










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.3 San José Jul./Sep. 2014
Bio-filtration capacity, oxygen consumption and ammonium excretion of Dosinia ponderosa and Chione gnidia (Veneroida: Veneridae) from areas impacted and non-impacted by shrimp aquaculture effluents
Bio-filtración, consumo de oxígeno y excreción amoniacal de Dosinia ponderosa y Chione gnidia (Veneroida: Veneridae), en áreas impactadas y no impactadas por efluentes de granjas camaroneras
Bio-filtración, consumo de oxígeno y excreción amoniacal de Dosinia ponderosa y Chione gnidia (Veneroida: Veneridae), en áreas impactadas y no impactadas por efluentes de granjas camaroneras
Karime Ramos-Corella1*, Luis Rafael Martínez-Córdova2*, Luis Fernando Enríquez-Ocaña2, Anselmo Miranda-Baeza3* & José Antonio López-Elías2
Abstract
Mollusks are some of the most important, abundant and diverse organisms inhabiting not only aquatic ecosystems, but also terrestrial environments. Recently, they have been used for bioremediation of aquaculture effluents; nevertheless, for that purpose it is necessary to analyze the capacity of a particular species. In this context, an experimental investigation was developed to evaluate the performance of two bivalves C. gnidia and D. ponderosa, collected from areas with or without shrimp aquaculture effluents. For this, the filtration capacity (as clearance rate) as well as the oxygen consumption and ammonia excretion rates were measured following standard methods. The clearance rate was significantly higher for D. ponderosa from impacted areas, when com- pared to C. gnidia, from both areas. Contrarily, the oxygen consumption was greater for C. gnidia from impacted areas compared to D. ponderosa from both areas. The same tendency was observed for the ammonia excretion with the highest rates observed for C. gnidia from impacted areas, whereas no differences were observed among D. ponderosa from both areas. The results suggest that both species developed different strategies to thrive and survive under the impacted conditions; D. ponderosa improved its filtration efficiency, while C. gnidia modified its oxygen consumption and ammonia excretion. We concluded that both species, and particularly D. ponderosa, can be used for bioremediation purposes.
Key words: mollusks, bio-filtration, environmental impact, shrimp aquaculture.
Resumen
Los moluscos son algunos de los organismos más importantes, abundantes y diversos que habitan no solo ecosistemas acuáticos sino también terrestres. Recientemente ellos han sido utilizados para la biorremediación de efluentes acuícolas; para este propósito, es necesario conocer la capacidad de especies particulares que funcionan como biorremediadores. En este contexto, se evaluó la eficiencia de filtración (medida como tasa de clarificación), así como las tasas de consumo de oxígeno y excreción amoniacal en los bivalvos D. pon- derosa y C. gnidia recolectados en áreas impactadas y no impactadas por efluentes de granjas camaroneras. La tasa de clarificación fue mayor para D. ponderosa procedente de áreas impactadas, comparada con la de C. fluctifraga en las dos áreas de recolecta. Contrariamente, la tasa de consumo de oxígeno fue superior en C. gnidia en las áreas impactadas al compararla con organismos de áreas no impactadas y con D. ponderosa de las dos áreas de recolecta. La tasa de excreción amoniacal siguió una tendencia similar con valores más altos para C. gnidia en áreas impactadas, mientras que no se observaron diferencias para D. ponderosa entre las áreas de recolecta. Los resultados sugieren que ambas especies desarrollan diferentes estrategias para adaptarse y sobrevivir bajo condiciones de impacto; D. ponderosa mejora su eficiencia de filtración y C. gnidia modifica su consumo de oxígeno y excreción amoniacal. Se concluye que ambas especies, pero sobre todo D. ponderosa pueden ser utilizadas con propósitos de biorremediación.
Palabras clave: acuacultura, bio-filtración, impacto ambiental, moluscos.
Aquaculture has continued growing world- wide in the new century. In only 50 years, its production passed from being almost insignificant to be equivalent to the fishery industry, and in 2010 it reached unprecedented volumes of more than 60 million tonnes (FAO, 2012). Regarding bivalve aquaculture (mainly filter-feeding), its production has also grown dramatically, quadruplicating the volumes in 30 years as reported by FAO (2012). Up to 2010, non-fed species represented one third of the global aquaculture production (FAO, 2012). As the world population grows, the demand for aquaculture products, including mollusk species, has been also growing in a similar proportion.
Despite their evident benefits, aquaculture is one of the most criticized activities world- wide, mainly because of the environmental impacts produced by their effluents in the receiving ecosystems, and causing eutrophication, hyper-nutrification, burrowing of benthic communities and the constant occurrence of epizooties (Martínez-Córdova, Martínez-Porchas, & Cortés-Jacinto, 2009; Martínez-Porchas & Martinez-Córdova, 2012).
Many strategies have been proposed or proven to minimize the effect of aquacul- ture effluents including: sedimentation lagoons (Martínez-Córdova & Enriquez-Ocaña, 2007), low or zero water exchange (Balasubramanian, Pillai, & Ravichandran, 2005), recirculation systems (Piedrahita, 2003), mangroves as nutrient sinks (Rivera-Monroy, Torres, Bahamon, Newmark, & Twilley, 1999), polyculture practices (Martínez-Cordova & Martinez-Porchas, 2006; Martínez-Porchas & Martinez-Cordova, 2012), and bioremediation (Paniagua-Michel & Garcia, 2003).
One of the most promising bioremediation strategies is the use of bivalve mollusks to remove suspended solids from the water column (Chopin, et al., 2001; Miranda, Lizarraga-Armenta, Rivas-vega, López-Elías, & Nieves-Soto, 2010; Peña-Messina, Martínez-Cordova, Bücke-Ramirez, Segovia-Quintero, & Zertuche-González, 2009; Martínez-Córdova, López-Elías, Martínez-Porchas, Bernal- Jaspeado, & Miranda-Baeza, 2011).
Mollusks are some of the most important, abundant and diverse organisms inhabiting not only aquatic ecosystems, but also terrestrial environments. In particular, bivalves are exclusively aquatic and most of them are filter- feeders; this means that they fed by filtration of water from which retain the organic nutritive portion, and discriminate and bio-deposit the inorganic fraction. Such capacity, make some bivalves good candidates for effluents bioremediation of aquaculture activities (Chavez- Croker & Oberque-Contreras, 2010).
The Gulf of California in Northwestern Mexico has a great diversity of bivalve mollusks (Keen, 1971); some of them have been evaluated as prospects for bioremediation of aquaculture effluents. Peña-Messina et al. (2009) evaluated the physiological filtration variables for Crassostrea cortesiensis and Anadara tuberculosa farmed in shrimp aquaculture effluents, and found that both species are good candidates to be considered for use as biofilters in aquaculture bioremediation. Martínez-Córdova, López-Elías, Leyva-Miranda, Armenta-Ayon, & Martinez-Porchas (2011) successfully used Chione fluctifraga for the bioremediation of shrimp farming effluents. Nieves-Soto, Enríquez-Ocaña, Piña-valdez, Maeda-Martínez, Almodóvar-Cebreros, & Acosta-Salmón (2011) found that A. tuberculosa can be grown in challenging environments such as shrimp ponds effluents and take advance of the cockle tolerance to extreme conditions. Enriquez-Ocaña, Nieves-Soto, Piña-valdez, Martínez-Córdova and Medina- Jasso (2012) evaluated the combined effect of temperature and salinity on the biofiltration capacity of C. corteziensis and found this bivalve have a greater filtration capacity at 32°C and 35PSU. Nieves-Soto, et al. (2013) documented the filtration and clarification rates and assimilation efficiency of Atrina tuberculosa under different combinations of temperature and food concentration.
Some coastal areas of the Gulf of California are now being impacted by shrimp aqua- culture effluents affecting some environmental variables such as salinity, dissolved oxygen, total and organic suspended solids concentration (turbidity) among others. Diverse species of bivalves thrive in those zones, and in spite of the impact, they exhibit a relatively good survival (Peña-Messina et al., 2009; Nieves-Soto et al., 2011). It is plausible to suppose that such species are adapted to those conditions by the development of some filtration strategies which permit them to thrive adequately. However, such conditions could also affect the oxygen consumption capacity and the ammonia excretion rate, which are considered as metabolism and stress indicators. The oxygen consumption in mollusks is affected by many diverse factors such as temperature, salinity and dissolved oxygen (Shumway & Koehn, 1982), as well as the concentration of suspended solids and turbidity (Alexander, Thorp, & Fell, 1994). The ammonia excretion could be affected by temperature (Saucedo, Ocampo, Monteforte, & Bervera, 2004), salinity (Bartberger & Pierce, 1976), and exposure to air (De vooys & De Zwaan, 1978). It has been demonstrated that ammonia excretion is closely related to stress- ful conditions such as those prevailing in impacted areas (Widdows et al., 1981, Comoglio, Gaxiola, Roque, Cuzon, & Amin, 2002).
The present study was focused on evaluating the filtration capacity (measured as the clarification rate), oxygen consumption and ammonia excretion of two of the most com- mercially-important bivalves, D. ponderosa and C. gnidia, collected from impacted and non-impacted areas of the coastal zone of Northwest Mexico.
Materials and Methods
Mollusk specimens were collected from two sites of the coastal zone, one impacted (28°40’2.58” N - 111°51’55.98” W), and the other non-impacted (28°56’1.80” N- 112°5’37.32” W) by shrimp aquaculture effluents. The samplings were made by scuba diving, using a manual hackle. It was not possible to collect organisms of similar sizes from both areas; in the non-impacted area only small specimens of D. ponderosa were found, while in the impacted area, only big organisms were possible to collect; contrarily, for C. gnidia smaller organisms were found in the impacted area as compared to the non-impacted.
A total of 40 individuals of each species were collected and immediately transported to the facilities of DICTUS, University of Sonora, México. These were placed into acclimation aquaria and maintained at 23°C temperature and 36psu salinity for seven days. The ethical rules to reduce both the stress and suffering of the studied organisms were observed during acclimation and experimentation. The clams were fed three times a day with a monoalgal culture of Chaetoceros muellerii from the same laboratory at a density of 100 000cel/mL. The water quality parameters (temperature, salinity, dissolved oxygen and pH), were monitored twice a day by means of a multi-parameter YSI Model 6600. The concentration of total ammonium nitrogen (TAN) was measured by spec-trophotometry using the Hach DR4000 and the routine and chemicals described in the manual.
After seven days, apparently healthy mollusks were randomly selected to be transferred to individual containers of different capacity (depending on the organisms size), to evaluate their filtration capacity as well as oxygen consumption and total ammonium excretion. The size for D. ponderosa from impacted areas ranged from 15 to 19cm, and from non impacted areas from 12 to 15cm. The size of C. gnidia from impacted areas ranged from 2.5 to 3.0cm; and from non impacted areas from 7.5 to 10.9cm. Both species reproduced along the year, mainly during last spring and the whole summer, which means that the selected individuals probably had different degree of gonadal development.
Clarification rates: The filtration capacity was evaluated by the measurement of the clarification rate. It was done by triplicate in a static system using experimental chambers of 5L for D. ponderosa and 2L for C. gnidia. One organism was placed into each one of the four cambers, and one more was used as control in which an empty valve of the corresponding organism was placed.
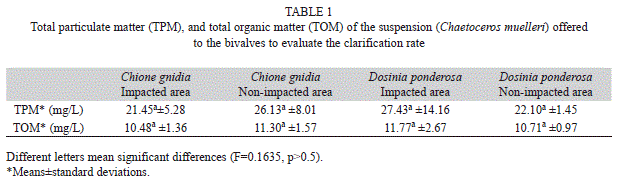
A suspension of the microalgae C. muelleri with an approximate cell density of 10x104cell/ mL were introduced in each chamber. Such low density was used to avoid the production of pseudo-feces (Berg, Fisher, & Landrum, 1996). The content of total particulate matter and organic matter of the suspension is presented in table 1. No significant differences in any of both parameters were observed (F=0.1635; p>0.05). To keep the suspension homogeneous, aeration through diffuser stones was applied, trying to maintain the diffusers not very close to the organisms to avoid stress (Fernández- Reiriz, Labarta, Albentosa, & Pérez-Camacho, 1998). A time of 40min was considered for the evaluation of the clarification rate of the clams, based on the experiences of previous studies in which was demonstrated that longer times may cause misestimations due to re-filtration processes (Riisgård, 2001). The time was counted from the moment in which the mollusks opened the valves and began to filtrate.
The clearancerate (CR) represents the water volume completely cleaned of suspended particles in a determined time, and was estimated as suggested by Jørgensen (1990) using the following formula:
CR= V [log Ci-log Cf·(0.434·T)-1] (1)
Where CR is the clarification rate in L/h per individual, v is the volume of the experimental chamber in L, Ci and Cf are the initial and final concentration of suspended particles, and T is the total time of clarification in hours. To get the dry weight, the clams were unshelled and the soft tissue weighed, dried for 48h in a stove at 90°C, and weighed again.
The concentration of suspended particles was measured in a Coulter Counter Beckman Z2.
The CR values were expressed per g DW through the following equation:
CR (g) = CR·(b 0.75)-1 (2)
Where CR (g) is the clarification rate standardized; CR is the clarification rate expressed per g DW, CR is the clarification rate as obtained from equation 1.
Oxygen consumption and ammonia excretion: To evaluate the oxygen consumption and ammonia excretion, the same organ- isms used for clarification rate (after 2 hours), were placed in containers hermetically closed, with oxygen saturated-seawater. The dissolved oxygen (DO), and total ammonia nitrogen (TAN) were measured at the beginning and after 4h; the DO by means of the multi-parameter YSI sonde and the TAN by spectrophotometry, using the HACH DR400 equipment.
The data of CR, DO and TAN were submitted to tests of normality of Lilliefors (Conover, 1999) and homoscedasticity of Bartlett (Zar, 1999). As the data were normal and homoscedastic, a two-way ANOVA was performed to establish differences among species and areas. A Tukey test was used to compare and rank the means.
Results
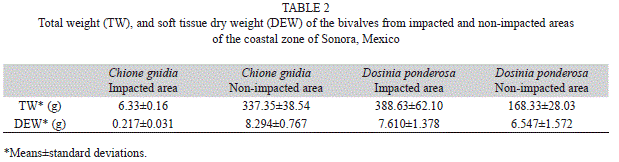
The means ± SD of total weight (TW) and dry soft tissue weight (DEW) of both species collected from the impacted and non-impacted areas are presented in table 2. The TW of C. gnidia was much greater for organisms from non-impacted areas as compared to those from impacted ones. Contrarily, D. ponderosa from impacted areas had a greater TW than the organisms from non-impacted areas. The same tendency was observed for the DEW, although for D. ponderosa, the differences were not significant.
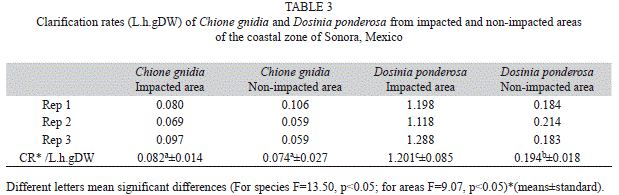
With regard to the clarification rate (CR), significant differences were found among the two evaluated species (Table 3). D. ponderosa showed much higher rates as compared to C. gnidia, independently of the areas. When compared the same species, but from different areas, C. gnidia did not observe differences in the CR, however D. ponderosa from impacted areas, had values much higher than those from non-impacted areas.
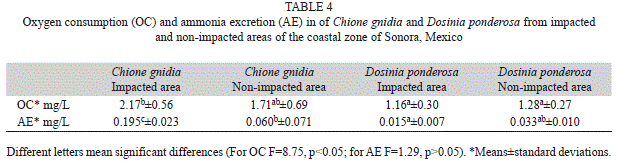
The means and SD of oxygen consumption and ammonia excretion recorded for the two species from both areas are presented in table 4. Significant differences in oxygen consumption were found among species, with C. gnidia from impacted areas, recording greater values than D. ponderosa from both areas. The same tendency was observed for ammonia excretion.
Discussion
When aquatic organisms with limited or any capacity of displacement are subjected to adverse conditions, they must develop strategies aimed to survive and thrive under those conditions. In the case of bivalve mollusks, some of these strategies could be the improvement of their filtration capacity, and modifications of the oxygen consumption and ammonia excretion. Nieves-Soto et al. (2013) demonstrated under laboratory conditions that the filtration and clarification rates of the pen shell A. tuberculosa varied significantly when temperature and food concentration were modified. They found that at 28°C (similar to the prevailing in our studied areas), the clarification rate was significantly higher when food concentration was higher. This suggest that bivalves we collected from impacted areas with high concentration of suspended solids (including microalgae), would have a higher clarification rate that those from non-impacted areas. This hypothesis was fulfilled only for D. ponderosa since the organisms from impacted areas recorded a mean CR more than six times greater than those from non-impacted areas. For C. gnidia the rates were very similar among both sites. A plausible explanation of these differences among species is the size and age of the organisms evaluated. The individuals of D. ponderosa were much larger and older (based on the growth rings) than the individuals of C. gnidia. This implies that the first were subjected for a longer time to the impacted conditions which probably allowed them to develop a better filtration strategy. Enriquez-Ocaña et al. (2012) reported for C. corteziensis a CR of 0.45mg/L.h.g.DW at 23°C and 35PSU. However, when temperature was 32°C the CR reached 1.607mg/L.h.g.DW. That last value was similar to the one found in the present study for D. ponderosa from the impacted areas, but much greater than that recorded for C. gnidia from the two areas, and D. ponderosa from non-impacted areas. Similar results were documented for A. tuberculosa by Nieves-Soto et al. (2011), who found high clarification rates at high temperature (30°C) and salinities of 30 and 40PSU. No specific studies approaching the filtration capacity of D. ponderosa have been reported in the scientific literature, which means that this is the first report about the subject.
The greater oxygen consumption (OC) recorded for C. gnidia when compared to D. ponderosa suggest that both species develop different strategies to thrive under impacted conditions; the first increased their FR and the second their OC. Alexander et al. (1994) reported that in the zebra mussel (Dreissena polymorpha), the OC was significantly affected by temperature and turbidity. Haure, Penisson, Bougrier, and Baud (1998) documented for Ostrea edulis, oxygen consumptions from 0.3 to 1.8mgO2/h.g, being greater at higher temperatures. The values we found are into this range except for C. gnidia from the impacted areas which was slightly higher (2.1mgO2/h.g).
The ammonia excretion (AE) of C. gnidia from the impacted areas was three fold-times greater compared to the same species from the non-impacted areas, while D. ponderosa had much lower rates. In this case it seems that the impacted condition of the areas influenced the AE only in C. gnidia but not in D. ponderosa. Since many years it is known that ammonia excretion is a response of the organisms to environmental stress and pollution (Bayne, Moore, Widdows, Livingstone, Salkeld, Crisp, Morris, Gray, Holden, Newell & McIntyre, 1979). Grant and Thorpe (1991) found that the soft-shell clam Mya arenaria, had a significant decrease in oxygen consumption and increase in ammonia excretion when was subjected to high turbidity. This finding partially coincide with our results of C. gnidia from the impacted areas, that with a high concentration of suspended solids (turbidity) showed a greater AE than the organisms collected from non- impacted areas. However, the OC results were different to those of the cited author.
As indicated by the results of the present study it can be concluded that the evaluated bivalves have developed different strategies to survive and thrive under environmental impact conditions produced by shrimp aquaculture discharges. D. ponderosa has improved its filtration capacity while C. gnidia has modified its oxygen consumption and ammonia excretion. These findings can be used to take advantage of the two species for practical purposes such as: bioremediation of aquaculture effluents, co-culture or polyculture with shrimp or any other species.
Acknowledgments
We want to thank two Mexican Government dependences: SEP-PROMEP and COFU- PRO INAPESCA for the financial support for this study.
References
Alexander, Jr. J. E., Thorp, J. H., & Fell, R. D. (1994). Tur- bidity and Temperature Effects on Oxygen Consumption in the Zebra Mussel (Dreissena polymorpha). Canadian Journal of Fisheries and Aquatic Sciences, 51, 179-184. [ Links ]
Balasubramanian, C. P., Pillai, S. M., & Ravichandran, P. (2005). Zero-water exchange shrimp farming systems (extensive) in the periphery of Chilka lagoon, Orissa, India. Aquaculture International, 12, 555-572. [ Links ]
Bartberger, C. A. & Pierce Jr., S.K. (1976). Relationship between ammonia excretion rates and hemolymph nitrogenous compounds of a euryhaline bivalve during low salinity acclimation. Biological Bulletin, 150, 1-14. [ Links ]
Bayne, B. L., Moore, M. N., Widdows, J., Livingstone, D. R., Salkeld, P.,Crisp, D. J., Morris, R. J., Gray, J. S., Holden, A. v., Newell, R. C., & McIntyre, A. D. (1979). Measurement of the Responses of Indivi- duals to Environmental Stress and Pollution, Studies with Bivalve Mollusks. Philosophical Transaction of Royal Society of London B, 286, 563-581. [ Links ]
Berg, D. J., Fisher, S. W., & Landrum, P. F. (1996). Clearance and processing of algal particles by Zebra Mussels (Dreissena polymorpha). Journal of Great Lakes Research, 22(3), 779-788. [ Links ]
Chavez-Crooker, P. & Oberque-Contreras, J. (2010). Biore-mediation of aquaculture wastes. Current Opinion in Biotechnology, 21, 313-317. [ Links ]
Chopin, T., Buschmann, A. H., Halling, Ch., Troell, M., Kautsky, N., Neori, A., Kraemer, G. P., Zertuche- González, J. A., Yarish, Ch., & Neefus, Ch. (2001). Integrating seaweeds into marine aquaculture systems, a key toward sustainability. Journal of Phycology, 37(6), 975-986. [ Links ]
Comoglio, L., Gaxiola, G., Roque, A., Cuzon, G., & Amin, O. (2002). The effect of starvation on re-feeding, digestive enzyme activity, oxygen consumption, and ammonia excretion in juvenile white shrimp Litopenaeus vannamei. Journal of Shellfish Research, 23, 243-249. [ Links ]
Conover, W. J. (1999). Practical nonparametric statistics. 3rd ed. New York, USA.: J Wiley & Sons. [ Links ]
De vooys, C. G. N. & De Zwaan, A. (1978). The rate of oxygen consumption and ammonia excretion by Mytilus edulis after various periods of exposure to air. Comparative Biochemistry and Physiology Part A Physiology, 60, 343-347. [ Links ]
Enriquez-Ocaña, L. F., Nieves-Soto, M., Piña-valdez, P., Martínez-Córdova, L. R., & Medina-Jasso, M. (2012). Evaluation of the combined effect of temperature and salinity on the filtration, clearance rate and assimilation efficiency of the mangrove oyster Crassostrea corteziensis (Hertlein, 1951). Archives of Biological Sciences, 64, 479-488. [ Links ]
FAO. (2012). El estado mundial de la pesca y la acuicultura 2012. Roma: FAO. [ Links ]
Fernández-Reiriz, M. J., Labarta, U., Albentosa, M., & Pérez-Camacho, A. (1998). Effect of Microalgal Diets and Commercial Wheat germ Flours on the Lipid Profile of Ruditapes decussatus Spat. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 119, 369-377. [ Links ]
Grant, J. & Thorpe, B. (1991). Effects of Suspended Sedi- ment on Growth, Respiration, and Excretion of the Soft-Shell Clam (Mya arenaria). Canadian Journal of Fisheries and Aquatic Sciences, 48, 1285-1292. [ Links ]
Haure, J., Penisson, C., Bougrier, S., & Baud, J. P. (1998). Influence of temperature on clearance and oxygen consumption rates of the flat oyster Ostrea edulis, determination of allometric coefficients. Aquaculture, 169, 211-224. [ Links ]
Jorgensen, C. B. (1990). Bivalve filter feeding, hydro- dynamics, bioenergetics, physiology and ecology. Fredensborg, Denmark: Olsen & Olsen. [ Links ]
Keen, M. (1971). Seashells of Tropical West America. Stanford, CA. USA.: Stanford University Press. [ Links ]
Martínez-Córdova, L. R. & Martinez-Porchas, M. (2006). Polyculture of the Pacific white shrimp, Litopenaeus vannamei, giant oyster, Crassostrea gigas, and blackclam, Chione fluctifraga in ponds in Sonora, Mexico. Aquaculture, 258, 321-326. [ Links ]
Martínez-Córdova, L. R. & Enríquez-Ocaña, L. F. (2007). Study of benthic fauna in the discharge lagoon of a shrimp farm with emphasis on the polychaetes. On Line Journal of Biological Sciences, 7(1), 12-17. [ Links ]
Martínez-Córdova, L. R., Martínez Porchas, M., & Cortés Jacinto, E. (2009). Camaronicultura Mexicana y Mundial, ¿Actividad sustentable o Industria Contaminante? Revista Internacional de Contaminación Ambiental, 25(3), 181-196. [ Links ]
Martínez-Córdova, L. R., López Elías, J. A., Martínez-Porchas, M., Bernal-Jaspeado, T., & Miranda Baeza, A. (2011). Studies on the bioremediation capacity of the adult black clam, Chione fluctifraga, of shrimp aquaculture effluents. Revista de Biología Marina y Oceanografía, 46, 105-113. [ Links ]
Martínez-Córdova, L. R., López-Elías, J. A., Leyva-Miranda, J. G., Armenta-Ayon, L., & Martinez-Porchas, M. (2011). Bioremediation and reuse of shrimp aquaculture effluents to farm whiteleg shrimp, Litopenaeus vannamei, a first approach. Aquaculture Research, 42, 1415-1423. [ Links ]
Martínez-Porchas, M. & Martínez-Córdova, L. R. (2012). World aquaculture, environmental impacts and troubleshooting alternatives. The Scientific World Journal, Recovered from http://www.hindawi.com/journals/ tswj/2012/389623/, o doi: 10.1100/2012/389623). [ Links ]
Miranda, A., Lizarraga-Armenta, J., Rivas-vega, M., López-Elías, J. A., & Nieves-Soto, M. (2010). Pacific Oyster, Crassostrea gigas, Cultured With Tilapia, Oreochromis mossambicus × Oreochromis niloticus in a Recirculation System. Journal of the World Aquaculture Society, 41, 764-772. [ Links ]
Nieves-Soto, M., Enríquez-Ocaña, F., Piña-valdez, P., Maeda-Martínez, A. N., Almodóvar-Cebreros, J. R., & Acosta-Salmón, H. (2011). Is the mangrove cockle Anadara tuberculosa a candidate for effluent bioremediation? Energy budgets under combined conditions of temperature and salinity. Aquaculture, 318, 434-438. [ Links ]
Nieves-Soto, M., Bückle-Ramírez, L. F., Piña-valdez, P., Medina-Jasso, M. A., Miranda-Baeza, A., Martínez- Córdova, L. R., & López-Elías, J. A. (2013). Combined effect of temperature and food concentration on the filtration and clarification rates and assimilation efficiency of Atrina tuberculosa Sowerby, 1835 (Mollusca, Bivalvia) under laboratory conditions. Archives of Biological Science, Belgrade, 65, 99-106. [ Links ]
Paniagua-Michel, J. & García, O. (2003). Ex-situ biore-mediation of shrimp culture effluent using constructed microbial mats. Aquacultural Engineering, 28, 131-139. [ Links ]
Peña-Messina, E., Martínez-Cordova, L. R., Bücke-Ramirez, L. F., Segovia-Quintero, M. A., & Zertuche- González, J. A. (2009). A preliminary evaluation of physiological filtration variables of Crassostrea corteziensis (Hertiein, 1951) and Anadara tuberculosa (Sowerby, 1833) in shrimp aquaculture effluents. Aquaculture Research, 40(15), 1750-1758. [ Links ]
Piedrahita, R. H. (2003). Reducing the potential environmental impact of tank aquaculture effluents through intensification and recirculation. Aquaculture, 226, 35-44. [ Links ]
Riisgård, H. U. (2001). On measurement of filtration rates in bivalves –the stony road to reliable data, review and interpretation. Marine Ecology Progress Series, 211, 275-291. [ Links ]
Rivera-Monroy, v. H., Torres, L. A., Bahamon, N., New- mark, F., & Twilley, R. R. (1999). The Potential Use of Mangrove Forests as Nitrogen Sinks of Shrimp Aquaculture Pond Effluents, the Role of Denitrification. Journal of the World Aquaculture Society, 30, 12-25. [ Links ]
Saucedo, P. E., Ocampo, L., Monteforte, M., & Bervera, H. (2004). Effect of temperature on oxygen consumption and ammonia excretion in the Calafia mother-of-pearl oyster, Pinctada mazatlanica (Hanley, 1856). Aquaculture, 229, 377-387. [ Links ]
Shumway, S. E. & Koehn, R. K. (1982). Oxygen Consumption in the American Oyster Crassostrea virginica. Marine Ecology Progress Series, 9, 59-68. [ Links ]
Widdows, J., Baynea, B. L., Donkina, P., Livingstonea, D. R., Lowea, D. M., Moorea, M. N., & Salkelda, P. N. (1981). Measurement of the responses of mussels to environmental stress and pollution in Sullom voe, a base-line study. Proceedings of the Royal Society of Edinburgh. Section B. Biological Sciences, 80, 323-338. [ Links ]
Zar, J. H. (1999). Biostatistical analysis. Englewood Cliffs, New Jersey, USA.: Prentice-Hall International. [ Links ]
Balasubramanian, C. P., Pillai, S. M., & Ravichandran, P. (2005). Zero-water exchange shrimp farming systems (extensive) in the periphery of Chilka lagoon, Orissa, India. Aquaculture International, 12, 555-572. [ Links ]
Bartberger, C. A. & Pierce Jr., S.K. (1976). Relationship between ammonia excretion rates and hemolymph nitrogenous compounds of a euryhaline bivalve during low salinity acclimation. Biological Bulletin, 150, 1-14. [ Links ]
Bayne, B. L., Moore, M. N., Widdows, J., Livingstone, D. R., Salkeld, P.,Crisp, D. J., Morris, R. J., Gray, J. S., Holden, A. v., Newell, R. C., & McIntyre, A. D. (1979). Measurement of the Responses of Indivi- duals to Environmental Stress and Pollution, Studies with Bivalve Mollusks. Philosophical Transaction of Royal Society of London B, 286, 563-581. [ Links ]
Berg, D. J., Fisher, S. W., & Landrum, P. F. (1996). Clearance and processing of algal particles by Zebra Mussels (Dreissena polymorpha). Journal of Great Lakes Research, 22(3), 779-788. [ Links ]
Chavez-Crooker, P. & Oberque-Contreras, J. (2010). Biore-mediation of aquaculture wastes. Current Opinion in Biotechnology, 21, 313-317. [ Links ]
Chopin, T., Buschmann, A. H., Halling, Ch., Troell, M., Kautsky, N., Neori, A., Kraemer, G. P., Zertuche- González, J. A., Yarish, Ch., & Neefus, Ch. (2001). Integrating seaweeds into marine aquaculture systems, a key toward sustainability. Journal of Phycology, 37(6), 975-986. [ Links ]
Comoglio, L., Gaxiola, G., Roque, A., Cuzon, G., & Amin, O. (2002). The effect of starvation on re-feeding, digestive enzyme activity, oxygen consumption, and ammonia excretion in juvenile white shrimp Litopenaeus vannamei. Journal of Shellfish Research, 23, 243-249. [ Links ]
Conover, W. J. (1999). Practical nonparametric statistics. 3rd ed. New York, USA.: J Wiley & Sons. [ Links ]
De vooys, C. G. N. & De Zwaan, A. (1978). The rate of oxygen consumption and ammonia excretion by Mytilus edulis after various periods of exposure to air. Comparative Biochemistry and Physiology Part A Physiology, 60, 343-347. [ Links ]
Enriquez-Ocaña, L. F., Nieves-Soto, M., Piña-valdez, P., Martínez-Córdova, L. R., & Medina-Jasso, M. (2012). Evaluation of the combined effect of temperature and salinity on the filtration, clearance rate and assimilation efficiency of the mangrove oyster Crassostrea corteziensis (Hertlein, 1951). Archives of Biological Sciences, 64, 479-488. [ Links ]
FAO. (2012). El estado mundial de la pesca y la acuicultura 2012. Roma: FAO. [ Links ]
Fernández-Reiriz, M. J., Labarta, U., Albentosa, M., & Pérez-Camacho, A. (1998). Effect of Microalgal Diets and Commercial Wheat germ Flours on the Lipid Profile of Ruditapes decussatus Spat. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 119, 369-377. [ Links ]
Grant, J. & Thorpe, B. (1991). Effects of Suspended Sedi- ment on Growth, Respiration, and Excretion of the Soft-Shell Clam (Mya arenaria). Canadian Journal of Fisheries and Aquatic Sciences, 48, 1285-1292. [ Links ]
Haure, J., Penisson, C., Bougrier, S., & Baud, J. P. (1998). Influence of temperature on clearance and oxygen consumption rates of the flat oyster Ostrea edulis, determination of allometric coefficients. Aquaculture, 169, 211-224. [ Links ]
Jorgensen, C. B. (1990). Bivalve filter feeding, hydro- dynamics, bioenergetics, physiology and ecology. Fredensborg, Denmark: Olsen & Olsen. [ Links ]
Keen, M. (1971). Seashells of Tropical West America. Stanford, CA. USA.: Stanford University Press. [ Links ]
Martínez-Córdova, L. R. & Martinez-Porchas, M. (2006). Polyculture of the Pacific white shrimp, Litopenaeus vannamei, giant oyster, Crassostrea gigas, and blackclam, Chione fluctifraga in ponds in Sonora, Mexico. Aquaculture, 258, 321-326. [ Links ]
Martínez-Córdova, L. R. & Enríquez-Ocaña, L. F. (2007). Study of benthic fauna in the discharge lagoon of a shrimp farm with emphasis on the polychaetes. On Line Journal of Biological Sciences, 7(1), 12-17. [ Links ]
Martínez-Córdova, L. R., Martínez Porchas, M., & Cortés Jacinto, E. (2009). Camaronicultura Mexicana y Mundial, ¿Actividad sustentable o Industria Contaminante? Revista Internacional de Contaminación Ambiental, 25(3), 181-196. [ Links ]
Martínez-Córdova, L. R., López Elías, J. A., Martínez-Porchas, M., Bernal-Jaspeado, T., & Miranda Baeza, A. (2011). Studies on the bioremediation capacity of the adult black clam, Chione fluctifraga, of shrimp aquaculture effluents. Revista de Biología Marina y Oceanografía, 46, 105-113. [ Links ]
Martínez-Córdova, L. R., López-Elías, J. A., Leyva-Miranda, J. G., Armenta-Ayon, L., & Martinez-Porchas, M. (2011). Bioremediation and reuse of shrimp aquaculture effluents to farm whiteleg shrimp, Litopenaeus vannamei, a first approach. Aquaculture Research, 42, 1415-1423. [ Links ]
Martínez-Porchas, M. & Martínez-Córdova, L. R. (2012). World aquaculture, environmental impacts and troubleshooting alternatives. The Scientific World Journal, Recovered from http://www.hindawi.com/journals/ tswj/2012/389623/, o doi: 10.1100/2012/389623). [ Links ]
Miranda, A., Lizarraga-Armenta, J., Rivas-vega, M., López-Elías, J. A., & Nieves-Soto, M. (2010). Pacific Oyster, Crassostrea gigas, Cultured With Tilapia, Oreochromis mossambicus × Oreochromis niloticus in a Recirculation System. Journal of the World Aquaculture Society, 41, 764-772. [ Links ]
Nieves-Soto, M., Enríquez-Ocaña, F., Piña-valdez, P., Maeda-Martínez, A. N., Almodóvar-Cebreros, J. R., & Acosta-Salmón, H. (2011). Is the mangrove cockle Anadara tuberculosa a candidate for effluent bioremediation? Energy budgets under combined conditions of temperature and salinity. Aquaculture, 318, 434-438. [ Links ]
Nieves-Soto, M., Bückle-Ramírez, L. F., Piña-valdez, P., Medina-Jasso, M. A., Miranda-Baeza, A., Martínez- Córdova, L. R., & López-Elías, J. A. (2013). Combined effect of temperature and food concentration on the filtration and clarification rates and assimilation efficiency of Atrina tuberculosa Sowerby, 1835 (Mollusca, Bivalvia) under laboratory conditions. Archives of Biological Science, Belgrade, 65, 99-106. [ Links ]
Paniagua-Michel, J. & García, O. (2003). Ex-situ biore-mediation of shrimp culture effluent using constructed microbial mats. Aquacultural Engineering, 28, 131-139. [ Links ]
Peña-Messina, E., Martínez-Cordova, L. R., Bücke-Ramirez, L. F., Segovia-Quintero, M. A., & Zertuche- González, J. A. (2009). A preliminary evaluation of physiological filtration variables of Crassostrea corteziensis (Hertiein, 1951) and Anadara tuberculosa (Sowerby, 1833) in shrimp aquaculture effluents. Aquaculture Research, 40(15), 1750-1758. [ Links ]
Piedrahita, R. H. (2003). Reducing the potential environmental impact of tank aquaculture effluents through intensification and recirculation. Aquaculture, 226, 35-44. [ Links ]
Riisgård, H. U. (2001). On measurement of filtration rates in bivalves –the stony road to reliable data, review and interpretation. Marine Ecology Progress Series, 211, 275-291. [ Links ]
Rivera-Monroy, v. H., Torres, L. A., Bahamon, N., New- mark, F., & Twilley, R. R. (1999). The Potential Use of Mangrove Forests as Nitrogen Sinks of Shrimp Aquaculture Pond Effluents, the Role of Denitrification. Journal of the World Aquaculture Society, 30, 12-25. [ Links ]
Saucedo, P. E., Ocampo, L., Monteforte, M., & Bervera, H. (2004). Effect of temperature on oxygen consumption and ammonia excretion in the Calafia mother-of-pearl oyster, Pinctada mazatlanica (Hanley, 1856). Aquaculture, 229, 377-387. [ Links ]
Shumway, S. E. & Koehn, R. K. (1982). Oxygen Consumption in the American Oyster Crassostrea virginica. Marine Ecology Progress Series, 9, 59-68. [ Links ]
Widdows, J., Baynea, B. L., Donkina, P., Livingstonea, D. R., Lowea, D. M., Moorea, M. N., & Salkelda, P. N. (1981). Measurement of the responses of mussels to environmental stress and pollution in Sullom voe, a base-line study. Proceedings of the Royal Society of Edinburgh. Section B. Biological Sciences, 80, 323-338. [ Links ]
Zar, J. H. (1999). Biostatistical analysis. Englewood Cliffs, New Jersey, USA.: Prentice-Hall International. [ Links ]
1. Programa de Postgrado en Biociencias de la Universidad de Sonora; karime_rc@hotmail.com
2. Departamento de Investigaciones Científicas y Tecnológicas de la Universidad de Sonora; Blvd Luis Donaldo Colosio s/n, entre Reforma y Sahuaripa, Edificio 7G, Hermosillo, Sonora, 83000, México; lmtz@guaymas.uson.mx, fenrquez@guayacan.uson.mx, jalopez@guayacan.uson.mx
3. Universidad Estatal de Sonora; Carr. a Huatabampo, Km. 5. Navojoa, Sonora, México, 85800; anselmo.miranda@ues.mx
Received 22-X-2013. Corrected 30-III-2014. Accepted 29-Iv-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}