










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.3 San José Jul./Sep. 2014
Delimitation of some neotropical laccate Ganoderma (Ganodermataceae): molecular phylogeny and morphology
Ddelimitación de algunos Ganoderma (Ganodermataceae)lacados neotropicales: filogenia molecular y morfología
Ddelimitación de algunos Ganoderma (Ganodermataceae)lacados neotropicales: filogenia molecular y morfología
Abstract
Ganoderma includes species of great economic and ecological importance, but taxonomists judge the current nomenclatural situation as chaotic and poorly studied in the neotropics. From this perspective, phylogenetic analyses inferred from ribosomal DNA sequences have aided the clarification of the genus status. In this study, 14 specimens of Ganoderma and two of Tomophagus collected in Brazil were used for DNA extraction, amplification and sequencing of the ITS and LSU regions (rDNA). The phylogenetic delimitation of six neotropical taxa (G. chalceum, G. multiplicatum, G. orbiforme, G. parvulum, G. aff. oerstedtii and Tomophagus colossus) was determined based on these Brazilian specimens and found to be distinct from the laccate Ganoderma from Asia, Europe, North America and from some specimens from Argentina. Phylogenetic reconstructions confirmed that the laccate Ganoderma is distinct from Tomophagus, although they belong to the same group. The use of taxonomic synonyms Ganoderma subamboinense for G. multiplicatum, G. boninense for G. orbiforme and G. chalceum for G. cupreum was not confirmed. However, Ganoderma parvulum was confirmed as the correct name for specimens called G. stipitatum. Furthermore, the name G. lucidum should be used only for European species. The use of valid published names is proposed according to the specimen geographical distribution, their morphological characteristics and rDNA analysis. Rev. Biol. Trop. 62 (3): 1197-1208. Epub 2014 September 01.
Key words: Agaricomycetes, phylogenetic taxonomy, rDNA sequences, species delimitation, neotropics.
Resumen
Ganoderma incluye especies de gran importancia económica y ecológica, sin embargo, su nomenclatura actual es caótica y poco estudiada en el neotrópico. En este estudio se utilizaron 14 muestras de Ganoderma y dos de Tomophagus recolectados en Brasil para la extracción de ADN, amplificación y secuenciación de las regiones ITS y LSU. La delimitación filogenética de seis táxones neotropicales fue discutida con base en especímenes brasileños y secuencias del GenBank. Estas especies mostraron ser distintas de los Ganoderma lacados de Asia, Europa, América del Norte y de algunos ejemplares de Argentina. Las reconstrucciones filogenéticas confirman que los Ganoderma lacados son distintos de Tomophagus, aunque pertenecen al mismo grupo. No se confirman los sinónimos de G. subamboinense a G. multiplicatum, de G. boninense a G. orbiforme y G. chalceum a G. cupreum. G. parvulum se confirma como el nombre correcto para G. stipitatum. G. lucidum sólo se debe utilizar para especies europeas. Por lo tanto, se propone el uso de nombres publicados válidamente de acuerdo con la distribución geográfica de las muestras, características morfológicas y análisis de ADNr.
Palabras clave: Agaricomycetes, taxonomía filogenética, secuencias de ADNr, delimitación de especie, neotrópico.
Ganoderma P. Karst. (Ganodermataceae, Agaricomycetes) is one of the largest genera of Polyporales and was described by Karsten (1881) based on Polyporus lucidus (Curtis) Fr. from Europe, a species with a laccate (shiny varnished looking) surface. The genus is characterized by double-walled basidiospores with truncate apex and ornamented endospore (Moncalvo & Ryvarden, 1997). The genus includes 80 species of wide geographic distribution with several tropical species and others restricted to temperate areas (Ryvarden, 2000; Kirk, Cannon, Minter, & Stalpers, 2008). Ryvarden (2004) reported the presence of 20 species in the neotropics, although Torres-Torres, Guzmán-Dávalos & Gugliotta (2012) and Gugliotta, Abrahão & Gibertoni (2013) listed 18 and 28 species, respectively, only in Brazil.
Being well known as decomposers and pathogens in tropical forests (Zakaria, Ali, Salleh, & Zakaria, 2009), species of this genus, mainly of the G. lucidum complex, produce bioactive compounds widely studied for preventing and relieving human diseases, such as several types of tumors and cancers, gastric ulcers, diabetes mellitus, hypertension and viral infections (Zhou et al., 2007). Besides, some medicinal effects were reported in manuscripts of the Chinese civilization more than 2 000 years ago (Hong & Jung, 2004; Seo & Kirk, 2000).
Although of significant ecological and biotechnological importance, the taxonomy of laccate Ganoderma has been questioned in recent years and poorly studied in the neotropics. Currently, taxonomists consider the nomenclatural situation as chaotic and suggest the necessity for a global revision due to the existence of multiple names for single species (Ryvarden, 1991; Ryvarden, 2004; Moncalvo & Ryvarden, 1997; Postnova & Skolotneva, 2010).
Usually, different morphological characteristics of the basidiospores (dimensions) and basidiomata (thickness of the cuticle and presence or absence of resinaceous deposits in the context) are widely used in an attempt to identify laccate species of Ganoderma (Seo & Kirk, 2000; Ryvarden, 2004; Torres-Torres & Guzmán-Dávalos, 2012; Torres-Torres, Guzmán-Dávalos, & Gugliotta, 2012). However, the use of molecular data, especially for phylogenetic studies based on ribosomal DNA sequences, combined with morphological studies has helped many authors to clarify the status of the genus (Moncalvo, Wang, & Hseu, 1995a; Moncalvo, Wang, & Hseu, 1995b; Gottlieb, Ferrer, & Wright, 2000; Smith & Sivasithamparam, 2000; Hong & Jung, 2004; Kaliyaperumal & Kalaichelvan, 2008; Cao, Wu, & Dai, 2012; Yang & Feng, 2013).
In Brazil, there are only a few studies regarding Ganoderma (Torrend, 1920; Loguercio-Leite, Groposo, & Halmenschlager, 2005; Torres-Torres et al., 2012) and none of these include a phylogenetic analysis. Thus, the aim of the present study was to determine phylogenetic relationships of the laccate Ganoderma based on sequence variation of the Internal Transcribed Spacer (ITS) and Large subunit (LSU) rDNA and to delimit the species’ occurrence within Brazilian territory.
Material and methods
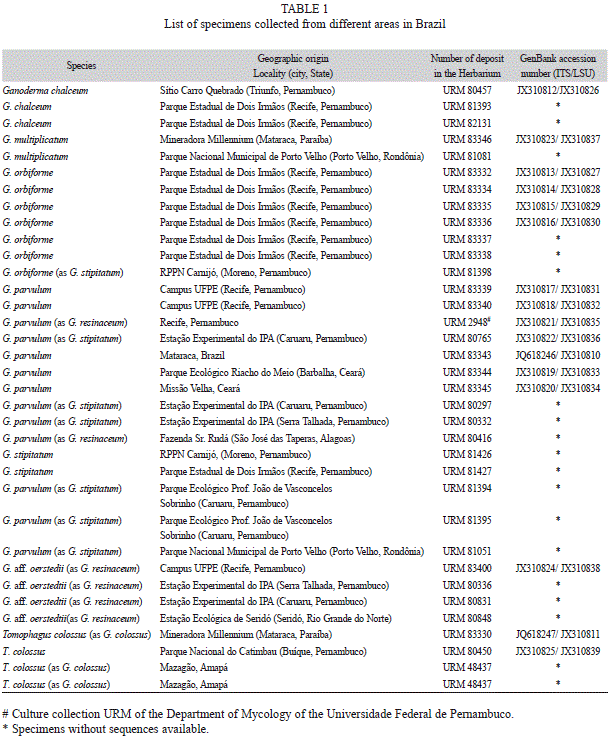
Morphological analysis: Thirty-one laccate specimens of Ganoderma and four of Tomophagus colossus (Fr.) Murrill were used in this study (Table 1). To observe the characteristics of the basidiospores, the hyphal system and the cuticle hyphae, free-hand thin sections of dried material were mounted in 5% KOH to ensure rehydration. Melzer’s reagent was used to test the amyloid reaction of the cuticle hyphae. Our data were then compared to the available literature (Bresadola, 1911; Gottlieb & Wright, 1999; Núñez & Ryvarden, 2000; Ryvarden, 2000; Ryvarden, 2004; Welti & Courtecuisse, 2010; Tham et al., 2012, Torres-Torres et al., 2012) and discussed in the text. All studied specimens were deposited in the herbarium Pe. Camille Torrend (URM), Department of Mycology, Universidade Federal de Pernambuco, Brazil.
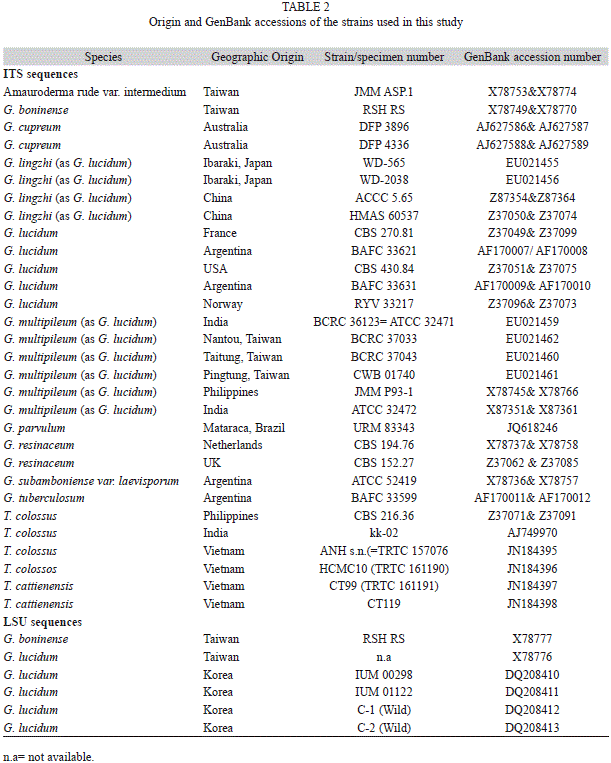
Genomic DNA extraction, polymerase chain reaction and sequencing: DNA was extracted using fragments of basidiomata (30-50mg) ground with a pestle in a porcelain mortar containing liquid nitrogen. The resulting powder was transferred to a tube containing 700μL of extraction buffer [CTAB 2%, 100mM Tris-HCl pH8, 1.4M NaCl, 20mM EDTA, 1% PVP (Rogers & Bendich, 1985)] and incubated at 65ºC for 30-40 min. DNA was purified with 700μL of chloroform-isoamyl alcohol (24:1), gently precipitated in 600μL of isopropanol and washed with 1mL of ethanol. Finally, the pellet was suspended in 50μL of ultrapure water (Góes-Neto, Loguercio-Leite, & Guerrero, 2005). The reaction mix and parameters for PCR amplification of the full ITS regions was according to Smith & Sivasithamparam (2000) using the primers ITS1 and ITS4 (White, Bruns, Lee, & Taylor, 1990). For LSU rDNA region, the amplification was performed with parameters and reagent concentrations following Góes-Neto, Loguercio-Leite & Guerrero (2005) using the primer pair LR0R and reverse LR7 (Moncalvo, Lutzoni, Rehner, Johnson, & Vilgalys, 2000). Negative controls containing all components of the reaction mix, except DNA, were used in each procedure to detect possible contamination. The amplification products of the sixteen laccate specimens (Table 1) were purified using the PureLink PCR Purification Kit (Invitrogen) and the purified products were sequenced at the Human Genome Research Center of the Universidade de São Paulo (USP, Brazil) in an ABI-310 Capillary Sequencer (PerkinElmer, Wellesley Massachusetts, USA). Cycle sequencing was carried out with primers ITS1 and ITS4 for ITS region and LR0R and LR5 for LSU region (Moncalvo et al., 2000). All sequences were deposited in GenBank (National Center for Biotechnology Information, Bethesda, Maryland, USA).
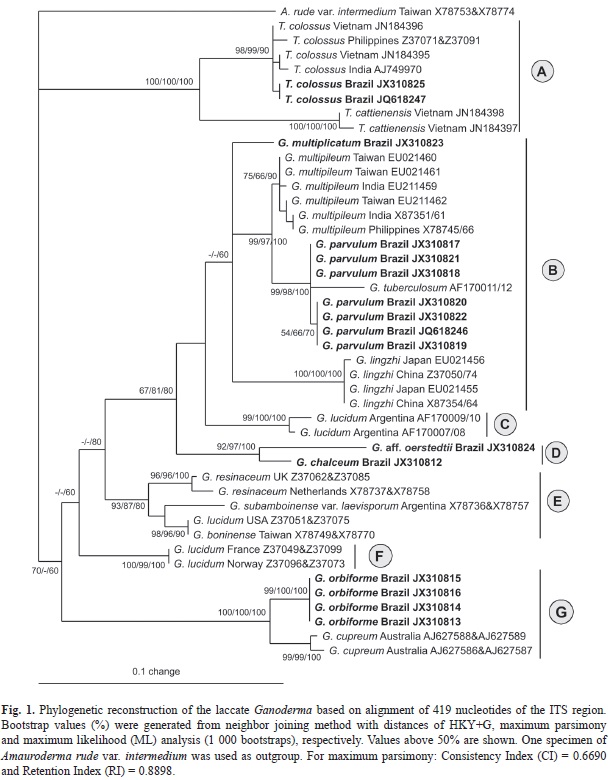
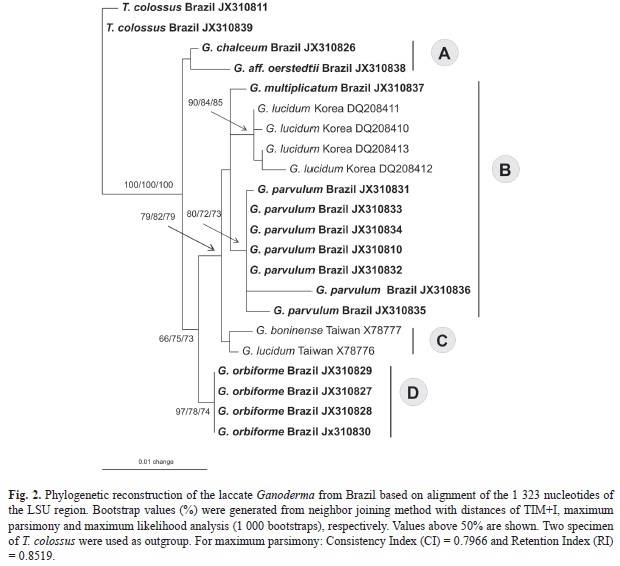
Phylogenetic analysis: Sixteen ITS and LSU rDNA sequences (14 of Ganoderma and two of T. colossus) of laccate Ganodermataceae were compared with other sequences retrieved from GenBank (Table 2). These sequences were aligned using ClustalX (Larkin, 2007), manually edited in BioEdit (Hall, 1999) and realigned to obtain the final alignment. Phylogenetic analyses and tree construction were performed separately for each locus. Neighbor joining (NJ) distances, maximum parsimony (MP) and maximum likelihood (ML) analyses were carried out using PAUP* version 4.0b10 (Swofford, 2002) and the support was evaluated using 1 000 bootstrap replicates. The NJ and ML analysis were based on HKY+G (ITS) and TIM+I (LSU) obtained from ModelTest 3.7 (Posada & Crandall, 1998), which computed the most likely patterns of phylogenetic evolution. Sequences from Amauroderma rude var. intermedium J. S. Furtado were used as outgroup for phylogenetic reconstruction based on ITS sequences and two specimens of T. colossus were used as outgroup for the LSU analysis.
Results
The ITS1-5.8S-ITS2 regions sequenced in this study varied in length from 548 to 571 nucleotides. The size of the ITS1 region did not differ markedly among the studied specimens, ranging from 199 to 205 nucleotides. This small variation was also observed for the ITS2 region and ranged between 192 and 201 nucleotides. The final alignment (ITS1 + ITS2) included 419 sites, with 266 constant sites (63%) and 153 variable (36%), of which 120 (28%) were parsimony informative.
The size of LSU sequences ranged from 1 320 to 1 322 nucleotides and aligned at 1 323 positions. Of these, 1 278 characters were constant, 21 characters were variable but parsimony uninformative, and 24 characters were parsimony informative. Although this study provided new sequences within the ITS and LSU regions, it is as yet impossible to perform multigene analysis due to the lack of other neotropical gene sequences from the species analyzed here.
The phylogenetic reconstruction performed with NJ, MP and ML analyses for ITS sequences showed basically the same topology and few differences in bootstrap values (Fig. 1). The same was observed for LSU analysis (Fig. 2). These results confirm that laccate Ganoderma is a monophyletic group although with low statistical support based on ITS analysis (NJ 70%; MP < 50% and ML 60%) and with high bootstrap values based on LSU analysis (NJ, MP and ML 100%). In the phylogenetic reconstruction based on ITS regions, seven clades were delimited (A, B, C, D, E, F and G). The Brazilian specimens of the six taxa studied (G. chalceum, G. multiplicatum, G. orbiforme, G. parvulum, G. aff. oerstedtii and T. colossus) were recovered in clades A, B, D and G, discussed below.
Discussion
The clade A formed a monophyletic lineage distinct from the laccate Ganoderma with strong statistical support (NJ, MP and ML = 100%) and included two species, T. colossus and T. cattienensis. Tomophagus colossus from Brazil grouped with representatives of the Asian species (NJ 98%, MP 99% and ML 90%), confirming that they are of the same species, but distinct from T. cattienensis. The genus Tomophagus was established first by Murrill (1905) and in the following decades was contested, being considered a confusing group. Later, molecular phylogenetic studies confirmed the genus as group well established in Ganodermataceae (Moncalvo, Wang, Wang, & Hseu, 1995c; Hong & Jung, 2004; Tham et al., 2012).
However, recent papers still mention Tomophagus as belonging to Ganoderma (Welti & Courtecuisse, 2010; Cao et al., 2012). Tomophagus currently has two species, T. colossus (= G. colossus) and T. cattienensis, both sharing the laccate (shiny) pilear surface, pale context with slightly dextrinoid skeletal hyphae, large basidiospores, and the striking chlamydospores, providing a unique combination of characters (Ryvarden, 2000; Ryvarden, 2004; Tham et al., 2012). Tomophagus colossus differs morphologically from T. cattienensis by having yellowish pilear surface while T. cattienensis has red to light brown pilear surface. Furthermore, T. cattienensis has a context that turns pale brown upon drying, instead of remaining creamy white as T. colossus, and shows slightly larger basidiospores [17.5-21.5 x 11.5-14.5μm (Tham et al., 2012) versus 16-19 x 10.5-12.5μm in T. colossus from Brazil. This size is consistent with Ryvarden’s (2004) descriptions].
Clade B was composed of specimens of G. multiplicatum, G. multipileum, G. parvulum, G. tuberculosum and G. lingzhi. Similar topology was also observed in the phylogenetic analysis based on LSU sequences for the available species (G. multiplicatum, “G. lucidum”, G. parvulum) (Fig. 2).
Ganoderma multiplicatum is a neotropical species described from Venezuela and is characterized by having amyloid, slightly tuberculate hyphal ends in the cuticle, and subglobose to ellipsoid basidiospores (7.5-8.5 x 5-6μm). The Brazilian specimens studied agree with the descriptions of Gottlieb & Wright (1999), Ryvarden (2000, 2004) and Torres-Torres, Guzmán-Dávalos, & Gugliotta (2012). Ryvarden (2000) recognized G. subamboinense Henn. as a synonym of G. multiplicatum, originally described from Brazil. Although morphologically similar [context with two or more black, resinous layers, hyphal ends in the cuticle generally amyloid and basidiospores 8-10 x 6-7μm (Gottlieb & Wright, 1999)], G. subamboinense Henn var. laevisporum Bazzalo & J.E. Wright ATCC 52419 (with a single sequence available for the species) were shown here to be distantly related (clade E), and thus should not be considered as synonyms.
Of the specimens here identified as G. parvulum by morphology and phylogenetic analysis, one was deposited in Herbarium URM as G. stipitatum (URM 80765) and another in the Culture Collection URM as G. resinaceum (URM 2948). For a long time, G. parvulum was considered synonym of G. stipitatum (Moncalvo & Ryvarden, 1997; Ryvarden, 2004). We agree that these are the same species, however, we follow the opinion of Torres-Torres et al. (2012) that the name G. parvulum Murrill 1902 should be used in preference to G. stipitatum (Murrill) Murrill 1908 (Fomes stipitatus Murrill 1903) as it was described earlier. Steyaert (1980) cited a wrong reference for the G. parvulum protologue and this was followed by Ryvarden (2004).
Ganoderma resinaceum URM 2948 is probably G. parvulum incorrectly identified. The basidiomata of G. resinaceum used for the original identification of the strain was not deposited in any Herbarium, preventing re-identification of the specimen. These two species are macromorphologically similar, and, microscopically, they both have smooth, weakly amyloid hyphal ends of the cuticle. However, G. resinaceum has larger basidiospores [9-11.5 x 5-7μm according to Ryvarden (2004) and 11.2-12.5 x 6.5-7.4μm according to Torres-Torres et al. (2012) versus 8-10 x 5-6μm in G. parvulum from Brazil)] and no black, resinous layers in the pale brown context (Ryvarden, 2004; Torres-Torres et al., 2012). The basidiospores of the Brazilian specimens of G. parvulum were similar to the description in Ryvarden (2004, as G. stipitatum). Furthermore, G. resinaceum was described on the basis of a specimen from France and G. parvulum was originally described from Brazil.
Similarly, the Argentinean specimen described as G. tuberculosum (BAFC 33599) (Gottlieb & Wright, 1999) also corresponds to G. parvulum. Ganoderma tuberculosum is characterized by a black, thick resinose band in the context similar to G. parvulum, but has longer basidiospores [10-11 x 7.5-9μm according to Welti & Courtecuisse (2010) versus 8-10 x 5-6μm in G. parvulum from Brazil]. Besides, G. tuberculosum does not have amyloid cuticle elements unlike G. parvulum, whose elements are distinctly amyloid. Gottlieb & Wright (1999) commented that the specimen BAFC 33599 was preliminarily identified as G. resinaceum, but was later revised to G. tuberculosum. However, they observed cuticle hyphal ends distinctly amyloid characteristic of G. parvulum and different from what was observed in the type of G. tuberculosum deposited in the Herbarium of the New York Botanical Garden in the study by Welti & Courtecuisse (2010). Yet, Gottlieb & Wright (1999) noted basidiospores ovoid to ellipsoid, 10-12 x 6-9μm, in three Argentinian specimens, including BAFC 33599, similar to the observed in G. tuberculosum. It is likely that the size of basidiospores is not a taxonomic criterion relevant for separating these two species, but the presence or absence of amyloid reaction of the cuticle elements is.
The specimens of G. parvulum and G. multiplicatum proved to be distinct from the specimens previously listed as G. lucidum for Asia (G. multipileum and G. lingzhi in figure 1; G. lucidum in Fig. 2) and from specimens still regarded as G. lucidum in South America, North America and Europe (clades C, E and F, respectively, in Fig. 1). Due to the high phenotypic plasticity of the subgenus Ganoderma, several species have been mistaken for G. lucidum strictu sensu. Torres-Torres et al. (2012) commented that Brazilian specimens of G. multiplicatum and G. parvulum have been mistaken for G. lucidum.
Although reports of G. lucidum in the neotropics are still found in the literature (Torres-Torres et al., 2012; Vasco-Palacios & Franco-Molano, 2013), in Herbaria records (http://emuweb.fieldmuseum.org/botany/crfResultsList.php, http://splink.cria.org.br) and in online databases (http://www.cybertruffle.org.uk/venefung/eng/index.htm, http://www.mycokey.com/Ecuador/EcuadorDB.htm, Gugliotta et al., 2013), we are of the opinion that this is a species restricted to Europe, as advocated by different authors (Moncalvo & Ryvarden, 1997; Cao et al., 2012). Thus, non-European G. lucidum corresponds to other species. Wang et al. (2009), using morphological and molecular data based on ITS sequences, observed that the specimens from tropical Asia identified preliminarily as G. lucidum were G. multipileum, whose type specimen is originally Asian. In a similar study, Cao, Wu, & Dai (2012), also using morphological and molecular studies based on ITS sequences and including strains widely cultivated in China, observed that G. lingzhi is the correct name for G. lucidum in East Asia. Similar approaches should be taken for all non-European G. lucidum.
In clade D, the specimen of G. chalceum grouped with one Brazilian specimen of G. aff. oerstedtii (URM83400) (NJ 92%, MP 97% and ML 100%, for ITS analysis), both belonging to the G. resinaceum complex. This clade can also be observed in the phylogenetic reconstruction based on LSU sequences, although with low statistical support (NJ, MP an ML < 50%). Basidiospores are smaller in G. chalceum [10-12 x 5-7μm versus 12-15 x 8-10μm in G. aff. oerstedtii (Ryvarden, 2004)] and the black resinous layer in the context is absent in G. aff. oerstedtii (present in G. chalceum). Thus, in this clade, the size of basidiospores and presence of resinous deposits in the context seem to be important characters for species delimitation. Ganoderma aff. oerstedii URM83400 was initially identified as G. resinaceum, but the use of G. resinaceum for South America specimens is not appropriate since two specimens of G. resinaceum (type locality: France) from Europe (clade E) are distinctly related. The material of G. aff. oerstedtii (URM83400) is scarce and more collections are desirable in order to confirm this species.
Clade G is composed by specimens of G. orbiforme and G. cupreum. All specimens of G. orbiforme clustered with high statistical support (ITS analysis: NJ 99%, MP and ML 100%; LSU analysis: NJ 97%, MP 78% and ML 74%) and were clearly distinct from the other laccate species.
The Index Fungorum databases and Ryvarden (2004) recognize G. boninense as synonym of G. orbiforme. However, the Mycobank database considers G. boninense as a distinct species, a conclusion supported by our study (G. boninense: clade E in ITS analysis, clade C in LSU analysis; G. orbiforme: clade G in ITS analysis, clade D in LSU analysis). Both species have similar basidiospore size [9-11μm in the Brazilian G. orbiforme versus 10-12μm (Núñez & Ryvarden, 2000) and 8.5-12μm (Chang, 1992) in G. boninense], and irregular, amyloid cuticle hyphal ends, but resinous layers are present in G. orbiforme and apparently absent in G. boninense (Chang, 1992; Núñez & Ryvarden, 2000). Ganoderma orbiforme was originally described from Guinea in Africa and also recorded in the neotropics, while G. boninense was originally described from Bonin Island in Japan and has been reported throughout the Pacific Islands and Sri Lanka, Australia, Taiwan, Japan and China (Chang, 1992; Moncalvo et al., 1995b). Apparently, the geographical distribution of the species was not considered relevant when the synonym was proposed.
Similarly, the Index Fungorum and MycoBank databases consider G. chalceum as a synonym for G. cupreum (clade E in ITS analysis). Both species have similar basidiospore size [(10-12 × 5-7μm in G. chalceum versus 8-11 × 5-7μm in G. cupreum (Bresadola, 1911)] and were originally described from western Africa. Contrary to G. cupreum, G. chalceum has been reported to the neotropics (Ryvarden, 2000; Ryvarden, 2004; Torres-Torres et al., 2012). The sequences of G. cupreum are of Australian origin and Smith & Sivasithamparam (2000) commented that more research was needed to verify that the isolates were correctly named. Moreover, there are no sequences of G. cupreum and G. chalceum originating from the type locality for better comparison.
In this study, we delimit six laccate taxa of Ganodermataceae collected in Brazil based on rDNA analyses with the support of morphological characters, mostly size of basidiospores, presence/absence of dark, resinous layers in the context, and presence/absence of amyloid hyphal ends in the cuticle. In addition, geographical distribution is also considered relevant as all Brazilian species differ from the previously known laccate Ganoderma from Asia, Europe, North and South America.
Acknowledgments
We are grateful to Allyne C. Gomes Silva for assistance in the identification of specimens and critical comments; the curator of Herbarium URM/UFPE, Leonor C. Maia, and curator of Micoteca URM/UFPE, Cristina M. de Souza Motta, for providing the specimens; Karl-Henrik Larsson (University of Oslo, Noway), Adriana de M. Gugliotta (Instituto de Botânica de São Paulo SP, Brazil) and Aristóteles Góes-Neto (Universidade Estadual de Feira de Santana BA, Brazil) for critical comments; Gladstone A. Silva for his help in DNA sequence analysis; Marcela Cárceres and Robert Lucking for reviewing the summary in Spanish. The first author also thanks the Fundação de Amparo a Ciência e Tecnologia do Estado de Pernambuco (FACEPE, IBPG-0677-2.02/09) for the scholarship and the Pós-Graduação em Biologia de Fungos (UFPE, Brazil). This research was supported by CNPq (Universal 470303/2009-3) and FACEPE (APQ-0444-2.03/08).
References
Bresadola, G. (1911). Fungi Congoenses. Annales Mycologici, 9, 266-276. [ Links ]
Cao, Y., Wu, S. H., & Dai, Y. C. (2012). Species clarification of the prize medicinal Ganoderma mushroom “Lingzhi”. Fungal Diversity, 56 (1), 49-62. [ Links ]
Chang, T. T. (1992). One terrestrial and some lignicolous Aphyllophorales (Basidiomycota) from Taiwan. Botanical Bulletin of Academia Sinica, 33, 327-334. [ Links ]
Góes-Neto, A., Loguercio-Leite, C., & Guerrero, R. T. (2005). DNA Extraction from frozen field-collected and dehydrated herbarium fungal basidiomata: perform of SDS and CTAB-based methods. Biotemas, 18(2), 19-32. (Recovered from https://periodicos.ufsc.br/index.php/biotemas/article/download/21410/19377). [ Links ]
Gottlieb, A. M. & Wright, J. E. (1999). Taxonomy of Ganoderma from southern South America: subgenus Ganoderma. Mycological Research, 103, 661-673. [ Links ]
Gottlieb, A. M., Ferrer, E., & Wright, J. E. (2000). rDNA analyses as an aid to the taxonomy of species of Ganoderma. Mycological Research, 104, 661-673. [ Links ]
Gugliotta, A. M., Abrahão, M. C., & Gibertoni, T. B. (2013). Polyporales in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. (Recovered from http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB92411). [ Links ]
Hall, T. A. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41, 95-98. (Recovered from http://jwbrown.mbio.ncsu.edu/JWB/papers/1999Hall1). [ Links ]
Hong, S. G. & Jung, H. S. (2004). Phylogenetic analysis of Ganoderma based on nearly complete mitochondrial small-subunit ribosomal DNA sequences. Mycologia, 96(4), 742-755. [ Links ]
Kaliyaperumal, M. & Kalaichelvan, P. T. (2008). Ganoderma australe from southern India. Microbiological Research, 163, 286-292. [ Links ]
Karsten, P. A. (1881). Enumeratio boletinearum et polyporarum fennicarum. Systemate novo dispositorum. Revue de Mycologie, 3, 16-19. [ Links ]
Kirk, P. M., Cannon, P. F., Minter, D. W., & Stalpers, J. A. (2008). Dictionary of the Fungi. 10º Ed, CAB International. [ Links ]
Kaliyaperumal, M. & Kalaichelvan, P. T. (2008). Ganoderma australe from southern India. Microbiological Research, 163, 286-292. [ Links ]
Larkin, M. A. (2007). Clustal W and Clustal X version 2.0. Bioinformatics, 23, 2947-2948. [ Links ]
Loguercio-Leite, C., Groposo, C., & Halmenschlager, M. A. (2005). Species of Ganoderma Karsten in a subtropical area (Santa Catarina State, Southern Brazil). Iheringia, 60, 135-139. [ Links ]
Moncalvo, J. M., Wang, H. H., & Hseu R. S. (1995a). Phylogenetic relationships in Ganoderma inferred from the internal transcribed spacers and 25S ribosomal DNA sequences. Mycologia, 87, 223-258. [ Links ]
Moncalvo, J. M., Wang, H. H., & Hseu, R. S. (1995b). Gene phylogeny of the Ganoderma lucidum complex based on ribosomal DNA sequences. Mycological Research, 99, 1489-1499. [ Links ]
Moncalvo, J. M., Wang, H. F., Wang, H. H., & Hseu, R. S. (1995c). The use of ribosomal DNA sequence data for species identification and phylogeny in the Ganodermataceae. In P. K. Buchanan, R. S. Hseu, & J. M. Moncalvo, (Ed.). Ganoderma, systematics, phytopathology and pharmacology (pp. 31-44). Proceedings of Contributed Symposium 59 A,B, 5th International Mycological Congress, Vancouver, August 14-21, 1994. Taipei. [ Links ]
Moncalvo, J. M. & Ryvarden, L. (1997). A nomenclatural study of the Ganodermataceae Donk. Fungiflora, Oslo, 11, 1-114. [ Links ]
Moncalvo, J. M., Lutzoni, F. M., Rehner, A. S., Johnson, J., & Vilgalys, R. (2000). Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences. Systematic Biology, 49, 278-305. [ Links ]
Murrill, W. A. (1908). Polyporaceae, Part 2. North American Flora, 9(2), 73-131. [ Links ]
Murrill, W. A. (1905). Tomophagus for Dendrophagus. Torreya, 5, 197. [ Links ]
Núñez, M. & Ryvarden, L. (2000). East Asian Polypores. Vol. 1. Fungiflora, Oslo. [ Links ]
Posada, D. & Crandall, K. A. (1998). Modeltest: testing the model of DNA substitution. Bioinformatics, 14, 817-818. [ Links ]
Postnova, E. & Skolotneva, E. S. (2010). Ganoderma lucidum complex: some individual groups of strains. Microbiology, 79(2), 63-71. [ Links ]
Rogers, S. O. & Bendich, A. J. (1985). Extraction of DNA from milligram amounts of fresh, herbarium, and mummified plant tissues. Plant Molecular Biology, 5, 69-76. [ Links ]
Ryvarden, L. (1991). Genera of polypores, nomenclature and taxonomy. Synopsis Fungorum, 5, 1-363. [ Links ]
Ryvarden, L. (2000). Studies in neotropical polypores 2: a preliminary key to neotropical species of Ganoderma with a laccate pileus. Mycologia, 92, 180-191. [ Links ]
Ryvarden, L. (2004). Neotropical Polypores. Part 1. Synopsis Fungorum, 19, 69-103. [ Links ]
Seo, G. S. & Kirk, P. M. (2000). Ganodermataceae: Nomenclature and classification. In P. D. Bridge & P. Holderness (Eds.), Ganoderma Disease of Perennial Crops (pp. 3-22). Wallingford, UK: CABI Publishing. [ Links ]
Smith, B. J. & Sivasithamparam, K. (2000). Isozymes of Ganoderma species from Australia. Mycological Research, 104(8), 952-961. [ Links ]
Steyaert, R. L. (1967). Les Ganoderma palmicoles. Bulletin du Jardin Botanique National de Belgique, 37(4), 465-492. [ Links ]
Swofford, D. L. (2002). PAUP*. Phylogenetic analysis using parsimony (*and other methods). Sunderland MA: Sinauer Associates. [ Links ]
Tham, L. X., Hung, N. L. Q. H., Duong, P. N. D., Hop, D. V., Dentinger, B. T. M., Moncalvo, J. M. (2012). Tomophagus cattienensis sp. nov., a new Ganodermataceae species from Vietnam: Evidence from morphology and ITS DNA barcodes. Mycological Progress, 11(3), 775-780. (Recovered from DOI:10.1007/s11557-011-0789-3). [ Links ]
Torrend, C. (1920). Les polyporacées du Brésil I. Le genre Ganoderma. Brotéria Série Botanica, 18, 23-43. [ Links ]
Torres-Torres, M. G., & Guzmán-Dávalos, L. (2012). The morphology of Ganoderma species with a laccate surface. Mycotaxon, 16, 201-216. Recovered from http://dx.doi.org/10.5248/119.201). [ Links ]
Torres-Torres, M. G., Guzmán-Dávalos, L., & Gugliotta, A. M. (2012). Ganoderma in Brazil: known species and new records. Mycotaxon, 121, 93-132. (Recovered from http://dx.doi.org/10.5248/121.93). [ Links ]
Vasco-Palacios, A., & Franco-Molano, A. E. (2013). Diversity of Colombian macrofungi (Ascomycota-Basidiomycota). Mycotaxon, 121, 499. [ Links ]
Wang, D. M., Wu, S. H., Su, C. H., Peng, J. T., Shih, Y. H., & Chen L. C. (2009). Botanical Studies, 50, 451-458. (Recovered from http://ejournal.sinica.edu.tw/bbas/content/2009/4/Bot504-08). [ Links ]
White, T. J., Bruns, T., Lee, S., & Taylor, J. W. (1990). Amplification and direct sequencing of ribosomal RNA genes for phylogenetics. In Innis, M. A., Gelfand, D. H., Sninsky, J. J., & White, T. J. PCR Protocols, a Guide to Methods and Applications. Academic Press, New York, 315-322. [ Links ]
Welti, S., & Courtecuisse, R. (2010). The Ganodermataceae in the French West Indies (Guadeloupe and Martinique). Fungal Diversity, 43, 103-126. (Recovered from DOI:10.1007/s13225-010-0036-2). [ Links ]
Yang, Z. L., & Feng, B. (2013). What is the Chinese “Lingzhi”? - A taxonomic mini-review. Mycology, 4(1), 1-4. (Recovered from DOI:10.1080/21501203.2013.774299). [ Links ]
Zakaria, L., Ali, N. S., Salleh B., & Zakaria, M. (2009). Molecular analysis of Ganoderma species from different hosts in Peninsula Malaysia. Journal of biological Sciences, 9(1), 12-20. (Recovered from: DOI:10.3923/jbs.2009.12.20). [ Links ]
Zhou, X., Lin, J., Yin, Y., Zhao, J., Sun, X., & Tang, K. (2007). Ganodermataceae: natural products and their related pharmacological functions. American Journal of Chinese Medicine, 35(4), 559-574. (Recovered from DOI: 10.1142/S0192415X07005065). [ Links ]
Cao, Y., Wu, S. H., & Dai, Y. C. (2012). Species clarification of the prize medicinal Ganoderma mushroom “Lingzhi”. Fungal Diversity, 56 (1), 49-62. [ Links ]
Chang, T. T. (1992). One terrestrial and some lignicolous Aphyllophorales (Basidiomycota) from Taiwan. Botanical Bulletin of Academia Sinica, 33, 327-334. [ Links ]
Góes-Neto, A., Loguercio-Leite, C., & Guerrero, R. T. (2005). DNA Extraction from frozen field-collected and dehydrated herbarium fungal basidiomata: perform of SDS and CTAB-based methods. Biotemas, 18(2), 19-32. (Recovered from https://periodicos.ufsc.br/index.php/biotemas/article/download/21410/19377). [ Links ]
Gottlieb, A. M. & Wright, J. E. (1999). Taxonomy of Ganoderma from southern South America: subgenus Ganoderma. Mycological Research, 103, 661-673. [ Links ]
Gottlieb, A. M., Ferrer, E., & Wright, J. E. (2000). rDNA analyses as an aid to the taxonomy of species of Ganoderma. Mycological Research, 104, 661-673. [ Links ]
Gugliotta, A. M., Abrahão, M. C., & Gibertoni, T. B. (2013). Polyporales in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. (Recovered from http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB92411). [ Links ]
Hall, T. A. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41, 95-98. (Recovered from http://jwbrown.mbio.ncsu.edu/JWB/papers/1999Hall1). [ Links ]
Hong, S. G. & Jung, H. S. (2004). Phylogenetic analysis of Ganoderma based on nearly complete mitochondrial small-subunit ribosomal DNA sequences. Mycologia, 96(4), 742-755. [ Links ]
Kaliyaperumal, M. & Kalaichelvan, P. T. (2008). Ganoderma australe from southern India. Microbiological Research, 163, 286-292. [ Links ]
Karsten, P. A. (1881). Enumeratio boletinearum et polyporarum fennicarum. Systemate novo dispositorum. Revue de Mycologie, 3, 16-19. [ Links ]
Kirk, P. M., Cannon, P. F., Minter, D. W., & Stalpers, J. A. (2008). Dictionary of the Fungi. 10º Ed, CAB International. [ Links ]
Kaliyaperumal, M. & Kalaichelvan, P. T. (2008). Ganoderma australe from southern India. Microbiological Research, 163, 286-292. [ Links ]
Larkin, M. A. (2007). Clustal W and Clustal X version 2.0. Bioinformatics, 23, 2947-2948. [ Links ]
Loguercio-Leite, C., Groposo, C., & Halmenschlager, M. A. (2005). Species of Ganoderma Karsten in a subtropical area (Santa Catarina State, Southern Brazil). Iheringia, 60, 135-139. [ Links ]
Moncalvo, J. M., Wang, H. H., & Hseu R. S. (1995a). Phylogenetic relationships in Ganoderma inferred from the internal transcribed spacers and 25S ribosomal DNA sequences. Mycologia, 87, 223-258. [ Links ]
Moncalvo, J. M., Wang, H. H., & Hseu, R. S. (1995b). Gene phylogeny of the Ganoderma lucidum complex based on ribosomal DNA sequences. Mycological Research, 99, 1489-1499. [ Links ]
Moncalvo, J. M., Wang, H. F., Wang, H. H., & Hseu, R. S. (1995c). The use of ribosomal DNA sequence data for species identification and phylogeny in the Ganodermataceae. In P. K. Buchanan, R. S. Hseu, & J. M. Moncalvo, (Ed.). Ganoderma, systematics, phytopathology and pharmacology (pp. 31-44). Proceedings of Contributed Symposium 59 A,B, 5th International Mycological Congress, Vancouver, August 14-21, 1994. Taipei. [ Links ]
Moncalvo, J. M. & Ryvarden, L. (1997). A nomenclatural study of the Ganodermataceae Donk. Fungiflora, Oslo, 11, 1-114. [ Links ]
Moncalvo, J. M., Lutzoni, F. M., Rehner, A. S., Johnson, J., & Vilgalys, R. (2000). Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences. Systematic Biology, 49, 278-305. [ Links ]
Murrill, W. A. (1908). Polyporaceae, Part 2. North American Flora, 9(2), 73-131. [ Links ]
Murrill, W. A. (1905). Tomophagus for Dendrophagus. Torreya, 5, 197. [ Links ]
Núñez, M. & Ryvarden, L. (2000). East Asian Polypores. Vol. 1. Fungiflora, Oslo. [ Links ]
Posada, D. & Crandall, K. A. (1998). Modeltest: testing the model of DNA substitution. Bioinformatics, 14, 817-818. [ Links ]
Postnova, E. & Skolotneva, E. S. (2010). Ganoderma lucidum complex: some individual groups of strains. Microbiology, 79(2), 63-71. [ Links ]
Rogers, S. O. & Bendich, A. J. (1985). Extraction of DNA from milligram amounts of fresh, herbarium, and mummified plant tissues. Plant Molecular Biology, 5, 69-76. [ Links ]
Ryvarden, L. (1991). Genera of polypores, nomenclature and taxonomy. Synopsis Fungorum, 5, 1-363. [ Links ]
Ryvarden, L. (2000). Studies in neotropical polypores 2: a preliminary key to neotropical species of Ganoderma with a laccate pileus. Mycologia, 92, 180-191. [ Links ]
Ryvarden, L. (2004). Neotropical Polypores. Part 1. Synopsis Fungorum, 19, 69-103. [ Links ]
Seo, G. S. & Kirk, P. M. (2000). Ganodermataceae: Nomenclature and classification. In P. D. Bridge & P. Holderness (Eds.), Ganoderma Disease of Perennial Crops (pp. 3-22). Wallingford, UK: CABI Publishing. [ Links ]
Smith, B. J. & Sivasithamparam, K. (2000). Isozymes of Ganoderma species from Australia. Mycological Research, 104(8), 952-961. [ Links ]
Steyaert, R. L. (1967). Les Ganoderma palmicoles. Bulletin du Jardin Botanique National de Belgique, 37(4), 465-492. [ Links ]
Swofford, D. L. (2002). PAUP*. Phylogenetic analysis using parsimony (*and other methods). Sunderland MA: Sinauer Associates. [ Links ]
Tham, L. X., Hung, N. L. Q. H., Duong, P. N. D., Hop, D. V., Dentinger, B. T. M., Moncalvo, J. M. (2012). Tomophagus cattienensis sp. nov., a new Ganodermataceae species from Vietnam: Evidence from morphology and ITS DNA barcodes. Mycological Progress, 11(3), 775-780. (Recovered from DOI:10.1007/s11557-011-0789-3). [ Links ]
Torrend, C. (1920). Les polyporacées du Brésil I. Le genre Ganoderma. Brotéria Série Botanica, 18, 23-43. [ Links ]
Torres-Torres, M. G., & Guzmán-Dávalos, L. (2012). The morphology of Ganoderma species with a laccate surface. Mycotaxon, 16, 201-216. Recovered from http://dx.doi.org/10.5248/119.201). [ Links ]
Torres-Torres, M. G., Guzmán-Dávalos, L., & Gugliotta, A. M. (2012). Ganoderma in Brazil: known species and new records. Mycotaxon, 121, 93-132. (Recovered from http://dx.doi.org/10.5248/121.93). [ Links ]
Vasco-Palacios, A., & Franco-Molano, A. E. (2013). Diversity of Colombian macrofungi (Ascomycota-Basidiomycota). Mycotaxon, 121, 499. [ Links ]
Wang, D. M., Wu, S. H., Su, C. H., Peng, J. T., Shih, Y. H., & Chen L. C. (2009). Botanical Studies, 50, 451-458. (Recovered from http://ejournal.sinica.edu.tw/bbas/content/2009/4/Bot504-08). [ Links ]
White, T. J., Bruns, T., Lee, S., & Taylor, J. W. (1990). Amplification and direct sequencing of ribosomal RNA genes for phylogenetics. In Innis, M. A., Gelfand, D. H., Sninsky, J. J., & White, T. J. PCR Protocols, a Guide to Methods and Applications. Academic Press, New York, 315-322. [ Links ]
Welti, S., & Courtecuisse, R. (2010). The Ganodermataceae in the French West Indies (Guadeloupe and Martinique). Fungal Diversity, 43, 103-126. (Recovered from DOI:10.1007/s13225-010-0036-2). [ Links ]
Yang, Z. L., & Feng, B. (2013). What is the Chinese “Lingzhi”? - A taxonomic mini-review. Mycology, 4(1), 1-4. (Recovered from DOI:10.1080/21501203.2013.774299). [ Links ]
Zakaria, L., Ali, N. S., Salleh B., & Zakaria, M. (2009). Molecular analysis of Ganoderma species from different hosts in Peninsula Malaysia. Journal of biological Sciences, 9(1), 12-20. (Recovered from: DOI:10.3923/jbs.2009.12.20). [ Links ]
Zhou, X., Lin, J., Yin, Y., Zhao, J., Sun, X., & Tang, K. (2007). Ganodermataceae: natural products and their related pharmacological functions. American Journal of Chinese Medicine, 35(4), 559-574. (Recovered from DOI: 10.1142/S0192415X07005065). [ Links ]
1.Departamento de Micologia, Programa de Pós-Graduação em Biologia de Fungos, Universidade Federal de Pernambuco, Av. Nelson Chaves s/n, CEP 50760-420, Recife, PE, Brazil; nelsonradar2005@hotmail.com
2.Departamento de Micologia, Programa de Pós-Graduação em Biologia de Fungos, Universidade Federal de Pernambuco, Av. Nelson Chaves s/n, CEP 50760-420, Recife, PE, Brazil;tatiana.gibertoni@pq.cnpq.br
3.Departamento de Micologia, Programa de Pós-Graduação em Biologia de Fungos, Universidade Federal de Pernambuco, Av. Nelson Chaves s/n, CEP 50760-420, Recife, PE, Brazil;elaine.malosso@ufpe.br
Received 22-XI-2013. Corrected 25-II-2014. Accepted 27-III-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}