










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.3 San José Jul./Sep. 2014
The morphology and bioactivity of the rice field cyanobacterium Leptolyngbya
Morfología y bioactividad de la cianobacteria Leptolyngbya en los campos de cultivo de arroz
Morfología y bioactividad de la cianobacteria Leptolyngbya en los campos de cultivo de arroz
Abstract
The genus Leptolyngbya comprises filamentous cyanobacteria that are important in rice fields. In the rhizosphere, cyanobacteria produce a variety of secondary metabolites such as auxins that are important in agriculture soil performance. To assess this, Leptolyngbya strain MMG-1, was isolated from the rhizosphere of rice plants and described. For this, the morphology of this strain was studied by light microscopy as well as by confocal laser scanning microscopy. Besides, the ability of this strain to synthesize an auxin-like bioactive compound was demonstrated under various culture conditions (different amounts of tryptophan; pH; different alternating light:dark periods; duration of the incubation). The auxin-like compound was extracted from the culture of Leptolyngbya strain MMG-1 and identified as indole-3-acetic acid (IAA) by thin layer chromatography (TLC) as well as by high performance liquid chromatography (HPLC). Our results showed that the strain required the precursor L-tryptophan for the synthesis of IAA. Leptolyngbya strain MMG-1 accumulated IAA intracellularly. The IAA secreted by Leptolyngbya strain MMG-1 was significantly correlated with the initial concentration of L-tryptophan in the medium, as well as with the duration of the incubation. The bioactivity of the secreted IAA was determined by its effect on the rooting pattern of Pisum sativum seedlings. The culture supernatant of Leptolyngbya strain MMG-1 stimulated the seedling lateral rooting, while it decreased root length. Hence, rhizospheric Leptolyngbya produced auxin under different conditions and affected the plants rooting pattern. Rev. Biol. Trop. 62 (3): 1251-1260. Epub 2014 September 01.
Key words: bioassay, Indole 3-Acetic Acid, Leptolyngbya, Pisum sativum, plant hormone.
Resumen
El género Leptolyngbya comprende cianobacterias filamentosas que son importantes en los campos de cultivo de arroz. En la rizosfera, las cianobacterias producen una variedad de metabolitos secundarios, tales como auxinas, que son importantes en el rendimiento de la agricultura del suelo. La cepa Leptolyngbya MMG-1, fue aislada de la rizosfera de plantas de arroz y se describe en este trabajo. La morfología de esta cepa se estudió por microscopía de luz, así como por microscopía confocal de barrido láser. Además, se estimó la capacidad de esta cepa para sintetizar el compuesto bioactivo auxina como se demostró en diversas condiciones de cultivo (diferentes cantidades de triptófano; pH; diferente luz alterna: períodos oscuros; duración de la incubación). La auxina se extrajo a partir del cultivo de la cepa Leptolyngbya MMG-1 y se identificó como ácido indol-3-acético (AA) por cromatografía de capa fina (TLC), así como por cromatografía líquida de alta resolución (HPLC). Nuestros resultados mostraron que la cepa requiere el precursor de L-triptófano para la síntesis de IAA. La cepa Leptolyngbya MMG-1 acumula intracelularmente IAA. El IAA secretada por la cepa Leptolyngbya MMG-1 se correlacionó significativamente con la concentración inicial de L-triptófano en el medio, así como con la duración de la incubación. La bioactividad de la IAA secretada se determinó por su efecto sobre el patrón de enraizamiento de plantas de semillero de Pisum sativum. El sobrenadante del cultivo de la cepa Leptolyngbya MMG-1 estimuló el enraizamiento lateral en la plántula, mientras que se redujo la longitud de la raíz. Por lo tanto, la producción de auxina por Leptolyngbya rizosférica afectó el crecimiento de las plantas.
Palabras clave: bioensayo, ácido indol-acético, Leptolyngbya, Pisum sativum, hormonas de plantas.
Cyanobacteria are important microorganisms in agricultural soils and especially in rice fields, where a large variety of them occur (Whitton, 2000). Cyanobacteria are oxygenic photoautotrophs and are known to produce a wide variety of secondary metabolites that may be important for a proper functioning of agricultural soils. In particular, filamentous cyanobacteria play multiple roles in the enhancement of soil fertility and thus in the improvement of crop plants. In the rhizosphere, cyanobacteria directly interact with plants through a variety of secondary metabolites. Phytohormones such as auxins are commonly used by rhizospheric microorganisms in order to interact with plants (Spaepen, Vanderleyden, & Remans, 2007). In response, the plants secrete a chemically rich mixture of organic and inorganic compounds in the form of root exudates that mainly serve to attract and assist the growth of microorganisms in rhizosphere (Lambers, Mougel, Jaillard, & Hinsinger, 2009). Rhizospheric microorganisms are efficient in detecting alteration of the environmental conditions that prevail in their surroundings. The efficiency of microorganisms to synthesize secondary metabolites such as auxins varies largely with the changes in environment (Bergman, Zheng, Klint, & Ran, 2008 ; Khalid, Arshad, & Zahir, 2006). Hence, rhizospheric microorganisms adapt optimally to the existing conditions in order to fully express their metabolic machinery (Bais, Weir, Perry, Gilroy, & Vivanco, 2006).
The genus Leptolyngbya comprises a variety of thin (<3μm) filamentous cyanobacteria, with their trichomes characterized by loose coils (Rippka & Herdman, 1992). Leptolyngbya species are common in a range of different environments such as marine, fresh water, soils and in archaeological sites (Wilmotte & Herdman, 2001). However, the taxonomic position of Leptolyngbya is controversial and the organisms belonging to it are difficult to identify (Bruno, Billi, Bellezza, & Albertano, 2009). In order to address this problem a more comprehensive approach is taken, known as a polyphasic approach, i.e. applying a combination of morphological, ultra-structural, molecular, and eco-physiological characteristics to identify the taxonomic status of ecologically relevant cyanobacteria (Castenholz & Norris, 2005). Leptolyngbya is abundantly present in rice paddies (Song, Martensson, Eriksson, Zheng, & Rasmussen, 2005). Species belonging to this genus are well-known for their ability to fix atmospheric nitrogen and this would thus enrich the soil with bound nitrogen.
This study was carried out in order to determine the morphology of Leptolyngbya and analyse its ability to synthesize auxins under certain cultivation conditions, and to test the effect of this cyanobacterial auxin on the development of roots of seedling.
Materials and Methods
Strain and growth conditions: Rhizospheric Leptolyngbya strain MMG-1 was isolated from roots of rice plants growing in the agricultural fields of the University of Punjab, Lahore, Pakistan (31°30’13” N - 74°17’47.05” E), in September 2006 (Ahmed, Stal, & Hasnain, 2010a). Axenic cultures of the strain (in triplicate) were photoautotrophically cultivated in BG-11 medium (Rippka & Herdman, 1992) for three weeks under constant culturing conditions with light (about 150µE/m2/s) provided by four 18W fluorescent lamps, under an alternating16:8 h light:dark cycle, and a temperature of 25-27°C without shaking (MLR-351H- Plant Growth Chamber, Sanyo North America Corp.) (Ahmed, Stal, & Hasnain, 2010a). Cultures were aseptically refreshed after every three weeks by transferring into freshly prepared BG-11 media.
Morphology: Leptolyngbya strain MMG-1was observed under a light microscope and by confocal laser scanning microscope (CLSM) following methods described previously (Ahmed, Stal, & Hasnain, 2010b). Two week-old cultures were dispersed in warm (45°C) liquid agarose (1% w/v) on an object glass, and after solidifying covered with a cover slip and observed with a light microscope (Zeiss Axiophot, Oberkochen, Germany) using differential interference contrast (DIC). The CLSM was a TCS-NT microscope (Leica, Heidelberg, Germany) equipped with an Argon-Krypton laser. Auto-fluorescence from cyanobacterial chlorophyll-a and phycobiliproteins was used for observation. The samples were excited by 488nm and emitted light was passed through a 590nm long-pass filter.
Chlorophyll-a determination: Chlorophyll-a was used to follow growth. It was extracted and determined according to Tandeau de Marsac and Houmard (1988). The cyanobacteria were harvested by centrifugation at 10 000g for 10 min at 4°C. The pellet was extracted with 80% methanol for 2h in the dark at 4°C. The extract was centrifuged (10 000g for 10min) and the absorbance of the supernatant was measured spectrophotometrically at 665nm against 80% methanol. The concentration of chlorophyll in the extract (µg/mL) was calculated as (OD665nm) ×13.9.
Biosynthesis of auxin-like compounds: Auxin biosynthesis was measured by using the Salkowski colorimetric method (Glickmann & Dessaux, 1995; Ahmed et al., 2010b).
Effect of the duration of the incubation and the initial concentration of L-Tryptophan on auxin synthesis: Auxin from the cultures was estimated after different incubation times as described previously with modification of medium (BG-11 medium instead of SAG medium) (Ahmed et al., 2010b). Six flaks containing 100mL of BG-11 medium supplemented with L-tryptophan were inoculated. A 7th flask did not receive an inoculum and was used as control. Every week one flask was harvested for the estimation of chlorophyll-a and for the production of auxin. The experiment was done using varying initial concentrations of L-tryptophan (250, 500, 750, 1 000, 1 250 and 1 500µg/mL). Each experiment was done in triplicate.
Effect of pH and light regime on auxin secretion: The effect of varying pH (5, 6, 7, 8 and 9) and light (16:8 and 8:16 h light:dark; continuous light and continuous dark) on the synthesis of auxin was studied in two separate experiments (Ahmed et al., 2010b). The Leptolyngbya strain MMG-1 was incubated for three weeks in BG-11 medium supplemented with 500µg/mL L-tryptophan under the above mentioned conditions. Each experiment was done in triplicate.
Extraction of released and cellular auxin: Released and intracellular auxin were extracted from the culture supernatant and the cyanobacterial cells, respectively (Ahmed et al., 2010b). All extracts were stored at -18°C and assayed within 48h.
Chromatographic analysis: Thin layer chromatography (TLC) of extracted IAA was done according to Gravel, Antoun, and Tweddell (2007). Whereas procedure of de-Bashan, Antoun, and Bashan (2008) was used for high performance liquid chromatography (HPLC) of extracted IAA.
Rooting assay: The auxin-like bioactivity from Leptolyngbya strain MMG-1 was demonstrated as the effect of the culture supernatant on the root growth of pea (P. sativum var. Climax) (Ahmed et al., 2010b). Certified pea seeds were surface sterilized and placed in Petri dishes containing two layers of Whatman filter paper. The filter sterilized supernatant of three-week-old cultures was added (1, 2.5 and 5mL) to the Petri dishes with the pea seeds and MilliQ water was added to make a total volume of 10mL. The seeds were germinated in the dark and then shifted to the growth chamber at a 16:8h light:dark (200µE/m2/s) regime. The number of roots and the root length of the seedlings were measured after 10 days of growth. The same procedure was applied for the filter-sterilized supernatants of cultures of the cyanobacterium of different age (1-6 weeks). IAA in the culture supernatants was measured colorimetrically with the Salkawski reagent. Each experiment was done in triplicate.
Data were statistically analyzed using the SPSS statistical package (version 20, IBM SPSS Inc, Chicago). Students t-test (Two tailed) was used to determine significant differences between the two groups and analysis of variance (ANOVA) was performed. The means were separated using Duncan’s multiple range test (p=0.05).
Results
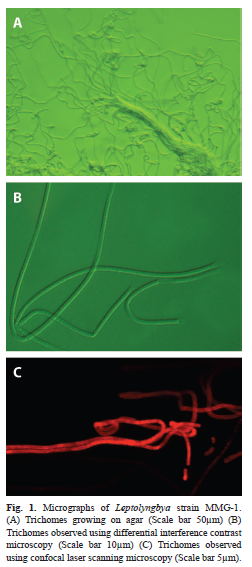
Strain characteristics: The trichomes of Leptolyngbya strain MMG-1 showed curly growth on BG-11 medium solidified with agarose (Fig. 1A). The width of the green trichomes was 1-1.5µm. A thick sheath surrounded the trichomes (Fig. 1B). Confocal laser scanning microscopy revealed the single cells of which the trichomes were composed (Fig. 1C).
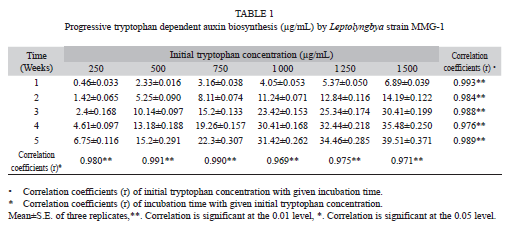
Impact of the duration of the incubation and the initial tryptophan concentration on auxin biosynthesis: The production of IAA by strain MMG-1 depended significantly on the concentration of tryptophan. The concentration of IAA in the supernatant gradually increased with time as well as with the supplemented amount of L-tryptophan in the medium, supported by highly significant correlation values in both cases (Table 1). The concentration of auxin in the medium reached 39µg/mL after five weeks and with 1 500µg/mL of tryptophan.
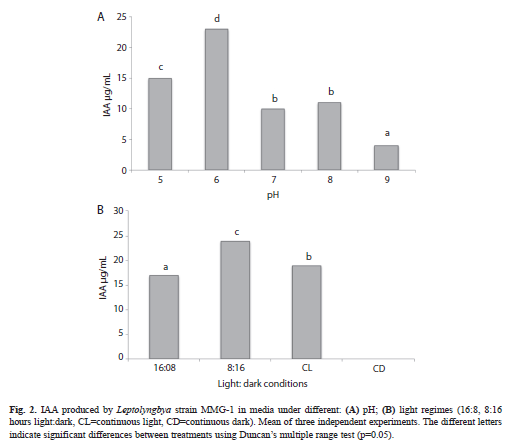
Effect of light and pH on IAA synthesis: A low pH significantly affected the synthesis of IAA by the strain MMG-1. The highest amount of IAA was found at pH 6 (23µg/mL) (Fig. 2A). The production of IAA was significantly higher (24µg/mL) under an alternating 8:16h light:dark regime than in cultures grown under other tested light regimes. Under continuous darkness no IAA was detected due to absence of growth (Fig. 2B).
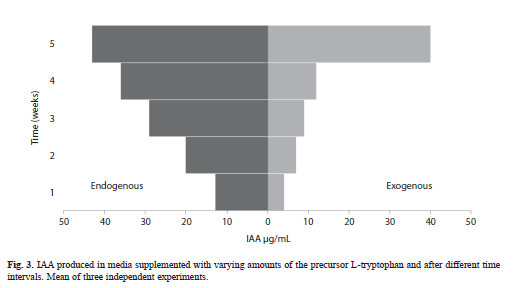
Cellular and exogenous IAA production: The production of exogenous and cellular IAA by strain MMG-1 was significantly different during any growth phase (t test, p=0.021). Although auxin was continuously released in the medium, the cyanobacterium accumulated IAA intracellularly (Fig. 3). Auxin accumulated intracellularly until five weeks of growth, when it reached 43µg/mL. Auxin that was released into the medium increased suddenly after five weeks of growth (233% as compared to week four).
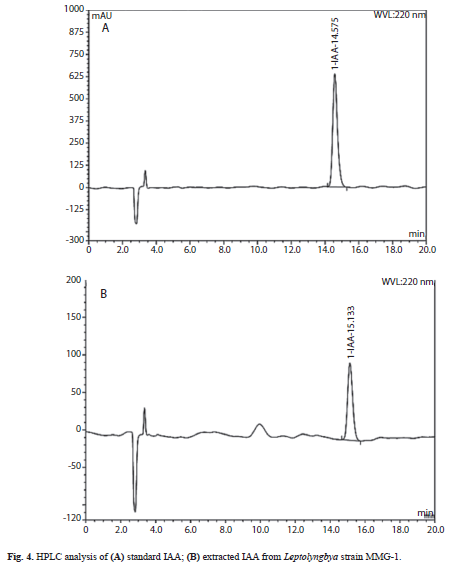
HPLC-TLC: Extracts of culture supernatant were analyzed by HPLC and thin layer chromatography (TLC) in order to confirm the presence of IAA. The TLC analysis gave the same Rf value (0.55-0.60) as the standard IAA. The retention time of the peak from the culture medium extract was similar to that of standard IAA when analysed by HPLC (Fig 4A y Fig. 4B).
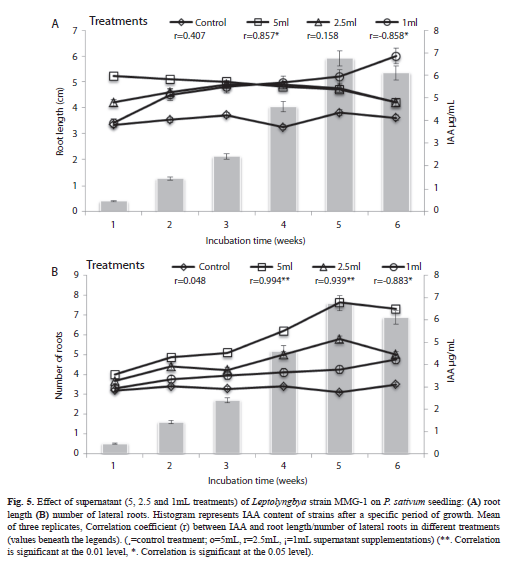
Effect of cyanobacteria culture medium on the number and length of lateral roots: The supernatant of strain MMG-1in less quantity resulted in an increased root length. However, the effect was more pronounced in supernatant with a low IAA concentration. The root length decreased with increased IAA in the medium as well as with the duration of the incubation (Fig. 5A). Supplementing 5mL of supernatant resulted in an increase of root length after one and two weeks, whereas1mL supernatant increased the root length after four, five and six weeks (53, 37 and 66%, respectively). The intensity of seedling rooting was positively affected by the addition of cyanobacterial culture supernatant with an optimum of 5mL. Strain MMG-1 supernatant enhanced the number of lateral roots by 146% after five weeks of incubation (Fig. 5B).
Discussion
Since the discovery of the rhizosphere microflora it became clear that these microorganisms play a vital role in growth and development of plants (Ortiz-Castro, Contreras-Cornejo, Macias-Rodriguez, & Lopez-Bucio, 2009). Cyanobacteria are an important group of microorganisms that modulate the growth of higher plants through a variety of secondary metabolites. Cyanobacteria physically and chemically interact with plants, for instance through the production of phytohormones.
The cyanobacterium Leptolyngbya strain MMG-1 was isolated from a rice paddy and produced the phytohormone auxin. TLC and HPLC analyses confirmed that IAA was produced. The Salkawski reagent is an easy, reliable and quick assay to detect and quantify IAA-like substances (Glickmann & Dessaux, 1995; Sergeeva, Liaimer, & Bergman, 2002). The most common pathway for IAA biosynthesis in plants and microorganisms is the indole-3-pyruvic acid pathway, which has L-tryptophan as an intermediate. This pathway has been described in many bacteria, including cyanobacteria, and the key enzyme (indole-3-pyruvate decarboxylase) of this pathway is encoded by ipdC (Sergeeva et al., 2002). This gene has been reported in many unicellular and filamentous cyanobacteria. Strain MMG-1 was unable to produce auxin in the absence of L-tryptophan. The amount of IAA that was produced depended on the initial amount of L-tryptophan supplied. The tryptophan to auxin conversion rate was independent of the supplemented tryptophan in the medium. The maximum conversion of 3.14% was obtained after five weeks of incubation and at an initial tryptophan concentration of 1 000µg/mL. Tryptophan has been detected inroot exudates of many plants. In Avena barbata tryptophan was exudated mainly near the root tips (Jaeger, Lindow, Miller, Clark, & Firestone, 1999). Tryptophan in the rhizosphere acts as a signal and as IAA precursor for the auxin-producing soil microorganisms (Hartmann, Schmid, van Tuinen, & Berg, 2009).
In the case of cyanobacteria, abiotic factors such as pH and light regime are crucial for the synthesis of IAA (Ahmed et al., 2010b). The production of larger quantities of IAA at lower pH may be attributed to the natural ability of microbes to efficiently express the IAA synthesis in rhizospheric conditions (Yuan, Liu, Saenkham, Kerr, & Nester, 2008). The light-dark regime of 8-16 hours gave the highest synthesis of IAA. This may be due to fact that IAA is sensitive to light and it may be degraded at light-dark regimes with longer light periods (Ahmed et al., 2010b).
Most important property of auxin is that they are effective on plant roots in minute concentrations. This property has been used to develop bioassays for the presence and activity of auxins (Barazani & Friedman, 1999). Sterile filtered culture supernatant affected the roots of P. sativum seedling. IAA resulted in a decrease of the root length, whereas it significantly enhanced the intensity of adventitious rooting in a time and concentration dependent manner (Ahmed et al., 2010b). Exploration of the mechanisms and conditions through which cyanobacteria maximize their secondary metabolites production should therefore allow their exploitation in biotechnology.
Acknowledgments
The Higher Education Commission of Pakistan is acknowledged for funding the visit of Mehboob Ahmed to the Netherlands Institute of Ecology (NIOO-KNAW) (IRSIP No.1-8 /HEC/HRD/2007/923). This is part of the PhD thesis of MA.
References
Ahmed, M., Stal, L. J., & Hasnain, S. (2010a). Association of non-heterocystous cyanobacteria with crop plants. Plant and Soil, 336(1-2), 363-375. [ Links ]
Ahmed, M., Stal, L. J., & Hasnain, S. (2010b). Production of indole-3-acetic acid by the cyanobacterium Arthrospira platensis strain MMG-9. Journal of Microbiology and Biotechnology, 20(9), 1259-1265. [ Links ]
Bais, H. P., Weir, T. L., Perry, L. G., Gilroy, S., & Vivanco, J. M. (2006). The role of root exudates in rhizosphere interactions with plants and other organisms. Annual Review of Plant Biology, 57, 233-266. [ Links ]
Barazani, O. & Friedman, J. (1999). Is IAA the major root growth factor secreted from plant-growth-mediating bacteria? Journal of Chemical Ecology, 25(10), 2397-2406. [ Links ]
Bergman, B., Zheng, W. W., Klint, J., & Ran, L. (2008). On the origin of plants and relations to contemporary cyanobacterial-plant symbioses. Plant Biotechnology, 25(3), 213-220. [ Links ]
Bruno, L., Billi, D., Bellezza, S., & Albertano, P. (2009). Cytomorphological and genetic characterization of troglobitic Leptolyngbya strains isolated from Roman hypogea. Applied and Environmental Microbiology, 75(3), 608-617. [ Links ]
Castenholz, R. W. & Norris, T. B. (2005). Revisionary concepts of species in the cyanobacteria and their applications. Algological Studies, 117(1), 53-69. [ Links ]
de-Bashan, L. E., Antoun, H., & Bashan, Y. (2008). Involvement of indole-3-acetic acid produced by the growth-promoting bacterium Azospirillum spp. in promoting growth of Chlorella vulgaris. Journal of Phycology, 44(4), 938-947. [ Links ]
Glickmann, E., & Dessaux, Y. (1995). A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Applied and Environmental Microbiology, 61(2), 793-796. [ Links ]
Gravel, V., Antoun, H., & Tweddell, R. J. (2007). Effect of indole-acetic acid (IAA) on the development of symptoms caused by Pythium ultimum on tomato plants. European Journal of Plant Pathology, 119(4), 457-462. [ Links ]
Hartmann, A., Schmid, M., van Tuinen, D., & Berg, G. (2009). Plant-driven selection of microbes. Plant and Soil, 321(1-2), 235-257. [ Links ]
Jaeger, C. H., Lindow, S. E., Miller, S., Clark, E., & Firestone, M. K. (1999). Mapping of sugar and amino acid availability in soil around roots with bacterial sensors of sucrose and tryptophan. Applied and Environmental Microbiology, 65(6), 2685-2690. [ Links ]
Khalid, A., Arshad, M., & Zahir, Z. A. (2006). Phytohormones: microbial production and applications. In N. Uphoff, A. S. Ball, E. Fernandes, H. Herren, O. Husson, M. Laing, C. Palm, J. Pretty, P. Sanchez, N. Sanginga, & J. Thies (Eds.), Biological Approaches to Sustainable Soil Systems (pp. 207-220). Boca Raton, Florida: Taylor & Francis/CRC. [ Links ]
Lambers, H., Mougel, C., Jaillard, B., & Hinsinger, P. (2009). Plant-microbe-soil interactions in the rhizosphere: an evolutionary perspective. Plant and Soil, 321(1), 83-115. [ Links ]
Ortiz-Castro, R., Contreras-Cornejo, H. A., Macias-Rodriguez, L., & Lopez-Bucio, J. (2009). The role of microbial signals in plant growth and development. Plant Signaling & Behavior, 4(8), 701-712. [ Links ]
Rippka, R. & Herdman, M. (1992). Pasteur Culture Collection of Cyanobacteria: Catalogue and Taxonomic Handbook. I. Catalogue of Strains, Institut Pasteur, Paris. [ Links ]
Sergeeva, E., Liaimer, A., & Bergman, B. (2002). Evidence for production of the phytohormone indole-3-acetic acid by cyanobacteria. Planta, 215(2), 229-238. [ Links ]
Song, T. Y., Martensson, L., Eriksson, T., Zheng, W. W., & Rasmussen, U. (2005). Biodiversity and seasonal variation of the cyanobacterial assemblage in a rice paddy field in Fujian, China. FEMS Microbiology Ecology, 54(1), 131-140. [ Links ]
Spaepen, S., Vanderleyden, J., & Remans, R. (2007). Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiology Reviews, 31(4), 425-448. [ Links ]
Tandeau de Marsac, N. & Houmard, J. (1988). Complementary chromatic adaptation: physiological conditions and action spectra. Methods in Enzymology, 167, 318-328. [ Links ]
Whitton, B. A. (2000). Soils and rice-fields. In B. A. Whitton & M. Potts (Eds.), Ecology of Cyanobacteria: Their Diversity in Time and Space (pp. 233-255). Dordrecht, The Netherlands: Kluwer Academic Publishers. [ Links ]
Wilmotte, A. & Herdman, M. (2001). Phylogenetic relationships among the cyanobacteria based on 16S rRNA sequences. In D. R. Boone, R. W. Castenholz, & G. M. Garrity (Eds.), Bergey’s Manual of Systematic Bacteriology (Vol. 1, pp. 487-599). New York: Springer. [ Links ]
Yuan, Z. C., Liu, P., Saenkham, P., Kerr, K., & Nester, E. W. (2008). Transcriptome profiling and functional analysis of Agrobacterium tumefaciens reveals a general conserved response to acidic conditions (pH 5.5) and a complex acid-mediated signaling involved in Agrobacterium-plant interactions. Journal of Bacteriology, 190(2), 494-507. [ Links ]
Ahmed, M., Stal, L. J., & Hasnain, S. (2010b). Production of indole-3-acetic acid by the cyanobacterium Arthrospira platensis strain MMG-9. Journal of Microbiology and Biotechnology, 20(9), 1259-1265. [ Links ]
Bais, H. P., Weir, T. L., Perry, L. G., Gilroy, S., & Vivanco, J. M. (2006). The role of root exudates in rhizosphere interactions with plants and other organisms. Annual Review of Plant Biology, 57, 233-266. [ Links ]
Barazani, O. & Friedman, J. (1999). Is IAA the major root growth factor secreted from plant-growth-mediating bacteria? Journal of Chemical Ecology, 25(10), 2397-2406. [ Links ]
Bergman, B., Zheng, W. W., Klint, J., & Ran, L. (2008). On the origin of plants and relations to contemporary cyanobacterial-plant symbioses. Plant Biotechnology, 25(3), 213-220. [ Links ]
Bruno, L., Billi, D., Bellezza, S., & Albertano, P. (2009). Cytomorphological and genetic characterization of troglobitic Leptolyngbya strains isolated from Roman hypogea. Applied and Environmental Microbiology, 75(3), 608-617. [ Links ]
Castenholz, R. W. & Norris, T. B. (2005). Revisionary concepts of species in the cyanobacteria and their applications. Algological Studies, 117(1), 53-69. [ Links ]
de-Bashan, L. E., Antoun, H., & Bashan, Y. (2008). Involvement of indole-3-acetic acid produced by the growth-promoting bacterium Azospirillum spp. in promoting growth of Chlorella vulgaris. Journal of Phycology, 44(4), 938-947. [ Links ]
Glickmann, E., & Dessaux, Y. (1995). A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Applied and Environmental Microbiology, 61(2), 793-796. [ Links ]
Gravel, V., Antoun, H., & Tweddell, R. J. (2007). Effect of indole-acetic acid (IAA) on the development of symptoms caused by Pythium ultimum on tomato plants. European Journal of Plant Pathology, 119(4), 457-462. [ Links ]
Hartmann, A., Schmid, M., van Tuinen, D., & Berg, G. (2009). Plant-driven selection of microbes. Plant and Soil, 321(1-2), 235-257. [ Links ]
Jaeger, C. H., Lindow, S. E., Miller, S., Clark, E., & Firestone, M. K. (1999). Mapping of sugar and amino acid availability in soil around roots with bacterial sensors of sucrose and tryptophan. Applied and Environmental Microbiology, 65(6), 2685-2690. [ Links ]
Khalid, A., Arshad, M., & Zahir, Z. A. (2006). Phytohormones: microbial production and applications. In N. Uphoff, A. S. Ball, E. Fernandes, H. Herren, O. Husson, M. Laing, C. Palm, J. Pretty, P. Sanchez, N. Sanginga, & J. Thies (Eds.), Biological Approaches to Sustainable Soil Systems (pp. 207-220). Boca Raton, Florida: Taylor & Francis/CRC. [ Links ]
Lambers, H., Mougel, C., Jaillard, B., & Hinsinger, P. (2009). Plant-microbe-soil interactions in the rhizosphere: an evolutionary perspective. Plant and Soil, 321(1), 83-115. [ Links ]
Ortiz-Castro, R., Contreras-Cornejo, H. A., Macias-Rodriguez, L., & Lopez-Bucio, J. (2009). The role of microbial signals in plant growth and development. Plant Signaling & Behavior, 4(8), 701-712. [ Links ]
Rippka, R. & Herdman, M. (1992). Pasteur Culture Collection of Cyanobacteria: Catalogue and Taxonomic Handbook. I. Catalogue of Strains, Institut Pasteur, Paris. [ Links ]
Sergeeva, E., Liaimer, A., & Bergman, B. (2002). Evidence for production of the phytohormone indole-3-acetic acid by cyanobacteria. Planta, 215(2), 229-238. [ Links ]
Song, T. Y., Martensson, L., Eriksson, T., Zheng, W. W., & Rasmussen, U. (2005). Biodiversity and seasonal variation of the cyanobacterial assemblage in a rice paddy field in Fujian, China. FEMS Microbiology Ecology, 54(1), 131-140. [ Links ]
Spaepen, S., Vanderleyden, J., & Remans, R. (2007). Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiology Reviews, 31(4), 425-448. [ Links ]
Tandeau de Marsac, N. & Houmard, J. (1988). Complementary chromatic adaptation: physiological conditions and action spectra. Methods in Enzymology, 167, 318-328. [ Links ]
Whitton, B. A. (2000). Soils and rice-fields. In B. A. Whitton & M. Potts (Eds.), Ecology of Cyanobacteria: Their Diversity in Time and Space (pp. 233-255). Dordrecht, The Netherlands: Kluwer Academic Publishers. [ Links ]
Wilmotte, A. & Herdman, M. (2001). Phylogenetic relationships among the cyanobacteria based on 16S rRNA sequences. In D. R. Boone, R. W. Castenholz, & G. M. Garrity (Eds.), Bergey’s Manual of Systematic Bacteriology (Vol. 1, pp. 487-599). New York: Springer. [ Links ]
Yuan, Z. C., Liu, P., Saenkham, P., Kerr, K., & Nester, E. W. (2008). Transcriptome profiling and functional analysis of Agrobacterium tumefaciens reveals a general conserved response to acidic conditions (pH 5.5) and a complex acid-mediated signaling involved in Agrobacterium-plant interactions. Journal of Bacteriology, 190(2), 494-507. [ Links ]
1. Department of Microbiology and Molecular Genetics, University of the Punjab, Lahore-54590, Pakistan;mehboob.mmg@pu.edu.pk
Department of Marine Microbiology, Royal Netherlands Institute for Sea Research (NIOZ), PO Box 140, 4400 AC Yerseke, The Netherlands.
2. Department of Marine Microbiology, Royal Netherlands Institute for Sea Research (NIOZ), PO Box 140, 4400 AC Yerseke, The Netherlands; lucas.stal@nioz.nl
Department of Aquatic Microbiology, IBED, University of Amsterdam, 1090 GE Amsterdam, The Netherlands.
3. Department of Microbiology and Molecular Genetics, University of the Punjab, Lahore-54590, Pakistan.
Women University Multan, Pakistan; genetic@brain.net.pk
Received 06-V-2013. Corrected 16-I-2014. Accepted 18-II-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}