










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.3 San José Jul./Sep. 2014
Macroperipatus ohausi: redescription and taxonomic notes on its status (Onychophora: Peripatidae)
Macroperipatus ohausi: redescripción y notas taxonómicas sobre su estatus (Onychophora: Peripatidae)
Macroperipatus ohausi: redescripción y notas taxonómicas sobre su estatus (Onychophora: Peripatidae)
Abstract
Onychophora have undergone a new wave of taxonomic revisions in the last three years. After a recent revision, only two species of Macroperipatus are registered from Brazil: Macroperipatus ohausi and Macroperipatus geayi. The former was only the second species of Onychophora to be described from Brazil, and is herein redescribed, illustrated, and its taxonomic status revised. Macroperipatus ohausi had only been collected at the type locality, Petrópolis, state of Rio de Janeiro, but has not been found there since its discovery. In 2009 we collected two specimens of this species in an Atlantic rain forest fragment in Nova Iguaçu in RPPN dos Petroleiros (also state of Rio de Janeiro). The color of the new specimens was interpreted and described based on the NBS/ISCC System of Color Designation, and corresponds only weakly to that described in earlier publications on M. ohausi. The identification of M. ohausi was based solely on its morphology and compared with its original description. In addition, a female syntype of M. ohausi deposited in the Zoologisches Museum (Hamburg) was examined. We reinvestigated its external morphology and concluded that the dorsal papillae have been misinterpreted in M. ohausi in previous studies of its integument. The new specimens examined are conspecific with M. ohausi mainly by having conical primary papillae, different sized accessory papillae, and by the number of legs. Our material resembles the original description of M. ohausi, for example, in spinous pad and nephridial tubercles, even though these features are not represented in the original description and rede- scription of M. ohausi; these features are further detailed here. We concluded that the dorsal papillae morphology of M. ohausi, shares diagnostic characters, for example, with representatives of the genera Peripatus and Epiperipatus. The dorsal papillae are rounded and not square as in the original description and redescription. Another argument that supports our conclusion is that the RPPN dos Petroleiros, the locality where the new samples were collected, belongs to the same mountain chain and Atlantic mosaic as the original material of M. ohausi. Furthermore, a male of M. ohausi is described here for the first time. Based on our morphological data, we transfered this species to the genus Epiperipatus, proposing a new combination, E. ohausi.
Key words: velvet worms, Peripatidae, taxonomy, geographical distribution, Brazil.
Resumen
Onychophora experimentó una nueva serie de revisiones taxonómicas en los últimos tres años. Después de una reciente revisión, sólo dos especies de Macroperipatus fueron registradas para Brasil: Macroperipatus ohausi y Macroperipatus geayi. La primera fue solo la segunda especie de Onychophora en ser descrita en Brasil y aquí es redescrita, ilustrada, y su estatus taxonómico revisado. Macroperipatus ohausi sólo se había recolectado en la localidad tipo, Petropolis, Estado de Rio de Janeiro, pero no se ha encontrado allí desde su descubrimiento. En 2009, recolectamos dos ejemplares de esta especie en un fragmento de la ‘Mata Atlantica’ en Nova Iguaçu en el RPPN de los Petroleiros (también en el estado de Rio de Janeiro). El color de los ejemplares nuevos fue interpretado y descrito, basado en el sistema NBS System/ISCC de designación de colores, y correspondió débilmente al descrito en publicaciones anteriores sobre M. ohausi. La identificación de M. ohausi se basó únicamente en su morfología, en comparación con la descripción original. Además, se examinó un sintipo hembra de M. ohausi depositado en Zoologisches Museum (Hamburgo). Nosotros reinvestigamos su morfología externa y concluimos que las papilas dorsales fueron mal interpretadas en M. ohausi en los estudios previos de su integumento. Los nuevos especímenes examinados son coespecíficos con M. ohausi principalmente por tener las papilas primarias cónicas, diferente tamaño de las papilas accesorias, y por el número de patas. Nuestro material se asemeja a la descripción original de M. ohausi, por ejemplo, la almohadilla espinosa y tubérculos nefridiales, aunque estas características no están representadas en la descripción original y redescripcion de M. ohausi, estas características también se detallan aquí. Llegamos a la conclusión de que la morfología de las papilas dorsales de M. ohausi, comparte caracteres de diagnóstico, por ejemplo, con los representantes de los géneros Peripatus y Epiperipatus. Las papilas dorsales son redondeadas y no cuadradas como en la descripción original y redescripción. Otro argumento que apoya nuestra conclusión es que la RPPN dos Petroleiros, la ubicación en la que se recogieron las nuevas muestras, pertenece a la misma cadena de montañas y al mosaico Atlántico, tal como el material original de M. ohausi. Por otra parte, se describe aquí por primera vez un macho de M. ohausi. Basado en nuestros datos morfológicos, transferimos esta especie al género Epiperipatus, que propone una nueva combinación, E. ohausi.
Palabras clave: gusanos de terciopelo, Peripatidae, taxonomía, distribución geográfica, Brasil.
Onychophorans, commonly referred to as velvet worms, are a small group of terrestrial invertebrates that are predominantly found in soil, leaf litter, or rotten wood. The species of Onychophora are arranged in two large families, which display a Pangean distribution (Morera-Brenes 2012, Murrienne et al. 2014). Currently 16 species are known from Brazil, belonging to four genera within the family Peripatidae (Peripatus Guilding, 1826, Epiperipatus Clark, 1913, Macroperipatus Clark, 1913, and Oroperipatus Cockerell, 1908). However, Sampaio-Costa, Chagas-Junior & Baptista (2009) recorded 24 morphospecies, of which half are undescribed.
The second oldest species described from Brazil is Peripatus ohausi Bouvier, 1900 col- lected in the city of Petrópolis (state of Rio de Janeiro). Bouvier (1900) gave a modest description of P. ohausi, but singular enough to distinguish it from other species known during that time. Its description was extended in Bouvier (1905) based only on the syntypes. Clark (1913) proposed the subgenus Macroperipatus to which P. ohausi was allocated. In 1975, Peck revised the New World Onychophora and considered Macroperipatus a valid genus of Peripatidae. In 1988, Read studied the integument of Macroperipatus ohausi (Bouvier, 1900) and showed that the base of the primary papillae is rounded rather than quadrangular, as in Macroperipatus torquatus (von Kennel, 1883), type species of Macroperipatus. Furthermore, Read (1988) proposed an approach to understand the morphology of the dorsal papillae using Scanning Electron Microscopy (SEM) in taxonomic studies of Peripatidae. He proposed to subdivide the genera of Peripatidae in three groups based on the scalation and mor- phology of dorsal papillae.
In a recent study of Brazilian peripatid onychophorans, Oliveira, Wieloch & Mayer (2010) revised two species previously classified as Macroperipatus –M. acacioi and M. machadoi Oliveira & Wieloch, 2005– and also showed that the base of the dermal papillae of these species are rounded rather than quadrangular, according to their original descriptions (Marcus & Marcus, 1955; Oliveira & Wieloch, 2005). In addition, the apical pieces of the primary papillae are well developed and, there- fore, they concluded that these two species were not representatives of Macroperipatus. Because of the presence of crural tubercles on only two pre-genital segments in males (Clark, 1913; Peck, 1975), the rounded dorsal papillae, and the low number of scale ranks in basal pieces of the primary papilla (Read, 1988), they included these two species in Epiperipatus.
As no specimen of M. ohausi had been collected in the last thirty years and its taxonomic status has not been reviewed, we redescribed and illustrated in this study its color in vivo, and the morphology of the dorsal integument using SEM. Furthermore, its distribution was extended and its taxonomic status revised.
Materials and Methods
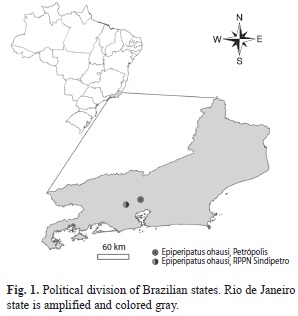
Material: The two specimens were col- lected in the Reserva Particular do Patrimônio Natural dos Petroleiros (RPPN dos Petroleiros) (22o35’44.83” S - 43o26’7.62” W), Nova Iguaçu, state of Rio de Janeiro (Brazil), about 28km from Petrópolis (Fig. 1). In respect of syntypes, some characters of the syntype (female) housed in the onychophoran collection of the Zoolo-gisches Museum Hamburg (Hamburg, Germany) were verified by Hilke Ruhberg (personal communication). Repository acronyms are as follows: MNRJ-Museu Nacional/UFRJ, Rio de Janeiro, State of Rio de Janeiro, Brazil and ZMH-Zoologisches Institut und Zoologisches Museum, Hamburg, Germany.
Fixation: Specimen (MNRJ 0056) was fixed directly in alcohol 70%. Specimen (MNRJ0058) was previously anesthetized in ethyl acetate and then fixed in alcohol 70%, according to the protocol of Monge-Nájera & Morera-Brenes (1994). After fixation, the specimens were sexed and examined under a stereomicroscope and with scanning electron microscope (SEM). SEM methods included: (1) Cleaning: pieces were rehydrated and cleaned with a 3% solution of Sodium Hypochlorite (NaClO) and distilled water. They were then dehydrated using an increasing alcoholic series: 30%, 50%, 70%, 80%, 90% and 100%. (2) Critical point: all the examined pieces with SEM were critical point dried in model Baltec CPD 030, and then covered by a coat of gold (approximately 200Å=2.10-10m) in Denton Desk Iv gold sputter model.
Nomenclature: Dental formula and descriptive terminology follows Morera- Brenes & Monge-Nájera (2010) and Oliveira et al. (2010), respectively.
Color study: Pictures were taken with a Fuji Finepix S1500 (4-mega pixels) camera. Coloration was described based on living specimens. Following Kury & Orrico (2006), standard names of the 267 Color Centroids of NBS/ ISCC Color System (Mundie, 1995) were used in descriptions and given between parentheses.
Results
Family Peripatidae
Genus Epiperipatus Clark, 1913
Epiperipatus ohausi (Bouvier, 1900) new combination
Peripatus ohausi Bouvier, 1900: 67, 1905: 204.
Peripatus (Macroperipatus) ohausi: Clark, 1913: 17.
Macroperipatus ohausi: Peck 1975: 347; Sampaio-Costa et al. 2009: 558; Oliveira et al. 2012:16.
Fig. 2-12.
Type material: (Syntypes: two females (ZMH and MNHN), BRAZIL, Petrópolis, 31. I. 1900, Dr. Ohaus leg. (Not examined).
Additional material examined: BRAZIL, state of Rio de Janeiro, Nova Iguaçu, Reserva Particular do Patrimônio Natural dos Petroleiros, 23.XII.2009, Guipponi, A.L.; Sampaio-Costa, C. leg. (1 female, MNRJ 0056); idem, 11.III.2010, Chagas-Jr, A.; Giupponi, A.L.; Kury, A.; Sampaio-Costa, C. leg. (1 male, MNRJ 0058).
Diagnosis: Antennae with 38 to 41 complete rings; outer jaw blade composed of one long curved principal tooth and an additional accessory tooth; inner jaw blade with one principal tooth, one accessory tooth, a diastema, and a series of 12 denticles; dorsal papillae rounded; basal piece of dorsal primary papillae conical; basal and apical pieces of dorsal pri- mary papillae with 7 to 8 and 3 to 4 scale ranks, respectively; leg pairs 27 to 29; ventral organs visible; anal glands of male indistinguishable.
Redescription
Measurements: Female (MNRJ 0056) 54mm long and 5mm wide, and male (MNRJ 0058) 29mm long, 3mm wide. Type (ZMH) 44mm long (syntype).
Color (for living specimens): Body background brownish gray (64) with dorsal light spots (primary papillae) pinkish gray (10). Dorsomedian furrow is pale violet (214), with light olive gray parallel diamonds (112). The head is moderate brown (58) and antennae are grayish to reddish purple (245) (Fig. 2). Dorsal side of legs is grayish to purplish pink (253), ventral side is pale purple (227), and the soles are yellowish white (92). The ventral side is dark pink (6) with light pink (4) ventral organs and light spots pinkish white (9), randomly dis- tributed (Fig. 3). One syntype (ZMH) is totally discolored.
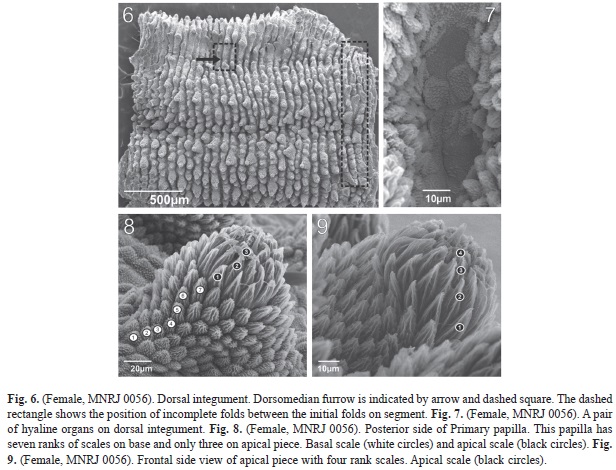
Integument: Twelve plicae per segment, seven of these cross over to ventral side. The incomplete plicae are interrupted over the legs (Fig. 6).
Hyaline organs: On dorsal side, between plicae and parallel to the dorsomedian furrow (Fig. 7).
Dorsomedian furrow: Slender, and starts at mid-point of head and finishes near anus.
Dorsal papillae: Female (MNRJ 0056). The dorsal papillae are close to each other and the size varies. Indistinct grooves can be present between two dorsal papillae (Fig. 6). The primary papillae are the biggest dorsal papillae (Fig. 6, 8 and 9); they are rarely in sequence or separated by accessory papillae. The number of accessory papillae varies between two primary papillae. The longest primary papillae are discolored with base diameter larger than plicae. These papillae are always found on alternate plicae in “longitudinal lines” present on dif- ferent aspects of dorsal side. Depending on its relative position to the dorsomedian furrow, these lines are composed of a different quantity of the largest primary papillae.
The conical basal piece of the primary papillae has seven (Fig. 8) or nine scale ranks. But nine scale ranks were observed only in the female (MNRJ 0056). These scales are lanceolate and increase in length near the top (Fig. 8). Constriction between the base and top are evident. The apical piece is spherical, asymmetrical and robust. The scales are narrow and as elongated as the taller scales of the base. The scales around the bristle are smaller and truncated (Fig. 8 and 9). The apical piece has three scale ranks on posterior side and four on anterior side.
The accessory papillae are conical and the smallest are the dorsal papillae. Usually the accessory papillae have many in dorsal plicae where the largest primary papillae are present (Fig. 6). The scales on the bottom are more elongated than on top.
Head. Without evident structures. Antennae. 38 to 41 complete rings. One syntype (ZMH) has 38 rings. In the collected female (MNRJ 0056) and male (MNRJ 0058) the first seven rings belong to the antennal tip. On both specimens between the 8th and 30th antennal rings there is a sequence of narrow, even and large odd rings. Frontal organs. Present at antennal base on ventral side with length about equal to four fused antennal papillae. Eyes. One on each antennal base. Mouth parts. Opening surrounded by one anterior oral lobe (unpaired lip) and seven pairs of oral lips. In each pair of oral lip there is a median series of small spines. One syntype (ZMH) has seven pairs of lips. One collected specimen (MNRJ 0056) presents a median series with 9 small spines, whereas the other (MNRJ 0058) presents 7 small spines. Jaws. (MNRJ 0056). Outer jaw blade. Composed of one long curved principal tooth and an additional accessory tooth (formula: 1/1) (Fig. 4). The accessory tooth is knife-shaped, curved at the top edge and straight at the bottom edge. Inner jaw blade. With one principal tooth, one accessory tooth, a diastema, and a series of 12 denticles (formula: 1/1/12) (Fig. 4 and 5). The accessory tooth of both blades is similar in shape.
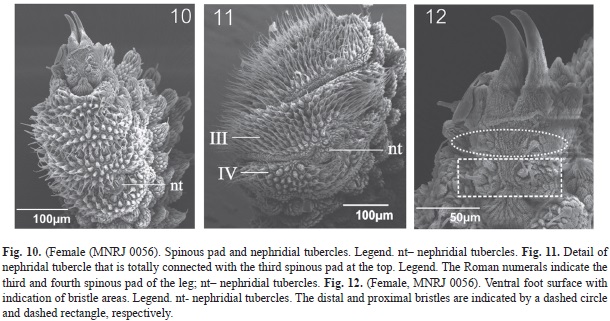
Legs: Female with 29 and male with 28 pairs of legs. Spinous pads. Fourth and fifth legs have four spinous pads with similar width, and length increases between the first three pads (Fig. 10 and 11). The length of the fourth spinous pad is similar to that of the second. Nephridial tubercle. Present on fourth and fifth pairs of legs, between 3rd and 4th spinous pads. It is totally connected with the third pad, articulating with the fourth, but is free from it (Fig. 10 and 11). Foot papillae. Feet have two anterior and one posterior foot papillae. ventral foot surface with six bristles: two pairs in proximal setiform ridge, and at least one bristle on each distal setiform ridge (Fig. 12). Ventral organs. They are clearly visible in vivo. Post-fixation they are represented by depressions on the ventral integument. Genital opening. Located between penultimate pair of legs in both sexes, but the morphology of the genital opening is different among the sexes. The genital opening in female is poorly preserved, but it seems to have two crossed furrows; the transversal one is only weakly visible. Moreover, the male has a single long furrow parallel to the main body.
Male characters: Crural papillae are pres- ent in one pre-genital pair of legs (a single tubercle per leg). Anal glands are only recog- nized by their corresponding open pores.
Type locality: Petrópolis, state of Rio de Janeiro, Brazil.
Distribution: State of Rio de Janeiro (Petrópolis and Nova Iguaçu, new record), Brazil.
Discussion
The Neotropical Onychophora are traditionally divided into two well-characterized groups named Andean and Caribbean peripatids. Our investigation shows that the two specimens reported in this study are peripatid onychophorans based on simple Caribbean group characteristics: (1) two anterior and one posterior foot papillae; (2) legs Iv and v with four spinous pads; and (3) nephridial tubercle openings between the 3rd and 4th pad. These characters are present in all representatives of Peripatus, Epiperipatus, Macroperipatus, Speleoperipatus Peck, 1975 and Plicatoperipatus Clark, 1913, but the presence of eyes and 12 plicae per segment distin- guishes them from Speleoperipatus and Plicatoperipatus, respectively.
Because of the conical basal piece with a roundish base and the spherical, asymmetrical and robust apical piece, the primary papillae of the analyzed specimens suggest a higher resemblance to Peripatus and Epiperipatus than to Macroperipatus. According to Read (1988), the quadrangular shape of dorsal papillae seen in M. torquatus is caused by the outline grooves. In addition, the scales of the basal piece are not arranged in ranks in this species (Oliveira et al., 2010). Therefore analyzed specimens do not belong to Macroperipatus.
The dorsal integument of all Macroperipatus species is not known in detail, although according to Read (1988) the dorsal primary papillae of M. acacioi, M. ohausi and M. guianensis (Evans, 1903) are rounded and contrast with quadrangular primary papillae of M. torquatus. In a recent review of two Brazilian species previously regarded as Macroperipatus–M. acacioi and M. machadoi– Oliveira et al. (2010) also showed that in these species the bases of the dermal papillae are roundish and covered with well-developed conical scales, which are arranged in several ranks. Therefore, Oliveira et al. (2010) transferred both species to the genus Epiperipatus. They also suggested that the dorsal papillae should be reinvestigated in other species of Macroperipatus by SEM.
Read (1988) registered one M. ohausi specimen, collected in 1981 by J. C. Diniz and C. R. Brandão, in Petrópolis. We contacted C. R. Brandão and he informed us of the exact collection point where the specimen was sampled (C. R. Brandão pers. comm.). We returned several times to this locality, but today this area is completely urbanized and we did not find any onychophoran there.
The two new specimens were collected 28km from Petrópolis, type locality of M. ohausi (Fig 1). The characters present in these specimens agree with the description of M. ohausi sensu Bouvier (1900, 1905), except for the number of leg pairs. The number of leg pairs is not fixed in Neotropical Peripatidae species (Bouvier, 1905; Campiglia & Laval- lard, 1973). The number of leg pairs in our specimens is 28 and 29, and the type is 27. Bouvier (1900, 1905) poorly described M. ohausi, based mainly on the description of the primary papillae morphology. Then, Read (1988) included the number of scale ranks in the description of this species based on recent specimens collected in 1981 by J. C. Diniz and C. R. Brandão in Petrópolis.
The dorsal primary papillae described by Read (1988, figs. A-D) were compared to the specimens from RPPN dos Petroleiros and also agree with them in morphology. The morphology of the dorsal primary papillae and the range of leg pairs show that Epiperipatus ohausi is more closely related to the species of Epiperipatus from the state of Minas Gerais (Southeastern Brazil): E. acacioi, E. machadoi, E. diadenoproctus Oliveira, Lacorte, Fonseca, Wieloch & Mayer, 2011; E. adenocryptus Oliveira et al. 2011; and E. paurognostus Oliveira et al. 2011. Epiperipatus ohausi has seven or eight scale ranks on the base of the primary papillae, while E. acacioi, E. machadoi, E. diadenoproctus, E. adenocryptus have 5-6, 5-8, 5-8 and 5-7, respectively. Four scale ranks on E. adenocryptus, three on E. ohausi and E. acacioi, and two on E. paurognostus were counted from the posterior side of the apical piece, but for E. diadenoproctus it was not described.
Epiperipatus ohausi, E. acacioi and E. machadoi have the asymmetrical apical piece in common, though it is spherical in the first species and ovate in latter two (although some E. acacioi specimens have a globular shape as well). The sensory bristles usually are needle-shaped and curved posteriorly, as observed in E. acacioi, E. machadoi and E. ohausi. However, it is spine-shaped in the cryptic species from Minas Gerais (Oliveira et al., 2011). The range of leg pairs is different among these six Brazilian Southeastern species. In increasing order, the ranges are: 27 to 29 in E. ohausi; 25 to 28 in E. acacioi (n=34); 26 to 29 in E. paurognostus (n=15, see Oliveira et al., 2011); 26 to 30 in E. adenocryptus (n=25, see Oliveira et al., 2011); E. diadenoproctus (n=62, see Oliveira et al., 2011), and 27 to 31 in E. machadoi (n=21). The anal glands are absent in males of E. ohausi, but the respective pores are present. These pores are observed on the ventral board of the anus. In E. acacioi and E. machadoi the anal glands seem also to be absent, while in E. adenocryptus, E. diadenoproctus and E. paurognostus these structures are present in the ventral board of the anus. The SEM primary papillae (Fig. 5, 7 and 8) analysis of M. ohausi showed that the base is not square-shaped as in M. torquatus, but rounded as in Peripatus and Epiperipatus.
The new record of E. ohausi has prompted us to redescribe and illustrate the color and integument of the species in detail. Furthermore, we reviewed the taxonomic status of this poorly known species.
Acknowledgments
We are grateful to Amanda Garcez da veiga for SEM operation at the Center for Scanning Electron Microscopy of the Museu Nacional, UFRJ. The establishment of this Center was made possible by a grant from Cenpes/Petrobras, and is part of the company’s Thematic Network for Marine Environmental Monitoring. We are grateful to Dr. Hilke Ruhberg for providing information about the type deposited in the Hamburg Museum. We are also grateful to Nivaldo Alves da Silva (present Sindipetro director) and Odirlei vieira da Fonseca for helping us find and collect onychophorans in RPPN dos Petroleiros. We are grateful for the additional comments made by colleagues who read the manuscript. This research was partially supported by grants from Fundação de Amparo à Pesquisa do Esta- do de São Paulo (process 2010/10388-8 and 2011/20211-0), awarded to both authors. This research is also part of the Project “Aracnídeos e Miriápodes da Mata Atlântica” (AMMA), of the ARACNOLAB (Laboratory of arachnids of the Museu Nacional/UFRJ).
References
Bouvier, E. L. (1900). Nouveau Péripate des environs de Rio-de-Janeiro. Bulletin de la Société Entomologique de France, 3, 66-68. [ Links ]
Bouvier, E. L. (1905). Monographie des Onychophores. Annales des Sciences Naturelles, Zoologie (9e Sér.), 2(1-3), 1-383. [ Links ]
Campiglia, S. & Lavallard, R. (1973). Contribution a la biologie de Peripatus acacioi Marcus et Marcus: II. variations du poids des animaux en fonction du sexe et du nombre des lobopodes. Boletim de Zoologia e Biologia Marinha [Nova Série], 30, 499-502. [ Links ]
Clark, A. H. (1913). A revision of the American species of Peripatus. Proceedings of the Biological Society of Washington, 26, 15-19. [ Links ]
Cockerell, T. D. A. (1908). Review of Monographie des Onychophores by E.L. BOUVIER. Science 27(694), 619-621. [ Links ]
Evans, R. (1903). On Peripatus guianensis (sp. nov.). Quarterly Journal of Microscopical Science, 47(186), 145-160. [ Links ]
Guilding, L. (1826). Mollusca Caribbaeana. Zoologica Journal, 2, 437-449. [ Links ]
Kury, A. B. & Orrico, v. G. D. (2006). A new species of Lacronia Strand, 1942 from the highlands of Rio de Janeiro (Opiliones, Gonyleptidae, Pachylinae). Revista Ibérica de Aracnologia, Zaragoza, 13, 147-153. [ Links ]
Marcus, E. & Marcus, E. (1955). A new Peripatus from Minas Gerais, Brazil. Anais da Academia Brasileira de Ciências, 27(2), 189-193. [ Links ]
Monge-Nájera, J. & Morera-Brenes, B. (1994). Morphological and physiological characteristics of two species of Epiperipatus from Costa Rica (Onychophora: Peripatidae). Revista de Biologia Tropical, 42(1/2), 181-188. [ Links ]
Morera-Brenes, B. & Monge-Nájera, J. (2010). A new giant species of placented worm and the mechanism by which onychophorans weave their nets (Onychophora: Peripatidae). Revista de Biología Tropical, 58(4), 1127-1142. [ Links ]
Morera-Brenes, B (2012). Los onicóforos, fósiles caminantes. El Salvador Ciencia & Tecnología, 17(23), [ Links ] 8-13.
Mundie, D. A. (1995). The NBS/ISCC Color System/ David A. Mundie Pittsburgh, PA: Polymath Systems 535.6 dc- 20. Retrieved from http://www.anthus.com/Colors/Cent.html. [ Links ]
Murienne, J., Daniels, S. R., Buckey, T.R., Mayer, G. & Giribet, G. (2014). A living fossil tale of Pangean biogeography. Proceedings of the Royal Society B: Biological Sciences, 281: 20132648. [ Links ]
Oliveira, I. & Wieloch, A. H. (2005). Macroperipatus machadoi sp. n. (Onychophora: Peripatidae) da Flo- resta Atlântica de Minas Gerais, Brasil. Lundiana 6(supp.), 61-66. [ Links ]
Oliveira, I., Read, v. M. S. J., & Mayer, G. (2012). A world checklist of Onychophora (velvet worms), with notes on nomenclature and status of names. ZooKeys, 211, 1-70. [ Links ]
Oliveira, I., Wieloch, A. H., & Mayer, G. (2010). Revised taxonomy and redescription of two species of the Peripatidae (Onychophora) from Brazil: a step towards consistent terminology of morphological characters. Zootaxa, 2493, 16-34. [ Links ]
Oliveira, I., Lacorte, G. A., Fonseca, C. G., Wieloch, A. H., & Mayer, G. (2011). Cryptic speciation in Brazilian Epiperipatus (Onychophora: Peripatidae) reveals an underestimated diversity among the peripatid velvet worms. Plos one, 6(6), e19973. [ Links ]
Peck, S. B. (1975). A review of the New World Onychophora with the description of a new cavernicolous genus and species from Jamaica. Psyche, 82(3-4), 341-358. [ Links ]
Read, v. M. S. J. (1988). The application of scanning electron microscopy to the systematics of the neotropical Peripatidae (Onychophora). Zoological Journal of the Linnean Society of London, 93(3), 187-223. [ Links ]
Sampaio-Costa, C., Chagas-Junior, A., & Baptista, R. L. C. (2009). Taxonomy and distribution of Brazilian Onychophora. Revista Brasileira de Zoologia, 26(3), 553-561. [ Links ]
von Kennel, J. (1883). Entwicklungsgeschichte von Peripatus. Zoologischer Anzeiger, 6(150), 531-537. [ Links ]
Bouvier, E. L. (1905). Monographie des Onychophores. Annales des Sciences Naturelles, Zoologie (9e Sér.), 2(1-3), 1-383. [ Links ]
Campiglia, S. & Lavallard, R. (1973). Contribution a la biologie de Peripatus acacioi Marcus et Marcus: II. variations du poids des animaux en fonction du sexe et du nombre des lobopodes. Boletim de Zoologia e Biologia Marinha [Nova Série], 30, 499-502. [ Links ]
Clark, A. H. (1913). A revision of the American species of Peripatus. Proceedings of the Biological Society of Washington, 26, 15-19. [ Links ]
Cockerell, T. D. A. (1908). Review of Monographie des Onychophores by E.L. BOUVIER. Science 27(694), 619-621. [ Links ]
Evans, R. (1903). On Peripatus guianensis (sp. nov.). Quarterly Journal of Microscopical Science, 47(186), 145-160. [ Links ]
Guilding, L. (1826). Mollusca Caribbaeana. Zoologica Journal, 2, 437-449. [ Links ]
Kury, A. B. & Orrico, v. G. D. (2006). A new species of Lacronia Strand, 1942 from the highlands of Rio de Janeiro (Opiliones, Gonyleptidae, Pachylinae). Revista Ibérica de Aracnologia, Zaragoza, 13, 147-153. [ Links ]
Marcus, E. & Marcus, E. (1955). A new Peripatus from Minas Gerais, Brazil. Anais da Academia Brasileira de Ciências, 27(2), 189-193. [ Links ]
Monge-Nájera, J. & Morera-Brenes, B. (1994). Morphological and physiological characteristics of two species of Epiperipatus from Costa Rica (Onychophora: Peripatidae). Revista de Biologia Tropical, 42(1/2), 181-188. [ Links ]
Morera-Brenes, B. & Monge-Nájera, J. (2010). A new giant species of placented worm and the mechanism by which onychophorans weave their nets (Onychophora: Peripatidae). Revista de Biología Tropical, 58(4), 1127-1142. [ Links ]
Morera-Brenes, B (2012). Los onicóforos, fósiles caminantes. El Salvador Ciencia & Tecnología, 17(23), [ Links ] 8-13.
Mundie, D. A. (1995). The NBS/ISCC Color System/ David A. Mundie Pittsburgh, PA: Polymath Systems 535.6 dc- 20. Retrieved from http://www.anthus.com/Colors/Cent.html. [ Links ]
Murienne, J., Daniels, S. R., Buckey, T.R., Mayer, G. & Giribet, G. (2014). A living fossil tale of Pangean biogeography. Proceedings of the Royal Society B: Biological Sciences, 281: 20132648. [ Links ]
Oliveira, I. & Wieloch, A. H. (2005). Macroperipatus machadoi sp. n. (Onychophora: Peripatidae) da Flo- resta Atlântica de Minas Gerais, Brasil. Lundiana 6(supp.), 61-66. [ Links ]
Oliveira, I., Read, v. M. S. J., & Mayer, G. (2012). A world checklist of Onychophora (velvet worms), with notes on nomenclature and status of names. ZooKeys, 211, 1-70. [ Links ]
Oliveira, I., Wieloch, A. H., & Mayer, G. (2010). Revised taxonomy and redescription of two species of the Peripatidae (Onychophora) from Brazil: a step towards consistent terminology of morphological characters. Zootaxa, 2493, 16-34. [ Links ]
Oliveira, I., Lacorte, G. A., Fonseca, C. G., Wieloch, A. H., & Mayer, G. (2011). Cryptic speciation in Brazilian Epiperipatus (Onychophora: Peripatidae) reveals an underestimated diversity among the peripatid velvet worms. Plos one, 6(6), e19973. [ Links ]
Peck, S. B. (1975). A review of the New World Onychophora with the description of a new cavernicolous genus and species from Jamaica. Psyche, 82(3-4), 341-358. [ Links ]
Read, v. M. S. J. (1988). The application of scanning electron microscopy to the systematics of the neotropical Peripatidae (Onychophora). Zoological Journal of the Linnean Society of London, 93(3), 187-223. [ Links ]
Sampaio-Costa, C., Chagas-Junior, A., & Baptista, R. L. C. (2009). Taxonomy and distribution of Brazilian Onychophora. Revista Brasileira de Zoologia, 26(3), 553-561. [ Links ]
von Kennel, J. (1883). Entwicklungsgeschichte von Peripatus. Zoologischer Anzeiger, 6(150), 531-537. [ Links ]
1. Departamento de Biologia e Zoologia, Instituto de Biociências, Universidade Federal de Mato Grosso. Avenidan Fernando Correa da Costa, 2367, Boa Esperança, CEP-78060-900, Cuiabá, MT, Brasil; amazonaschagas@gmail.com
2. Departamento de Zoologia, Instituto de Biociências, Universidade de São Paulo. Rua do Matão-Travessa 14, nº 321, Cidade Universitária, São Paulo, SP, Brasil. CEP 05508-090; csampaioc@gmail.com
Received 20-vIII-2013. Corrected 06-Iv-2014. Accepted 05-v-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}