










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.3 San José Jul./Sep. 2014
Species diversity and activity of insectivorous bats in three habitats
in La Virgen de Sarapiquí, Costa Rica
Amanda Cormier
Abstract: Pineapple farms make up 45 000ha of Costa Rican landscape and are the second most exported crop. This is economically beneficial for the Costa Ricans, but greatly affects the natural flora and fauna because it is such a low growing crop. This study examined the differences in insectivorous bat species diversity and activity in the habitat gradient between the forest in Tirimbina Biological Reserve in La Virgen de Sarapiquí, 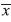 =22.6), in comparison to the border (=39.9), and the forest (=44.2) (p=0.0028). Agricultural environments affected and lowered bat presence. Also, a greater mean number of bats recorded between 1900-1930hrs compared to 1730-1800hrs, coincided with the setting of the sun and beginning of bat activity. More research is need throughout the night and the year to establish clearer patterns of bat use and activity in different habitats. Rev. Biol. Trop. 62 (3): 939-946. Epub 2014 September 01.
=22.6), in comparison to the border (=39.9), and the forest (=44.2) (p=0.0028). Agricultural environments affected and lowered bat presence. Also, a greater mean number of bats recorded between 1900-1930hrs compared to 1730-1800hrs, coincided with the setting of the sun and beginning of bat activity. More research is need throughout the night and the year to establish clearer patterns of bat use and activity in different habitats. Rev. Biol. Trop. 62 (3): 939-946. Epub 2014 September 01.
Key words: bat activity,
Resumen
Diversidad y actividad de especies de murciélagos insectívoros en tres hábitats de La Vírgen, de Sarapiquí, Costa Rica. Hay 45 000ha de piñeras en Costa Rica. La piña es la segunda fruta de mayor exportación en el país, por lo cual es muy importante para la economía. Este cultivo beneficia al país en términos económicos; no obstante, tiene efectos negativos en la flora y la fauna de áreas naturales al ser una planta herbácea de baja altura. Esta investigación examinó la diversidad y actividad de los murciélagos insectívoros en el gradiente entre un bosque (Reserva Biológica Tirimbina) y una piñera (Finca Corsicana) en la Virgen, Sarapiquí, Costa Rica. Durante cuatro semanas, puse grabadoras en tres sitios (bosque, borde de bosque y piñera) para grabar las llamadas de los murciélagos. Después analicé las llamadas para identificar las especies. Encontré cuatro familias y 19 especies. Hubo una reducción en el número de pases de murciélagos (número de veces que un murciélago vuela al lado de la grabadora) en la piñera ( =22.6), en comparación con el borde de bosque ( =39.9) y el bosque ( =44.2) (p=0.0028). El ambiente agrícola afectó y redujo la presencia de murciélagos. Familias diferentes parecen estar adaptadas a hábitats diferentes. Finalmente, un mayor número de murciélagos fueron registrados entre las 19:00 y 19:30 hrs, comparados con las 17:30-18:00 hrs, lo cual coincide con el atardecer y el inicio de la actividad nocturna de los murciélagos. Se necesita más investigación, tanto en horas nocturnas, como del resto del año, para entender mejor los patrones de actividad y el uso de diferentes hábitats.
Palabras clave: murciélagos insectívoros, actividad de murciélagos, grabadoras ultrasónicas, gradiente de hábitats, bosque tropical lluvioso, Costa Rica.
Conservation efforts are being made all over the world to help protect the environment from human destruction. In order for these conservation efforts to be implemented, there must be an understanding of: a) how changes to the landscape affect the flora and the fauna, and b) how to correct the problem or at least create a balance between the needs of humans and the natural life. This study focuses on how the gradation in diversity between a biological reserve (protected forest), the border of the forest, and a pineapple farm affect bat species diversity and activity.
Pineapple is the second most exported crop in
Miller, Arnett, and Lacki (2003) examined 56 bat studies on habitat management of forest-roosting bats and concluded that without roosting areas, bats will leave. Thus conservation efforts should be made to maintain roosting areas. It is highly beneficial to farmers to keep bats in an area because insectivorous bats greatly decrease pests and frugivorous and nectivorous bats help with pollination. It has been estimated that a colony of big brown bats (Eptesicus fuscus) can eat 1.3 million insects a year, or each bat can eat 4 to 8g per night. In monetary terms this is around $74/acre that farmers in south-central
Of 1 116 species of the bats in the world, 113 are found in Costa Rica and about 60 are known in the rural town of La Virgen de Sarapiquí, where this study was conducted (Laval & Rodríguez-Herrera, 2002). Around 75% of all bat species are insectivores, the rest eat fruit, nectar, fish, birds, or blood (Bat Conservation International, Inc., 2011). Even though insectivorous bats make up the majority of bat species, there are fewer studies on insectivorous bats in comparison to other types because insectivorous bats usually fly higher than mist nets, which are typically used in bat studies. The solution to this problem is recording devices.
Within the past 15 years there has been an increase in sonic recorder use to identify bat species difficult to catch in nets. Initially, research groups worked to identify all sounds made by each bat species by capturing them and recording calls of each species in open rooms. With species recordings, they were able to identify bats in the field with recording equipment (O’Farrell & Miller, 1999; Rydell, Arita, Santos, & Granados, 2002; Jung, Kalko, & von Helversen, 2007). Evidence of the usefulness of recording calls was shown in a study (Sampaio, Kalko, Bernard, Rodríguez-Herrera, & Handley, 2003) that focused on bat species adaptability to new surroundings in the
The use of recording devices is possible because the majority of bat species use echolocation to navigate and find food during the night (Rodríguez-Herrera & Montero, 2009). The bats send out the call and use the echo that bounces back to form an image of their surroundings, called an eco-image. This eco-image includes information about the form and the texture of an object. The frequency of these calls is higher than the human range, which stops at 20kHz. One bat can call as many as 400 000 times per night. Bats use two types of calls, the frequency-modulated (fm), which is in a wide frequency band for a short period of time, and the quasi-constant frequency (qcf), which is in a narrow frequency band for a longer period of time (Schnitzler & Kalko, 1998). Each species’ fm and qcf calls are different, which is how recording devices can be used to identify different bat species.
The main objectives of this study were to determine species richness in a gradient of disturbance in La Virgen de Sarapiquí. Through the use of sonic recorders, this study aimed to identify bat species in order to determine their activity and diversity within: a) the forest at Tirimbina Biological Reserve, b) the edge of this forest, and c) the edge of the pineapple farm, Finca Corsicana.
This will help build a better understanding of the interaction between bats and humans based on the effects of agriculture and create a better understanding of how best to protect bat habitats.
Materials and methods
Study area: This study took place in La Virgen de Sarapiquí, Heredia, in northcentral
Habitat sites: There were three main habitat sites: 1) the forest, 2) forest border, and the 3) pineapple farm. The forest was part of Tirimbina Biological Reserve, which is 98.55% forest, 1.15% open area; mostly comprised of walking trails, and 0.30% wetlands. The forested area is made up of 85% primary forest and 15% secondary forest and these include tropical pre-montane forest that transitions into basal and tropical forest. The west border of the reserve is made up of the
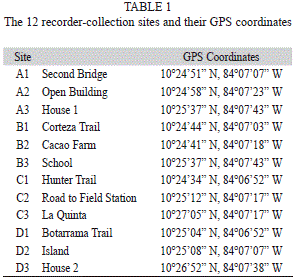
Recorder-collection sites: At each of the three habitat sites there were four recorder-collection sites (one used per week). During week A (March 11, 2013 to March 15, 2013) one recorder was placed on the tower of the second bridge within the forest (A1), the second within an open building at the edge of the forest (A2), and the third at house 1 along the edge of the pineapple farm (A3). During week B (March 18, 2013 to March 22, 2013) one recorder was placed just off the Corteza trail, which is past the bridge (B1), the second in the cacao plantation, which is along the river (B2), and the third on the side of a building part of Escuela Las Palmitas next to the pineapple farm (B3). During week C (April 8, 2013 to April 12, 2013) one recorder was placed on the Hunter trail (C1), the second was placed on the road to Tirimbina’s field station (C2), and the third at La Quinta hotel (C3). During week D (April 15, 2013 to April 19, 2013) the first recorder was placed within the forest off the Botarrama trail (D1), the second on an area known as the island, which divides the river in two (D2), and the third at house 2 near the pineapple farm (D3).
Bat call collection: Each week one recorder (Song Meter SM2BAT+, Wildlife Acoustics,
Bat call identification: During daylight hours I analyzed the recordings using Raven Pro 1.5, (The Cornell Lab of Ornithology,
To identify bat species, I compared the peak frequency, the length of the call, and the shape of the call to known calls (O’Farrell & Miller, 1997; Fenton, Rydell, Vonhof, Eklof, & Lancaster, 1999; Kossl, Mora, Coro, & Vater, 1999; O’Farrell & Miller, 1999; O’Farrell, Miller, & Gannon, 1999; Siemers, Kalko, & Schnitzler, 2001; Mora, Macías, Vater, Coro, & Kossl, 2004; Jung et al., 2007; MacSwiney et al., 2008; Pio, Clarke, MacKie, & Racey, 2010; Jung & Kalko, 2011). Peak frequency is the frequency at the maximum power of the call. These researchers have captured different species and recorded their calls in order to accurately identify the species. Many of the articles mentioned above contained pictures of the sounds that I compared to the images in Raven Pro 1.5.
Time constraints hindered the analysis of all 12h from each night. Instead I analyzed the calls only from 1730-1800hrs and 1900-1930hrs for each night.
Differences between means of the variable, “number of passes”, were estimated between: a) the three habitats and two time periods; b) bat families and two time periods; c) bat families and three habitats; and d) four weeks and three habitats, with parametric two-way ANOVA. Differences between means of the variable, “number of species”, were estimated between: a) three habitats and two time periods with parametric two-way ANOVA. Assumptions were tested prior to execution of parametric ANOVAS (Sokal & Rohlf, 1995). Tests were executed with Statgraphics statistical software (www.statgraphics.com).
Results
In 94 30-min segments over 4wks, several trends appeared. There were fewer bat passes for the pineapple farm ( =22.6) than for the forest ( =44.2) or border ( =39.9) (F=5.69, d.f.=2, 312, p=0.0028, Fig. 1). There was a significant increase in the mean number of bat passes between 1730-1800hrs ( =25.4) and 1900-1930hrs ( =44.3) (F=11.71, d.f.=1, 313, p=0.0001). This trend was consistent for all habitats (F=0.65, d.f.=2, 309, p=0.5241, Fig. 1).
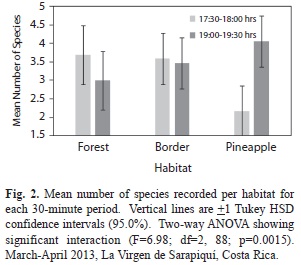
There was also a significant difference in the mean number of species per habitat and 30-min period (F=6.98, d.f.=2, 88, p=0.0015, Fig. 2). At 1730-1800hrs, there were more species in the forest ( =3.69) and border ( =3.59) than during 1900-1930hrs (forest =3.0, border =3.47). The opposite occurred at the pineapple site where there were fewer species during 1730-1800hrs ( =2.18) than during 1900-1930hrs ( =4.06).
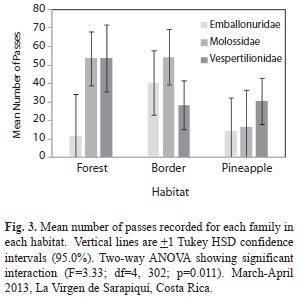
Four families of bats were found: Emballonuridae, Molossidae, Vespertillionidae, and only two passes of Moormopidae (removed for statistical testing). Of the four families, 19 different species were identified (Cormura brevirostris, Cynomops greenhalli, Cyttarops alecto, Diclidurus albus, Eptesicus brasiliensis, Eptesicus furinalis, Euderma maculatum , Eumops spp, Lasiurus ega, Molossus currentium, Molossus molossus, Myotis albescens, Myotis nigricans, Peropteryx kappleri, Peropteryx macrotis, Pteronotus parnellii, Rhogeessa io, Saccopteryx bilineata, Saccopteryx leptura). There was a significant difference in mean number of bat passes per family with Emballonuridae having the least ( =24.0), then Vespertillionidae ( =34.3), and Molossidae with the greatest mean number of passes ( =45.9) (F=4.32, d.f.=2, 308, p=0.0141). When families were compared to the three habitats there was a significant difference (F=3.33, d.f.=4, 302, p=0.0109). The mean number of bat passes for Emballonuridae was greatest at the border sites ( =40.5) and the smallest in the forest ( =11.5, Fig. 3). Mean number of passes of Molossidae was greatest at the border as well ( =54.4) and smallest in the pineapple farm ( =16.4, Fig. 3). In contrast, the greatest mean number of passes of Vespertillionidae was in the forest ( =53.7) and the least in the border habitat ( =28.3).
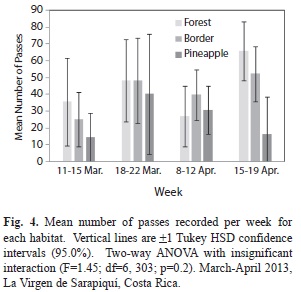
There was no significant difference in mean number of passes at each habitat between weeks (F=1.45, d.f.=6, 303, p=0.939, Fig. 4). For all four weeks the mean number of passes per habitat increased, decreased and, except for the pineapple farm, increased again in week D.
Between the two 30-min periods was an hour of time. This hour created a significant difference in the activity of the different families (F=6.61, d.f.=2, 305, p=0.0015, Fig. 5). There were fewer passes for the Emballonuridae ( =11.57) and the Molossidae ( =23.06) during 1730-1800hrs than between 1900-1930hrs (Emballonuridae =35.53, Molossidae =64.40). Mean number of passes of Vespertilionidae changed little and unlike the other two families, more passes occurred during 1730-1800hrs ( =36.14) than during1900-1930hrs ( =32.64).
Discussion
The main objective of this study was to examine the effect of habitat gradation on bats and results showed a difference in bat activity with the greatest mean number of passes in the forest and decreasing to the smallest mean number of passes in the pineapple farm. No significant difference existed in mean number of species between habitats, but a significant difference existed in mean number of passes per family in each habitat.
Based on different calls, different families seemed better suited to different habitats. For example, Vespertilionidae use fm calls with a long frequency and short duration, making them suited for forest habitats, where the greatest number of passes occurred (Schnitzler & Kalko, 1998). Voigt and Holderied (2012) found Molossidaes better suited for open spaces partly because of their physical conformation (body size and wing length) and their calls are qcf, almost cf, which means they have a small change in frequency and a longer length. In their study of M. currentium and M. sinaloe in open and closed spaces, the M. currentium performed better in the dense forest-like areas than the M. sinaloe. In my study there were a number of M. currentium and no M. sinaloe, which may explain why there was a greater number of Molossidae found in the forest and the border than the pineapple farm. Alpízar, Víquez, Hong, Rodríguez-Herrera, and González-Maya (2012) in Tirimbina, with bat species in dense and open spaces within the forest, found greater numbers of Emballonuridae in the forest than the open spaces, but overall there was more species richness in open spaces (Jaccard=0.308). I found more Emballonuridae in the border than the other two habitats. Emballonuridae use gcf calls, which have both a large change in frequency and a long length of time making them ideally suited for border spaces (Schnitzler & Kalko, 1998).
While there was no significant difference in number of passes per habitat between weeks, there were some interesting trends that can be explained by the individual recorder-collection sites. In week B, the mean number of passes in the forest and the border were almost the same (forest =48.07, border =48.14). During week B the forest recorder was placed low to the ground, which could have prevented the microphone from picking up all of the calls and the border site was in the cocoa plantation, which is along the =27.0). The forest recorder in week D was the furthest from the trail than in the other weeks creating a denser environment which may be why there is the highest mean number of passes at this site ( =65.60).
The last major trend was the increase of mean number of passes between 1900-1930hrs when compared to 1730-1800hrs (p=0.0001). Bats are nocturnal species and the sun sets around 1730hrs, so by 1900hrs there has been more time for the bats to wake up and head out to feed. Jung & Kalko (2010) noticed a decrease in bat activity after 2030hrs in their human populated site, but no change throughout the night in their forest site. More analysis would have to be done of the bat calls from my study in order to clearly notice a trend in bat activity throughout the night.
Acknowledgments
I would like to thank Bernal Rodríguez-Herrera for helping to guide me through this project with his vast knowledge of bats. Also, I would like to thank Christopher Vaughan and Michael McCoy for their help in editing and advising during the study. Judy Magnan arranged my home stay for 2mo and Alejandra Loaiza was instrumental in helping to collect all of my equipment. Additionally, Priscilla Alpízar helped in teaching me how to use the computer programs and how to identify the species based on the images. Finally, I would like to thank all of Tirimbina’s staff for their encouragement throughout the data collection process, especially Emmanuel Pequeño for helping me put out and take down the recorders every week.
References
Alpízar, P., Víquez, L. R., Hong, F., Rodríguez-Herrera, B., & González-Maya, J. F. (2012). Efecto de los claros de bosque en la composición de murciélagos insectívoros en la Reserva Biológica La Tirimbina, Sarapiquí, Costa Rica. Revista Biodiversidad Neotropical, 2(2), 138-142. [ Links ]
Bat Conservation International, Inc. (2013). Bat Conservation International, Inc. Retrieved from Bat Conservation International website. Retrieved from http://www.batcon.org. [ Links ]
Boyles, J. G., Cryan, P. M., McCracken, G. F., & Kunz, T. H. (2011). Economic importance of bats in agriculture. Science, 332, 41-42. [ Links ]
Estrada-Villegas, S., Meyer, C. F. J., & Kalko, E. K. V. (2010). Effects of tropical forest fragmentation on aerial insectivorous bats in a land-bridge island system. Biological Conservation, 143, 597-608. [ Links ]
Fenton, M. B., Rydell, J., Vonhof, M. J., Eklof, J., & Lancaster, W. C. (1999). Constant-frequency and frequency-modulated components in the echolocation calls of three species of small bats (Emballonuridae, Thyropteridae, and Vespertilionidae). Canadian Journal of Zoology, 77(12), 1891-1900. [ Links ]
Ingwersen, W. W. (2012). Life cycle assessment of fresh pineapple from
Jung, K. & Kalko, E. K. V. (2010). Where forest meets urbanization: Foraging plasticity of aerial insectivorous bats in an anthropogenically altered environment. Journal of Mammalogy, 91(1), 144-153. [ Links ]
Jung, K. & Kalko, E. K. V. (2011). Adaptability and vulnerability of high flying Neotropical aerial insectivorous bats to urbanization. Diversity and Distributions, 17(2), 262-274. [ Links ]
Jung, K., Kalko, E. K. V., & von Helversen, O. (2007). Echolocation calls in
Kossl, M., Mora, E.,
Laval, R. K. & Rodríguez-Herrera, B. (2002). Murciélagos de Costa Rica. San José, Costa Rica: Editorial INBio. [ Links ]
MacSwiney, M. C., Clarke, F. M., & Racey, P. A. (2008). What you see is not what you get: The role of ultrasonic detectors in increasing inventory completeness in Neotropical bat assemblages. Journal of Applied Ecology, 45, 1364-1371. [ Links ]
Miller, D. A., Arnett, E. B., & Lacki, M. J. (2003). Habitat management for forest-roosting bats of
Mora, E. C., Macías, S., Vater, M.,
O’Farrell, M. J. & Miller, B. W. (1997). A new examination of calls of some Neotropical bats (Emballonuridae and Mormoopidae). Journal of Mammalogy, 78(3), 954-963. [ Links ]
O’Farrell, M. J. & Miller, B. W. (1999). Use of vocal signatures for the inventory of free-flying Neotropical bats. Biotropica, 31(3), 507-516. [ Links ]
O’Farrell, M. J., Miller, B. W., & Gannon, W. L. (1999). Qualitative identification of free-flying bats using the Anabat detector. Journal of Mammalogy, 80(1), 11-23. [ Links ]
Pio, D. V. V., Clarke, F. M., MacKie,
Rodríguez-Herrera, B. & Montero, K. (2009). El ultrasonido de los murciélagos: Historia y retos. Boletín Oficial del Proyecto de Conservación de Aguas y Tierras ProCAT, 2(2), 19-24. [ Links ]
Rydell, J., Arita, H. T.,
Sampaio, E. M., Kalko, E. K. V., Bernard, E., Rodríguez-Herrera, B., & Handley, C. O., Jr. (2003). A biodiversity assessment of bats (Chiroptera) in a tropical lowland rainforest of central
Schnitzler, H. U. & Kalko, E. K. V. (1998). How echolocating bats search and find food. In T. H. Kunz & P. A. Racey, (Eds.), Bat biology and conservation (pp. 183-196).
Siemers, B. M, Kalko, E. K. V., & Schnitzler, H. U. (2001). Echolocation behavior and signal plasticity in the Neotropical bat Myotis nigricans (Schinz, 1821) (Vespertilionidae): A convergent case with European species of Pipistrellus? Behavioral Ecology and Sociobiology, 50, 317-328. [ Links ]
Sokal, R. R. & Rohlf, F. J. (1995). Biometry: The principles and practice of statistics in biological research.
Tirimbina Biological Reserve. (2010). Retrieved from Tirimbina website: http://www .tirimbina.org). [ Links ]
Voigt, C. C. & Holderied, M. W. (2012). High maneuvering costs force narrow-winged molossid bats to forage in open space. Journal of Comparative Physiology, 182, 415-424. [ Links ]
Received 20-III-2014. Corrected 20-IV-2014. Accepted 26-V-2014.


{kind=link}
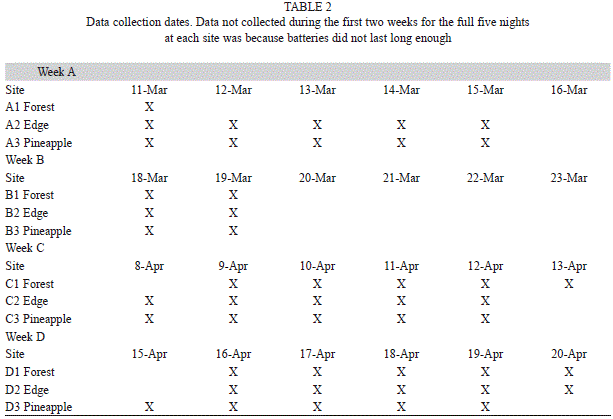{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}