countries, there are just a few studies in Panama (Arias et al., 1986; Arias, Jorge, Lee, Barrantes, & Inaba, 1988), Nicaragua (Llerena et al., 2012; Llerena et al., 2013) and Costa Rica (reporting some alleles of a Costa Rican Amerindian population) (Jorge & Arias, 1995). CYP2D6 genetic polymorphisms studies have been carried out in Latin American countries supported by the CEIBA. FP Consortium of the Ibero-American Network of Pharmacogenetics & Pharmacogenomics (RIBEF) (de Andrés et al., 2013; Rodeiro et al., 2012), including Ecuadorians (Dorado et al., 2012), Mexicans (Sosa-Macías, Dorado, Alanis- Bañuelos, Llerena, & Lares-Asseff, 2010) and Cubans (González et al., 2008; Llerena et al., 2012; Llerena et al., 2013; Peñas-Lledó,
Dorado, Pacheco, González, & Llerena, 2009). However, this will constitute the first report of a Costa Rican population including groups from different ethnic backgrounds.
The present study aimed to determine the frequency of UMs and Poor Metabolizers (PMs) in a Costa Rican population, as well as to determine whether there are differences in CYP2D6-predicted phenotype frequencies among three Costa Rican groups with different ethnic backgrounds. Additionally, the frequency of PMs and UMs obtained in this study was compared with published data from Ibero-American populations, and finally, this study also aimed to describe allele frequencies among different Costa Rican ethnic groups.
Materials and methods
Subjects: The study comprised 385 healthy individuals belonging to three ethnic groups: Amerindian (n=197), Afro-Caribbean (n=49) and mestizo (n=139).
The DNA samples were obtained from a DNA biobank of the
All the samples were already classified according to the ethnic origin and previous studies (see inclusion criteria), codified, and stored with an ID. Demographic data of these populations are available elsewhere (Barrantes, 1989; Madrigal, 2006; Morera et al., 2003).
The inclusion criteria were:
Phenotype features: For the Amerindian population, the criteria were copper-colored skin, straight hair, slanted eyes, and short stature; in the Afro-Caribbean population, dark skin, curly hair, flat nose, and prominent cheekbones were the criteria; the Mestizo population comprised all those subjects not included in any of the aforementioned groups.
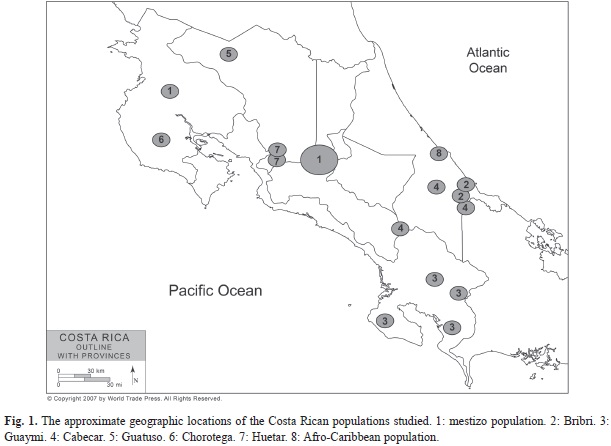
Places of residence: For the Amerindian group, the places of residence were Matambu Indian locality (Chorotega), the South and the Pacific area (Guaymi), the
Atlantic Talamanca and the Pacific area (Cabecar), the Talamanca area (Bribri), the Quitirrisi and Zapaton Indian localities (Huetar), and the Margarita and Tonjibe Guatuso Indian localities (Guatuso or Maleku). The samples of the Afro- Caribbean population were collected from volunteers living in the Atlantic coastal region of Limon. The mestizo population was selected from people living in the Guanacaste region, and in the Western or Central Valley of Costa Rica (Fig. 1). The inclusion of an individual in a group excluded that individual from being part of any other population.
Genetic markers such as blood group systems: ABO (O for Amerindians and B for the Afro-Caribbeans), rhesus, MNS, P, Kell, Kidd, Duffy, Diego, and Lewis; plasmatic protein systems (albumin, transferrin, haptoglobin, and ceruloplasmin) (Barrantes et al., 1990; Bieber, Bieber, Rodewald, & Barrantes, 1996); single nucleotide polymorphisms (SNPs) (Herrmann et al., 2002); microsatellites (Wang et al., 2007); mitochondrial DNA (Santos et al., 1994) and chromosome Y (Ruiz- Narváez et al., 2005).
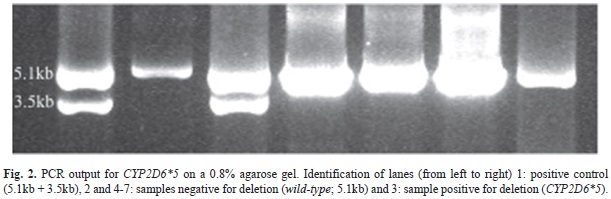
Genotyping procedure: For genotyping, the CEIBA.FP Consortium methodology was followed. To detect the presence of allelic variants harboring a CYP2D6*5 gene deletion or a duplication, long range (XL)-PCR was performed as described in detail elsewhere (Fig. 2) (Dorado et al., 2005). Subjects positive for a duplication or deletion were further characterized for gene copy number with the TaqMan assay Hs00010001_cn, which specifically amplifies exon 9 sequences and does not amplify CYP2D7 or CYP2D8 pseudogenes or CYP2D6/CYP2D7 hybrids alleles carrying CYP2D7 exon 9 sequences. Genotype analysis for the CYP2D6*2 (-1584 C>G), *3 (2549A>del), *4 (100 C>T, 1846G>A), *6 (1707 T>del), *10 (100 C>T), *17 (1023 C>T), *29 (3183 G>A), *35 (-1584 C>G and 31 G>A), and *41 (2988 G>A) allelic variants was carried out on genomic DNA, using commercially available TaqMan assays as previously described (Dorado et al., 2012). To discriminate among CYP2D6*1xN, *2xN, *4xN and *10xN alleles, a 10kb long XL-PCR fragment was generated from duplication-positive subjects and tested for respective SNPs by an established PCR-RFLP approach (Dorado et al., 2005).
Predicted hydroxylation capacity group:
In order to extrapolate genetic data to metabolic phenotype information, an activity score was utilized as previously described (Gaedigk et al., 2008; Llerena et al., 2012).
Zero value was assigned to CYP2D6*3, *4, *4XN, *5, *6 variants; 0.5 to each copy of CYP2D6*10, *17, *29, *41 alleles; one was assigned to CYP2D6 wt, *2, *35, and more than two (depending on the number of copies) to the multiplication of the active alleles (wtxN, *2xN). Individuals with zero active genes were
classified as poor metabolizers (PMs), and those with more than two active gene copies were classified as Ultra-rapid Metabolizers (UMs) (Gaedigk et al., 2008; Llerena et al., 2012).
The differences in CYP2D6 allele frequencies were compared using the X2-test and/or Fishers exact test. P-values <0.05 were regarded as statistically significant. Hardy-Weinberg equilibrium was determined by comparing the genotype frequencies with the expected values using a contingency table X2 statistic with Yates correction. Statistical analyses were performed using the STATISTICA 4.3 (Stat- Soft,
Results
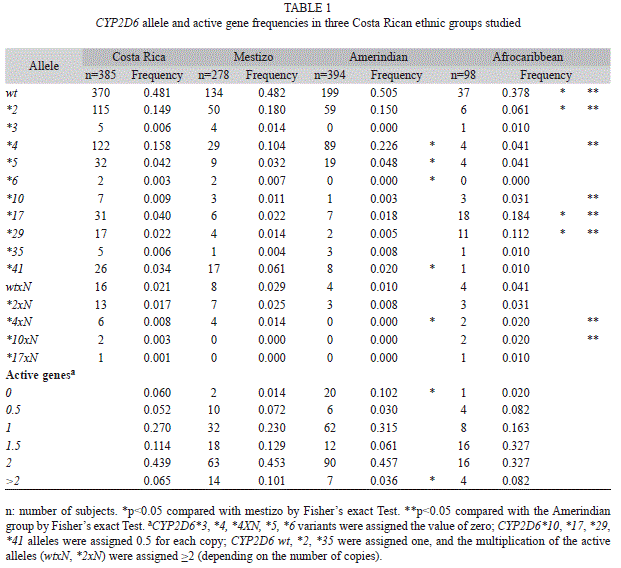
CYP2D6 allele frequencies are given in table 1. Multiplications of active genes (wtxN, *2xN) were present in 15 individuals of the mestizo population and in seven of each of the Amerindian and Afro-Caribbean populations (Table 1).
Null activity alleles *4 and *5 frequencies in the Amerindian population were higher (22.6% and 4.8%, respectively) than in the mestizo group (10.4% and 3.2%, respectively; p<0.05) (Table 1). The frequencies of alleles with decreased activity (*17 and *29) in the Afro-Caribbean population were higher (18.4% and 11.2%) than in the other two populations (p<0.05), and the CYP2D6*10 frequency in the Amerindian population (0.3%) was lower than in the Afro-Caribbean (3.1%; p<0.05).
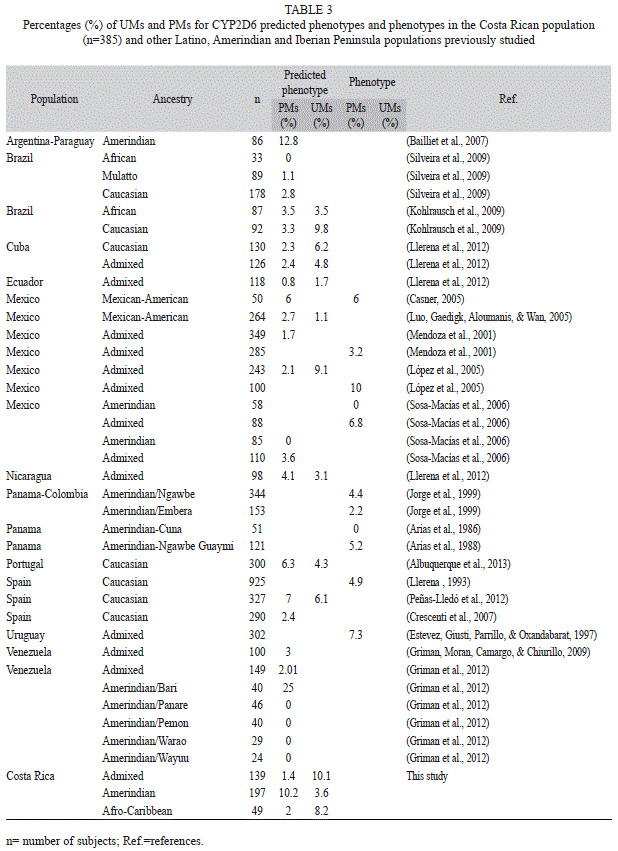
The CYP2D6 frequencies for each activity score group are given in table 1. The entire Costa Rican population frequency of PMs and UMs were 6% and 6.5% respectively.
As expected, the percentage of UMs in the mestizo population (10.1%) was higher than in the Amerindian population (3.6%, p<0.05) (Table 1). However, the frequency of individuals classified as PMs (zero active genes) was higher in Amerindians (10.2%) than in the mestizo population (1.4%, p<0.05). The frequency
of UMs (8.2%) and PMs (2%) of the Afro-Caribbean population was not different to any of the Costa Rican populations studied.
The frequencies of CYP2D6 genotypes are listed in table 2. The mestizo population showed more diversity concerning genotypes in comparison with the other Costa Rican populations studied. In all three groups, the most frequently found CYP2D6 genotypes belonged to the classification of two active genes (Table
2). Published data from Latin American populations is used to compare results with the Costa Rican populations (Table 3).
Discussion
To the best of our knowledge, this is the first study in a Costa Rican population that examined the predicted metabolic phenotype frequencies of CYP2D6 (UMs and PMs) in three ethnic groups. The entire Costa Rican population frequency of PMs (6%) is consistent with the Portuguese (Albuquerque et al., 2013), the Mexican-American (Casner, 2005), and the Colombian mestizo (Isaza, Henao, López, & Cacabelos, 2000) populations. Likewise, the frequency of UMs for the Costa Rican population is similar to those reported for the Spanish population (6.1%) (Peñas-Lledó et al., 2012).
Considering the ethnicity of the analyzed populations, the frequency of UMs in the mestizo group (10.1%) is similar to those reported in a Spanish population (6.1%) (Peñas-Lledó et al., 2012), those of a Mexican admixed population (9.1%) (López, Guerrero, Jung-Cook, & Alonso, 2005), and it is lightly greater than the percentage of UMs determined with debrisoquine in a Spanish population (5.2%, p=0.053) (Llerena et al., 2009). Moreover, the high frequency of PMs in the Costa Rican Amerindian population (10.2%) is similar to that reported in an Amerindian population from
Despite the small number of individuals in the Afro-Caribbean group, the frequencies of PMs and UMs are comparable to those of Brazilian populations with African ancestry (Kohlrausch et al., 2009; Silveira, Canalle, Scrideli, Queiroz, & Tone, 2009).
Individuals carrying two inactive alleles will produce a non-functional protein. Therefore, all of them will likewise be determined as PMs in phenotyping studies. However, discordance between the identification of Ums by molecular methods and phenotype has been reported (Llerena et al., 2012; Løvlie, Daly, Molven, Idle, & Steen, 1996), so the predicted phenotype estimation needs to be confirmed with phenotyping studies.
Variability of CYP2D6 alleles was found within this Costa Rican population, in accordance with the finding that the Costa Rican population is genetically heterogeneous (Morera et al., 2003; Morera & Barrantes, 2004). The high frequency of PMs in the Amerindian group can mainly be accounted for by the presence of the null allele CYP2D6*4 in this population (22.6%) at a frequency similar to those found in Amerindian populations of Argentina-Paraguay
(17.8%) (Bailliet et al., 2007). However, it is higher than those reported in Panamanian Embera (14%) and Ngwabe (17.1%) populations (p<0.05) (Jorge, Eichelbaum, Griese, Inaba, & Arias, 1999).
Regarding reduced activity alleles, the CYP2D6*17 and CYP2D6*29 frequencies in the Afro-Caribbean population were similar to those reported for an Afro-American population (Gaedigk, Bradford, Marcucci, & Leeder, 2002), in agreement with their African ancestry (
frequency in the Amerindian group (0.3%) was similar to that of the Tepehuano (0%) (Sosa- Macías et al., 2006, 2010) and Mapuche (1.8%) (Muñoz et al., 1998) populations, but lower than other Amerindian populations (7.1% and 6.9%; p<0.05) (Bailliet et al., 2007; Jorge et al., 1999).
A limitation of this study was that the Lumber of individuals in the Afro-Caribbean population was low (n=49). Moreover, the inclusion criteria did not include ancestry informative markers analysis (AIMs). In this sense, further studies involving genetic markers of ancestry are needed in the Costa Rican population. It is
necessary to remark that even though this study reports allele frequencies of CYP2D6 for Costa Ricans, they might not be representative of the population and might have been influenced by random effects.
Furthermore, the study of populations with complex ancestry such as Latin American populations comprises the fact that novel or rare variants (Fohner et al., 2013; Gaedigk et al., 2010) might appear, leading to poor metabolism or reduced function (Montané-Jaime et al., 2013). In the future, sequencing the CYP2D6
gene in these populations might be suitable to detect relevant genetic variants.
Given the percentage of UMs found in the mestizo and Afro-Caribbean population and of PMs in the Amerindian group, it might be appropriate to follow available guidelines that provide information relating to the interpretation of CYP2D6 genotype test results to guide the dosing of different drugs (Crews et al.,
2012; Hicks et al., 2013). Likewise, the information provided by this study supports that it might be appropriate to consider the development of drug treatment guidelines taking into account population ethnic background, meaning specific alleles of the population tested, to improve drug safety and efficacy in Costa Rican and Latin American populations.
In conclusion, we report here for the first time the frequency of PMs (6%) and Ums (6.5%) in a Costa Rican population. Secondly, we found a difference between the frequency of predicted UM and PM phenotype across ethnicity in Costa Ricans.
Acknowledgments
CCG was supported by a fellowship of the
The research was supported by Gobierno de Extremadura, Consejería de Empleo, Empresa e Innovación, and Fondo Social Europeo (FSE) fellowship PD10199 (MEGN) and a grant from AEXCID 13IA002. The project was coordinated in the CEIBA.FP Consortium of the Ibero-American Network of Pharmacogenetics
& Pharmacogenomics (RIBEF): Graciela E. Moya (Buenos Aires, Argentina), Eduardo Tarazona-Santos (Belo Horizonte, Brazil), Alba P. Sarmiento (Bogotá, Colombia), Ramiro Barrantes (San José, Costa Rica), Idania Rodeiro, Luis R. Calzadilla (La Habana, Cuba), Enrique Terán (Quito, Ecuador), Rocío Ortiz-López (Nuevo León, México), Marisol López-López (Mexico City, Mexico), Martha G. Sosa-Macías (Durango, Mexico), Ronald Ramírez-Roa (León, Nicaragua), Manuela Grazina (Coimbra, Portugal), Adrián LLerena (Badajoz, Spain), Francisco E. Estévez-Carrizo (Montevideo, Uruguay).
References
Albuquerque, J., Ribeiro, C., Naranjo, M. E., Llerena, A., Grazina, M., & CEIBA.FP Consortium. (2013). Characterization of CYP2D6 genotypes and metabolic profiles in the Portuguese population: pharmacogenetic implications. Personalized Medicine, 10(7), 709-718. [ Links ]
Arias, T. D., Jorge, L., Lee, D., Guerrero de Solís, N., Escobar, J., Barrantes, R., & Inaba, T. (1986). Biotransformation of sparteine in the Cuna American Indians of Panama. Revista médica de Panamá, 11(3), 199-209. [ Links ]
Arias, T. D., Jorge, L. F., Lee, D., Barrantes, R., & Inaba, T. (1988). The oxidative metabolism of sparteine in the Cuna Amerindians of Panama: absence of evidence for deficient metabolizers. Clinical Pharmacology and Therapeutics, 43(4), 456-465. [ Links ]
Azofeifa, J., Hahn, M., Ruiz, E., Hummerich, L., Morales, A. I., Jiménez, G., & Barrantes, R. (2004). The STR polymorphism (AAAAT)n within the intron 1 of the tumor protein 53 (TP53) locus in 17 populations of different ethnic groups of Africa,
Bailliet, G., Santos, M. R., Alfaro, E. L., Dipierri, J. E., Demarchi, D. A., Carnese, F. R., & Bianchi, N. O. (2007). Allele and genotype frequencies of metabolic genes in Native Americans from
Barrantes, R., Smouse, P. E., Neel, J. V., Mohrenweiser, H. W., & Gershowitz, H. (1982). Migration and genetic infrastructure of the Central American Guaymi and their affinities with other tribal groups. American Journal of Physical Anthropology, 58(2), 201-214. [ Links ]
Barrantes, R. (1989). Biodemografía de las poblaciones amerindias de Costa Rica. Vínculos, 15, 59-68. [ Links ]
Barrantes,R., Smouse, P. E., Mohrenweiser, H. W., Gershowitz, H., Azofeifa,J., Arias, T. D., & Neel, J. V. (1990). Microevolution in lower Central America: genetic characterization of the Chibcha-speaking groups of Costa Rica and country-region Panama, and a consensus taxonomy based on genetic and linguistic affinity. American Journal of Human Genetics, 46(1), 63-84. [ Links ]
Barrantes, R. (1993a). Diversidad genética y mezcla racial en los amerindios de Costa Rica y Panamá. Revista de Biología Tropical, 41(3), 379-384. [ Links ]
Barrantes, R. (1993b). Estructura de poblaciones. In Evolución en el trópico: los amerindios de Costa Rica y Panamá (pp. 51-85). San José, Costa Rica: Editorial de la Universidad de Costa Rica. [ Links ]
Bieber, H., Bieber, S. W., Rodewald, A., & Barrantes, R. (1996). Microevolution and genetic affinities among six Amerindian tribes of lower
Bradford, L. D. (2002). CYP2D6 allele frequency in European Caucasians, Asians, Africans and their descendants. Pharmacogenomics, 3(2), 229-243. [ Links ]
Casner, P. R. (2005). The effect of CYP2D6 polymorphisms on dextromethorphan metabolism in Mexican Americans. Journal of Clinical Pharmacology, 45(11), 1230-1235. [ Links ]
Crescenti, A., Mas, S., Gassó, P., Baiget, M., Bernardo, M., & Lafuente, A. (2007). Simultaneous genotyping of CYP2D6*3, *4, *5 and *6 polymorphisms in a Spanish population through multiplex long polymerase chain reaction and minisequencing multiplex single base extension analysis. Clinical and Experimental Pharmacology & Physiology, 34(10), 992-997. [ Links ]
Crews, K. R., Gaedigk, A., Dunnenberger, H. M., Klein, T. E., Shen, D. D., Callaghan, J. T., Kharasch, E. D., & Skaar, T. C. (2012). Clinical Pharmacogenetics Implementation Consortium. Clinical Pharmacogenetics Implementation Consortium (CPIC) guidelines for codeine therapy in the context of cytochrome P450 2D6 (CYP2D6) genotype. Clinical Pharmacology and Therapeutics, 91(2), 321-326. [ Links ]
CYP Alleles Nomenclature Database. (2013). The Human Cytochrome P450 (CYP) Allele Nomenclature Database. Retrieved from http://www.cypalleles.ki.se/ cyp2d6.htm
1. Genetics Section, School of Biology, University of Costa Rica, 2060 San Pedro, San José, Costa Rica;
ccg004@gmail.com, gerardo.jimenez@ucr.ac.cr, ramiro.barrantes@ucr.ac.cr
2. CICAB Clinical Research Centre,
megleznaranjo@gmail.com, allerena@unex.es


{kind=link}
{kind=link}
{kind=link}
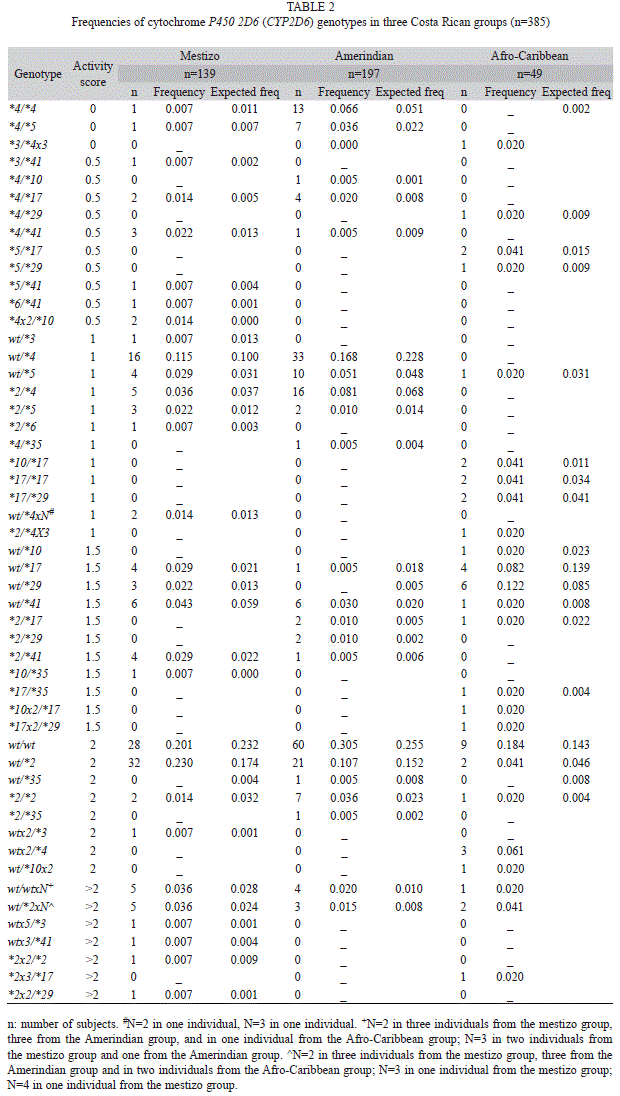{kind=link}
{kind=link}