










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.3 San José Jul./Sep. 2014
Seed rain under native and non-native tree species in the Cabo Rojo National Wildlife Refuge, Puerto Rico
Lluvia de semillas bajo especies de árboles nativos y no-nativos en el Refugio de Vida Silvestre de Cabo Rojo, Puerto Rico
Lluvia de semillas bajo especies de árboles nativos y no-nativos en el Refugio de Vida Silvestre de Cabo Rojo, Puerto Rico
Abstract
Seed dispersal is a fundamental process in plant ecology and is of critical importance for the restoration of tropical communities. The lands of the Cabo Rojo National Wildlife Refuge (CRNWR), formerly under agriculture, were abandoned in the 1970s and colonized mainly by non-native tree species of degraded pastures. Here we described the seed rain under the most common native and non-native trees in the refuge in an attempt to determine if focal tree geographic origin (native versus non-native) influences seed dispersal. For this, seed rain was sampled for one year under the canopies of four native and four non-native tree species common in this refuge using 40 seed traps. No significant differences were found for the abundance of seeds, or their diversity, dispersing under native versus non-native focal tree species, nor under the different tree species. A significantly different seed species composition was observed reaching native versus non-native focal species. However, this last result could be more easily explained as a function of distance of the closest adults of the two most abundantly dispersed plant species to the seed traps than as a function of the geographic origin of the focal species. We suggest to continue the practice of planting native tree species, not only as a way to restore the community to a condition similar to the original one, but also to reduce the distances needed for effective dispersal. Rev. Biol. Trop. 62 (3): 1129-1136. Epub 2014 September 01.
Key words: seed dispersal, non-native, restoration, dispersal limitation, tropical dry forest.
Resumen
La dispersión de semillas es fundamental para la restauración de comunidades tropicales. Los terrenos del Refugio de Vida Silvestre de Cabo Rojo, anteriormente cultivado, fueron abandonados en los ‘70 y colonizados principalmente por árboles no-nativos. Describimos la lluvia de semillas debajo de las cuatro especies más comunes de árboles nativos y de las cuatro especies de árboles no-nativos de este refugio para evaluar si la procedencia del árbol influye en la dispersión de las semillas. Se recogieron semillas en 40 trampas de semillas; debajo de cinco individuos de cada especie. No encontramos diferencias significativas en abundancia de semillas o en su diversidad, tanto al comparar nativos versus no-nativos, como al comparar entre especies de árboles. Por otro lado, si se presentaron diferencias en la composición de semillas dispersadas debajo de nativos versus debajo de no-nativos. Sin embargo, encontramos evidencia de que ese último resultado pudo deberse más bien a la cercanía del adulto más cercano de la especie dispersada que a la procedencia de la especie de árbol estudiado. Sugerimos continuar la práctica de sembrar árboles nativos, tanto para restaurar la comunidad a una condición más similar a la original como para disminuir las distancias necesarias para una dispersión más efectiva.
Palabras clave: dispersión de semillas, no autóctona, restauración, limitación de dispersión, bosque tropical seco.
Seed dispersal is usually the premier demographic process allowing plants to find adequate sites for their recruitment and eventual reproduction (Nathan & Muller-Landau, 2000, Wenny, 2001) and for avoiding the usually damaging effects of landing in the neighborhood of their parental plants (Howe & Smallwood, 1982). Seed dispersal has been extensively studied in tropical moist and wet ecosystems (Holl, Loik, Lin, & Samuels, 2000), but relatively little is known about this process in tropical and subtropical dry forests (Bullock & Solis-Magallanes, 1990, Campbell, Lynam, & Hatto, 1990, Ceccon & Hernandez, 2009, Vieira & Scariot, 2006) even though these ecosystem types occupy approximately 40% of the tropics and subtropics (Murphy & Lugo, 1986). Recent studies have highlighted the importance of seed dispersal for the recolonization of the original plant communities in abandoned tropical agricultural lands (see review by Holl et al., 2000).
The Subtropical Dry Forest lifezone occupies 14% of the area of Puerto Rico (Ewel & Whitmore, 1973). The Cabo Rojo National Wildlife Refuge (CRNWR), located within the Subtropical Dry Forest, was established in an area with a long history of agricultural disturbance (mostly for grazing) but later abandoned in the 1970’s. Since its abandonment a mixture of native and non-native plant species have colonized the area (Zuill, 1985). Refuge managers and conservationists are concerned about the dominance by the non-native component of this colonizing vegetation.
Non-native plant species have been long considered a threat to native biota (Heywood, 1989). Some of these species have been implicated in modifying their newly colonized habitats in respect to ecosystem properties like hydrology, nutrient dynamics, and disturbance regimes, or by directly competing with the native species. Although several studies have documented that native species regenerate under the canopy of non-native species (see review by Parrotta, Turnbull & Jones, 1997) we have not found any studies testing the relative influence of native and non-native focal species on seed dispersal. This study intends to describe the seed rain arriving under a range of native and non-native trees in the CRNWR. More specifically, this study addressed the following questions: (1) does seed rain abundance, diversity and composition differ under native and non-native trees?, and (2) do the different focal tree species differentially influence the seed rain?
Materials and methods
Study area: The study area is the Eastern portion of the CRNWR. It has an area of 238ha and its geographic center is located at latitude 17°58’35” N-67°10’10” W, in the municipality of Cabo Rojo, Puerto Rico. Mean annual precipitation is 908mm and mean annual temperature is 26°C (NOAA, 2002). Ewel & Whitmore (1973) classified this area as Subtropical Dry Forest.
For about two centuries prior to 1967 these lands were used for grazing and agriculture. In 1967 the United States established a station of the Foreign Broadcast Information Service at the site. Even so, grazing continued until 1978 when a refuge manager was contracted for the area, four years after the Foreign Broadcast Service ended its activities and the lands were transferred to the US Fish and Wildlife Service. Since 1978 the existing vegetation has been developing with a strong component of non-native species. Refuge managers have engaged in interplanting native species in an attempt to restore the vegetation to a more natural character but without removing most of the existing vegetation.
Data collection: In order to sample the seed rain under trees, traps were constructed with a funnel-shaped form made of a plastic mesh with holes of approximately 1mm wide. These were attached to square PVC frames (0.5m x 0.5m); held at 0.3 to 0.4m above the ground.
Four to six trees were sought of each of eight tree species: four native species (Bucida buceras, Guaiacum officinale, Hymenaea courbaril, and Guazuma ulmifolia) and four non-native species (Melicoccus bijugatus, Pithecellobium dulce, Prosopis pallida, and Tamarindus indica). The 8 species are zoochorous. Selected individual trees had trunks of diameters larger than 10cm and were located near trails to facilitate removing seeds from all traps in one day. Although we understand that several tree characteristics, like tree height and crown width, may influence dispersal, we did not seek to control for such characteristics; nevertheless, the variability we observed in this respect did not seem of concern.
One trap was placed under each tree at approximately halfway between the trunk and the edge of the crown on the side of the tree where its crown seemed widest. Seeds were collected during one full year (February 2004 to January 2005). Traps were visited weekly during the first month and at variable frequency from then on; less frequently when fewer seeds were collected and more frequently when more seeds were collected.
Trapped material was processed at the laboratory. Non-seed material was discarded. Seeds belonging to the species of the focal tree were discarded, as it was too uncertain if these seeds were dispersed from other individuals or just fell from the focal tree. All other apparently viable seeds were then processed for identification by comparison to seeds found still attached to their corresponding plants nearby, by comparison to published drawings, or by comparison to material found at the Herbarium of the Mayaguez Campus of the University of Puerto Rico (MAPR). All other seeds that could not be identified by the above procedures were assigned to different morphospecies and sent to experts at the Herbario Nacional de Mexico and to the Smithsonian Institution in Washington, DC. Nomenclature follows Liogier and Martorell (2002). Collected seeds were deposited at MAPR for further reference (barcode numers MAPR36868-MAPR36896).
Infostat (2005) was used for univariate and bivariate analyses; PCORD (McCune & Mefford, 1999) was used for multivariate analyses. Shapiro-Wilks test was used to determine normality for dispersed seed abundance; because of lack of normality these data were square-root transformed (v(y+0.5)) prior to statistical analyses. Significance was tested at a=0.05 level.
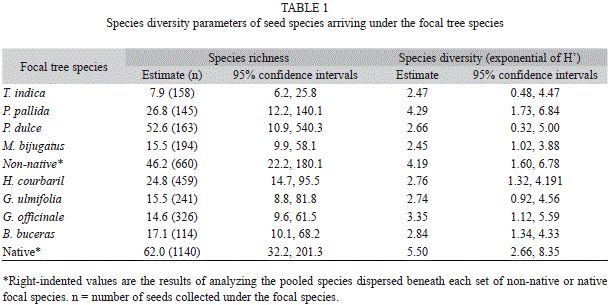
A nested ANOVA was used to examine differences in mean numbers of dispersed seeds over the course of the sampled year between native and non-native focal species (groups) and among the different focal tree species (subgroups). Significance for differences in species richness and species diversity (i.e., exponent of Shannon’s H’, also referred to as effective number of species sensu Jost (2006) were examined by parameter estimation using program SPADE (Chao & Cheng, 2003).
Compositional affinities were evaluated with a Flexible Beta cluster analysis using the Bray-Curtis distance on the square-root transformed abundance data and beta=-0.25. McCune and Grace (2002) recommend the Flexible Beta algorithm with beta set to -0.25 because it is a space conserving linkage method and is compatible with the Bray-Curtis distance (a distance measure appropriate for species by samples matrices). Dispersed species occurring in fewer than two traps were considered rare and removed from the species by trap matrix prior to the cluster analysis to avoid the tendency of rare species to have excessive influence on the results (McCune & Grace, 2002).
After obtaining the results of the above analyses traps were visited to explore the possible effect of dispersal distance on the two species with the highest influence in the cluster analysis results (Pilosocereus royenii and Leucaena leucocephala). Twenty-eight of the 40 traps were visited (not all traps could be relocated because of vandalism) to measure the distances from the traps to the closest mature individual of the two above mentioned species; plant maturity was determined as those with at least the size of the smallest individual found with fruits. We sought for mature adults not farther than 15m from the traps because of the difficulty in observing true absences farther away.
Results
A total of 1 800 seeds belonging to 29 morphospecies were collected in the traps; 99 seeds belonging to eight morphospecies could not be identified to genus or species. The families best represented, with 93% of the collected seeds, were: Leguminosae, Cactaceae, Myrtaceae, and Oleaceae. It is worth mentioning that the Myrtaceae seeds (356), belonging to just one species (Psidium guajava), were collected in only one dispersal event, seemingly the result of defecation by a feral monkey. The family represented by the most species was the Leguminosae with four species.
Although 55% more seeds arrived under native versus non-native focal trees such difference was not statistically significant (F=1.65, df=1.6, p=0.246). No statistically significant differences were found for comparisons of abundance of seeds dispersed under the different species of focal trees either (F=0.54, df=6.32, p=0.775).
No statistically significant difference was found in paired t tests for mean number of native versus non-native seeds arriving under either native (n=20, p=0.130) or non-native focal tree species (n=20, p=0.438). Frequently dispersed species of seeds with the largest numbers of seeds dispersed under native focal trees were the non-native L. leucocephala (426) and the native P. royenii (127), while P. royenii (359) and the non-native Jasminum fluminense (157) arrived in largest numbers under non-native focal trees.
Among the identified species of seeds, 17 species were collected under native focal tree species (8 natives and 9 non-natives) while 14 species were collected under non-native focal tree species (6 natives and 8 non-natives); 21 species were dispersed under both native and non-native focal species. However, there were no statistical differences for species richness or species diversity of seeds dispersed under native versus non-native focal tree species or among species of focal trees (Table 1).
The flexible beta classification (Fig. 1) returned two groups of traps with clear affinities for native versus non-native focal trees. The first group of traps consisted of 13 traps, 11 of which were under native focal trees, while the other group was composed of 23 traps, 16 of which were under non-native focal trees. An Indicator Species Analysis determined that the non-native L. leucocephala was dispersed in significantly larger numbers and frequency under native focal trees (p<0.001), while native P. royenii (p<0.001), and the non-natives J. fluminense (p<0.05), and P. dulce (p<0.05) were dispersed in significantly larger numbers and frequency under non-native focal trees.
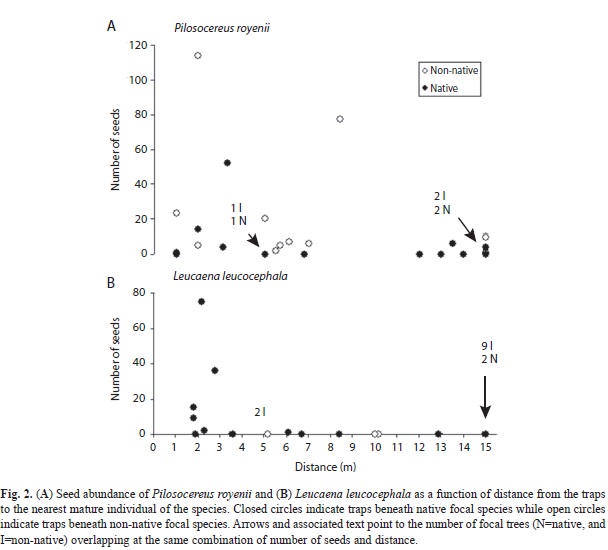
Among the above mentioned indicator species, L. leucocephala and P. royenii were the most abundantly and widely dispersed in this study; we looked further into the possible patterns of dispersal of these species by measuring the distances between each trap and their closest mature individual. Figure 2A and figure 2B suggest the expected negative association between distance to mature individuals and number of dispersed seeds, but they also show a tendency for traps that received more seeds of P. royenii to be under non-native focal tree species, while the traps that received more seeds of L. leucocephala tended to be under native focal tree species.
Discussion
The above results indicated that neither the geographical origin of the focal species or the taxonomic identity of the focal species influenced the measured dispersal parameters. These results suggest as well, that the dispersal vectors of the species found in the traps are not influenced by the identity or the geographical origin of the focal species below which the traps were situated. Vectors other than animals (e.g., wind, gravity) can not be expected to be influenced by the geographical origin of the focal tree species or by their taxonomic identities, but our data suggest that animal vectors were not influenced either by focal species identity or geographical origin.
Animal vectors may visit trees for a number of purposes: feeding, resting, sexual display, or roosting (Cardoso da Silva, Uhl, & Murray, 1996). During any of these activities fruits or seeds consumed elsewhere may be dropped below the visited trees. Our results suggest that it is equally likely that an animal vector will visit native and non-native focal trees for any or all of such activities. It is possible that if there was ever any reason to discourage these animal vectors from visiting these non-native focal species such reasons do not exist anymore.
The only dispersal parameter significantly associated with the geographical origin of the focal species was dispersed seed species composition. However, this result was influenced mostly by a handful of species. Among the two most abundant of those species, P. royenii (a zoochorous native columnar cactus) was selectively dispersed to a non-native focal species while L. leucocephala (a non-native legume small tree) was being selectively dispersed to native focal species. This apparent paradox can be more easily explained by the tendency, probably accidental, for mature individuals of those species to be closer to focal trees of a different geographical origin. Therefore, geographical origin of the focal species may not be the factor influencing such compositional pattern.
P. royenii is a fleshy fruited species likely to be dispersed by birds and other vertebrates. However, as is usually the case, most seeds of plants exhibiting any dispersal syndrome are dispersed near the parent plants (Clark, Macklin & Wood, 1998). Therefore, if mature individuals of P. royenii tended to be closer to traps under non-native focal species it was likely to be selectively found in traps of such geographic origin. Alternatively, it is quite common to find P. royenii in the shade of P. pallida trees throughout the Subtropical Dry Forest lifezone in Puerto Rico (personal observation) suggesting that P. pallida may serve as a nurse plant for P. royenii. If that is the case in CRNWR then mature individuals of P. royenii are more likely to be found close to that non-native focal species in this study.
L. leucocephala is a dry fruited species, being dispersed by physical mechanisms. Its seeds or pods can be transported by winds (Parrotta, 2000) but also through an explosive mechanism as the pod dries quickly under the appropriate weather conditions. Thus, we would not expect it to show differences in dispersal to traps under native versus non-native trees. The observed pattern can be explained by the fact that mature individuals of this species happened to be located closer to traps placed beneath the native trees chosen for this study than to those beneath the non-native trees, thus resulting in their closer association to species of such geographical origin.
The observations on the two species mentioned above suggest that they have short dispersal kernels and that they may be dispersal limited; more detailed studies can provide conclusive evidence on this.
The results of this study also have implications for management of the vegetation of the CRNWR. The geographical origin of the species in this study has no clear influence on the seed rain parameters studied here. On the contrary, evidence (although probably spurious) was found for the spatial association between non-native focal trees and the seed rain of the native P. royenii. Such association may be mediated by the zoochorous nature of the focal species which may attract dispersers carrying seeds of natives (Parrotta et al., 1997). Therefore, removing these non-native tree species may have undesirable results for the dispersal of at least that native species. We admit that we do not have conclusive evidence for such undesirable result, therefore we suggest following Zavaleta, Hobbs and Mooney (2001) advice to do a pre-eradication assessment before attempting removal of the non-native trees studied here.
Moreover, the apparent dispersal limitation of at least two of the most abundantly dispersed species in this study suggests that the current practice of planting native species throughout the CRNWR is an appropriate restoration practice. The more seed sources of native species there are the more likely the vegetation will be restored to the species composition originally present in this area. Moreover, it is of great importance to continue such planting practices considering that the most abundant tree of the refuge (P. pallida), shows little further regeneration on its own in the refuge, probably owing to the fact that this species is shade intolerant (Skolmen, 1990). If P. pallida actually disappears naturally from this refuge the planted natives will likely replace P. pallida.
Acknowledgments
The CRNWR provided materials, labor, and logistic support for this study. We also thank Joseph Schwagerl, Carlos Betancourt, and several students of the second author’s Plant Ecology class for their assistance in the field.
References
Bullock, S. H. & Solis-Magallanes, J. A. (1990). Phenology of canopy trees of a tropical deciduous forest in Mexico. Biotropica, 22, 22-35. [ Links ]
Campbell, B., Lynam, T., & Hatto, J. C. (1990). Small-scale in the recruitment of forest species during succession in tropical dry forest, Mozambique. Vegetatio, 87, 51-57. [ Links ]
Cardoso da Silva, J. M., Uhl, C., & Murray, G. (1996). Plant succession, landscape management, and the ecology of frugivorous birds in abandoned Amazonian pastures. Conservation Biology, 10, 491-503. [ Links ]
Ceccon, E. & Hernández, P. (2009). Seed rain dynamics following disturbance exclusion in a secondary tropical dry forest in Morelos, Mexico. Revista de Biología Tropical, 57, 257-269. [ Links ]
Chao, A. & Shen, T. J. (2003) Program SPADE (Species Prediction And Diversity Estimation). Institute of Statistics, National Tsing Hua University, Hsin-Chu, Taiwan. Recovered from chao.stat.nthu.edu.tw. [ Links ]
Clark, J. S., Macklin, E., & Wood, L. (1998). Stages and spatial scales of recruitment limitation in southern appalachian forests. Ecological Monographs, 68, 213-235. [ Links ]
Ewel, J. J. & Whitmore, J. L. (1973). The ecological life zones of Puerto Rico and the U.S Virgin Islands. Río Piedras, Puerto Rico: Institute of Tropical Forestry [ Links ]
Heywood, V. H. (1989). Patterns, extents and modes of invasions by terrestrial plants. In J. A. Drake, H. A. Mooney, F. di Castri, R. H. Grooves, F. J. Kruger, M. Rejmanek, & M. Williamson (Eds.), Biological invasions: a global perspective (pp. 31-60). John Wiley: Chichester, UK. [ Links ]
Holl, K. D., Loik, M. E., Lin, E. H. V., & Samuels, I. A. (2000). Tropical montane forest restoration in Costa Rica: overcoming barriers to dispersal and establishment. Restoration Ecology, 8, 339-349. [ Links ]
Howe, H. F. & Smallwood, J. (1982). Ecology of seed dispersal. Annual Reviews of Ecology and Systematics, 13, 201-228. [ Links ]
InfoStat (2005). InfoStat versión 2005. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Córdoba, Argentina. [ Links ]
Jost, L. (2006). Entropy and diversity. Oikos, 113, 363-375. [ Links ]
Liogier, H. A. & Martorell, L. F. (2002). Flora of Puerto Rico and adjacent islands: a systematic synopsis. Río Piedras, Puerto Rico: Editorial de la Universidad de Puerto Rico. [ Links ]
McCune, B. & Grace, J. B. (2002). Analysis of ecological communities. Oregon, USA: MJM. [ Links ]
McCune, B. & Mefford, M. J. (1999). PC-ORD. Multivariate Analysis of Ecological Data. Version 4.34. MjM Software, Gleneden Beach, Oregon, U.S.A. [ Links ]
Murphy, P. G. & Lugo, A. E. (1986). Ecology of tropical dry forest. Annual Reviews of Ecology and Systematics,17, 67-88. [ Links ]
Nathan, R. & Muller-Landau, H. C. (2000). Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends in Ecology & Evolution, 15, 278-285. [ Links ]
NOAA (2002). Divisional Normals and Standard Deviations of Temperature, Precipitation, and Heating and Cooling Degree Days 1971 – 2000 (and previous normals periods). US Department of Commerce. National Oceanic and Atmospheric Administration. Climatography of the United States No 85. [ Links ]
Parrotta, J. A. (2000). Leucaena leucocephalla. In J. K. Francis & C. A. Lowe (Eds.). Silvics of native and exotic trees of Puerto Rico and the Caribbean Islands. USA: Department of Agriculture, Forest Service International Institute of Tropical Forestry. [ Links ]
Parrotta, J. A., Turnbull, J. W., & Jones, N. (1997). Catalyzing native forest regeneration on degraded tropical lands. Forest Ecology & Management, 99, 1-7. [ Links ]
Skolmen, R. G. (1990). Prosopis pallida. In R. M. Burns & B. H. Honkala (Eds.), Silvics of North America, Volumen 2. Hardwoods. Washintong DC, USA.: Department of Agriculture, Forest Service. [ Links ]
Vieira, D. L. M. & Scariot, A. (2006). Principles of natural regeneration of tropical dry forests for restoration. Restoration Ecology, 14, 11-20. [ Links ]
Wenny, D. G. (2001). Advantages of seed dispersal: a re-evaluation of directed dispersal. Evolutionary Ecological Research, 3, 51-74. [ Links ]
Zavaleta, E. S., Hobbs, R. J., & Mooney, H. A. (2001). Viewing invasive species removal in a whole-ecosystem context. Trends in Ecology & Evolution, 16, 454-459. [ Links ]
Zuill, H. (1985). The trees of a selected area of the national wildlife refuge, Cabo Rojo, Puerto Rico: structure and composition. Boquerón, PR: U.S. Fish and Wildlife Service. [ Links ]
Campbell, B., Lynam, T., & Hatto, J. C. (1990). Small-scale in the recruitment of forest species during succession in tropical dry forest, Mozambique. Vegetatio, 87, 51-57. [ Links ]
Cardoso da Silva, J. M., Uhl, C., & Murray, G. (1996). Plant succession, landscape management, and the ecology of frugivorous birds in abandoned Amazonian pastures. Conservation Biology, 10, 491-503. [ Links ]
Ceccon, E. & Hernández, P. (2009). Seed rain dynamics following disturbance exclusion in a secondary tropical dry forest in Morelos, Mexico. Revista de Biología Tropical, 57, 257-269. [ Links ]
Chao, A. & Shen, T. J. (2003) Program SPADE (Species Prediction And Diversity Estimation). Institute of Statistics, National Tsing Hua University, Hsin-Chu, Taiwan. Recovered from chao.stat.nthu.edu.tw. [ Links ]
Clark, J. S., Macklin, E., & Wood, L. (1998). Stages and spatial scales of recruitment limitation in southern appalachian forests. Ecological Monographs, 68, 213-235. [ Links ]
Ewel, J. J. & Whitmore, J. L. (1973). The ecological life zones of Puerto Rico and the U.S Virgin Islands. Río Piedras, Puerto Rico: Institute of Tropical Forestry [ Links ]
Heywood, V. H. (1989). Patterns, extents and modes of invasions by terrestrial plants. In J. A. Drake, H. A. Mooney, F. di Castri, R. H. Grooves, F. J. Kruger, M. Rejmanek, & M. Williamson (Eds.), Biological invasions: a global perspective (pp. 31-60). John Wiley: Chichester, UK. [ Links ]
Holl, K. D., Loik, M. E., Lin, E. H. V., & Samuels, I. A. (2000). Tropical montane forest restoration in Costa Rica: overcoming barriers to dispersal and establishment. Restoration Ecology, 8, 339-349. [ Links ]
Howe, H. F. & Smallwood, J. (1982). Ecology of seed dispersal. Annual Reviews of Ecology and Systematics, 13, 201-228. [ Links ]
InfoStat (2005). InfoStat versión 2005. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Córdoba, Argentina. [ Links ]
Jost, L. (2006). Entropy and diversity. Oikos, 113, 363-375. [ Links ]
Liogier, H. A. & Martorell, L. F. (2002). Flora of Puerto Rico and adjacent islands: a systematic synopsis. Río Piedras, Puerto Rico: Editorial de la Universidad de Puerto Rico. [ Links ]
McCune, B. & Grace, J. B. (2002). Analysis of ecological communities. Oregon, USA: MJM. [ Links ]
McCune, B. & Mefford, M. J. (1999). PC-ORD. Multivariate Analysis of Ecological Data. Version 4.34. MjM Software, Gleneden Beach, Oregon, U.S.A. [ Links ]
Murphy, P. G. & Lugo, A. E. (1986). Ecology of tropical dry forest. Annual Reviews of Ecology and Systematics,17, 67-88. [ Links ]
Nathan, R. & Muller-Landau, H. C. (2000). Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends in Ecology & Evolution, 15, 278-285. [ Links ]
NOAA (2002). Divisional Normals and Standard Deviations of Temperature, Precipitation, and Heating and Cooling Degree Days 1971 – 2000 (and previous normals periods). US Department of Commerce. National Oceanic and Atmospheric Administration. Climatography of the United States No 85. [ Links ]
Parrotta, J. A. (2000). Leucaena leucocephalla. In J. K. Francis & C. A. Lowe (Eds.). Silvics of native and exotic trees of Puerto Rico and the Caribbean Islands. USA: Department of Agriculture, Forest Service International Institute of Tropical Forestry. [ Links ]
Parrotta, J. A., Turnbull, J. W., & Jones, N. (1997). Catalyzing native forest regeneration on degraded tropical lands. Forest Ecology & Management, 99, 1-7. [ Links ]
Skolmen, R. G. (1990). Prosopis pallida. In R. M. Burns & B. H. Honkala (Eds.), Silvics of North America, Volumen 2. Hardwoods. Washintong DC, USA.: Department of Agriculture, Forest Service. [ Links ]
Vieira, D. L. M. & Scariot, A. (2006). Principles of natural regeneration of tropical dry forests for restoration. Restoration Ecology, 14, 11-20. [ Links ]
Wenny, D. G. (2001). Advantages of seed dispersal: a re-evaluation of directed dispersal. Evolutionary Ecological Research, 3, 51-74. [ Links ]
Zavaleta, E. S., Hobbs, R. J., & Mooney, H. A. (2001). Viewing invasive species removal in a whole-ecosystem context. Trends in Ecology & Evolution, 16, 454-459. [ Links ]
Zuill, H. (1985). The trees of a selected area of the national wildlife refuge, Cabo Rojo, Puerto Rico: structure and composition. Boquerón, PR: U.S. Fish and Wildlife Service. [ Links ]
1. Ponce School of Medicine and Health Sciences, AIDS Research Program, PO Box 7004, Ponce, PR 00732-7004; andrea.ag186@gmail.com
2. Universidad de Puerto Rico, Departamento de Biología, PO Box 9000, Mayagüez, PR 00681-9000. Tel: 787-832-4040-ext 3913; jdchinea@yahoo.com
Received 13-I-2014. Corrected 04-IV-2014. Accepted 05-V-2014.


{kind=link}
{kind=link}
{kind=link}