










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.3 San José Jul./Sep. 2014
Seasonal dynamics of the shoreline vegetation in the Zapatosa floodplain lake complex, Colombia
Dinámica estacional de la vegetación litoral en el complejo lagunar inundable Zapatosa, Colombia
Dinámica estacional de la vegetación litoral en el complejo lagunar inundable Zapatosa, Colombia
Abstract
Floodplain lakes and associated wetlands in tropical dry climates are controlled by pronounced and severe seasonal hydrologic fluctuations. We examined the plant community response to a bimodal flooding pattern in the Zapatosa Floodplain Lake Complex (ZFLC), Northern Colombia. We measured floristic and quantitative change in four sampling periods emphasizing seasonal differences in plant abundance and life-form structure. Of 79 species identified in the lake complex, 52 were used to characterize eight community types via classification and ordination procedures. Results showed that community structure does not change significantly during the flooding/receding stages. But maximum drawdown phase significantly disrupts the aquatic community structure and the exposed shorelines become colonized by ruderal terrestrial plants. Early rainfalls at the beginning of the wet season are emphasized as an important feature of plant regeneration and community development. The general strategy of the ZFLC vegetation can be framed into the flood pulse concept of river-floodplain systems. Thus, plant communities are mainly responding to disturbances and destruction events imposed by extreme water level fluctuations. Rev. Biol. Trop. 62 (3): 1073-1097. Epub 2014 September 01.
Key words: Cesar River, flood pulse, littoral, macrophytes, Magdalena River, shallow lakes, bimodal seasonality.
Resumen
Las ciénagas y humedales asociados a sistemas fluviales en climas tropicales secos son controlados por fluctuaciones hidrológicas estacionales pronunciadas. En este trabajo se examinó la respuesta de la comunidad vegetal a un patrón bimodal de inundaciones en el complejo de ciénagas de Zapatosa (ZFLC), al norte de Colombia. Se estudiaron los cambios florísticos y cuantitativos de la vegetación en cuatro periodos de muestreo destacando diferencias estacionales en las abundancias y la estructura biotipológica de las plantas. De las 79 especies identificadas en el complejo lagunar, 52 se utilizaron para caracterizar ocho tipos de comunidades mediante tácticas multivariadas de clasificación y ordenación. Los resultados revelan que la estructura de las comunidades no cambia de manera significativa durante los periodos de inundación/sequía. Únicamente la época de máxima sequía altera la estructura de las comunidades acuáticas y el litoral expuesto es colonizado por plantas ruderales terrestres. Las precipitaciones iniciales al inicio de la temporada de lluvias se destacan como una característica importante para la regeneración de las plantas y el desarrollo subsiguiente de las comunidades. La estrategia de sobrevivencia general de la vegetación acuática del ZFLC puede enmarcarse en el concepto de pulso de inundación de los sistemas fluviales inundables. Por lo tanto las comunidades vegetales responden principalmente a los disturbios y la destrucción impuestos por las fluctuaciones extremas del nivel del agua.
Palabras clave: Rio Cesar, pulso de inundación, litoral, macrófitos, Rio Magdalena, ciénagas, estacionalidad bimodal.
Floodplains play an important role regarding the biogeochemistry and ecology of tropical river systems. Plant and animal life of this particular system are adapted to the seasonal variations in space and time caused by extreme water level fluctuations. This seasonality also represents an important factor in the speciation of several aquatic and wetland plants (Haynes & Holm-Nielsen, 1989). The dynamic character of these systems gives rise to complex primary and secondary production processes that are difficult to predict in detail, but nutrient input from rivers and streams are important in general, as a significant contribution can be expected following rainfall from the surrounding watershed. Nutrient cycling proceeds mainly through heterotrophic processes via bacteria, fungi, micro-zooplankton and shredding-invertebrates (Melack & Fisher, 1990; Wantzen, Yule, Mathooko, & Pringle, 2008; Warfe et al., 2011). The interactions between rivers and their surrounding floodplain motivated Junk, Bayley, and Sparks (1989) to propose the ‘flood pulse concept’ as a main force controlling the biota of tropical floodplains. They considered the floodplain as the area periodically inundated by the waters of rivers, lakes, lagoons, and/or underground water, or by direct precipitation. The concept aims to explain the physical and chemical conditions that obligate the biota to respond through different phenological, morphological, anatomical and/or physiological adaptations to predictable hydrological cycles. Given its alternation between aquatic and terrestrial phases, the flooded area of the shoreline is defined as ‘aquatic-terrestrial transitional zone’ (ATTZ). Recently the pulse concept was reviewed by Junk and Wantzen (2004), as likewise expanded to temperate rivers (Tockner, Malard, & Ward, 2000) and lakes (Wantzen, Junk, & Rothhaupt, 2008). Another emerging concept related to the flood pulse is the homogenization effect of the inundation process mediated through increasing lateral and longitudinal connectivity. In this case hydrological connectivity, referring here to water mediated transfer of matter, energy, and/or organisms within or between elements of the hydrologic cycle (Pringle, 2001), homogenizes the limnological (physical, chemical) and biological characteristics of contiguous water bodies along river floodplain systems (Thomaz, Bini, & Bozelli, 2007).
The herbaceous vegetation of tropical floodplain systems is very heterogeneous, highly productive and plays an important role in nutrient cycling. For example, in the middle Amazon it occupies approximately 0.5% of the basin but contributes 5% of the total organic matter to the downstream transport (Hedges et al., 1986). Plant communities in tropical seasonally flooded wetlands and floodplain lakes have been thoroughly studied for Northern Australia (Casanova & Brock, 2000; Finlayson, 2005; Warfe et al., 2011), India (Unni, 1971; Middleton, 1999; van der Valk, Middleton, Williams, Mason, & Davis, 1993), and Africa (Rees, 1978; Müller & Deil, 2005). In tropical and subtropical South America studies on the different aspects of the flooding cycles and their effect on aquatic vegetation were carried out in the Orinoco floodplains of Venezuelan (Castroviejo & López, 1985; Rial, 2000; 2006), the Amazon (Junk, 1970; 1986; Junk & Piedade, 1993; 1997; Ferreira, Piedade, Wittmann, & Franco, 2010; Piedade et al., 2010), the Pantanal (Do Prado, Heckman, & Martins, 1994; Frey, 1995; Fortney et al., 2004; Pott & Pott, 2004; Kufner, Scremin-Dias, & Guglieri-Caporal, 2011; Pott, Pott, Lima, Moreira, & Oliveira, 2011), coastal Brazil (Rolon, Lacerda, Maltchik, & Guadagnin, 2008), and particularly the Paraná due to increasing river impoundments (e.g., Franceschi, Torres, Prado, & Lewis, 2000; Murphy et al., 2003; Thomaz, Souza, & Bini, 2003; Santos & Thomaz, 2007; Sabattini & Lallana, 2008; Santos & Thomaz, 2008; Padial et al., 2009; Thomaz, Carvalho, Padial, & Kobayashi, 2009; Sousa, Thomaz, & Murphy, 2011).
Additionally, aquatic and wetland vegetation of tropical floodplains are considered of prime importance for a variety of biota. For instance, macroinvertebrates living on macrophytes and in the root zones of floating mats are using this habitat for shelter and food resource, serving also as primary food supply for fish and other vertebrates (primarily amphibians, reptilians and birds; Junk, 1973; Poi de Neiff & Carignan, 1997; Poi de Neiff, 2003; Takeda, Souza-Franco, Melo, & Monkolski, 2003). Furthermore, aquatic vegetation represents a key factor in the selection of habitats by fish (Chick & McIvor, 1997; Gomes, Bulla, Agostinho, Vasconcelos, & Miranda, 2012), serving as refuge to avoid predators (Chick & McIvor, 1997; Warfe & Barmuta, 2004), as spawning habitat, and as nurseries for initial life stages and juveniles (Grenouillet & Pond, 2001; Bulla, Gomes, Miranda, & Agostinho, 2011).
The ‘Zapatosa Floodplain Lake Complex’ (ZFLC) and its associated wetlands, part of an even larger floodplain system called the Mompox Depression, is considered one of the most important freshwater ecosystems of the Magdalena River basin in Northern Colombia. In spite of its importance, hydrobiological information on the river basin is practically inexistent, dealing mostly with the local inventory and ecology of fish assemblages (Ríos-Pulgarín, Jiménez-Segura, Palacio, & Ramírez Restrepo, 2008), aquatic and wetland plants (Schmidt-Mumm, 1987), and a generalized overview on macrophyte vegetation (Rangel-Ch., 2010). In the Magdalena River fish stocks have collapsed due to overexploitation and basin mismanagement, and the relationship between fish migration and the bimodal flood pattern of the river is very poorly understood, following Galvis and Mojica (2007) who also highlighted the urgent need to study the floristic composition and ecology of the floating mats and inundated forests for better understanding fish migration dynamics.
In this study, we aimed on identifying the influence of the distinct bimodal seasonal flooding and drought cycle spanning over one year. Using transects, we examined the shoreline of the ZFLC with emphasis on seasonal differences in macrophyte abundances and life-form structure. The specific objectives of this study were to: (1) compare and characterize the composition and community structure of the littoral herbaceous vegetation between four ecologically distinct dry/wet flooding phases within the ZFLC; (2) determine the existence of a zonation pattern in the plant community composition along the spatial inundation gradient; and (3) provide baseline data for management that can be used to evaluate long-term floristic and structural changes in the aquatic vegetation. Related to objective (2), we expected that the environmental filters associated with the bimodal flooding and drawdown phases would greatly shape the seasonal plant assemblages, and hypothesize that the inundation event increases the similarity of the vegetation structure with increasing flooding.
Materials adn methods
Study site: The Zapatosa lake complex (8°58’-9°16’ N, 73°40’-73°58’ W), part of the Mompox tectonic depression (~6 500km2) in the Caribbean savannas, represents one of the most important and extensive floodplain systems of Northern Colombia (Fig. 1). Four large rivers flow into the Mompox Depression: the Magdalena River, with an average water discharge of ~4 100m3/s at the depression entrance, Cauca River (~2 300m3/s), San Jorge River (~270m3/s) and Cesar River (~200m3/s). At the exit of the depression, the Magdalena River has an average discharge of ~7 400m3/s. Sediment load along the Magdalena River shows an increasing trend downstream, from 51MT/yr at the upper course to 144MT/yr in the lower reach. The mean specific sediment yield for the whole Magdalena basin is approximately 689±528t/Km2/yr (Restrepo & Syvitski, 2006). In addition, the Magdalena River appears to have the highest sediment yield of the large rivers along the Caribbean and Atlantic coasts of South America. It is noteworthy to mention that the dissolved load is of the same magnitude as the Orinoco River, similar to the Paraná River, and ten times lower than that of the Amazonas River (Restrepo & Kjerfve, 2000; 2004).
The rain pattern and water level fluctuation in the ZFLC follows a bimodal tetra-seasonal regime, although water level rises with a lag of around one month after initiation of the wet season. The first and smaller flood (ca. 2.6m over average water level) occurs between May and July, the second between October and December (ca. 5m over average water level). A lesser drought, the Caribbean mid-summer drought (‘veranillo’) caused by the intensification and expansion of the North Atlantic subtropical high pressure cell (Poveda, Waylen, & Pulwarty, 2006; Gamble, Parnell, & Curtis, 2008), occurs between July and September, while the main dry season takes place between January and April. The Cesar River flows directly into the Zapatosa floodplain lake where it forms an internal delta creating a series of water bodies of different sizes. At the South-Western end, the lake drains into the Magdalena River through the Cesar channel and a series of minor streams, lagoons and wetlands. During high waters, the Magdalena river back-flows into the lake through this channel and adjacent wetlands, contributing to the hydrological cycle to a considerable extent. The lake itself has a mean extension of ~35 000ha, with an increment of ~14 500ha during high waters. Belonging to the Magdalena and Cesar districts, the lake has an altitude of ca. 30m above sea level, mean annual temperatures between 27°C and 30°C, and an annual rainfall that varies from 1 000 to 2 600mm (Rangel-Ch., 2007). Appendix 1, summarizing raw data from Viña et al. (1991), provides some limnological parameters for the four sampling seasons.
Sampling: Vegetation sampling was carried out during four field trips between June 29-July 2 (Jul-1990), October 16-19 (Oct-1990), February 26-March 2 (Feb-1991), and May 20-23 (May-1991). Based on rain fall fluctuations, the four periods were defined as minor dry season or mid-summer drought (Jul-90), mayor wet season (Oct-90), mayor dry season (Feb-91), and minor wet season (May-91). The 74 sample sites where selected during a preceding recognition trip, and locations were chosen in consent on habitat qualities meeting the requirements for different groups of biota (fish, phytoplankton, and macroinvertebrates associated to the root zone of macrophytes). Additionally, shorelines densely cowered by trees or lacking macrophytes due to bank erosion or slumping were excluded from vegetation sampling. Site characteristics representing different habitat conditions are: wave exposed shorelines (sites 2, 4, 11, 17, 18, 20, 21), sheltered shorelines (sites 1, 3, 7, 8, 9, 10, 12, 19, 22), small lagoons (sites 13, 14, 15), and locations subjected to river flow (sites 5, 6, 16). All lakes and lagoons are directly connected to the Cesar River channel (Fig. 1).
Sites 1 to 16 were sampled by means of transects on a regular basis during all four seasons. We also sampled an additional number of transects to broaden our knowledge on distribution and to complement the vegetation inventory. Therefore, site 17 was additionally sampled during Oct-90, Feb-91 and May-91; sites 18, 21 and 22 during Oct-90; sites 19 and 20 during Oct-90 and May-91 respectively (Fig. 1). Similar to the sampling design applied by Schmidt-Mumm and Vargas (2012), transects were placed along the inundation gradient. The lengths of transects varied from 30 to 40m for the July-90 sampling, but were maintained constant at 30m for the remaining seasons. Each transect consisted of 1m2 quadrats laid out contiguously on the aquatic-terrestrial gradient and then further sub-sampled by four 0.25m2 plots. A total of 2 279 quadrats were sampled during the four seasons. Using a canoe to access the aquatic vegetation, ‘rooted frequency’ (Greig-Smith, 1983) of plants (presence/absence) was recorded for each subplot. Voucher specimens of almost all the plant collections made in the floodplain lakes were deposited at the Herbario Nacional Colombiano (COL) and Herbario Pontificia Universidad Javeriana (HPUJ). We thereon made identification of specimens in the field, except for plants not previously collected and those that required further confirmation in the laboratory. Two species of Lemna where found in the study area (Landolt & Schmidt-Mumm, 2009), however due to difficulties in distinguishing Lemna aequinoctialis from L. valdiviana in the field, we quantified booth species as Lemna spp.
Life forms: Life-form and growth form are two complementary concepts that first classify aquatic vegetation into a number of broad categories dependent on the relation of the plants to substratum and water level, and then they are grouped according to comparable structure and similar relations to the physical environment. Here we adapted a simplified scheme of Sculthorpe (1985) and Hutchinson (1975). The percent frequency (%) was used to compare the following life form categories: (1) free floating (includes the acropleustophytes and mesopleustophytes), (2) rooted floating-leafed (ephydates), (3) rooted submerged (hyphydates), (4) rooted floating (floating-helophytes; creeping plants and trailing vines, also the reptohelophytes of Wiegleb (1991), (5) rooted emergent (erect-helophytes; plants erect or decumbent), and (6) ‘terrestrials’ which include all those plants growing on the exposed shore-line during the dry season.
Classification of the shoreline vegetation was completed in three steps following the approach of Jensén and van der Maarel (1980) and Schmidt-Mumm and Vargas (2012):
1.Primary classification of the quadrats per transects in order to find local plant groups characteristic for each transect (local zonation patterns). Here we classified quadrats into ‘transect groups’ for each transect.
2.Secondary classification of the groups obtained in the primary classification for each sampling period. The aim is to find clusters of groups that characterize the overall zonation pattern of the particular sampling period. Here we classified the ‘transect groups’ into clusters for each sampling period independently.
3.Final classification of the clusters from the independent sampling periods into generalized ‘community types’.
For the primary and secondary classification we performed a ‘Two-Way Indicator Species Analysis’ (TWINSPAN) procedure on PC-ORD (McCune & Mefford, 2011). In both cases pseudospecies cut levels 0, 2, 5, 10 and 20 were used. One of the problems with TWINSPAN classification is the assumption of a strong gradient that dominates the data structure and the failure to identify secondary gradients (van Groenewoud, 1992; Belbin & McDonald, 1993). However, for the present case we expected that each transect reflects the strong underlying aquatic-terrestrial gradient and that each group represents a portion of the local zonation pattern on the particular transitional zone. The same strong underlying gradient for the ‘clusters of transect groups’ was also assumed in the secondary classification. The primary classification is based on raw data, here represented by the presence of species in the sub-samples in each 1m2 quadrat. For the secondary classification we also used raw data summing the number of sub-samples for each species in the respective group. Species occurring in less than 4% of the groups were excluded from the analysis.
For the final classification in the resulting clusters a percent frequency (%) of species was calculated as a ratio of the number of occupied sub-samples and the total number of sub-samples in the cluster. However, since more than one strong underlying gradient is assumed, final classification was performed by flexible clustering (ß=-0.25) and the Bray-Curtis distance measure using PC-ORD (McCune & Mefford, 2011). Species in less than 5% occurrences where excluded from the species by community matrix. Indicator Species Analysis (ISA) was applied to each cluster level and the lowest average p-value was used as guideline to prune the dendrogram at the most informative level (Dufrêne & Legendre, 1997; McCune & Grace, 2002). Final decision of the cutting level was also based on the ordination output. The resulting clusters are then summarized by calculating a synoptic ‘mean percent frequency’ for each species in each of the final clusters. These clusters are here considered as final ‘Community Types’.
ISA was also used to determine indicator species that characterize the final groups. The Indicator Value method combines information on the concentration of species abundance in a particular community type and the faithfulness of occurrence of a species in a particular community type. Resulting indicator value for each species in each community type was tested for statistical significance by a Monte Carlo randomization procedure. A total of 1 000 randomized runs were used in PC-ORD to determine the significance (p=0.05) of the indicator values (McCune & Mefford, 2011). By definition, indicator species are considered the most characteristic species of each community type, found mostly in a single community type of the classification and present in the majority of the communities belonging to that community type (Dufrêne & Legendre, 1997; Legendre & Legendre, 2012).
NMDS ordination with Bray-Curtis distance method was used to examine the spatial and temporal variations in the species by ‘Community Type’ data matrix. We selected the autopilot mode with the slow and thorough analysis of PC-ORD (McCune & Mefford, 2011). For the preliminary runs a random starting configuration number and 250 randomized runs was used for Monte Carlo tests. The number of axes and starting configuration resulting from the preliminary analysis was used to rerun NMDS with the option of 250 iterations, 50 runs on real data, and varimax rotation. Tie handling of the distance matrix was not penalized in both cases (autopilot and final run). We used after the fact correlations (r2) between distances in the ordination space and distances in the original space (Bray-Curtis distance) to calculate the proportion of variance represented in the ordination axes (McCune & Grace, 2002).
Starting from the secondary classification, we used the Multi-response Permutation Procedure (MRPP) in program PC-ORD (McCune & Mefford, 2011) to test for differences in the compositional structure between the four sampling periods. MRPP was based on a Bray-Curtis distance matrix and n/sum (n) to weight groups. We rank transformed the distance matrix to make the MRPP results more analogous in theory to NMDS. MRPP provides a permutation p-value for the test of group differences and an A-value for within-group homogeneity (McCune & Grace, 2002).
Results
Floristic composition and life forms: A total of 79 species in 60 genera and 33 families were identified within, and in the surroundings of the ZFLC (see Appendix 2 for a complete species list). Of these 43% were Monocots, 48% Eudicots, 3% unranked Angiosperms (Nymphaeales), and 6% ferns. Most species belong to the family Poaceae and Cyperaceae. Fifty two species (66%) were recorded in 74 transects, and 34 of these were considered as aquatic or wetland plants. Most of the aquatic and wetland plants have a worldwide distribution, 44% of the species are represented on more than one continent, 12% are limited to the American continent, and 44% are restricted to the Neotropical area. Polygonum caucanum is the only species with a distribution restricted to the Magdalena and Cauca valley in Colombia.
Species richness per transect ranged from 7 to 14 for July-90 (average=11 spp.), 3 to 15 for October-90 (average=11 spp.), 6 to 19 for February-91 (average=11 spp.), and 5 to 18 for May-91 (average=11 spp.). Most frequent species are Paspalum repens (95%), Eichhornia crassipes (81%), Ludwigia helminthorrhiza (80%), Neptunia oleracea (73%), Salvinia auriculata (70%), Aeschynomene indica (69%), Pistia stratiotes (62%), and Lemna spp. (59%). Two species, Ipomoea subrevoluta Choisy and Cleome parviflora H.B.K., where reported only once for all the sampling sites. Other rare species not listed in table 1 are Salvinia sprucei Kuhn, Najas arguta H.B.K. and Leersia hexandra Sw.
We classified the 52 species into five major life forms types, resulting in 19% of the plants grouped as free floating (pleustophytes), 2% as rooted submerged (hyphydates), 13% as rooted floating (floating-helophytes), and 33% as rooted emergent (erect-helophytes). The remaining 33% were considered as terrestrials. No rooted floating-leaved plants were recorded for the sample units. Abundances (percent frequency) of free floating plants decreased from Jul-90 to Oct-90 and from Oct-90 to Feb-91, and then increased from Feb-91 to May-91 (Fig. 2). A similar trend was also observed for the rooted floating plants. Rooted emergent plants slightly increased from Jul-90 to Oct-90, strongly decreased from Oct-90 to Feb-91, and then increased from Feb-91 to May-91. Terrestrial plants at the shoreline peaked during Feb-91, later decreasing but not disappearing during May-91 (Fig. 2). Rooted submerged plants were recorded only in two transects with low frequency during Jun-90 and therefore are not presented in figure 2.
Community structure: TWINSPAN classifications, first by transects and then by the resulting groups produced 17 ‘clusters’ for the July 1990 sampling period, 12 for October 1990, 20 for February 1991, and 13 for May 1991. The 52 species by 62 ‘cluster’ data matrix was the starting point for the final classification and ordination analysis. Eight species were found in less than 5% of the clusters and are excluded from further analysis.
Eight ‘Community Types (CT)’ and 26 indicator species were recognized by ISA pruning the cluster dendrogram (Fig. 3, Table 1). Cluster partitioning consistency was confirmed by the ordination analysis (Fig. 4a). The community types are, organized on an aquatic-terrestrial gradient and characterized by the two highest indicator values: I) Salvinia auriculata-Wolffiella lingulata free-floating communities; II) Persicaria hispida-Caperonia castaneifolia herbaceous floating mat; III) Aeschynomene indica-Paspalum repens graminoid floating mat; IV) Vigna dominated graminoid floating mat; V) Brachiaria mutica reed-swamps; VI) Persicaria glabra forb-community; VII) Heliotropium indicum-Ambrosia peruviana episodic forb community; VIII) Mimosa dormiens-Eragrostis sp. episodic forb community.
The NMDS ordination resulted in a relative stable, two dimensional solution, with a high but acceptable final stress of 14.58 and final instability of 0.00. The proportion of variation represented by the two axes was 70%, with 48% on axis one and 22% on axis two, respectively (Fig. 4a). The first NMDS ordination axis, showing a consistent wet to dry spatial and seasonal gradient, places the aquatic communities (CT-I, CT-II, CT-III, and CT-IV) at the left end and the terrestrial dry season shoreline community (CT-VIII) at the right end of the axis. The second axis segregates two types of rooted emergent communities (CT-V and CT-VI), representing the higher end of the aquatic-terrestrial transitional zone. Community type descriptions, based on indicator species and plant strategies (Table 1), follow below.
I. Salvinia auriculata-Wolffiella lingulata free-floating communities: This community type comprises 29% of the total sampled area and occupies the permanently flooded part of the aquatic-terrestrial gradient. Life form structure of the community is characterized by emergent and submerged pleustophytes. Significant indicator species included Salvinia auriculata, Wolffiella lingulata, Pistia stratiotes, Wolffia columbiana, Eichhornia crassipes, and Lemna spp.
II. Persicaria hispida-Caperonia castaneifolia herbaceous floating mat: A total of 2% of the surveyed area belongs to this community and is localized on the seasonally flooded shore zone. Life and growth form structure of this community is shaped by rooted emergent and floating plants. Significant indicator species include Persicaria hispida, Caperonia castaneifolia, and Ludwigia helminthorrhiza.
III. Aeschynomene indica-Paspalum repens graminoid floating mat: Representing the most extensive community type of the floodplain, this floating mat is localized on permanently to seasonally inundated shores and comprises 41% of the sampled area. Life and growth form structure of the community is characterized by rooted floating grasses, and floating compound leaved herbs. Significant indicator species are Aeschynomene indica and Paspalum repens.
IV. Vigna luteola dominated graminoid floating mat: A total of 7% of the total samples belong to this community type. This community frequently invades Eichhornia crassipes free floating mats or Paspalum repens floating mats usually found on semi-permanently to seasonally flooded shores. Community growth form structure is determined by the underlying matrix of free floating herbs or rooted floating grasses that are interwoven by the vine Vigna luteola. The only significant indicator species is Vigna luteola.
V. Brachiaria mutica reed-swamps. This community type comprises 4% of the 2 279 sampled plots and is located on the uppermost part of the seasonally flooded zone on the shoreline. The physiognomy of the community is distinguished by rooted emergent tall grasses capable of forming dense stands. The only significant indicator species is Brachiaria mutica; an invasive grass introduced from Africa for cattle ranching and it is currently well established in wetlands throughout the whole Magdalena valley.
VI. Persicaria glabra forb-community: A total of 2% of the sampled area belongs to this community type. As in the previous community, it is located on the seasonally flooded part of the shore and the physiognomy is characterized by rooted emergent tall herbs. One significant indicator species is defined by Persicaria glabra.
VII. Heliotropium indicum-Ambrosia peruviana episodic forb community: This community type comprises 5% of the sampled sites, and colonizes exposed shores during low water level. Substrate consists mainly of moist mud-covered sandy terrain with a thin layer of litter remaining from the preceding macrophytes vegetation. Significant indicator species include Heliotropium indicum, Ambrosia peruviana, and Physalis angulata.
VIII. Mimosa dormiens-Eragrostis sp. episodic forb community: Situated on exposed shores during low water level, this community type comprised 9% of the total samples. Plants of this community are pioneer herbs or low grasses that usually colonize the upper and dryer sandy part of the shore-line. Significant indicator species include Mimosa dormiens, Eragrostis sp., Portulaca oleracea, Alternanthera caracasana, Eragrostis hypnoides, Heliotropium procumbens, Euphorbia serpens, Evolvulus sp., and Fimbristylis vahlii.
Seasonal variations of community types: The NMDS ordination in figure 4b illustrates the compositional differences of assemblages related to seasonal flooding and drought. Convex hulls for groups of the July 1990, October 1990 and May 1991 sampling periods strongly overlap, indicating small changes in community structure. Corresponding centroids (representing a multi-dimensional average) of the July 1990, October 1990 and May 1991 sampling periods are also close and only the February 1991 sampling period distinctively differs from the remaining centroids. Pairwise comparison of all four sampling periods by the MRPP test confirms significant differences of the February 1991 vegetation with the remaining periods (Table 2). The differences are mainly generated by the presence of the dry season terrestrial shoreline community types (Fig. 4a). However results of MRPP should be interpreted with caution since sampling periods are not completely independent. There is also a varying degree of dependence. While the October 1990 community structure is a direct consequence of the previous (July 1990) vegetation, the February 1991 community structure is not completely determined by the previous sampling period. In this case a significant portion of the communities are represented by terrestrial newcomers invading the exposed shoreline. The same applies to the May 1991 vegetation partially resulting from randomly dispersed seeds. Therefore only the rooting perennial community types are continually present during the whole dry-wet season. Quantitative changes in the structure of the vegetation types through the sampling period are presented in figure 5. They were given in relation to the mean fluctuations of water level (m) and rain fall (mm). The percent frequency illustrates the fractions of the community types present at each survey period.
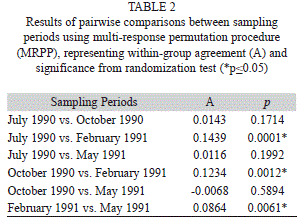
Figure 5 reveals a slight reduction in the abundances of the free floating community (CT-I) from July 1990 to October 1990, followed by a stronger reduction in the abundances from October 1990 to February 1991. After the dry season (February 1991) a gradual recovery of the free floating vegetation could be observed during May 1991. Community Types II and III follow a similar trend. Community Type IV increased from July to October 1990, disappeared during the dry season in February 1991, and reappeared during the filling stage in May 1991. Community Types V and VI displayed similar abundances with small variations during the whole season. Finally the dry season shoreline Community Types VII and VIII reached a peak during February 1991 and gradually disappeared towards May 1991.
Discussion
On a worldwide basis and with approximately 984 species the Neotropics rank first in vascular macrophyte species diversity (Chambers, Lacoul, Murphy, & Thomaz, 2008). The number of aquatic and wetland species recorded in this survey (61 taxa, excluding dry season terrestrials) is relatively low compared to the 120 species reported by Schmidt-Mumm (1987) for the middle Magdalena River basin. Lists of macrophytes have also been compiled for several lowland freshwater environments in South America. For instance, around 54 aquatic and wetland species were reported for the Amazon floodplain near Manaus (Junk & Piedade, 1993), 321 for the coastal area of the State of Rio Grande do Sul (Irgang & Gastal-Junior, 1996), 117 for the Southern Parana State (Cervi, Bona, Moço, & Von Linsingen, 2009), 153 for the Upper Paraná River floodplain (Ferreira, Mormul, Thomaz, Pott, & Pott, 2011), 247 for the Pantanal in Mato Grosso (Pott & Pott, 2011), 279 for the Pantanal (Pott et al., 2011), and 67 for the Bolivian Pantanal (Frey, 1995). There are many differences in these lists as a result of the collecting effort, areas covered, and changes in the taxonomy, making direct comparisons with the ZFLC difficult. Also notable is the rare occurrence of rooted submerged species in the ZFLC. We only found Najas arguta sporadically during the whole sampling season in sheltered places with a high transparency and low wave action. Flood pulses, high sediment load and the dominance of free floating communities and floating mats in the littoral zone can explain the lack of rooted submerged communities in the ZFLC. Similar conditions were also reported for certain floodplain lakes in the San Jorge River of Colombia (Rangel-Ch., 2010), and the Upper Paraná River (Padial et al., 2009; Thomaz et al., 2009). Another life form directly avoiding the ZFLC are the rooted floating leaved plants. Nymphaea species are mostly confined to nearby ponds and marshes that seldom dry out and where wave action is low.
Community structure: In general the vegetation classification summarizes communities representing four sampling periods and a variety of habitat types, ranging from river shorelines, permanent flooded lakes and lagoons to shallow seasonal flooded marshes. All lakes, lagoons and marshes are connected to the Cesar River, thus directly influenced by its flooding regime. The dominant aquatic and wetland vegetation of the ZFLC consists mainly of free floating (Salvinia auriculata-Wolffiella lingulata) and rooted floating communities (Aeschynomene indica-Paspalum repens, Persicaria hispida-Caperonia castaneifolia). The remaining community types are less conspicuous in the floodplain area presenting a scattered distribution throughout the system. With the exception of a preliminary characterization for the San Jorge floodplain lakes (Rangel-Ch., 2010), information on aquatic and wetland vegetation of Colombian lowland floodplain lakes are actually lacking. However, phytosociological classifications using the ‘Zürich-Montpellier’ approach (Kent, 2012) are available and summarized for the Venezuelan Llanos, Pantanal in Bolivia, and Western Paraguay (Castroviejo & López, 1985; Galán de Mera & Navarro, 1992; Galán de Mera & Vicente Orellana, 2006; Galán de Mera & Linares, 2008). A comparison with these schemes is complicated due to the different sampling and classification methods used. On the other hand, Murphy et al. (2003), using the TWINSPAN procedure, recognized the existence of three major aquatic vegetation types for the upper Paraná River. Two communities are dominated by Eichhornia azurea and characterized by the free floating Limnobium laevigatum or Salvinia species, the third one by rooted floating leaved and submerged species determined by Nymphaea amazonum and Myriophyllum aquaticum.
Studies on the existence of zonation patterns along a depth gradient in Neotropic floodplain lakes resulted in different outcomes. For example, Rial (2000) described the macrophyte vegetation of different lakes and streams in the Venezuelan Llanos and concluded that zonation patterns do occur but are strongly influenced by habitat type, local topography and flooding regime. In the Pantanal Do Prado et al. (1994) studied the floristic changes within communities as hydrological conditions change seasonally, and concluded that very distinct species aggregations could occur. However, they found less clearly defined spatial distribution of species along inundation gradients. Also in the Pantanal, Fortney et al. (2004) revealed that zonation can occur in far lakes but not in lakes near the Paraguay River where habitats are subject to strong currents and removal of sediments from substrate. Similar findings are also presented by Padial et al. (2009) for the Paraná River. Here the flood disturbance affected macrophyte species richness, assemblage structure and zonation patterns differently according to the localization of the floodplain lakes within the river sub-systems.
For the ZFLC only a simple and weak zonation pattern can be evidenced along the inundation gradient. The most conspicuous zonation is formed between Salvinia auriculata-Wolffiella lingulata (CT-I) and Aeschynomene indica-Paspalum repens (CT-III). By locally reducing current action and wind-wave disturbances, Paspalum repens floating mats also provide the micro-habitats favorable for the growth of floating plants. In the Cesar River this is particularly notable in sites with slow to moderate flow. Furthermore, during high waters, current stagnates due to the backflow from the Magdalena River, and macrophytes extend locally on the entire river channel (e.g., Eichhornia crassipes, Paspalum repens, Ludwigia helminthorrhiza). Less clearly defined zonation patterns occur when only one community type occupies the shoreline. In this case vegetation stands of Salvinia auriculata-Wolffiella lingulata (CT-I), Persicaria hispida-Caperonia castaneifolia (CT-II), Aeschynomene indica-Paspalum repens (CT-III), or Persicaria glabra (CT-VI) dominate the inundation gradient. On the other hand free floating mats may detach through wind and wave action, temporarily forming zonation patterns with other vegetation types (e.g., Persicaria hispida-Caperonia castaneifolia, Persicaria glabra).
Floating mats have been reported from a number of floodplains around South America: the Middle Paraná River (Tur, 1972), Amazon River (Junk, 1970; 1983) and Brazilian Pantanal (Pivari, Pott, & Pott, 2008). Initial development of floating mats in the ZFLC follows similar patterns as those described for these areas. However, in spite of the presence of mat forming precursor communities like Salvinia auriculata-Wolffiella lingulata (CT-I), Aeschynomene indica-Paspalum repens (CT-III) or Vigna luteola (CT-IV), there is no secondary colonization of floating mats in the ZFLC that progressively leads to the building of floating islands capable of carrying shrubs and trees. Relatively stable water levels and protected shorelines are a main factor which contributes to the development and sustenance of extensive peat forming floating mats (Fortney et al., 2004; Azza, Denny, van de Koppel, & Kansiime, 2006; John, Sylas, Paul, & Unni, 2009). Probably the extensive gentle sloping littoral around the ZFLC and the fact that the surrounding wetlands are flooded by shallow waters, both of which are mostly subjected twice a year to droughts and wetting, explain the absence of peat forming floating islands in the studied area. High temperatures and continually fluctuating water level conditions enhance decomposition processes (Junk & Howard-Williams, 1984; Polunin, 1984; Padial & Thomaz, 2006; Cunha-Santino & Bianchini Jr, 2009), hindering the accumulation of organic layers that lead to the formation of compact floating mats. Burning of nearby grasslands that can extend to the floodplain and cattle ranching during the dry season also destroy part of the vegetation, reducing the accumulation of litter.
Seasonal variation: Flooding pulses are a common feature of many tropical rivers (Junk et al., 1989; Tockner et al., 2000; Junk & Wantzen, 2004) and floodplain lakes (Wantzen et al., 2008). They also homogenize the biological, limnological, physical and chemical characteristics of contiguous water bodies by increasing hydrological connectivity (Junk & Wantzen, 2004; Thomaz et al., 2007). In the case of the ZFLC free floating and rooted floating macrophytes exhibit intense growth during the period of increasing water level and during floods, while gradually decreasing when the water receded from the floodplain. Community structure and species composition did not vary significantly during these phases. But during the dry season, when extensive parts of the shorelines where exposed to drought, the community structure became significantly different from previous phases. Not even the mid-summer drought had as significant a disrupting effect on vegetation structure.
Our results indicated that the homogenizing effect does not increase steadily during flooding, and that community composition is determined from the start of the wet season. Similar results have also been reported by Rial (2006) for the Venezuelan Llanos. In accordance with the findings of Osland, González, and Richardson (2011), we considered that early rainfall periods at the beginning of the wet season initiate seed germination and re-sprouting of plants, therefore setting the community structure of the vegetation that will establish and persist during the following flooding phase. Aquatic and wetland plants surviving the dry season in permanently inundated areas of the lake and in lagoons also contribute to the fast re-colonization of shorelines. Thus the floristic composition of the vegetation is determined from the onset of the wet season and increasing or receding water levels will not substantially change the community structure during the flooding cycle. But maximum low water level during the dry season will reset the scenario for the next vegetation cycle, which is determined by the enduring seed banks and the vegetative reproduction capability of dry-tolerant species. There is also evidence that the distribution of several macrophytes is rearranged during the flooding process (Boschilia, Oliveira, & Thomaz, 2008; Sousa et al., 2011). Further research is required to test this hypothesis as well as to determine the wetting regimes (rain, flooding) that support viable dormant seeds germination and rhizomes sprouting in these systems.
While water fluctuations are certainly a dominant driving force in the dynamics of the ZFLC vegetation, perennial communities of Persicaria hispida-Caperonia castaneifolia (CT-II), Brachiaria mutica (CT-V), and Persicaria glabra (CT-VI) are less affected by these variations. These communities persist during the entire season with few changes in their abundances, thus contributing, in low intensity, to the homogenization process of contiguous water bodies. On the other hand, individual plants or patches of free floating macrophytes are passively spread by wind and currently increasing the similarity within and between aquatic habitats during flooding. Additionally, the success of Eichhornia crassipes in colonizing habitats subjected to wide and erratic fluctuations in water level appears to be related to its ability to modify morphological traits according to the availability of nutrients and water level (Neiff, Casco, & Poi de Neiff, 2008).
Hydrological changes between the flooding and receding waters occur on a temporal continuity following a bimodal pattern for the ZFLC. Based on the existing literature for South American floodplain lakes, very little are known about fluctuations in plant community structure and the bimodal flooding patterns. One can expect differences in seasonal patterns between geographical regions, but further comparative studies are needed in order to increase the knowledge about wetland vegetation and its response to similar flood pulses.
Next we present a summary of the vegetation processes in the ZFLC in order to provide consistent information for the developing of sound management and restoration plans. In this case, we adopted the hydrological pulses defined by Esteves (1998) as dry phase, flooding phase, high water phase, and receding phase.
I. Dry phase: This phase corresponds to the dry season and occurs during December and January to March of each year. As the water recedes, the biomass produced by the aquatic and semi-aquatic vegetation dries and decays. As a result, large amounts of dead biomass accumulate in the exposed sediments and their decomposition contributes to an increase in the nutritional status of the shoreline (Ferreira et al., 2010; Piedade et al., 2010). For most hydrophytes it represents a period of dormancy, where plants survive in the form of seeds, rhizomes and/or desiccation resistant buds. However, areas permanently flooded during the dry season are serving as a shelter for aquatic and wetland plants. Other parts of the dry and exposed shorelines are rapidly colonized by ruderal terrestrial plants, and survival span of this vegetation is limited by the start of the next flooding phase.
The distribution patterns of terrestrial plants on the shoreline are determined by short-term water level fluctuations, which are dictated by temporal rainfalls in the Cesar and Magdalena river catchment area. The expanding and contracting action of the water level on the steeper shorelines can lead to a concentrated seed deposition and germination, locally leading to the formation of distinct vegetation bands. Growth and exuberance of the terrestrial plant communities also relies on the physical and nutrient conditions of the substrate. Thus, exposed and well drained shorelines with gravel to sandy and nutrient poor substrates are generally colonized by Mimosa dormiens-Eragrostis sp. communities (CT-VIII), while fine sediments enriched with detritus or other organic material are usually occupied by the Heliotropium indicum-Ambrosia peruviana communities types (CT-VII).
II. Flooding phase: Persistent rainfalls, which start during March, initiate the wet season, yet water level of the CZFL only begins to raise approximately one month later when excess water of the Magdalena and Cesar Rivers start to flood the basin. Seed germination and vegetative regeneration by rhizomes or stolons of the rooted emergent plants are triggered by early-wet season rainfalls (Osland et al., 2011), leading to a fast re-growth of the mat forming vegetation. Low-lying flats and swales of the floodplain (Junk, 1986) become inundated by runoff water from heavy rains, forming shallow lakes and lagoons where rooted floating plants start to build up dense populations of Paspalum repens, Ludwigia helminthorrhiza, Neptunia oleracea and Aeschynomene indica. Similarly, a fast development of free floating plants consisting of Eichhornia crassipes, Salvinia auriculata, and Pistia stratiotes can be observed in these habitats. It is also during this phase when remaining litter from the aquatic and wetland vegetation of the previous season and decaying terrestrial herbaceous plants starts to decompose in shallow waters, generating a local reduction of O2 and development of H2S. However, with raising water level this condition will persist only for a few weeks. These conditions do not occur at the final phase of the mid-summer drought in August.
III. High water phase: This phase represents the maximum flooding period and occurs between October and November in each year. The previous mid-summer drought during August causes only a minor reduction in the overall vegetation cover. Thereafter the Aeschynomene indica-Paspalum repens community types (CT-III) slightly increases without exceeding the exuberance of the prior flooding phase (May-June). However the open spaces not occupied by graminoid floating mats are filled by free floating communities of the Salvinia auriculata-Wolffiella lingulata (CT-I) or Vigna luteola (CT-IV) type. This observation agrees with the findings of van der Valk (2005), Maltchik, de Oliveira, Rolon, and Stenert (2005) and Maltchik, Rolon, and Schott (2007) that small water-level fluctuations (~50cm) in wetlands may lead to changes in species dominance, moreover if fast clonal growing species are involved. The occasional appearance of the free floating species Salvinia sprucei and Phyllanthus fluitans can also be observed before the completion of the flooding cycle. Similar events are also reported by Junk (1970) for the Middle Amazon and Sabattini and Lallana (2008) for the middle Paraná River. It is also during this phase that wind and wave action detaches portions of the free floating mats leading to errant floating islands drifting in channels, rivers and lakes.
IV. Receding phase: After maximum floods during November a continuous drawdown is initiated ending with the lowest water level in February. Thus, if rising water level represents an increasing homogenization process (Thomaz et al., 2007), drawdown could be seen as an inverse progression toward heterogeneization through floodplain fragmentation. Receding water also affects the littoral in different ways, retreating faster and more extensively on gentle sloping terrain than on steep sloping shorelines. On the other hand the receding phase can be associated to a flowering and seed producing stage of aquatic and wetland species. In certain species the ability to reproduce is related to the age of the plant, requiring only short vegetative periods before flowering (Sculthorpe, 1985). For instance, in the Middle Amazon flowering of Paspalum repens initiates during the middle of the rain season and ends in the middle of the dry season (Junk, 1970), while sporocarp production of Salvinia auriculata is associated with habitats that dry up (Coelho, Lopes, & Sperber, 2005). In the ZFLC the later also has been observed for Salvinia sprucei and Azolla microphylla. However, floral initiation frequently occurs at the end of maximum vegetative growth, and may also occur in response to water temperature and chemistry (Junk, 1983; Ghosh, Santra, & Mukherjee, 1993).
Acknowledgments
The first author acknowledges Gerardo Viña Vizcaino and Ricardo Sandoval (Control Ambiental, Ecopetrol-DCC) for logistic support; Alberto Ramírez and Efrain Rondon (Environmental Biologists) provided assistance in the field; Roberto Jaramillo M. and José Luis Fernández Alonso (Herbario Nacional Colombiano COL, Bogotá) for help in species identification. Also thanks to all those fishers who paddled the canoe and introduced the first author to the less accessible and remote parts of the Zapatosa floodplain lakes. Field research was supported by contract No. 5-2201-0321 and 5-2201-0082 with the Colombian Petroleum Company (Ecopetrol-DCC), Cúcuta.
References
APG III. (2009). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society, 161(2), 105-121. [ Links ]
Azza, N., Denny, P., van de Koppel, J., & Kansiime, F. (2006). Floating mats: their occurrence and influence on shoreline distribution of emergent vegetation. Freshwater Biology, 51(7), 1286-1297. [ Links ]
Belbin, L. & McDonald, C. (1993). Comparing three classification strategies for use in ecology. Journal of Vegetation Science, 4(3), 341-348. [ Links ]
Boschilia, S. M., Oliveira, E. F., & Thomaz, S. M. (2008). Do aquatic macrophytes co-occur randomly? An analysis of null models in a tropical floodplain. Oecologia, 156(1), 203-214. [ Links ]
Bulla, C. K., Gomes, L. C., Miranda, L. E., & Agostinho, A. A. (2011). The ichthyofauna of drifting macrophyte mats in the Ivinhema River, upper Paraná River basin, Brazil. Neotropical Ichthyology, 9(2), 403-409. [ Links ]
Casanova, M. T. & Brock, M. A. (2000). How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecology, 147(2), 237-250. [ Links ]
Castroviejo, S. & López, G. (1985). Estudio y descripción de las comunidades vegetales del “Hato El Frío” los llanos de Venezuela. Memoria de la Fundación La Salle de Ciencias Naturales, 45(124), 79-151. [ Links ]
Cervi, A. C., Bona, C., de Chiara Moço, M. C., & von Linsingen, L. (2009). Macrófitas aquáticas do Município de General Carneiro, Paraná, Brasil. Biota Neotropica, 9(3), 215-222. [ Links ]
Chambers, P. A., Lacoul, P., Murphy, K. J., & Thomaz, S. M. (2008). Global diversity of aquatic macrophytes in freshwater. Hydrobiologia, 595, 9-26. [ Links ]
Chick, J. H. & McIvor, C. C. (1997). Habitat selection by three littoral zone fishes: effects of predation pressure, plant density and macrophyte type. Ecology of Freshwater Fish, 6(1), 27-35. [ Links ]
Coelho, F. F., Lopes, F. S., & Sperber, C. F. (2005). Persistence strategy of Salvinia auriculata Aublet in temporary ponds of Southern Pantanal, Brazil. Aquatic Botany, 81(4), 343-352. [ Links ]
Cunha-Santino, M. B. C. & Bianchini Jr, I. (2009). Humificação e mineralização de macrófitas aquáticas: uma revisão sobre esses processos. Oecologia Brasiliensis, 13(04), 665-675. [ Links ]
Do Prado, A. L., Heckman, C. W., & Martins, F. R. (1994). The seasonal succession of biotic communities in wetlands of the tropical wet-and-dry climatic zone: II. The aquatic macrophyte vegetation in the Pantanal of Mato Grosso, Brazil. Internationale Revue der gesamten Hydrobiologie und Hydrographie, 79(4), 569-589. [ Links ]
Dufrêne, M. & Legendre, P. (1997). Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs, 67, 345-366. [ Links ]
Esteves, F. A. (1998). Considerations on the ecology of wetlands with emphasis on brasilian floodplain ecosystems. Oecologia Brasiliensis, 4, 111-135. [ Links ]
Ferreira, C. S., Piedade, M. T. F., Wittmann, A. O. D., & Franco, A. C. (2010). Plant reproduction in the Central Amazonian floodplains: challenges and adaptations. AoB Plants, plq009, 1-14. [ Links ]
Ferreira, F. A., Mormul, R. P., Thomaz, S. M., Pott, A., & Pott, V. J. (2011). Macrophytes in the upper Paraná river floodplain: checklist and comparison with other large South American wetlands. Revista de Biología Tropical, 59(2), 541-556. [ Links ]
Finlayson, C. M. (2005). Plant ecology of Australia’s tropical floodplain wetlands: A review. Annals of Botany, 96(4), 541-555. [ Links ]
Fortney, R. H., Benedict, M., Gottgens, J. F., Walters, T. L., Leady, B. S., & Rentch, J. (2004). Aquatic plant community composition and distribution along an inundation gradient at two ecologically-distinct sites in the Pantanal region of Brazil. Wetlands Ecology and Management, 12(6), 575-585. [ Links ]
Franceschi, E. A., Torres, P. S., Prado, D. E., & Lewis, J. P. (2000). Disturbance, succession and stability: a ten year study of temporal variation of species composition after a catastrophic flood in the river Paraná, Argentina. Community Ecology, 1(2), 205-214. [ Links ]
Frey, R. (1995). Flora and vegetation of “Las Piedritas” and the margin of Laguna Cáceres, Puerto Suárez, Bolivian Pantanal. Bulletin of the Torrey Botanical Club, 122(4), 314-319. [ Links ]
Galán de Mera, A. & Linares, E. P. (2008). Datos sobre la vegetación de los humedales de América del Sur. De las sabanas bolivianas a los llanos del Orinoco (Venezuela). Acta Botanica Malacitana, 33, 271-288. [ Links ]
Galán de Mera, A. & Navarro, G. (1992). Comunidades vegetales acuáticas del Paraguay occidental. Caldasia, 17(1), 35-46. [ Links ]
Galán de Mera, A. & Vicente Orellana, J. A. (2006). Aproximación al esquema sintaxonómico de la vegetación de la región del Caribe y América del Sur. Anales de Biología, 28, 3-27. [ Links ]
Galvis, G. & Mojica, J. I. (2007). The Magdalena River fresh water fishes and fisheries. Aquatic Ecosystem Health & Management, 10(2), 127-139. [ Links ]
Gamble, D. W., Parnell, D. B., & Curtis, S. (2008). Spatial variability of the Caribbean mid-summer drought and relation to north Atlantic high circulation. International Journal of Climatology, 28(3), 343-350. [ Links ]
Ghosh, S. K., Santra, S. C., & Mukherjee, P. K. (1993). Phenological studies in aquatic macrophytic plants of lower Gangetic Delta, West Bengal, India. Feddes Repertorium, 104(1/2), 93-111. [ Links ]
Gomes, L. C., Bulla, C. K., Agostinho, A. A., Vasconcelos, L. P., & Miranda, L. E. (2012). Fish assemblage dynamics in a Neotropical floodplain relative to aquatic macrophytes and the homogenizing effect of a flood pulse. Hydrobiologia, 685(1), 97-107. [ Links ]
Greig-Smith, P. (1983). Quantitative plant ecology. Studies in ecology (3rd ed., Vol. 9). Berkeley: University of California Press. [ Links ]
Grenouillet, G. & Pond, D. (2001). Juvenile fishes in macrophyte beds: influence of food resources, habitat structure and body size. Journal of Fish Biology, 59(4), 939-959. [ Links ]
Haynes, R. R. & Holm-Nielsen, L. B. (1989). Speciation of Alismatidae in the neotropics. In L. B. Holm-Nielsen, I. C. Nielsen, & H. Balslev (Eds.), Tropical forests. Botanical dynamics, speciation and diversity (pp. 211-219). London: Academic Press. [ Links ]
Hedges, J. I., Clark, W. A., Quay, P. D., Richey, J. E., Devol, A. H., & Santos, U. d. M. (1986). Compositions and fluxes of particulate organic material in the Amazon River. Limnology and Oceanography, 31(4), 717-738. [ Links ]
Hutchinson, G. E. (1975). A treatise on limnology, Vol. III: Limnological botany. New York: Wiley. [ Links ]
Irgang, B. E. & Gastal-Junior, C. V. S. (1996). Macrófitas aquáticas da planície costeira do RS. Porto Alegre, Brazil: CPG-Botânica/UFRGS. [ Links ]
Jensén, S. & van der Maarel, E. (1980). Numerical approaches to lake classification with special reference to macrophyte communities. Vegetatio, 42(1/3), 117-128. [ Links ]
John, C. M., Sylas, V. P., Paul, J., & Unni, K. S. (2009). Floating islands in a tropical wetland of peninsular India. Wetlands Ecology and Management, 17(6), 641-653. [ Links ]
Junk, W. J. (1970). Investigations on the ecology and production biology of the “floating meadows” (Paspalo-Echinochloetum) on the Middle Amazon. 1. The floating vegetation and its ecology. Amazoniana, 2, 449-495. [ Links ]
Junk, W. J. (1973). Investigations on the ecology and production biology of the “floating meadows” (Paspalo-Echinochloetum) on the Middle Amazon. 2. The aquatic fauna in the root zone of floating vegetation. Amazoniana, 4, 9-102. [ Links ]
Junk, W. J. (1983). Ecology of swamps on the Middle Amazon. In A. J. P. Gore (Ed), Ecosystems of the world: Vol. 4. Mires: swamp, bog, fen and moor. Regional studies (1st ed., pp. 269-294). Amsterdam: Elsevier. [ Links ]
Junk, W. J. (1986). Aquatic plants of the Amazon system. In B. R. Davies & K. F. Walker (Eds), Monographiae biologicae: Vol. 60. The ecology of river systems (pp. 319-337). Dordrecht: Junk. [ Links ]
Junk, W. J., Bayley, P. B., & Sparks, R. E. (1989). The flood pulse concept in river-floodplain systems. In D. P. Dodge (Ed.), Canadian Special Publication of Fisheries and Aquatic Sciences: Vol. 106. Proceedings of the International Large River Symposium (LARS) (pp. 110-127). [ Links ]
Junk, W. J. & Howard-Williams, C. (1984). Ecology of aquatic macrophytes in Amazonia. In H. Sioli (Ed.), Monographiae biologicae: Vol. 56. The Amazon. Limnology and landscape ecology of a mighty tropical river and its basin (pp. 269-293). Dordrecht, Netherlands: Kluwer Academic Publishers. [ Links ]
Junk, W. J. & Piedade, M. T. F. (1993). Herbaceous plants of the Amazon floodplain near Manaus: species diversity and adaptions to the flood pulse. Amazoniana, 12(3/4), 467-484. [ Links ]
Junk, W. J. & Piedade, M. T. F. (1997). Plant life in the floodplain with special reference to herbaceous plants. In W. J. Junk (Ed.), Ecological studies: Vol. 126. The central Amazon floodplain. Ecology of a pulsing system (pp. 147-185). Berlin: Springer. [ Links ]
Junk, W. J. & Wantzen, K. M. (2004). The flood pulse concept: new aspects, approaches and applications - an update. In R. L. Welcomme & T. Petr (Eds.), Sustaining livelihoods and biodiversity in the new millennium. Ecoregion conservation for freshwater systems, with a focus on large rivers (pp. 117-140). Bankok: FAO Regional Office for Asia and the Pacific, RAP Publication 2004/17. [ Links ]
Kent, M. (2012). Vegetation description and data analysis: A practical approach (2. ed.). Chichester, West Sussex: Wiley. [ Links ]
Kufner, D. C. L., Scremin-Dias, E., & Guglieri-Caporal, A. (2011). Composição florística e variação sazonal da biomassa de macrófitas aquáticas em lagoa de meandro do Pantanal. Rodriguésia, 62(4), 803-812. [ Links ]
Landolt, E. & Schmidt-Mumm, U. (2009). Lemnaceae. Flora de Colombia, 24, 4-54. [ Links ]
Legendre, P. & Legendre, L. (2012). Numerical ecology (3rd ed.). Oxford: Elsevier. [ Links ]
Maltchik, L., de Oliveira, G. R., Rolon, A. S., & Stenert, C. (2005). Diversity and stability of aquatic macrophyte community in three shallow lakes associated to a floodplain system in the south of Brazil. Interciencia, 30(3), 166-170. [ Links ]
Maltchik, L., Rolon, A. S., & Schott, P. (2007). Effects of hydrological variation on the aquatic plant community in a floodplain palustrine wetland of southern Brazil. Limnology, 8(1), 23-28. [ Links ]
McCune, B. & Grace, J. B. (2002). Analysis of ecological communities. Gleneden Beach, Oregon, U.S.A.: MjM Software Design. Retrieved from http://www.gbv.de/dms/hebis-darmstadt/toc/106964135.pdf [ Links ]
McCune, B. & Mefford, M. J. (2011). PC-ORD. Multivariate Analysis of Ecological Data. Version 6. Gleneden Beach, Oregon, U.S.A.: MjM Software Design. [ Links ]
Melack, J. M. & Fisher, T. R. (1990). Comparative limnology of tropical floodplain lakes with an emphasis on the Central Amazon. Acta Limnologica Brasiliensia, 3, 1-48. [ Links ]
Middleton, B. (1999). Succession and herbivory in monsoonal wetlands. Wetlands Ecology and Management, 6(4), 189-202. [ Links ]
Müller, J. V. & Deil, U. (2005). The ephemeral vegetation of seasonal and semi-permanent ponds in tropical West Africa. Phytocoenologia, 35(2), 327-388. [ Links ]
Murphy, K. J., Dickinson, G., Thomaz, S. M., Bini, L. M., Dick, K., Greaves, K., Wingfield, R. A. (2003). Aquatic plant communities and predictors of diversity in a sub-tropical river floodplain: the upper Rio Paraná, Brazil. Aquatic Botany, 77(4), 257-276. [ Links ]
Neiff, J. J., Casco, S. L., & Poi de Neiff, A. (2008). Response of Eichhornia crassipes (Pontederiaceae) to water level fluctuations in two lakes with different connectivity in the Paraná River floodplain. Revista de Biología Tropical, 56(2), 613-623. [ Links ]
Osland, M. J., González, E., & Richardson, C. J. (2011). Coastal freshwater wetland plant community response to seasonal drought and flooding in northwestern Costa Rica. Wetlands, 31(4), 641-652. [ Links ]
Padial, A. A., Carvalho, P., Thomaz, S. M., Boschilia, S. M., Rodrigues, R. B., & Kobayashi, J. T. (2009). The role of an extreme flood disturbance on macrophyte assemblages in a Neotropical floodplain. Aquatic Sciences, 71(4), 389-398. [ Links ]
Padial, A. A. & Thomaz, S. M. (2006). Effects of flooding regime upon the decomposition of Eichhornia azurea (Sw.) Kunth measured on a tropical, flow-regulated floodplain (Paraná River, Brazil). River Research and Applications, 22(7), 791-801. [ Links ]
Piedade, M. T. F., Junk, W. J., D`Ângelo, S. A., Wittmann, F., Schöngart, J., Barbosa, K. M. D. N., & Lopes, A. (2010). Aquatic herbaceous plants of the Amazon floodplains: state of the art and research needed. Acta Limnologica Brasiliensia, 22(02), 165-178. [ Links ]
Pivari, M. O., Pott, V. J., & Pott, A. (2008). Macrófitas aquáticas de ilhas flutuantes (baceiros) nas sub-regiões do Abobral e Miranda, Pantanal, MS, Brasil. Acta Botanica Brasilica, 22(2), 563-571. [ Links ]
Poi de Neiff, A. (2003). Macroinvertebrates living on Eichhornia azurea Kunth in the Paraguay River. Acta Limnologica Brasiliensia, 15(1), 55-63. [ Links ]
Poi de Neiff, A. & Carignan, R. (1997). Macroinvertebrates on Eichhornia crassipes roots in two lakes of the Paraná River floodplain. Hydrobiologia, 345(2/3), 185-196. [ Links ]
Polunin, N. V. (1984). The decomposition of emergent macrophytes in fresh water. Advances in Ecological Research, 14, 115-166. [ Links ]
Pott, A. & Pott, V. J. (2004). Features and conservation of the Brazilian Pantanal wetland. Wetlands Ecology and Management, 12(6), 547-552. [ Links ]
Pott, V. J. & Pott, A. (2011). Species diversity, distribution, and biomass of aquatic macrophytes of the Pantanal. In W. J. Junk, C. J. Da Silva, C. Nunes da Cunha, & K. M. Wantzen (Eds.), The Pantanal: Ecology, biodiversity and sustainable management of a large neotropical seasonal wetland (pp. 257-279). Sofia: Pensoft Publishers. [ Links ]
Pott, V. J., Pott, A., Lima, L. C. P., Moreira, S. N., & Oliveira, A. K. M. (2011). Aquatic macrophyte diversity of the Pantanal wetland and upper basin. Brazilian Journal of Biology, 71(1 supl. 1), 255-263. [ Links ]
Poveda, G., Waylen, P. R., & Pulwarty, R. S. (2006). Annual and inter-annual variability of the present climate in northern South America and southern Mesoamerica. Palaeogeography, Palaeoclimatology, Palaeoecology, 234(1), 3-27. [ Links ]
Pringle, C. M. (2001). Hydrologic connectivity and the management of biological reserves: a global perspective. Ecological Applications, 11(4), 981-998. [ Links ]
Rangel-Ch., J. O. (Ed.). (2007). Estudio de inventario de fauna, flora, descripción biofísica y socioeconómica y línea de base ambiental Ciénaga de Zapatosa: Informe final de actividades. Bogotá: Corpocesar-Universidad Nacional de Colombia. Retrieved from http://www.corpocesar.gov.co/files/Informe%20Final%20de%20Actividades%20Zapatosa.pdf [ Links ]
Rangel-Ch., J. O. (2010). Vegetación acuática, caracterización inicial. In J. O. Rangel-Ch. (Ed.), Colombia diversidad biótica IX. Ciénagas de Córdoba: Biodiversidad, ecología y manejo ambiental (pp. 325-339). Bogotá D. C.: Instituto de Ciencias Naturales. Universidad Nacional de Colombia. [ Links ]
Rees, W. A. (1978). The ecology of the Kafue Lechwe: soils, water levels and vegetation. Journal of Applied Ecology, 15(1), 163-176. [ Links ]
Restrepo, J. D. & Kjerfve, B. (2000). Magdalena river: interannual variability (1975-1995) and revised water discharge and sediment load estimates. Journal of Hydrology, 235(1-2), 137-149. [ Links ]
Restrepo, J. D. & Kjerfve, B. (2004). The Pacific and Caribbean rivers of Colombia: water discharge, sediment transport and dissolved loads. In L. D. Lacerda, R. E. Santelli, E. K. Duursma, & J. J. Abrão (Eds.), Environmental science. Environmental geochemistry in tropical and subtropical environments (pp. 235-251). Berlin: Springer. [ Links ]
Restrepo, J. D. & Syvitski, J. P. M. (2006). Assessing the effect of natural controls and land use change on sediment yield in a major andean river: the Magdalena drainage basin, Colombia. Ambio: A Journal of the Human Environment, 35(2), 65-74. [ Links ]
Rial, B. A. (2000). Aspectos cualitativos de la zonación y estratificación de comunidades de plantas acuáticas en un humedal de los Llanos de Venezuela. Memoria de la Fundación La Salle de Ciencias Naturales, 60(153), 69-85. [ Links ]
Rial, B. A. (2006). Variabilidad espacio-temporal de las comunidades de plantas acuáticas en un humedal de los Llanos de Venezuela. Revista de Biología Tropical, 54(2), 403-413. [ Links ]
Ríos-Pulgarín, M. I., Jiménez-Segura, L. F., Palacio, J. A., & Ramírez Restrepo, J. J. (2008). Comunidad de peces en la Ciénaga de Ayapel, Río Magdalena (Córdoba) Colombia: cambios espacio-temporales en su asociación. Actualidades Biológicas, 30(88), 29-53. [ Links ]
Rolon, A. S., Lacerda, T., Maltchik, L., & Guadagnin, D. L. (2008). Influence of area, habitat and water chemistry on richness and composition of macrophyte assemblages in southern Brazilian wetlands. Journal of Vegetation Science, 19, 221-228. [ Links ]
Sabattini, R. A. & Lallana, V. H. (2008). Aquatic macrophytes. In M. H. Iriondo, J. C. Paggi, & M. J. Parma (Eds.), The Middle Paraná River. Limnology of a Subtropical Wetland (pp. 205-226). Berlin: Springer. [ Links ]
Santos, A. M. & Thomaz, S. M. (2007). Aquatic macrophytes diversity in lagoons of a tropical floodplain: The role of connectivity and water level. Austral Ecology, 32, 177-190. [ Links ]
Santos, A. M. & Thomaz, S. M. (2008). Short term fluctuations and succession trends in tropical floodplain vegetation measured by correspondence analysis. Brazilian Archives of Biology and Technology, 51(4), 781-791. [ Links ]
Schmidt-Mumm, U. (1987). Lista comentada de los macrófitos acuáticos y palustres de la región de Barrancabermeja (Santander). Pérez-Arbelaezia, 1(4/5), 43-53. [ Links ]
Schmidt-Mumm, U. & Vargas, O. (2012). Comunidades vegetales de las transiciones terrestre-acuáticas del páramo de Chingaza, Colombia. Revista de Biología Tropical, 60(1), 35-64. [ Links ]
Sculthorpe, C. D. (1985). The biology of aquatic vascular plants. Königstein: Koeltz. [ Links ]
Sousa, W. T., Thomaz, S. M., & Murphy, K. J. (2011). Drivers of aquatic macrophyte community structure in a Neotropical riverine lake. Acta Oecologica, 37(5), 462-475. [ Links ]
Takeda, A. M., de Souza-Franco, G. M., de Melo, S. M., & Monkolski, A. (2003). Invertebrados associados às macrófitas aquáticas da planície de inundação do alto rio Paraná (Brasil). In S. M. Thomaz & L. M. Bini (Eds.), Ecologia e manejo de macrófitas aquáticas (pp. 243-260). Maringá: EDUEM. [ Links ]
Thomaz, S. M., Bini, L. M., & Bozelli, R. L. (2007). Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia, 579(1), 1-13. [ Links ]
Thomaz, S. M., Carvalho, P., Padial, A. A., & Kobayashi, J. T. (2009). Temporal and spatial patterns of aquatic macrophyte diversity in the Upper Paraná River floodplain. Brazilian Journal of Biology, 69(2), 617-625. [ Links ]
Thomaz, S. M., Souza, D. C., & Bini, L. M. (2003). Species richness and beta diversity of aquatic macrophytes in a large subtropical reservoir (Itaipu Reservoir, Brazil): the influence of limnology and morphometry. Hydrobiologia, 505(1-3), 119-128. [ Links ]
Tockner, K., Malard, F., & Ward, J. V. (2000). An extension of the flood pulse concept. Hydrological Processes, 14(16-17), 2861-2883. [ Links ]
Tur, N. M. (1972). Embalsados y camalotales de la región isleña del Paraná Medio. Darwiniana, 17, 397-407. [ Links ]
Unni, K. S. (1971). An ecological study of the macrophytic vegetation of the Doodhadhari Lake, Raipur, M. P., India 1. Distribution and seasonal change in aquatic plants. Hydrobiologia, 37(1), 139-155. [ Links ]
van der Valk, A. G. (2005). Water-level fluctuations in North American prairie wetlands. Hydrobiologia, 539(1), 171-188. [ Links ]
van der Valk, A. G., Middleton, B. A., Williams, R. L., Mason, D. H., & Davis, C. B. (1993). The biomass of an Indian monsoonal wetland before and after being overgrown with Paspalum distichum L. Vegetatio, 109(1), 81-90. [ Links ]
van Groenewoud, H. (1992). The robustness of correspondence, detrended correspondence, and TWINSPAN analysis. Journal of Vegetation Science, 3(2), 239-246. [ Links ]
Viña, G., Ramírez, A. G., Lamprea, L., Garzón, B., Schmidt-Mumm, U., Rondon, E., & Florez, C. (1991). Ecología de la Ciénaga de Zapatosa y su relación con un derrame de petróleo. Informe Control Ambiental. Cúcuta. [ Links ]
Wantzen, K. M., Junk, W. J., & Rothhaupt, K. O. (2008). An extension of the floodpulse concept (FPC) for lakes. Hydrobiologia, 613(1), 151-170. [ Links ]
Wantzen, K. M., Yule, C. M., Mathooko, J. M., & Pringle, C. M. (2008). Organic matter processing in tropical streams. In D. Dudgeon (Ed.), Aquatic ecology series. Tropical stream ecology (1st ed., pp. 43-64). London, Burlington, MA: Academic Press. [ Links ]
Warfe, D. M. & Barmuta, L. (2004). Habitat structural complexity mediates the foraging success of multiple predator species. Oecologia, 141(1), 171-178. [ Links ]
Warfe, D. M., Pettit, N. E., Davies, P. M., Pusey, B. J., Hamilton, S. K., Kennard, M. J., & Halliday, I. A. (2011). The ‘wet-dry’ in the wet-dry tropics drives river ecosystem structure and processes in northern Australia. Freshwater Biology, 56(11), 2169-2195. [ Links ]
Wiegleb, G. (1991). Die Lebens- und Wuchsformen der makrophytischen Wasserpflanzen und deren Beziehungen zur Ökologie, Verbreitung und Vergesellschaftung der Arten. Tuexenia, 11, 135-147. [ Links ]
Azza, N., Denny, P., van de Koppel, J., & Kansiime, F. (2006). Floating mats: their occurrence and influence on shoreline distribution of emergent vegetation. Freshwater Biology, 51(7), 1286-1297. [ Links ]
Belbin, L. & McDonald, C. (1993). Comparing three classification strategies for use in ecology. Journal of Vegetation Science, 4(3), 341-348. [ Links ]
Boschilia, S. M., Oliveira, E. F., & Thomaz, S. M. (2008). Do aquatic macrophytes co-occur randomly? An analysis of null models in a tropical floodplain. Oecologia, 156(1), 203-214. [ Links ]
Bulla, C. K., Gomes, L. C., Miranda, L. E., & Agostinho, A. A. (2011). The ichthyofauna of drifting macrophyte mats in the Ivinhema River, upper Paraná River basin, Brazil. Neotropical Ichthyology, 9(2), 403-409. [ Links ]
Casanova, M. T. & Brock, M. A. (2000). How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecology, 147(2), 237-250. [ Links ]
Castroviejo, S. & López, G. (1985). Estudio y descripción de las comunidades vegetales del “Hato El Frío” los llanos de Venezuela. Memoria de la Fundación La Salle de Ciencias Naturales, 45(124), 79-151. [ Links ]
Cervi, A. C., Bona, C., de Chiara Moço, M. C., & von Linsingen, L. (2009). Macrófitas aquáticas do Município de General Carneiro, Paraná, Brasil. Biota Neotropica, 9(3), 215-222. [ Links ]
Chambers, P. A., Lacoul, P., Murphy, K. J., & Thomaz, S. M. (2008). Global diversity of aquatic macrophytes in freshwater. Hydrobiologia, 595, 9-26. [ Links ]
Chick, J. H. & McIvor, C. C. (1997). Habitat selection by three littoral zone fishes: effects of predation pressure, plant density and macrophyte type. Ecology of Freshwater Fish, 6(1), 27-35. [ Links ]
Coelho, F. F., Lopes, F. S., & Sperber, C. F. (2005). Persistence strategy of Salvinia auriculata Aublet in temporary ponds of Southern Pantanal, Brazil. Aquatic Botany, 81(4), 343-352. [ Links ]
Cunha-Santino, M. B. C. & Bianchini Jr, I. (2009). Humificação e mineralização de macrófitas aquáticas: uma revisão sobre esses processos. Oecologia Brasiliensis, 13(04), 665-675. [ Links ]
Do Prado, A. L., Heckman, C. W., & Martins, F. R. (1994). The seasonal succession of biotic communities in wetlands of the tropical wet-and-dry climatic zone: II. The aquatic macrophyte vegetation in the Pantanal of Mato Grosso, Brazil. Internationale Revue der gesamten Hydrobiologie und Hydrographie, 79(4), 569-589. [ Links ]
Dufrêne, M. & Legendre, P. (1997). Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs, 67, 345-366. [ Links ]
Esteves, F. A. (1998). Considerations on the ecology of wetlands with emphasis on brasilian floodplain ecosystems. Oecologia Brasiliensis, 4, 111-135. [ Links ]
Ferreira, C. S., Piedade, M. T. F., Wittmann, A. O. D., & Franco, A. C. (2010). Plant reproduction in the Central Amazonian floodplains: challenges and adaptations. AoB Plants, plq009, 1-14. [ Links ]
Ferreira, F. A., Mormul, R. P., Thomaz, S. M., Pott, A., & Pott, V. J. (2011). Macrophytes in the upper Paraná river floodplain: checklist and comparison with other large South American wetlands. Revista de Biología Tropical, 59(2), 541-556. [ Links ]
Finlayson, C. M. (2005). Plant ecology of Australia’s tropical floodplain wetlands: A review. Annals of Botany, 96(4), 541-555. [ Links ]
Fortney, R. H., Benedict, M., Gottgens, J. F., Walters, T. L., Leady, B. S., & Rentch, J. (2004). Aquatic plant community composition and distribution along an inundation gradient at two ecologically-distinct sites in the Pantanal region of Brazil. Wetlands Ecology and Management, 12(6), 575-585. [ Links ]
Franceschi, E. A., Torres, P. S., Prado, D. E., & Lewis, J. P. (2000). Disturbance, succession and stability: a ten year study of temporal variation of species composition after a catastrophic flood in the river Paraná, Argentina. Community Ecology, 1(2), 205-214. [ Links ]
Frey, R. (1995). Flora and vegetation of “Las Piedritas” and the margin of Laguna Cáceres, Puerto Suárez, Bolivian Pantanal. Bulletin of the Torrey Botanical Club, 122(4), 314-319. [ Links ]
Galán de Mera, A. & Linares, E. P. (2008). Datos sobre la vegetación de los humedales de América del Sur. De las sabanas bolivianas a los llanos del Orinoco (Venezuela). Acta Botanica Malacitana, 33, 271-288. [ Links ]
Galán de Mera, A. & Navarro, G. (1992). Comunidades vegetales acuáticas del Paraguay occidental. Caldasia, 17(1), 35-46. [ Links ]
Galán de Mera, A. & Vicente Orellana, J. A. (2006). Aproximación al esquema sintaxonómico de la vegetación de la región del Caribe y América del Sur. Anales de Biología, 28, 3-27. [ Links ]
Galvis, G. & Mojica, J. I. (2007). The Magdalena River fresh water fishes and fisheries. Aquatic Ecosystem Health & Management, 10(2), 127-139. [ Links ]
Gamble, D. W., Parnell, D. B., & Curtis, S. (2008). Spatial variability of the Caribbean mid-summer drought and relation to north Atlantic high circulation. International Journal of Climatology, 28(3), 343-350. [ Links ]
Ghosh, S. K., Santra, S. C., & Mukherjee, P. K. (1993). Phenological studies in aquatic macrophytic plants of lower Gangetic Delta, West Bengal, India. Feddes Repertorium, 104(1/2), 93-111. [ Links ]
Gomes, L. C., Bulla, C. K., Agostinho, A. A., Vasconcelos, L. P., & Miranda, L. E. (2012). Fish assemblage dynamics in a Neotropical floodplain relative to aquatic macrophytes and the homogenizing effect of a flood pulse. Hydrobiologia, 685(1), 97-107. [ Links ]
Greig-Smith, P. (1983). Quantitative plant ecology. Studies in ecology (3rd ed., Vol. 9). Berkeley: University of California Press. [ Links ]
Grenouillet, G. & Pond, D. (2001). Juvenile fishes in macrophyte beds: influence of food resources, habitat structure and body size. Journal of Fish Biology, 59(4), 939-959. [ Links ]
Haynes, R. R. & Holm-Nielsen, L. B. (1989). Speciation of Alismatidae in the neotropics. In L. B. Holm-Nielsen, I. C. Nielsen, & H. Balslev (Eds.), Tropical forests. Botanical dynamics, speciation and diversity (pp. 211-219). London: Academic Press. [ Links ]
Hedges, J. I., Clark, W. A., Quay, P. D., Richey, J. E., Devol, A. H., & Santos, U. d. M. (1986). Compositions and fluxes of particulate organic material in the Amazon River. Limnology and Oceanography, 31(4), 717-738. [ Links ]
Hutchinson, G. E. (1975). A treatise on limnology, Vol. III: Limnological botany. New York: Wiley. [ Links ]
Irgang, B. E. & Gastal-Junior, C. V. S. (1996). Macrófitas aquáticas da planície costeira do RS. Porto Alegre, Brazil: CPG-Botânica/UFRGS. [ Links ]
Jensén, S. & van der Maarel, E. (1980). Numerical approaches to lake classification with special reference to macrophyte communities. Vegetatio, 42(1/3), 117-128. [ Links ]
John, C. M., Sylas, V. P., Paul, J., & Unni, K. S. (2009). Floating islands in a tropical wetland of peninsular India. Wetlands Ecology and Management, 17(6), 641-653. [ Links ]
Junk, W. J. (1970). Investigations on the ecology and production biology of the “floating meadows” (Paspalo-Echinochloetum) on the Middle Amazon. 1. The floating vegetation and its ecology. Amazoniana, 2, 449-495. [ Links ]
Junk, W. J. (1973). Investigations on the ecology and production biology of the “floating meadows” (Paspalo-Echinochloetum) on the Middle Amazon. 2. The aquatic fauna in the root zone of floating vegetation. Amazoniana, 4, 9-102. [ Links ]
Junk, W. J. (1983). Ecology of swamps on the Middle Amazon. In A. J. P. Gore (Ed), Ecosystems of the world: Vol. 4. Mires: swamp, bog, fen and moor. Regional studies (1st ed., pp. 269-294). Amsterdam: Elsevier. [ Links ]
Junk, W. J. (1986). Aquatic plants of the Amazon system. In B. R. Davies & K. F. Walker (Eds), Monographiae biologicae: Vol. 60. The ecology of river systems (pp. 319-337). Dordrecht: Junk. [ Links ]
Junk, W. J., Bayley, P. B., & Sparks, R. E. (1989). The flood pulse concept in river-floodplain systems. In D. P. Dodge (Ed.), Canadian Special Publication of Fisheries and Aquatic Sciences: Vol. 106. Proceedings of the International Large River Symposium (LARS) (pp. 110-127). [ Links ]
Junk, W. J. & Howard-Williams, C. (1984). Ecology of aquatic macrophytes in Amazonia. In H. Sioli (Ed.), Monographiae biologicae: Vol. 56. The Amazon. Limnology and landscape ecology of a mighty tropical river and its basin (pp. 269-293). Dordrecht, Netherlands: Kluwer Academic Publishers. [ Links ]
Junk, W. J. & Piedade, M. T. F. (1993). Herbaceous plants of the Amazon floodplain near Manaus: species diversity and adaptions to the flood pulse. Amazoniana, 12(3/4), 467-484. [ Links ]
Junk, W. J. & Piedade, M. T. F. (1997). Plant life in the floodplain with special reference to herbaceous plants. In W. J. Junk (Ed.), Ecological studies: Vol. 126. The central Amazon floodplain. Ecology of a pulsing system (pp. 147-185). Berlin: Springer. [ Links ]
Junk, W. J. & Wantzen, K. M. (2004). The flood pulse concept: new aspects, approaches and applications - an update. In R. L. Welcomme & T. Petr (Eds.), Sustaining livelihoods and biodiversity in the new millennium. Ecoregion conservation for freshwater systems, with a focus on large rivers (pp. 117-140). Bankok: FAO Regional Office for Asia and the Pacific, RAP Publication 2004/17. [ Links ]
Kent, M. (2012). Vegetation description and data analysis: A practical approach (2. ed.). Chichester, West Sussex: Wiley. [ Links ]
Kufner, D. C. L., Scremin-Dias, E., & Guglieri-Caporal, A. (2011). Composição florística e variação sazonal da biomassa de macrófitas aquáticas em lagoa de meandro do Pantanal. Rodriguésia, 62(4), 803-812. [ Links ]
Landolt, E. & Schmidt-Mumm, U. (2009). Lemnaceae. Flora de Colombia, 24, 4-54. [ Links ]
Legendre, P. & Legendre, L. (2012). Numerical ecology (3rd ed.). Oxford: Elsevier. [ Links ]
Maltchik, L., de Oliveira, G. R., Rolon, A. S., & Stenert, C. (2005). Diversity and stability of aquatic macrophyte community in three shallow lakes associated to a floodplain system in the south of Brazil. Interciencia, 30(3), 166-170. [ Links ]
Maltchik, L., Rolon, A. S., & Schott, P. (2007). Effects of hydrological variation on the aquatic plant community in a floodplain palustrine wetland of southern Brazil. Limnology, 8(1), 23-28. [ Links ]
McCune, B. & Grace, J. B. (2002). Analysis of ecological communities. Gleneden Beach, Oregon, U.S.A.: MjM Software Design. Retrieved from http://www.gbv.de/dms/hebis-darmstadt/toc/106964135.pdf [ Links ]
McCune, B. & Mefford, M. J. (2011). PC-ORD. Multivariate Analysis of Ecological Data. Version 6. Gleneden Beach, Oregon, U.S.A.: MjM Software Design. [ Links ]
Melack, J. M. & Fisher, T. R. (1990). Comparative limnology of tropical floodplain lakes with an emphasis on the Central Amazon. Acta Limnologica Brasiliensia, 3, 1-48. [ Links ]
Middleton, B. (1999). Succession and herbivory in monsoonal wetlands. Wetlands Ecology and Management, 6(4), 189-202. [ Links ]
Müller, J. V. & Deil, U. (2005). The ephemeral vegetation of seasonal and semi-permanent ponds in tropical West Africa. Phytocoenologia, 35(2), 327-388. [ Links ]
Murphy, K. J., Dickinson, G., Thomaz, S. M., Bini, L. M., Dick, K., Greaves, K., Wingfield, R. A. (2003). Aquatic plant communities and predictors of diversity in a sub-tropical river floodplain: the upper Rio Paraná, Brazil. Aquatic Botany, 77(4), 257-276. [ Links ]
Neiff, J. J., Casco, S. L., & Poi de Neiff, A. (2008). Response of Eichhornia crassipes (Pontederiaceae) to water level fluctuations in two lakes with different connectivity in the Paraná River floodplain. Revista de Biología Tropical, 56(2), 613-623. [ Links ]
Osland, M. J., González, E., & Richardson, C. J. (2011). Coastal freshwater wetland plant community response to seasonal drought and flooding in northwestern Costa Rica. Wetlands, 31(4), 641-652. [ Links ]
Padial, A. A., Carvalho, P., Thomaz, S. M., Boschilia, S. M., Rodrigues, R. B., & Kobayashi, J. T. (2009). The role of an extreme flood disturbance on macrophyte assemblages in a Neotropical floodplain. Aquatic Sciences, 71(4), 389-398. [ Links ]
Padial, A. A. & Thomaz, S. M. (2006). Effects of flooding regime upon the decomposition of Eichhornia azurea (Sw.) Kunth measured on a tropical, flow-regulated floodplain (Paraná River, Brazil). River Research and Applications, 22(7), 791-801. [ Links ]
Piedade, M. T. F., Junk, W. J., D`Ângelo, S. A., Wittmann, F., Schöngart, J., Barbosa, K. M. D. N., & Lopes, A. (2010). Aquatic herbaceous plants of the Amazon floodplains: state of the art and research needed. Acta Limnologica Brasiliensia, 22(02), 165-178. [ Links ]
Pivari, M. O., Pott, V. J., & Pott, A. (2008). Macrófitas aquáticas de ilhas flutuantes (baceiros) nas sub-regiões do Abobral e Miranda, Pantanal, MS, Brasil. Acta Botanica Brasilica, 22(2), 563-571. [ Links ]
Poi de Neiff, A. (2003). Macroinvertebrates living on Eichhornia azurea Kunth in the Paraguay River. Acta Limnologica Brasiliensia, 15(1), 55-63. [ Links ]
Poi de Neiff, A. & Carignan, R. (1997). Macroinvertebrates on Eichhornia crassipes roots in two lakes of the Paraná River floodplain. Hydrobiologia, 345(2/3), 185-196. [ Links ]
Polunin, N. V. (1984). The decomposition of emergent macrophytes in fresh water. Advances in Ecological Research, 14, 115-166. [ Links ]
Pott, A. & Pott, V. J. (2004). Features and conservation of the Brazilian Pantanal wetland. Wetlands Ecology and Management, 12(6), 547-552. [ Links ]
Pott, V. J. & Pott, A. (2011). Species diversity, distribution, and biomass of aquatic macrophytes of the Pantanal. In W. J. Junk, C. J. Da Silva, C. Nunes da Cunha, & K. M. Wantzen (Eds.), The Pantanal: Ecology, biodiversity and sustainable management of a large neotropical seasonal wetland (pp. 257-279). Sofia: Pensoft Publishers. [ Links ]
Pott, V. J., Pott, A., Lima, L. C. P., Moreira, S. N., & Oliveira, A. K. M. (2011). Aquatic macrophyte diversity of the Pantanal wetland and upper basin. Brazilian Journal of Biology, 71(1 supl. 1), 255-263. [ Links ]
Poveda, G., Waylen, P. R., & Pulwarty, R. S. (2006). Annual and inter-annual variability of the present climate in northern South America and southern Mesoamerica. Palaeogeography, Palaeoclimatology, Palaeoecology, 234(1), 3-27. [ Links ]
Pringle, C. M. (2001). Hydrologic connectivity and the management of biological reserves: a global perspective. Ecological Applications, 11(4), 981-998. [ Links ]
Rangel-Ch., J. O. (Ed.). (2007). Estudio de inventario de fauna, flora, descripción biofísica y socioeconómica y línea de base ambiental Ciénaga de Zapatosa: Informe final de actividades. Bogotá: Corpocesar-Universidad Nacional de Colombia. Retrieved from http://www.corpocesar.gov.co/files/Informe%20Final%20de%20Actividades%20Zapatosa.pdf [ Links ]
Rangel-Ch., J. O. (2010). Vegetación acuática, caracterización inicial. In J. O. Rangel-Ch. (Ed.), Colombia diversidad biótica IX. Ciénagas de Córdoba: Biodiversidad, ecología y manejo ambiental (pp. 325-339). Bogotá D. C.: Instituto de Ciencias Naturales. Universidad Nacional de Colombia. [ Links ]
Rees, W. A. (1978). The ecology of the Kafue Lechwe: soils, water levels and vegetation. Journal of Applied Ecology, 15(1), 163-176. [ Links ]
Restrepo, J. D. & Kjerfve, B. (2000). Magdalena river: interannual variability (1975-1995) and revised water discharge and sediment load estimates. Journal of Hydrology, 235(1-2), 137-149. [ Links ]
Restrepo, J. D. & Kjerfve, B. (2004). The Pacific and Caribbean rivers of Colombia: water discharge, sediment transport and dissolved loads. In L. D. Lacerda, R. E. Santelli, E. K. Duursma, & J. J. Abrão (Eds.), Environmental science. Environmental geochemistry in tropical and subtropical environments (pp. 235-251). Berlin: Springer. [ Links ]
Restrepo, J. D. & Syvitski, J. P. M. (2006). Assessing the effect of natural controls and land use change on sediment yield in a major andean river: the Magdalena drainage basin, Colombia. Ambio: A Journal of the Human Environment, 35(2), 65-74. [ Links ]
Rial, B. A. (2000). Aspectos cualitativos de la zonación y estratificación de comunidades de plantas acuáticas en un humedal de los Llanos de Venezuela. Memoria de la Fundación La Salle de Ciencias Naturales, 60(153), 69-85. [ Links ]
Rial, B. A. (2006). Variabilidad espacio-temporal de las comunidades de plantas acuáticas en un humedal de los Llanos de Venezuela. Revista de Biología Tropical, 54(2), 403-413. [ Links ]
Ríos-Pulgarín, M. I., Jiménez-Segura, L. F., Palacio, J. A., & Ramírez Restrepo, J. J. (2008). Comunidad de peces en la Ciénaga de Ayapel, Río Magdalena (Córdoba) Colombia: cambios espacio-temporales en su asociación. Actualidades Biológicas, 30(88), 29-53. [ Links ]
Rolon, A. S., Lacerda, T., Maltchik, L., & Guadagnin, D. L. (2008). Influence of area, habitat and water chemistry on richness and composition of macrophyte assemblages in southern Brazilian wetlands. Journal of Vegetation Science, 19, 221-228. [ Links ]
Sabattini, R. A. & Lallana, V. H. (2008). Aquatic macrophytes. In M. H. Iriondo, J. C. Paggi, & M. J. Parma (Eds.), The Middle Paraná River. Limnology of a Subtropical Wetland (pp. 205-226). Berlin: Springer. [ Links ]
Santos, A. M. & Thomaz, S. M. (2007). Aquatic macrophytes diversity in lagoons of a tropical floodplain: The role of connectivity and water level. Austral Ecology, 32, 177-190. [ Links ]
Santos, A. M. & Thomaz, S. M. (2008). Short term fluctuations and succession trends in tropical floodplain vegetation measured by correspondence analysis. Brazilian Archives of Biology and Technology, 51(4), 781-791. [ Links ]
Schmidt-Mumm, U. (1987). Lista comentada de los macrófitos acuáticos y palustres de la región de Barrancabermeja (Santander). Pérez-Arbelaezia, 1(4/5), 43-53. [ Links ]
Schmidt-Mumm, U. & Vargas, O. (2012). Comunidades vegetales de las transiciones terrestre-acuáticas del páramo de Chingaza, Colombia. Revista de Biología Tropical, 60(1), 35-64. [ Links ]
Sculthorpe, C. D. (1985). The biology of aquatic vascular plants. Königstein: Koeltz. [ Links ]
Sousa, W. T., Thomaz, S. M., & Murphy, K. J. (2011). Drivers of aquatic macrophyte community structure in a Neotropical riverine lake. Acta Oecologica, 37(5), 462-475. [ Links ]
Takeda, A. M., de Souza-Franco, G. M., de Melo, S. M., & Monkolski, A. (2003). Invertebrados associados às macrófitas aquáticas da planície de inundação do alto rio Paraná (Brasil). In S. M. Thomaz & L. M. Bini (Eds.), Ecologia e manejo de macrófitas aquáticas (pp. 243-260). Maringá: EDUEM. [ Links ]
Thomaz, S. M., Bini, L. M., & Bozelli, R. L. (2007). Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia, 579(1), 1-13. [ Links ]
Thomaz, S. M., Carvalho, P., Padial, A. A., & Kobayashi, J. T. (2009). Temporal and spatial patterns of aquatic macrophyte diversity in the Upper Paraná River floodplain. Brazilian Journal of Biology, 69(2), 617-625. [ Links ]
Thomaz, S. M., Souza, D. C., & Bini, L. M. (2003). Species richness and beta diversity of aquatic macrophytes in a large subtropical reservoir (Itaipu Reservoir, Brazil): the influence of limnology and morphometry. Hydrobiologia, 505(1-3), 119-128. [ Links ]
Tockner, K., Malard, F., & Ward, J. V. (2000). An extension of the flood pulse concept. Hydrological Processes, 14(16-17), 2861-2883. [ Links ]
Tur, N. M. (1972). Embalsados y camalotales de la región isleña del Paraná Medio. Darwiniana, 17, 397-407. [ Links ]
Unni, K. S. (1971). An ecological study of the macrophytic vegetation of the Doodhadhari Lake, Raipur, M. P., India 1. Distribution and seasonal change in aquatic plants. Hydrobiologia, 37(1), 139-155. [ Links ]
van der Valk, A. G. (2005). Water-level fluctuations in North American prairie wetlands. Hydrobiologia, 539(1), 171-188. [ Links ]
van der Valk, A. G., Middleton, B. A., Williams, R. L., Mason, D. H., & Davis, C. B. (1993). The biomass of an Indian monsoonal wetland before and after being overgrown with Paspalum distichum L. Vegetatio, 109(1), 81-90. [ Links ]
van Groenewoud, H. (1992). The robustness of correspondence, detrended correspondence, and TWINSPAN analysis. Journal of Vegetation Science, 3(2), 239-246. [ Links ]
Viña, G., Ramírez, A. G., Lamprea, L., Garzón, B., Schmidt-Mumm, U., Rondon, E., & Florez, C. (1991). Ecología de la Ciénaga de Zapatosa y su relación con un derrame de petróleo. Informe Control Ambiental. Cúcuta. [ Links ]
Wantzen, K. M., Junk, W. J., & Rothhaupt, K. O. (2008). An extension of the floodpulse concept (FPC) for lakes. Hydrobiologia, 613(1), 151-170. [ Links ]
Wantzen, K. M., Yule, C. M., Mathooko, J. M., & Pringle, C. M. (2008). Organic matter processing in tropical streams. In D. Dudgeon (Ed.), Aquatic ecology series. Tropical stream ecology (1st ed., pp. 43-64). London, Burlington, MA: Academic Press. [ Links ]
Warfe, D. M. & Barmuta, L. (2004). Habitat structural complexity mediates the foraging success of multiple predator species. Oecologia, 141(1), 171-178. [ Links ]
Warfe, D. M., Pettit, N. E., Davies, P. M., Pusey, B. J., Hamilton, S. K., Kennard, M. J., & Halliday, I. A. (2011). The ‘wet-dry’ in the wet-dry tropics drives river ecosystem structure and processes in northern Australia. Freshwater Biology, 56(11), 2169-2195. [ Links ]
Wiegleb, G. (1991). Die Lebens- und Wuchsformen der makrophytischen Wasserpflanzen und deren Beziehungen zur Ökologie, Verbreitung und Vergesellschaftung der Arten. Tuexenia, 11, 135-147. [ Links ]
1.Department of Limnology and Oceanography, University of Vienna, Althanstraße 14, A-1090 Vienna, Austria;udo.schmidt-mumm@univie.ac.at
2.Department of Limnology and Oceanography, University of Vienna, Althanstraße 14, A-1090 Vienna, Austria;georg.janauer@univie.ac.at
Received 29-IX-2013. Corrected 20-I-2014. Accepted 18-II-2014.


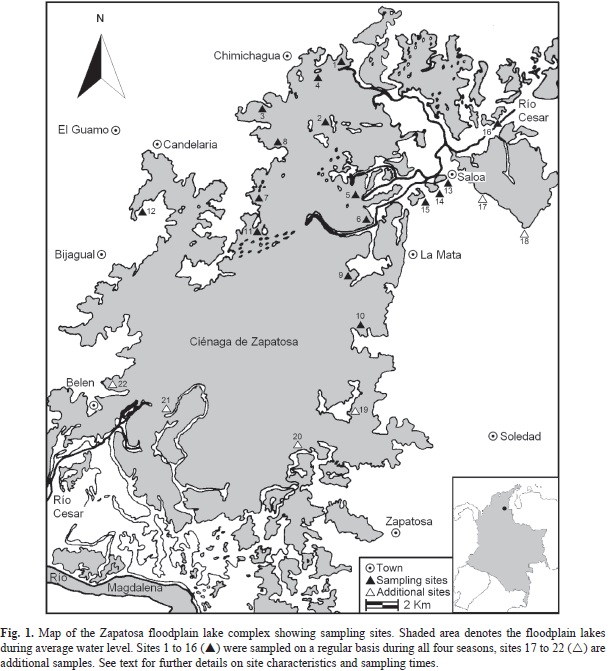{kind=link}
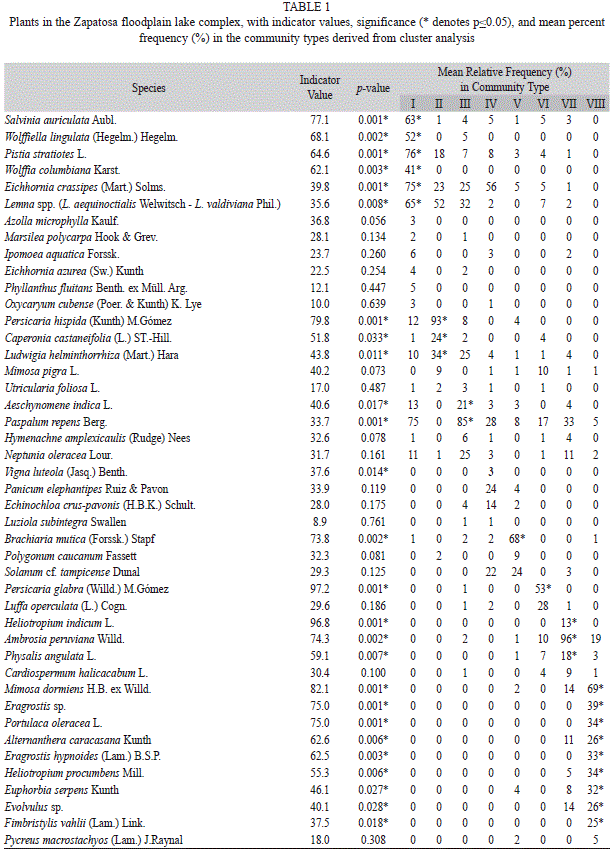{kind=link}
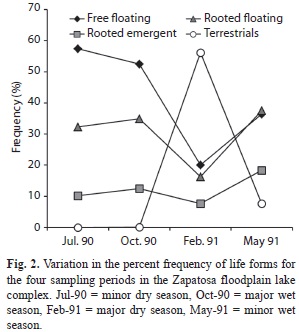{kind=link}
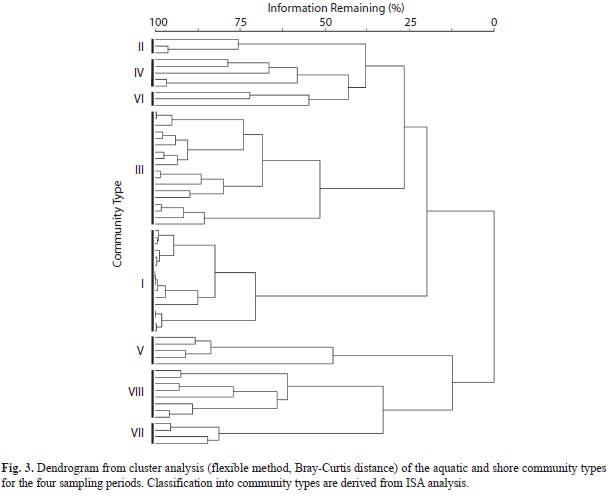{kind=link}
{kind=link}
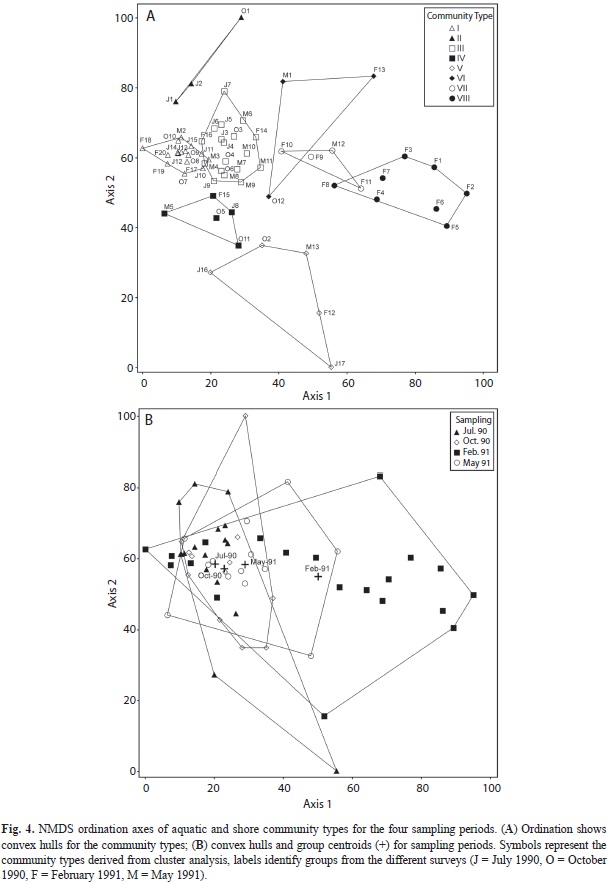{kind=link}
{kind=link}