










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.3 San José Jul./Sep. 2014
Microbial diversity and digestive enzyme activities in the gut of earthworms found in sawmill industries in Abeokuta, Nigeria
Diversidad microbiana y actividad de enzimas digestivas en el intestino de las lombrices de tierra en los aserraderos de Abeokuta, Nigeria
Diversidad microbiana y actividad de enzimas digestivas en el intestino de las lombrices de tierra en los aserraderos de Abeokuta, Nigeria
Abstract
The growing demand for wood has resulted in large volumes of wood wastes that are daily released to the soil from the activities of sawmills in South-Western Nigeria. In an attempt to setup a bioremediation model for sawdust, this study therefore aimed at evaluating microbial diversity, and the level of digestive enzymes in the gut of earthworms (Eudrilus eugeniae, Libyodrilus violaceous and Hyperiodrilus africanus) of sawmill origin. Four major sawmills located in Abeokuta (7o9’12” N - 3o19’35” E), namely Lafenwa, Sapon, Isale-Ake and Kotopo sawmills were used for this study. The arboretum of the Federal University of Agriculture, Abeokuta was used as control. Gut microbial analysis was carried out using the pour-plate method while digestive enzyme activities in the earthworm guts were done by the spectrophotometric method. Higher microbial counts (28.5±0.1x103-97.0±0.1x103cfu for bacteria and 7.0±0.1x103-96.0±0.1x103cfu for fungi) and microbial diversity were recorded in the gut of earthworms of the sawmill locations than those of the control site (17.5±0.1x103cfu for bacteria and 4.5±0.1x103cfu for fungi). Streptococcus mutans and Proteus spp.were common in the gut of E. eugeniae, and L. violaceous from the study sawmills, while Streptococcus mutans were also identified in H. africanus, but absent in the gut of E. eugeniae from the control site. Cellulase (48.67±0.02mg/g) and lipase (1.81±0.01mg/g) activities were significantly higher (p<0.05) in the gut of earthworms from the control site than those of the study sawmills. Furthermore, amylase (α and β) activity was highest in the gut of earthworms from the sawmills. Variations observed in the gut microbial and digestive enzyme activities of earthworms from the study sawmills as compared to the control site suggests that earthworms, especially E. eugeniae, could be a better organism for use as bioremediator of wood wastes. Rev. Biol. Trop. 62 (3): 1241-1249. Epub 2014 September 01.
Key words: earthworm, gut microflora, digestion, gut enzymes, sawdust, bioremediation.
Resumen
La creciente demanda de madera ha resultado en grandes volúmenes de residuos de madera que se lanzan diariamente a la tierra, procedentes de las actividades de los aserraderos en el suroeste de Nigeria. Se realizó este estudio en un intento de configurar un modelo de biorremediación de aserrín, con el objetivo de evaluar la diversidad microbiana y el nivel de las enzimas digestivas en el intestino de las lombrices de tierra (Eudrilus eugeniae, Libyodrilus violáceo y Hyperiodrilus africanus) de origen aserradero. Para este estudio se utilizaron cuatro grandes aserraderos ubicados en Abeokuta (7°9’12 ‘’ N - 3°19’35 ‘’ E), a saber: Lafenwa, Sapon, Isale-Ake y Aserraderos Kotopo. El arboreto de la Universidad Federal de Agricultura, Abeokuta se utilizó como control. El análisis microbiano se llevó a cabo utilizando el método de vertido de placa, mientras que las actividades de enzimas digestivas en los intestinos de las lombrices se realizaron por el método espectrofotométrico. Los recuentos microbianos más altos fueron 28.5±97.0±0.1x103-0.1x103cfu para las bacterias y 7.0±0.1x103-96.0±0.1x103cfu para los hongos y la diversidad microbiana registrada en el intestino de las lombrices de tierra de los lugares de aserraderos y las del sitio de control fueron: 17.5±0.1x103cfu para bacterias y 4.5±0.1x103cfu para hongos. Streptococcus mutans y Proteus spp. eran comunes en el intestino de E. eugeniae, y L. violaceous de los aserraderos de estudio, mientras que también se identificaron Streptococcus mutans en H. africanus, que estuvieron ausentes en el intestino de E. eugeniae en el sitio de control. Actividades de celulasa (48.67±0.02mg/g) y lipasa (1.81±0.01mg/g) fueron significativamente mayores (p<0.05) en el intestino de las lombrices de tierra en el sitio de control que las de los aserraderos de estudio. Por otra parte, la actividad de la amilasa (α y β) fue mayor en el intestino de las lombrices de tierra de los aserraderos. Las variaciones observadas en la microbiota intestinal y la actividad de las enzimas digestivas de las lombrices de tierra de los aserraderos de estudio, en comparación con el sitio de control, sugieren que las lombrices de tierra, especialmente E. eugeniae, podrían ser un mejor organismo para su uso como biorremediador de residuos de madera.
Palabras clave: lombriz de tierra, microflora intestinal, digestión, enzimas intestinales, aserrín, biorremediación.
Sawmilling is a major enterprise providing direct and indirect employment for thousands of people in the tropical rain forest region of Nigeria where there is abundance of trees (Ihekwaba, Nwafor, & Adienbo, 2009). Sawdust which is the major residue of sawmilling operations is a by-product of wood processing and is generally regarded as waste (Lennox, Abriba, Alabi, & Akubuenyi, 2010).
It is well established that earthworms (Class Oligochaetae; Phylum Annelida), play important roles in the terrestrial ecosystem (Debasmita, Mishra, & Mohapatra, 2011). Some of these roles include soil aeration (Olayinka et al., 2011; Owa, Dedeke, & Yeye, 2002), humus formation (Renu, Pandey, Bisht, Kandpal, & Kausha, 2006) and organic matter recycling (Satchell, 1967). It has also been reported that while earthworms use organic matter as their nutrient source, the microorganisms ingested along with these nutrient actually produce the enzymes that make the nutrients available for the worm’s use (Lee, 1985; Owa, Olowoparija, Aladesida, & Dedeke, 2013).
The ability of the microbes to survive the enteric condition of the gut of earthworms is very important (Owa et al., 2013). Hornor & Mitchell (1981) suggested that the ingested microbial populations play a key role in earthworm nutrition by helping in the breakdown of organic matter, particularly the components that the earthworms cannot utilize in their natural state.
The microbial activities in the gut and cast of earthworms was described by Devi et al. (2009) to be essential for the degradation of organic wastes which results in the release of nutrients to plants. Earthworms transform organic waste constituents into more useful forms by grinding and digesting with the aid of aerobic and anaerobic microflora (Maboeta & Van Rensberg, 2003). This biological decomposition of organic matter has been described to be mediated by a variety of biochemical processes in which enzymes play a key role (Garcia, Hernandez, Costa, Ceccanti, & Ciardi, 1992; Vuorinen, 1999).
Sawmills are a very common industry in the South-Western part of Nigeria. These industries release high volumes of sawdust which have cellulose, hemicelluloses and lignin as its major constituent. This sawdust forms the bulk of wastes released during sawmilling activities. The sawdust is usually burnt in piles in open air in different locations around the vicinities of sawmills (Dosunmu & Ajayi, 2002; Adelagun, Berezi, & Akintunde, 2012) in the dry seasons and consequently pollute the atmosphere. During the wet season, the sawdust produced are usually spread over the sawmill soils.
Earthworms are the most abundant and common soil fauna found around sawmills (Bamidele et al., unpublished). The activities of these earthworms were believed to be connected to their role in the degradation of sawdust as well as soil humidification and their pedobiological roles in the study sawmills. In order to ascertain these with the aim of setting up a bioremediation model for sawdust using earthworms, there is the need to compare the activities of digestive enzymes and microflora in the gut of earthworms from the vicinities of sawmills with those of a natural site, devoid of sawmilling activities. This study therefore aims at evaluating the effect of waste products of sawmill origin on the gut microflora and gut digestive enzymes of the earthworms from sawmill areas of Abeokuta, Southwestern Nigeria.
Materials and Methods
Experimental site: Four major sawmills located in Abeokuta, South-Western Nigeria namely Lafenwa (7o09’44” N - 3o19’35” E), Sapon (7o09’12” N - 3o20’49” E), Isale-Ake (7o09’48” N - 3o21’23” E) and Kotopo (7o11’05” N - 3o25’39” E) sawmills were selected and used for this study. They are very busy in activities and supply most of the processed wood and wood products used in Abeokuta and neighbouring towns. Earthworm samples were collected from each of the study sawmill respectively. Earthworms collected from the arboretum of the Federal University of Agriculture, Alabata, Abeokuta (7o 10’00” N - 3o 02’00” W) were used as control.
Earthworm sample collection: Earthworms were collected according to the method described by Owa et al. (2013). The soil was carefully turned using a spade while the earthworms were handpicked into containers and transported to the laboratory where they were washed with clean sterile water. The worms were kept under refrigeration for three to four hours in order to kill them without causing any harm or alteration to the microbial and digestive enzyme activities of the gut.
The earthworm species were identified by an earthworm taxonomist, Dr. Aladesida, A.A. of the Department of Biological Sciences, Federal University of Agriculture, Abeokuta. The earthworm species used were Libyorilus violaceous (Beddard, 1891), Hyperiodrilus africanus, (Beddard, 1890) and Eudrilus eugeniae, (Kinberg, 1866). The earthworm species varied according their location as follows:
• Control site: Eudrilus eugeniae, (Kinberg, 1866)
• Kotopo sawmill: Eudrilus eugeniae, (Kinberg, 1866)
• Sapon sawmill: Eudrilus eugeniae, (Kinberg, 1866)
• Lafenwa sawmill: Libyodrilus violaceous (Beddard, 1891)
• Isale Ake sawmill: Hyperiodrilus africanus, (Beddard, 1890)
The variation in the earthworm species used became inevitable because the distribution of earthworm species found in the different locations were not the same all through.
The earthworm specimens were reposited in the museum collection of the Biological Sciences Laboratory, Federal University of Agriculture, Abeokuta, Nigeria.
Dissection of earthworms: Each specimen to be dissected was washed in sterile distilled water and pinned down horizontally on sterilized dissecting board with the dorsal part downward. The ventral part was cut open longitudinally along the earthworm using sterilized dissecting kits (Owa et al., 2013). The guts were removed for microbial and digestive enzyme analyses of the gut content.
Gut microbial analysis: All glass wares and kits used were sterilized using aseptic method. The work bench and the used areas were sterilized with 90% ethanol. A spirit lamp was lit up to ensure that the air of the vicinity of the work bench was free of contaminants.
The media used were Nutrient agar (NA) for bacteria isolation while Potato Dextrose agar (PDA) was used for fungi isolation. A volume of 1mL of homogenized earthworm guts was dissolved in 9mL of sterile distilled water. This was then followed by serial dilution into 9mL of sterile distilled water into different dilution factors of 10-1, 10-2 and 10-3. A total of 1mL of each of these dilutions was subsequently inoculated into sterile plates after which the sterilized prepared media were poured into each of the plates. Bacteria plates were incubated at 37oC for 18 to 48 hours, while fungal plates were incubated at 27oC for five days. Colony forming units (cfu) were determined by visual counting (Ademolu & Idowu, 2011). The distinct colonies were sub-cultured to get a pure culture. The identification of the bacteria colonies was based on classification schemes of Harrigan & McCance (1976) and Sneath, Mair, Sharpe, & Holt (1986). The identification was based essentially on morphology and biochemical reactions. Fungi genus was determined through morphological criteria using identification keys such as the description of mycelia and of asexual reproduction forms (Domsch, Gams, & Anderson 1980). Further identification was carried out according to Kreger-venrij (1984) by pseudomycelium formation and pattern of sugar fermentation (Glucose, lactose, Sucrose, maltose and fructose).
Gut Digestive Enzymes Analysis
Cellulase activity: A total of 1g of frozen earthworm gut tissue was ground with 20mL of 1/15M dibasic sodium phosphate (K2HPO4) in a mortar maintained at 5oC with crushed ice and cellulose activity determined according to the method of Nokrans (1957).
Total amylase (α and β) activity: Enzyme extract was prepared by grinding 1g of the earthworm guts with 20mL 1/10M sodium acetate buffer pH 5.0 in a mortar maintained at 5oC with crushed ice and the buffer extract centrifuged at 18 000g for 30 minutes at 2oC. The α amylase activity was determined according to the method described by Wilson (1971) while β amylase activity was determined according to the method of Swain & Dekker (1966);
Lipase activity: Enzyme extracts were prepared in the same way as those of total amylase activity. Lipase activity in the extracts was determined using the method of Yong & Wood (1977).
Proteinase activity: Enzyme extracts were prepared in a manner similar to those of total amylase activity except that 20mL of 0.05M sodium phosphate buffer pH 6.0 were used as the extracting buffer. Proteinase activity in the enzyme extracts was determined using the method of McDonald & Chen (1965).
Data collected was subjected to statistical analyses which included descriptive statistics and Analysis of Variance (ANOVA) using the Statistical Package for Social Sciences (SPSS) version 16.0. Post Hoc test was done using LSD (Least Significant different), p-value was set at 0.05.
Results
Microbial Count: Microbial load count in the gut of the earthworm samples showed that the gut of E. eugeniae from sawmills had higher microbial count compared to those from the control site (Table 1).
The highest bacteria count in the gut of E. eugeniae was obtained from Sapon sawmill (54±0.1x103cfu) while it was lowest (17.5±0.1x103cfu) in those of the control site. Bacterial counts were significantly different (p<0.05) in the gut of E. eugeniae from Kotopo sawmill, Sapon sawmill and the control site (Table 1). A count of 82.5 x 103cfu was made in the gut of H. africanus from Isale-Ake sawmill, while L. violaceous from Lafenwa sawmill recorded 97±0.1x103cfu (Table 1).
Fungal count (96±0.1 x 103cfu) was significantly higher (p<0.05) in the gut of E. eugeniae from Kotopo sawmill than those of Sapon sawmill (10±0.1x103cfu) and control site (4.5±0.1x103cfu) (Table 1).
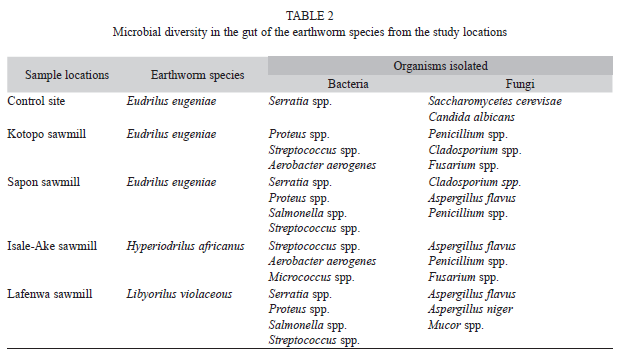
Microbial Diversity: A total of five bacteria species were present in the gut of E. eugeniae from the study locations (Table 2). Serratia spp., Proteus spp., Salmonellae spp. and Streptococcus spp. and Aerobacter aerogenes were identified in the gut of E. eugeniae from Kotopo and Sapon sawmills while only Serratia spp. was identified from those of the control site.
Streptococcus mutans and Proteus spp. was found to be common in the gut of E. eugeniae from the study sawmills but absent in those of the control site. Streptococcus mutans was also identified in H. africanus from Isale Ake sawmill while both Streptococcus mutans and Proteus spp. were identified in L. violaceous from Lafenwa sawmill.
Two genera of yeast (Candida albicans and Saccharomycetes cerevisae) were isolated from the gut of E. eugeniae from the control site. On the other hand, four genera of moulds (Aspergillus flavus, Penicillium spp., Fusarium spp., and Cladosporium spp.) were isolated from the gut of E. eugeniae from the study sawmills. Penicillium spp. and Cladosporium spp. were present in the gut of E. eugeniae from both Kotopo and Sapon sawmills. Aspergillus spp. was also discovered in the gut of H. africanus from Isale ake sawmill and L. violaceous from Lafenwa sawmill.
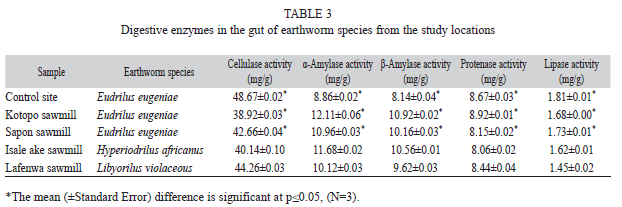
Gut digestive enzymes: The result of the gut digestive enzyme activities from the study locations is presented in table 3. Cellulase activity recorded the highest values of all the enzymes (38.92±0.03mg/g to 48.67±0.02mg/g) in the gut of the earthworms from the study locations while lipase activity had the lowest values (1.45±0.02mg/g to 1.81±0.01mg/g). Cellulase activity was highest in the gut of E. eugeniae from the control site than those of the study sawmills. L. violaceous from Lafenwa sawmill had cellulase activity of 44.26 ±0.03mg/g while 40.14 ±0.1mg/g was recorded for H. africanus from Isale Ake sawmill.
The values for α amylase in the gut of E. eugeniae followed the same trend with those of β amylase. The lowest values of 8.86±0.02mg/g and 8.14±0.04mg/g were recorded for α and β amylase in the control site respectively, while samples from Kotopo sawmill with the highest value of 12.11±0.06mg/g for α-amylase also recorded the highest value of 10.92±0.02mg/g for β-amylase.
Proteinase activity in the earthworm guts ranged between 8.06±0.02mg/g and 8.92±0.01mg/g. The highest lipase activity (1.81±0.01mg/g) was recorded in the gut of E. eugeniae from the control site and this follows the order: control>Sapon sawmill>Kotopo sawmill. Values obtained for each of the digestive enzymes were significantly different (p<0.05) (Table 3).
Discussion
The result of the microbial load count in the gut of the earthworms indicated more bacteria and fungal load in the gut of E. eugeniae from the study sawmills than those from the control site. Microbial load was also high in L. violaceous obtained from Lafenwa swmill and H. africanus from Isale Ake sawmill. Higher microbial load in the gut of earthworms from the study sawmills could be responsible for the utilization of sawdust as a source of carbon and energy in the gut of these earthworms. This corroborates the findings of Godliving (2002) that bacteria and fungi degrade wood dust. Dosoretz, Chenn, & Greth (1990) also reported the reduction in carbon content of sawdust when subjected to microbial degradation.
It was also observed that more bacteria and fungi species (diversity) were present in the gut of E. eugeniae from the study sawmills than those from the control site. Microbial diversity was high in L. violaceous obtained from Lafenwa swmill and H. africanus from Isale Ake sawmill. Sawmills by nature generate a lot of wastes-saw dusts, wood off cuts, wood backs, plain shavings, wood rejects, among others (Dosunmu & Ajayi, 2002). The more microflora diversity found in the gut of earthworms from the study sawmills could be needed to aid the digestion of wood particles as well as degrading other pollutants arising from wood processing activities. This increase in microbial diversity in the gut of the earthworms from the study sawmills might as well be due to the utilization of the abundant sawdust in the sawmills, which could serve as a cheap source of nutrient for the earthworms’ use.
Streptococcus mutans and Proteus spp. were present in the gut of E. eugeniae from the study sawmills but absent in the gut of those from the control site. These microbes were also present in the gut of L. violaceous obtained from Lafenwa sawmill while Streptococcus mutans was present in H. africanus from Isale Ake sawmill. Streptococcus mutans has been reported to be highly efficient in the degradation of cellulose in sawdust (Lennox et al., 2010). Dutkiewicz, Krysinska-Traczyk, Prazmo, Skorska, & Sitkowska (2001) also identified Streptococcus mutans in the air of Polish sawmills. There is the probability that Streptococcus mutans could be of great significance in the degradation/digestion of wood dust in the gut of the earthworms from the study sawmills. The mutualism of Streptococcus spp. with casts and gut of L. violaceous have been documented (Idowu, Edema, & Adeyi 2005; Dedeke, Omemu, Aladesida, & Museliu, 2010).
Parthasarathi, Ranganathan, Anandi, & Zeyer (2007) identified the presence of Proteus spp. in cowdung and pressmud, but absent in the gut of E. eugeniae reared on them. It could therefore be inferred that Proteus spp. was selectively ingested by these earthworms as supplement for the lingo-cellulotic digestion of wood from sawdust.
The gut of E. eugeniae from the study sawmills harbours a great fungal diversity, mainly moulds while on the other hand, only yeast was present in those from the control site. The presence of yeast isolates (Saccharomycetes cerevisae and Candida albicans) in the gut of E. eugeniae from the control site and their absence from the gut of those from the study sawmills could be an indication that yeast might be of little or no significance in the break down/digestion of wood particles from sawdust in the gut of the earthworms.
Aspergillus species, Fusarium spp. and Penicillium spp. were the common fungal genera in the gut of E. eugeniae from the study sawmills but were absent in those of the control site. These microorganisms were also present in the gut of H. africanus from Isale Ake sawmill while Aspergillus spp. was present in L. violaceous from Lafenwa sawmill. These micro-organisms have been associated with the degradation of cellulose in sawdust and wood shavings (Lennox et al., 2010; Hameed, Khoder, & Farag, 1999). Aspergillus species and Penicillium spp. have also been reported to be present in the air of Polish sawmills (Dutkiewicz et al., 2001) and in the gut of tropical earthworm Glyphodrillus tuberosus isolated from organic rice fields (Debasmita et al., 2011). Their presence might be in order to aid organic breakdown of wood and ligno-cellulotic materials in the soil of the study sawmills. This compares favourably with the report of Parthasarathi & Ranganathan (1998) that sawdust contains more cellulose, lignin and cellulotic enzyme producing microbes which are Aspergilli and Fusarium spp. as well as proteolytic enzyme producing microbes (Aspergillus niger and Aspergillus flavus).
Fairly well established that the gut of earthworm is a specialized microhabitat of enhanced microbial activities in soils (Karsten & Drake 1995; Idowu et al., 2005), the difference in microbial species (bacteria and fungi) isolated from the gut of earthworms from the control site and those of the study sawmills, in the present study, suggests that the microhabitats of the earthworm guts were carefully selected to suit their environment and also compensate for the digestion of the available substrate.
Cellulase activities was higher than α and β amylase, proteinase and lipase activities in the gut of earthworms from all the study locations. This is not unexpected as earthworms feed mainly on plants and plant materials and these have higher percentage of cellulose. Higher cellulase activities than other digestive enzymes in earthworms had been reported by Devi et al. (2009) for normal and vermicomposting earthworms. Zhang, Li., Shen, Wang, & Sun (2000) also recorded higher cellulase activity than protease and phosphatease in Eisenia fetida.
Cellulase activities was highest in the gut of E. eugeniae from the control site than those from the study sawmills. Enzyme activity, of the various factors is influenced by type of food (Parthasarathi & Ranganathan, 2000). The variation in cellulase activities recorded in the gut of E. eugeniae from the control site to those from the study sawmills could be associated with their ecological categories and feeding habitat (Zhang et al., 2000). The earthworms of the control site feed mainly on leaves and tender stem litters while those from the study sawmills feed mainly on sawdust particles incorporated in soils. Sawdust is made up of three major components; cellulose, hemicelluloses and lignin (Erikson, Blanchette, & Ander, 1990) lignin being the most recalcitrant and protects the cellulose and hemicellulose from enzymatic attack by some microorganisms (Bonnarme & Jeffries, 1998). The recalcitrant nature of lignin in sawdust which forms the bulk of organic substrate in the sawmill soils could as well have resulted in the low activities of cellulase recorded in the gut of E. eugeniae from the study sawmills as compared to those of the control site. Shi, Shi, Wang, Lu, & Yan (2007) reported a reduction in the cellulase activity in the gut of earthworms treated with deltametrine pesticides. Therefore, several other pollutants arising as a result of sawmilling activities could as well be responsible for the low cellulase activities in the gut of the earthworms from the study sawmills as compared to those from the control site.
Based on the variations observed in the microbial and digestive enzyme activities in the gut of earthworms from the study sawmills as compared to the control site, earthworms especially E. eugeniae could be a better organism for use as a bioremediator of wood wastes.
Acknowledgment
We wish to acknowledge Aladesida Adeyinka of the Department of Biological Sciences, Federal University of Agriculture, Abeokuta who freely identified the earthworm species used for this research work.
References
Adelagun, R. O., Berezi, E. P., & Akintunde, O. A. (2012). Air Pollution In A Sawmill Industry: The Okobaba (Ebute-meta, Lagos) Experience. Journal of Sustainable Developmental and Environmental Protection, 2(2), 29-36. [ Links ]
Ademolu, K. O. & Idowu, A. B. (2011). Occurrence and distribution of microflora in the gut regions of the Variegated grasshopper Zonocerus variegatus (Orthoptera: Pyrgomorphidae) during development. Zoological Studies, 50(4), 409-415. [ Links ]
Bonnarme, D. & Jeffries, T. W. (1998). Mn (11) regulation of lignin peroxidise and mangares-dependent peroxidise from lignin-degrading white-rot fungi. Applied Environmental Microbiology, 56, 210-217. [ Links ]
Debasmita, C., Mishra, C. S. K., & Mohapatra, P. K. (2011). Diversity of Bacteria and Fungi in the Gut of the Tropical Earthworm Glyphodrillus tuberosus Isolated from Conventional and Organic Rice Fields. Journal of Pharmacology and Toxicology, 6(3), 303-311. [ Links ]
Dedeke, G. A., Omemu, O., Aladesida, A. A., & Museliu, F. (2010). Comparative Microbial Analysis of Earthworm Casts Collected From Ikenne, Ogun State, Nigeria. Ethiopian Journal of Environmental Studies and Management, 3(3), 57-63. [ Links ]
Devi, S. H., Vijayalakshmi, K., Jyotsna, K. P., Shaheen, S. K., Jyothi, K., & Rani, M. S. (2009). Comparative assessment in enzyme activities and microbial populations during normal and vermicomposting. Journal of Environmental Biology, 30(6), 1013-1017. [ Links ]
Domsch, K. H., Gams, W., & Anderson, T. H. (1980). Compendium of soil fungi. London: Academic Press. [ Links ]
Dosoretz, G. G., Chenn, H. C., & Greth, H. E. (1990). Effect of environmental conditions of extracellular protease activity in lignolytic cultures of phanerochaetae chrusosporium. Applied and Environmental Microbiology, 56: 395-400. [ Links ]
Dosunmu, O. O. & Ajayi, A. B. (2002). Problems and management of sawmill waste in Lagos. Proceedings of International Environmental Pollution Control and Waste Management (EPCOWN 2002), pp. 271-278. [ Links ]
Dutkiewicz, J., Krysinska-Traczyk, E., Prazmo, Z., Skorska, C., & Sitkowska, J. (2001). Exposure to airborne microorganisms in Polish sawmills. Annals of Agricultural and Environmental Medicine, 8, 71-80. [ Links ]
Erikson, K. E., Blanchette, R. A., & Ander, P. (1990). Microbial and enzymatic degradation of wood and wood components. New York: Springer Verlag. [ Links ]
Garcia, C., Hernandez, T., Costa, F., Ceccanti, B., & Ciardi, C. (1992). Changes in ATP content, enzyme activity and inorganic nitrogen species during composting of organic wastes. Canadian Journal of Soil Science, 72, 243-253. [ Links ]
Godliving, M. Y. (2002). Continuous production of lignin degrading enzymes by Bjer kandora adusta immobilized on polyurethane foam. Biotechnology Letters, 24, 173-177. [ Links ]
Hameed, A. A., Khoder, M. I., & Farag, S. A. (1999). Organic dust and gaseous contaminants at wood working shops. Journal of Environmental Monitoring, 2, 73-76. [ Links ]
Harrigan, W. F. & McCance, M. E. (1976). Laboratory methods in fords and dairy microbiology. London: Academic Press. [ Links ]
Hornor, S. G. & Mitchell, M. J. (1981). Effect of earthworm E. fetida (Oligochaeta) on fluxes of volatile carbon sulfur compounds from sewage sludge. Soil Biology and Biochemistry, 13, 347-372. [ Links ]
Idowu, A. B., Edema, M. O., & Adeyi, A. O. (2006). Distribution of bacteria and fungi in the earthworm Libyodrillus violaceous (Annelida: Oligochaeta), a native earthworm from Nigeria. Revista de Biología Tropical, 54(1), 49-58. [ Links ]
Ihekwaba, A. E., Nwafor, A., & Adienbo, O. M. (2009). Lung Function Indices in Primary and Secondary Sawmill Workers in Port Harcourt Nigeria. African Journal of Applied Zoology and Environmental Biology, 11, 101-105. [ Links ]
Karsten, G. R. & Drake, H. L. (1995). Comparative assessment of the aerobic and anaerobic microfloras of earthworm and forest soils. Applied and Environmental Microbiology, 61(3), 1039-1044. [ Links ]
Kreger-venrij, W. (1984). The yeast- a taxonomic syudy. 3rd Ed., Amsterdam: Elservier Science Publisher. [ Links ]
Lee, K. E. (1985). Earthworms: Their ecology and relationship with soil and land use. Australia: Academic Press. [ Links ]
Lennox, J. A., Abriba, C., Alabi, B. N., & Akubuenyi, F. C. (2010). Comparative degradation of sawdust by microorganisms isolated from it. African Journal of Microbiology Research, 4(13), 1352-1355. [ Links ]
Maboeta, M. S. & Van Rensburg, L. (2003). Vermicomposting of industrially produced wood chips and sewage sludge utilizing Eisenia foetida. Ecotoxicology and Environmental Safety, 56, 265-270. [ Links ]
McDonald, C. E. & Chen, L. L. (1965). Lowry modification of the folin reagent for determination of proteinase activity. Analytical Biochemistry, 10, 175-177. [ Links ]
Nokrans, B. (1957). Studies of β-glucosidase and cellulose splitting enzymes from Polyporus annosus. French Physiological Plantarum, 10, 198-213. [ Links ]
Olayinka, O. T., Idowu, A. B., Dedeke, G. A., Akinloye, O. A., Ademolu, K. O., & Bamgbola, A. A. (2011). Earthworm as Bio-indicator of Heavy Metal Pollution around Lafarge, WapcoCement Factory, Ewekoro, Nigeria. Proceedings of the Environmental Management Conference, Federal University of Agriculture, Abeokuta, Nigeria, 489-496. [ Links ]
Owa, S. O., Dedeke, G. A., & Yeye, A. J. (2002). Earthworm cast characteristics under Bahama grass and the question of why earthworms cast. African Journal of Science and Technology, 3(1-2), 33-35. [ Links ]
Owa, S. O., Olowoparija, S. B., Aladesida, A., & Dedeke, G. A. (2013). Enteric bacteria and fungi of the Eudrilid earthworm Libyodrilus violaceus. African Journal of Agricultural Research, 8(17), 1760-1766. [ Links ]
Parthasarathi, K., & Ranganathan, L. S. (1998). Pressmud vermicasts are the ‘hot spots’ of fungi and bacteria. Ecology, Environment and Conservation, 4, 81-86. [ Links ]
Parthasarathi, K. & Ranganathan, L. S. (2000). Profiles of enzyme activity in the gut of Lampito mauritii and Eudrilus eugeniae reared on various substrates. Tropical Ecology 41(2), 251-254. [ Links ]
Parthasarathi, K., Ranganathan, L. S., Anandi V., & Zeyer, J. (2007). Diversity of microflora in the gut and casts of tropical composting earthworms reared on different substrates. Journal of Environmental Biology, 28(1), 87-97. [ Links ]
Renu, B., Pandey, H., Bisht, S. P., Kandpal, B., & Kaushal, B. R. (2006). Feeding and casting activities of the earthworm (Octolasion tyrtaeum) and their effects on crop growth under laboratory conditions. Tropical Ecology, 47(2), 291-294. [ Links ]
Satchell, J. E. (1967). Lumbricidae. In A. Burges & F. Raw (Eds.), Soil biology (pp. 259-322). London: Academic. [ Links ]
Shi, Y., Shi, Y., Wang, X., Lu, Y., & Yan, S. (2007). Comparative effects of lindane and deltamethrin on mortality, growth, and cellulase activity in earthworms (Eisenia fetida). Pesticide Biochemistry and Physiology, 89, 31-38. [ Links ]
Sneath, P. H. A., Mair, N. S., Sharpe, M. E., & Holt, J. G. (1986). Bregey’s manual of systematic bacteriology. (Vol. 2). Baltimore: Williams and Wilkins. [ Links ]
Swain, R. R. & Dekker, E. E. (1966). Seed germination studies I. Purification and properties of an amylase from the cotyledons germinating peas. Biochimia Biophysica Acta, 122, 75-86. [ Links ]
Vuorinen, A. H. (1999). Phosphatases in horse and chicken manure composts. Compost Science and Utilization, 7, 47-54. [ Links ]
Wilson, A. M. (1971). Amylase synthesis and stability in crested wheatgrass seed at low water potentials. Plant Physiology, 48, 541-546. [ Links ]
Yong, F. M. & Wood, B. J. (1977). Microbiology and biochemistry of ‘soy’-‘sauce’ fermentation. Advanced Applied Microbiology, 17, 157-194. [ Links ]
Zhang, B., Li, G., Shen, T., Wang, J., & Sun, Z. (2000). Changes in microbial biomass C, N, and P and enzyme activities in soil incubated with the earthworms Metaphire guillelmi or Eisenia fetida. Soil Biology and Biochemistry, 32, 2055-2062. [ Links ]
Ademolu, K. O. & Idowu, A. B. (2011). Occurrence and distribution of microflora in the gut regions of the Variegated grasshopper Zonocerus variegatus (Orthoptera: Pyrgomorphidae) during development. Zoological Studies, 50(4), 409-415. [ Links ]
Bonnarme, D. & Jeffries, T. W. (1998). Mn (11) regulation of lignin peroxidise and mangares-dependent peroxidise from lignin-degrading white-rot fungi. Applied Environmental Microbiology, 56, 210-217. [ Links ]
Debasmita, C., Mishra, C. S. K., & Mohapatra, P. K. (2011). Diversity of Bacteria and Fungi in the Gut of the Tropical Earthworm Glyphodrillus tuberosus Isolated from Conventional and Organic Rice Fields. Journal of Pharmacology and Toxicology, 6(3), 303-311. [ Links ]
Dedeke, G. A., Omemu, O., Aladesida, A. A., & Museliu, F. (2010). Comparative Microbial Analysis of Earthworm Casts Collected From Ikenne, Ogun State, Nigeria. Ethiopian Journal of Environmental Studies and Management, 3(3), 57-63. [ Links ]
Devi, S. H., Vijayalakshmi, K., Jyotsna, K. P., Shaheen, S. K., Jyothi, K., & Rani, M. S. (2009). Comparative assessment in enzyme activities and microbial populations during normal and vermicomposting. Journal of Environmental Biology, 30(6), 1013-1017. [ Links ]
Domsch, K. H., Gams, W., & Anderson, T. H. (1980). Compendium of soil fungi. London: Academic Press. [ Links ]
Dosoretz, G. G., Chenn, H. C., & Greth, H. E. (1990). Effect of environmental conditions of extracellular protease activity in lignolytic cultures of phanerochaetae chrusosporium. Applied and Environmental Microbiology, 56: 395-400. [ Links ]
Dosunmu, O. O. & Ajayi, A. B. (2002). Problems and management of sawmill waste in Lagos. Proceedings of International Environmental Pollution Control and Waste Management (EPCOWN 2002), pp. 271-278. [ Links ]
Dutkiewicz, J., Krysinska-Traczyk, E., Prazmo, Z., Skorska, C., & Sitkowska, J. (2001). Exposure to airborne microorganisms in Polish sawmills. Annals of Agricultural and Environmental Medicine, 8, 71-80. [ Links ]
Erikson, K. E., Blanchette, R. A., & Ander, P. (1990). Microbial and enzymatic degradation of wood and wood components. New York: Springer Verlag. [ Links ]
Garcia, C., Hernandez, T., Costa, F., Ceccanti, B., & Ciardi, C. (1992). Changes in ATP content, enzyme activity and inorganic nitrogen species during composting of organic wastes. Canadian Journal of Soil Science, 72, 243-253. [ Links ]
Godliving, M. Y. (2002). Continuous production of lignin degrading enzymes by Bjer kandora adusta immobilized on polyurethane foam. Biotechnology Letters, 24, 173-177. [ Links ]
Hameed, A. A., Khoder, M. I., & Farag, S. A. (1999). Organic dust and gaseous contaminants at wood working shops. Journal of Environmental Monitoring, 2, 73-76. [ Links ]
Harrigan, W. F. & McCance, M. E. (1976). Laboratory methods in fords and dairy microbiology. London: Academic Press. [ Links ]
Hornor, S. G. & Mitchell, M. J. (1981). Effect of earthworm E. fetida (Oligochaeta) on fluxes of volatile carbon sulfur compounds from sewage sludge. Soil Biology and Biochemistry, 13, 347-372. [ Links ]
Idowu, A. B., Edema, M. O., & Adeyi, A. O. (2006). Distribution of bacteria and fungi in the earthworm Libyodrillus violaceous (Annelida: Oligochaeta), a native earthworm from Nigeria. Revista de Biología Tropical, 54(1), 49-58. [ Links ]
Ihekwaba, A. E., Nwafor, A., & Adienbo, O. M. (2009). Lung Function Indices in Primary and Secondary Sawmill Workers in Port Harcourt Nigeria. African Journal of Applied Zoology and Environmental Biology, 11, 101-105. [ Links ]
Karsten, G. R. & Drake, H. L. (1995). Comparative assessment of the aerobic and anaerobic microfloras of earthworm and forest soils. Applied and Environmental Microbiology, 61(3), 1039-1044. [ Links ]
Kreger-venrij, W. (1984). The yeast- a taxonomic syudy. 3rd Ed., Amsterdam: Elservier Science Publisher. [ Links ]
Lee, K. E. (1985). Earthworms: Their ecology and relationship with soil and land use. Australia: Academic Press. [ Links ]
Lennox, J. A., Abriba, C., Alabi, B. N., & Akubuenyi, F. C. (2010). Comparative degradation of sawdust by microorganisms isolated from it. African Journal of Microbiology Research, 4(13), 1352-1355. [ Links ]
Maboeta, M. S. & Van Rensburg, L. (2003). Vermicomposting of industrially produced wood chips and sewage sludge utilizing Eisenia foetida. Ecotoxicology and Environmental Safety, 56, 265-270. [ Links ]
McDonald, C. E. & Chen, L. L. (1965). Lowry modification of the folin reagent for determination of proteinase activity. Analytical Biochemistry, 10, 175-177. [ Links ]
Nokrans, B. (1957). Studies of β-glucosidase and cellulose splitting enzymes from Polyporus annosus. French Physiological Plantarum, 10, 198-213. [ Links ]
Olayinka, O. T., Idowu, A. B., Dedeke, G. A., Akinloye, O. A., Ademolu, K. O., & Bamgbola, A. A. (2011). Earthworm as Bio-indicator of Heavy Metal Pollution around Lafarge, WapcoCement Factory, Ewekoro, Nigeria. Proceedings of the Environmental Management Conference, Federal University of Agriculture, Abeokuta, Nigeria, 489-496. [ Links ]
Owa, S. O., Dedeke, G. A., & Yeye, A. J. (2002). Earthworm cast characteristics under Bahama grass and the question of why earthworms cast. African Journal of Science and Technology, 3(1-2), 33-35. [ Links ]
Owa, S. O., Olowoparija, S. B., Aladesida, A., & Dedeke, G. A. (2013). Enteric bacteria and fungi of the Eudrilid earthworm Libyodrilus violaceus. African Journal of Agricultural Research, 8(17), 1760-1766. [ Links ]
Parthasarathi, K., & Ranganathan, L. S. (1998). Pressmud vermicasts are the ‘hot spots’ of fungi and bacteria. Ecology, Environment and Conservation, 4, 81-86. [ Links ]
Parthasarathi, K. & Ranganathan, L. S. (2000). Profiles of enzyme activity in the gut of Lampito mauritii and Eudrilus eugeniae reared on various substrates. Tropical Ecology 41(2), 251-254. [ Links ]
Parthasarathi, K., Ranganathan, L. S., Anandi V., & Zeyer, J. (2007). Diversity of microflora in the gut and casts of tropical composting earthworms reared on different substrates. Journal of Environmental Biology, 28(1), 87-97. [ Links ]
Renu, B., Pandey, H., Bisht, S. P., Kandpal, B., & Kaushal, B. R. (2006). Feeding and casting activities of the earthworm (Octolasion tyrtaeum) and their effects on crop growth under laboratory conditions. Tropical Ecology, 47(2), 291-294. [ Links ]
Satchell, J. E. (1967). Lumbricidae. In A. Burges & F. Raw (Eds.), Soil biology (pp. 259-322). London: Academic. [ Links ]
Shi, Y., Shi, Y., Wang, X., Lu, Y., & Yan, S. (2007). Comparative effects of lindane and deltamethrin on mortality, growth, and cellulase activity in earthworms (Eisenia fetida). Pesticide Biochemistry and Physiology, 89, 31-38. [ Links ]
Sneath, P. H. A., Mair, N. S., Sharpe, M. E., & Holt, J. G. (1986). Bregey’s manual of systematic bacteriology. (Vol. 2). Baltimore: Williams and Wilkins. [ Links ]
Swain, R. R. & Dekker, E. E. (1966). Seed germination studies I. Purification and properties of an amylase from the cotyledons germinating peas. Biochimia Biophysica Acta, 122, 75-86. [ Links ]
Vuorinen, A. H. (1999). Phosphatases in horse and chicken manure composts. Compost Science and Utilization, 7, 47-54. [ Links ]
Wilson, A. M. (1971). Amylase synthesis and stability in crested wheatgrass seed at low water potentials. Plant Physiology, 48, 541-546. [ Links ]
Yong, F. M. & Wood, B. J. (1977). Microbiology and biochemistry of ‘soy’-‘sauce’ fermentation. Advanced Applied Microbiology, 17, 157-194. [ Links ]
Zhang, B., Li, G., Shen, T., Wang, J., & Sun, Z. (2000). Changes in microbial biomass C, N, and P and enzyme activities in soil incubated with the earthworms Metaphire guillelmi or Eisenia fetida. Soil Biology and Biochemistry, 32, 2055-2062. [ Links ]
1. Biological Sciences Department, Federal University of Agriculture, P.M.B. 2240, Abeokuta, Nigeria; julius.bamidele@yahoo.com
2. Biological Sciences Department, Federal University of Agriculture, P.M.B. 2240, Abeokuta, Nigeria.
3. Biological Sciences Department, Federal University of Agriculture, P.M.B. 2240, Abeokuta, Nigeria.
4. Microbiology Department, Federal University of Agriculture, P.M.B. 2240, Abeokuta, Nigeria.
Received 13-I-2014. Corrected 20-iii-2014. Accepted 29-iv-2014.


{kind=link}
{kind=link}
{kind=link}