










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.3 San José Jul./Sep. 2014
Beach almond (Terminalia catappa, Combretaceae) seed production and predation by scarlet macaws (Ara macao) and variegated squirrels (Sciurus variegatoides)
Producción de semillas y depredación del almendro de playa (Terminalia catappa, Combretaceae) por lapas rojas (Ara macao) y ardillas chizas (Sciurus variegatoides)
Producción de semillas y depredación del almendro de playa (Terminalia catappa, Combretaceae) por lapas rojas (Ara macao) y ardillas chizas (Sciurus variegatoides)
Abstract
Knowledge of ecological impacts of exotic beach almond (Terminalia catappa) in the central Pacific of Costa Rica are little known, but studies have found this species to be a potentially important food source for endangered scarlet macaws (Ara macao). In this study, reproductive phenology and seed predation by variegated squirrels (Sciurus variegatoides) and scarlet macaws were measured during March and April 2011 on beaches of central Pacific coastal Costa Rica. Seed productivity and predation levels were quantified on a weekly basis for 111 beach almond trees to assess the importance of beach almond as a food source for scarlet macaws and the extent of resource partitioning between seed predators. Seed production of the trees was great (about 194 272 seeds) and approximately 67% of seeds were predated by seed predators. Macaws consumed an estimated 49% of seeds while squirrels consumed 18%. Additionally, evidence of resource partitioning between squirrels and macaws was found. Scarlet macaws preferred to feed on the northern side and edge of the canopy while squirrels preferred to feed on the southern and inside parts of the canopy. Both species ate most seeds on the ocean side of the tree. Despite the status of this tree as an exotic species, the beach almond appears to be an important resource for scarlet macaw population recovery. The resource produced by this tree should be taken into account as reforestation efforts continue in Costa Rica. Rev. Biol. Trop. 62 (3): 929-938. Epub 2014 September 01.
Key words: Ara macao, beach almond, Costa Rica, phenology, scarlet macaw, seed predation, Sciurus variegatoides, Terminalia catappa, variegated squirrel.
Resumen
El conocimiento de los impactos ecológicos del almendro de playa exótico (Terminalia catappa) en el Pacífico Central de Costa Rica son poco conocidos, pero los estudios han encontrado que esta especie es una fuente de alimento potencialmente importante para la lapa roja (Ara macao), en peligro de extinción. En este estudio, se midieron la fenología reproductiva y la depredación de semillas por las ardillas (Sciurus variegatoides) y lapas rojas durante marzo y abril 2011 en las playas de la costa Pacífica Central de Costa Rica. Los niveles de productividad y depredación de semillas se cuantificaron semanalmente para 111 almendros de playa, para evaluar la importancia del almendro de playa como fuente de alimento para la lapa roja y el grado de repartición de recursos entre los depredadores de semillas. La producción de semillas de los árboles fue alta (cerca de 194 272 semillas) y aproximadamente el 67% de las semillas fueron comidas por los depredadores, las lapas rojas consumen un estimado de 49% de las semillas, mientras que las ardillas consumen el 18%. Adicionalmente, se encontró evidencia de la repartición de recursos entre las ardillas y las lapas. Las lapas rojas prefieren alimentarse en el lado norte y el borde de la copa, mientras que las ardillas prefieren las regiones del sur y el interior de la copa. Ambas especies se comieron la mayoría de las semillas en la parte del árbol con lado al mar. A pesar de la situación de este árbol como una especie exótica, la playa de almendras parece ser un recurso importante para la recuperación de la población de lapas rojas. El recurso que produce este árbol debe tomarse en cuenta para continuar con los esfuerzos de reforestación en Costa Rica.
Palabras clave: Ara macao, almendro de playa, Costa Rica, fenología, lapa roja, depredación de semillas, Sciurus variegatoides, Terminalia catappa, ardilla chiza.
The role of, and response to, exotic species has recently become a hotly debated topic (Davis et al., 2011). While exotic species are often viewed as detrimental to native communities, it is important to consider what the actual effects of an exotic species are in this age of rapid environmental change. In many cases introduced species do indeed result in negative environmental changes (Vitousek, D’Antonio, Loope, & Westbrooks, 1996), but in some cases, there is evidence that an introduced species can improve conditions for some native species. Changes generated by introduced species such as pest control, pollination, competitive and predator release, and trophic subsidies can result in benefits to native species, especially in environments where climate and land use is changing rapidly, resulting in modified native communities (Rodriguez, 2006; Schlaepfer, Sax, & Olden, 2010).
In this case, the scarlet macaw (Ara macao L.) in Costa Rica, suffered severe population declines in the 1980s and early 90´s due primarily to deforestation and poaching (Wiedenfeld, 1994; Vaughan, Nemeth, Cary, & Temple, 2005). This population decline resulted in the listing of scarlet macaws as threatened on the Appendix I of the Convention on International Trade in Endangered Species (CITES). However, over the last 20 years, habitat restoration, local education, and poaching control have resulted in considerable growth of the Costa Rican scarlet macaw population (Vaughan et al., 2005). As the macaw population has been recovering, much about their behavior and ecology has been investigated, including diet (Vaughan, Nemeth, & Marineros, 2006). Most notably, recent macaw diet studies have indicated a large reliance on exotic species, with 52% of their diets consisting of exotic species while one specific exotic tree, Terminalia catappa (L.) makes up about 36% of the diet (Matuzak, Bezy, & Brightsmith, 2008).
Tropical forest, especially dry forest in Central America has been has been severely reduced primarily by deforestation for cattle grazing (Quesada & Stoner, 2003). When grazing occurs, forests have less regeneration of many native trees that macaws feed on (Stern, Quesada, & Stoner, 2002) and as this reduction in native food sources has occurred, T. catappa has been widely planted in reforestation efforts, especially along beaches (Vaughan et al., 2006).
Terminalia catappa is native to Southeast Asia but, due to ocean currents and humans, it has spread to almost all tropical areas. While T. Catappa is common throughout Costa Rica, high tolerance to wind and salt stress makes it most prevalent on beaches. On beaches, T. catappa is known to prevent erosion of beach sand, provide food and shelter for humans and animals, and has potential for commercial cultivation for its seeds (Thomson & Evans, 2006). Based on observations, the seed production peak of T. catappa coincides with that of many other dry forest species, occurring during the dry season (January-May) (Frankie, Baker, & Opler, 1974).
Scarlet macaws, as seed predators, are also important in the reproduction of T. catappa. In addition to macaws, variegated squirrels (Sciurus variegatoides Ogilby, 1839), ants, and various bats are known to feed on T. catappa seeds (Zuchowski, 2007). Of these other seed predators, squirrels are the most common (personal observation). In utilizing the same resource as scarlet macaws, it would be expected that these predators would interact in some way, either competing for or partitioning the resource (MacArthur, 1958).
The objective of this study was to investigate the role and the effects of an exotic species as a food source for threatened scarlet macaws in Costa Rica and how interaction with variegated squirrels affects macaw use of the resource. Specific objectives were to investigate 1) production and predation levels of T. catappa seeds during the dry season and 2) seed predator feeding patterns and interactions on T. catappa trees.
Materials and Methods
Study site: This study was performed on the central Pacific coast of Costa Rica between the Tárcoles River and the resort Punta Leona (9°45’ N - 84°40’ W). The study area is characterized by the transition between dry and humid tropical forest (Tosi, 1969), but is now dominated by agriculture and small forest patches. The climate is warm and humid, with a mean annual temperature of 25-30°C and mean annual precipitation of 2.5-3.3m and distinct wet (May-December) and dry (January-April) seasons (Coen, 1983).
Seven beaches were studied; Playas Azul, Tárcoles, Pita, Agujas, Limoncito, Manta, and Blanca (Fig. 1). Playas Azul, Tárcoles, and Pita are primarily residential, while playa Agujas has a section developed for tourism while most is unoccupied by humans. Playas Limoncito, Manta, and Blanca are part of the resort Punta Leona and playas Manta and Blanca are frequented by resort guests, while Limoncito is small and isolated, with little human presence.
Tree selection and characteristics: Judgment sampling was used to get a wide array of trees of different sizes, locations, and surrounding characteristics. Number of trees selected per beach varied by beach length and was as follows; Azul (n=14), Tárcoles (n=20), Pita (n=20), Agujas (n=24), Limoncito (n=5), Manta (n=16), and Blanca (n=12). Both the total tree height and the height of the leaved canopy (total height - height of trunk without leaves) were measured with a clinometer. Additionally, a tape measure was used to measure the crown diameter (distance from edge to edge of the crown). A combination of crown height and diameter was used to calculate crown volume.
Seed production data: Seed production was monitored on a weekly basis on all trees studied. To quantify the reproductive phenology, the number of mature seeds, immature seeds, and flower heads were counted in three randomly selected sections of approximately 1m3 per tree. A mature seed was defined as a seed the size where predators were likely to consume it, determined by observations of seeds already eaten. Immature seeds were seeds smaller than the seeds frequently consumed by predators. Flower heads were defined as any flowering head containing flowers with petals.
Seed predation data: Seed predation was measured on a weekly basis in 0.5m2 plots. Eight plots per tree were organized with four plots on the interior, approximately 0.5m from the center of the crown and four plots around the edge of the crown. Plots were oriented North, South, East, and West, with two plots in each direction (Fig. 1). Plots 1, 3, 5, and 7 were considered interior plots while plots 2, 4, 6, and 8 were considered exterior plots. Plots 1 and 2 were considered North plots, plots 3 and 4 were considered East plots, plots 5 and 6 were considered South plots, and plots 7 and 8 were considered West plots. Preliminary trials indicated that there was negligible post-dispersal predation of entire seeds or empty shells, and that the exocarp of the seeds remained green for approximately one week. Thus, only seeds with some green exocarp left on their outer shell were considered during weekly sampling.
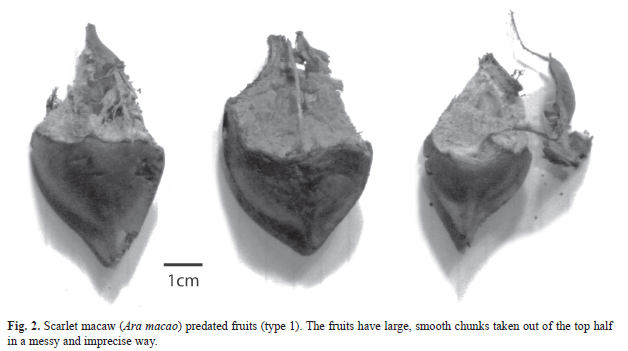
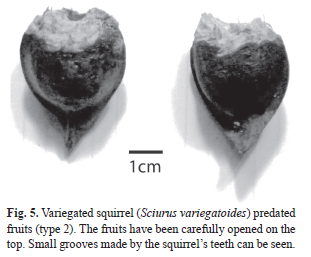
Seeds from each plot were categorized by 5 seed predation classes based on shell markings. These classes included; macaw predation, macaw rejection, squirrel predation, squirrel rejection, and no predation. Predated seeds were defined as opened seeds where the embryo had been eaten (Fig. 2, Fig. 3, Fig. 4 and Fig. 5). Rejected seeds were defined as seeds showing evidence of being picked by a predator, but not opened to expose the inner part (Fig. 6, Fig. 7). No predation seeds were defined as those that showed no sign of predation. Macaws and squirrels each had two methods of opening seeds (Fig. 2, Fig. 3, Fig. 4 and Fig. 5).
Total seed production and predation were calculated for each tree based on the seed, flower, seed predation, and seed rejection density obtained during sampling, combined with the total volume of the crown and area under the crown for each tree. For analytical purposes, the no predation category consisted of seeds that were rejected by predators and seeds that showed no signs of predation.
Contingency tables and Chi-squared tests (Sokal & Rohlf, 1995) were used to analyze trends in preferred feeding location of seed predators along with the association between seed predators. Observed predation frequency, split by plot under each tree and by beach, was compared to the expected frequency. When predation occurred, differences in mean number of seeds predated between plots and beaches for both macaws and squirrels were analyzed by one-way parametric ANOVAs. For these ANOVAs, the data were log-transformed and a posteriori comparisons between means were analyzed using the Tukey method. For interpretability, means are reported without transformation.
Results
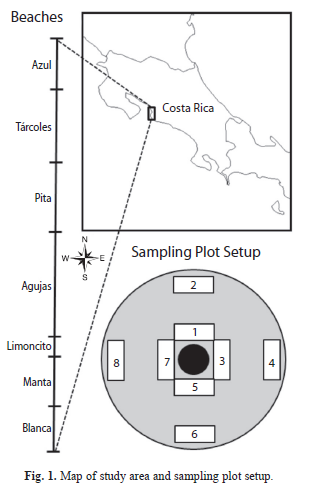
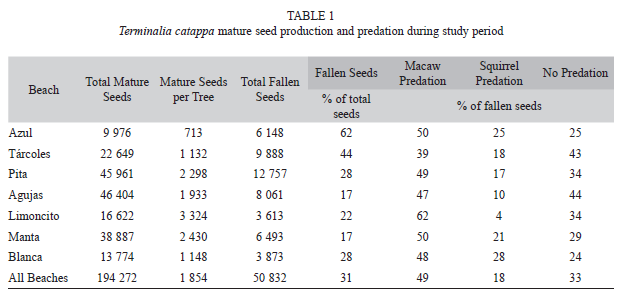
Seed production and predation: Across 111 trees, approximately 194 272 mature seeds were produced during the 2-mo study period (
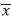 =1 854 seeds/tree). A total 24 294 seeds were predated by scarlet macaws, 9 857 seeds rejected by macaws, 8 814 seeds predated by squirrels, 2 448 seeds rejected by squirrels, and 5 419 seeds that fell without signs of predation during the study period. This makes for a total of 50 832 seeds that fell from trees during the study period, 31% of the total mature seeds produced. Of those that fell, 33% had not been consumed by predators. The proportion of seeds that fell varied by beach, with Northern beaches (Azul and Tárcoles) having a higher proportion of seeds produced that fell compared to other beaches. However, Azul differed from Tárcoles in that a higher proportion of fallen seeds were predated (Table 1).
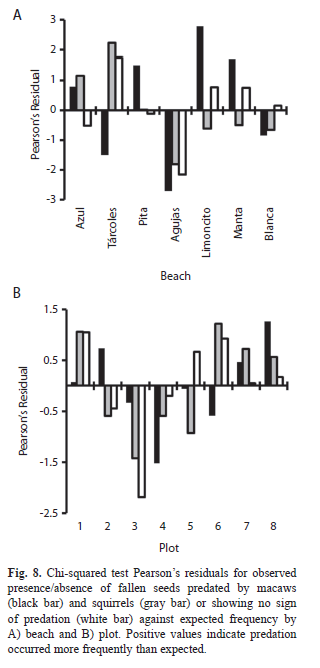
=1 854 seeds/tree). A total 24 294 seeds were predated by scarlet macaws, 9 857 seeds rejected by macaws, 8 814 seeds predated by squirrels, 2 448 seeds rejected by squirrels, and 5 419 seeds that fell without signs of predation during the study period. This makes for a total of 50 832 seeds that fell from trees during the study period, 31% of the total mature seeds produced. Of those that fell, 33% had not been consumed by predators. The proportion of seeds that fell varied by beach, with Northern beaches (Azul and Tárcoles) having a higher proportion of seeds produced that fell compared to other beaches. However, Azul differed from Tárcoles in that a higher proportion of fallen seeds were predated (Table 1). Predator preference and interaction, beach: Both the frequency and amount of predation varied significantly by beach for macaws and squirrels. Macaw predation was more frequent than expected on Azul, Pita, Limoncito, and Manta and less frequent than expected on Tárcoles and Agujas (X2=48.7, df=6, p<0.001) (Fig. 8A). Squirrels, on the other hand, preferred to feed on Azul and Tárcoles beaches while feeding less frequently than expected on the more Southern beaches (X2=15.8, df=6, p=0.01) (Fig. 2A). Notably, macaws and squirrels differed in feeding preferences on Tárcoles (squirrel preferred), Pita, Limoncito, and Manta (macaw preferred) beaches and were similar on Azul, Agujas, and Blanca. Seeds that were not predated were found more often than expected on Tárcoles, Limoncito, and Manta and less frequently on Agujas (X2=21.9, df=6, p=0.001) (Fig. 8A).
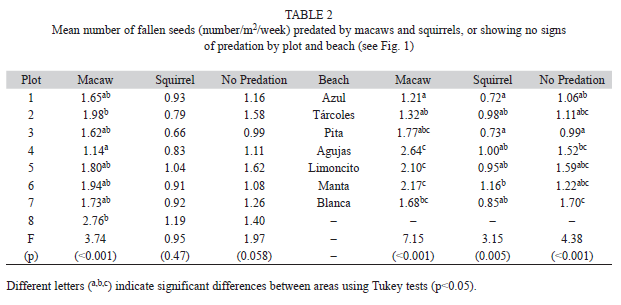
When predation did occur, the amount varied by beach for each seed predator. For macaws, significantly more seeds were consumed on Southern beaches (Agujas, Limoncito, Manta, and Blanca) while fewer seeds were consumed on the Northern beaches (Azul, Tárcoles, and Pita) (Table 2). Squirrel predation was consistently less than macaw predation and significantly more predation occurred on Manta compared to Azul and Pita beaches. Significantly more intact seeds fell on Agujas and Blanca compared to Azul and Pita beaches (Table 2).
Tárcoles was the only beach where macaws didn’t consume approximately 50% of the predated seeds. Squirrels, on the other hand, consumed a very low proportion of predated seeds on Agujas and Limoncito compared to other beaches (Table 1).
Predator preference and interaction, plot: There were fewer significant differences in predation frequency by plot. Observed macaw predation frequency was not significantly different than expected (X2=10.5, df=7, p=0.16), although there was a tendency to feed more often above plots 2, 7, and 8, and feed less frequently above plots 3, 4, and 6. Squirrel predation frequency also did not vary significantly from expected (X2=10.5, df=7, p=0.16) but there was a tendency to feed more frequently in plots 1 and 6-8 and less frequently than expected in plots 2-5. The observed frequency of intact seeds did differ significantly from expected (X2=21.9, df=6, p=0.001), with the most notable difference being a much lower frequency observed in plot 3 (Fig. 8B).
When macaws did feed, the amount of seeds eaten above each plot differed significantly, with higher amount of predation in plots 2 and 8 compared to plot 4. While not significantly different, there was also a tendency to eat more seeds in plot 6. Overall, macaws tended to consume more seeds around the edge of the canopy on the ocean side. Squirrel predation did not differ significantly by plot although there were slightly more seeds consumed above plots 1, 5, and 8. Plot was marginally significant in in number of seeds that fell without predation (Table 2).
Discussion
Regarding scarlet macaw predation on beach almond, the high proportion of T. catappa seeds in scarlet macaw diets (Matuzak et al., 2008) resulted in the consumption of almost half of the seeds produced by T. catappa. This is very similar to a study by Villaseñor-Sanchez, Dirzo, & Renton (2010) that found a 43% seed predation rate by Amazona finschi on Astronium graveolens and a study by Coates-Estrada, Estrada, & Meritt (1993) that found a mean predation rate of 51% by Amazona autumnalis on Stemmadenia donnell-smithii. These examples are, however, much greater than other studies of seed predation by parrots. A study of scarlet macaw predation of Brazil nut (Bertholletia excelsa) revealed that macaws consumed only 10% of the seed crop (Trivedi, Cornejo, & Watkinson, 2004). The predation rate reported in this study, however, only includes seeds that had fallen and there were many seeds left to fall in most trees at the end of the study.
Macaws preferred to feed on three beaches; Blanca, Manta, and Limoncito. All three of these beaches are located within a resort/club called Punta Leona, but are the furthest beaches from the macaw roosting site in the Guacalillo Mangrove Reserve (Vaughan et al., 2005) to the north of the study area. This would not be expect based on optimal foraging theory (MacArthur & Pianka, 1966), but could be a result of the conservation efforts made by Punta Leona. The resort has planted many T. catappa trees on its grounds and throughout the whole central Pacific coast of Costa Rica. Additionally, the beaches of Punta Leona are part of the ecological blue flag program in Costa Rica, which is only awarded to the healthiest beaches of Costa Rica.
Squirrels, on the other hand, had preferences for the most human-dominated beaches. Azul, Tárcoles, and Pita beaches are all residential, with houses adjacent to the beach. Squirrels are often associated with humans, so they are better adapted to the disturbances common on developed beaches. Interestingly, the number of macaw feeding events was not necessarily lower on residential beaches, but the number of seeds eaten was. This indicates that macaws spend less time feeding on residential beaches, even though they visit them about the same amount as other beaches. This was probably observed because macaws were more likely to be disturbed by human presence.
Regarding preferred feeding locations, both seed predators fed more often over the ocean side (plot 8) and less often away from the ocean (plot 4). This is likely because the main method of natural propagation of T. catappa is through the ocean currents and tides (Thomson & Evans, 2006), making it advantageous to produce a large number of seeds on the ocean side. This larger concentration of seeds would draw more seed predators. Plot 4, on the other hand, faces away from the ocean and is often engulfed by other trees, decreasing the light that reaches that side.
The preferred feeding locations in trees provide some evidence of resource partitioning between seed predators in this study. There was a weak division in predation between the north and south sides of the tree canopy, where macaws tended to feed on the northern side while squirrels preferred to feed on the southern side of the tree. This indicates that macaws may prefer to feed more often in the morning as they fly from their roosting site to the north of the study area. There was also a slight tendency for squirrels to prefer feeding on the inside of the tree canopy, while macaws tended to feed on the edge of the canopy. This resource partitioning is likely a result of morphological differences between species. Macaws fly from tree to tree landing on outer branches. Alternatively, squirrels tend to climb the trunk to access the crown, making it easier to consume seeds along the interior of the tree.
This study suggests that T. catappa seeds are a very important food resource for scarlet macaws during the study period (March-April, 2011). As macaw populations have been increasing in the central Pacific of Costa Rica (Vaughan et al., 2005), T. catappa has become an important part of their diet (Matuzak et al., 2008). Additionally, this study reveals high T. catappa seed predation by macaws along beaches. As a tree that grows quickly and produces many seeds, T. catappa planting projects, which are occurring throughout the Central Pacific of Costa Rica, appear to be important in macaw recovery. Additionally, T. catappa seeds are important for other seed predators, and current predation rates suggest that enough food is produced to support both squirrels and macaws.
This system is an example of how exotic species can interact with and influence native populations. In this case, this exotic species is likely integral in supporting an endangered species, and facilitating their recovery, while possibly beneficial to beach communities (Thomson & Evans, 2006). This does not, however, mean that the overall effects of T. catappa on the surrounding community should not be investigated to assess whether the species is harming other elements of the community.
The importance of Punta Leona resort as an example to other beaches, is shown by its success and, thus should be a model for other beaches. Despite being very populated, Punta Leona had the greatest macaw presence, which indicates that beach health and cleanliness is likely more important than human presence. Not only should reforestation of native trees along with T. catappa take place, but measures should be taken to clean up beaches and reduce pollution in the area.
During March and April of 2011, the following conclusions were reached: a) T. catappa trees on the central Pacific coast of Costa Rica produced a large number of seeds where natural food sources may have been scarce; b) there was evidence of resource partitioning of the seed resource between squirrels and macaws; and c) T. catappa trees were a very important component of scarlet macaw diets and the resource size allowed squirrels and macaws to persist on the same resource.
Acknowledgments
Thanks goes to the Associated Colleges of the Midwest, Costa Rica program staff for coordinating this study abroad program. Kathleen Shea provided important feedback on this project. Also, thanks to my (JJH) host families during my stay in Costa Rica. Finally, thanks to Punta Leona resort, especially Arnulfo Villalobos Arias for his assistance in the field. I would also like to thank The Parrot Society UK for funding different aspects of the project.
References
Coates-Estrada, R., Estrada, A., & Meritt, D. (1993). Foraging by parrots (Amazona autumnalis) on fruits of Stemmadenia donnell-smithii (Apocynaceae) in the tropical rainforest of Los Tuxtlas, Mexico. Journal of Tropical Ecology, 9, 121-124. [ Links ]
Coen, E. (1983). Climate. In D. Janzen (Ed.). Costa Rican Natural History (pp. 35-46). Chicago, IL, USA: University of Chicago. [ Links ]
Davis, M., Chew, M. K., Hobbs, R. J., Lugo, A. E., Ewel, J. J., Vermeij, G. J., & Briggs, J. C. (2011). Don’t judge species on their origins. Nature, 474(1), 53-154. [ Links ]
Frankie, G. W., Baker, H. G., & Opler, P. A. (1974). Comparative phenological studies of trees in tropical wet and dry forests in lowlands of Costa Rica. Journal of Ecology, 62, 881-919. [ Links ]
MacArthur, R. H. (1958). Population ecology of some warblers of northeastern coniferous forests. Ecology, 39, 599-619. [ Links ]
MacArthur, R. H. & Pianka, E. R. (1966). On the optimal use of a patchy environment. American Naturalist, 100, 603-609. [ Links ]
Matuzak, G. D., Bezy, M. B., & Brightsmith, D. J. (2008). Foraging ecology of parrots in a modified landscape: Seasonal trends and introduced species. The Wilson Journal of Ornithology, 120, 353-365. [ Links ]
Quesada, M. & Stoner, K. E. (2003). Threats to the conservation of tropical dry forest in Costa Rica. In G. W. Frankie, A. Mata, & S. B. Vinso, (Eds.). Biodiversity Conservation in Costa Rica: Learning the lessons in the seasonal dry forest (pp. 266-280). Berkeley, CA, USA: University of California. [ Links ]
Rodriguez, L. F. (2006). Can invasive species facilitate native species? Evidence of how, when, and why these impacts occur. Biological Invasions, 8, 927-939. [ Links ]
Schlaepfer, M. A., Sax, D. F., & Olden, J. D. (2010). The potential conservation value of non-native species. Conservation Biology, 25, 428-437. [ Links ]
Sokal, R. R. & Rohlf, F. J. (1995). Biometry: The principles and practice of statistics in biological research. New York, USA: W. H. Freeman and Co. [ Links ]
Stern, M., Quesada, M., & Stoner, K. E. (2002). Changes in composition and structure of a tropical dry forest following intermittent cattle grazing. Revista de Biologia Tropical, 50(3/4), 1021-1034. [ Links ]
Thomson, L. A. J. & Evans, B. (2006). Terminalia catappa (tropical almond), ver. 2.2. Species Profiles for Pacific Island Agroforestry. Permanent Agriculture Resources (PAR). Hōlualoa, HI, USA. Retrieved from http://agroforestry.net/tti/T.catappa-tropical-almond.pdf [ Links ]
Tosi, J. (1969). Ecological map of Costa Rica. San José, Costa Rica: Tropical Science Center. [ Links ]
Trivedi, M. R., Cornejo, F. H., & Watkinson, A. R. (2004). Seed predation on Brazil nuts (Bertholletia excelsa) by macaws (Psittacidae) in Madre de Dios, Peru. Biotropica, 36, 118-122. [ Links ]
Vaughan, C., Nemeth, N. M., Cary, J., & Temple, S. (2005). Response of a scarlet macaw (Ara macao) population to conservation practices in Costa Rica. Bird Conservation International, 15, 119-130. [ Links ]
Vaughan, C., Nemeth. N., & Marineros. L. (2006). Scarlet Macaw, (Ara macao), diet in Central Pacific Costa Rica. Revista de Biología Tropical, 54, 919-926. [ Links ]
Villaseñor-Sanchez, E., Dirzo, I. R., & Renton, K. (2010). Importance of the lilac-crowned parrot in pre-dispersal seed predation of Astronium graveolens in a Mexican tropical dry forest. Journal of Tropical Ecology, 26, 227-236. [ Links ]
Vitousek, P. M., D’Antonio, C. M., Loope, L. L., & Westbrooks, R. (1996). Biological invasions as global environmental change. American Scientist, 84, 468-478. [ Links ]
Wiedenfeld, D. (1994). A new subspecies of scarlet macaw (Ara macao) and its status and conservation. Ornitologia Neotropical, 5, 99-104. [ Links ]
Zuchowski, W. (2007). Tropical plants of Costa Rica. Ithaca, New York, USA: Cornell University. [ Links ]
Coen, E. (1983). Climate. In D. Janzen (Ed.). Costa Rican Natural History (pp. 35-46). Chicago, IL, USA: University of Chicago. [ Links ]
Davis, M., Chew, M. K., Hobbs, R. J., Lugo, A. E., Ewel, J. J., Vermeij, G. J., & Briggs, J. C. (2011). Don’t judge species on their origins. Nature, 474(1), 53-154. [ Links ]
Frankie, G. W., Baker, H. G., & Opler, P. A. (1974). Comparative phenological studies of trees in tropical wet and dry forests in lowlands of Costa Rica. Journal of Ecology, 62, 881-919. [ Links ]
MacArthur, R. H. (1958). Population ecology of some warblers of northeastern coniferous forests. Ecology, 39, 599-619. [ Links ]
MacArthur, R. H. & Pianka, E. R. (1966). On the optimal use of a patchy environment. American Naturalist, 100, 603-609. [ Links ]
Matuzak, G. D., Bezy, M. B., & Brightsmith, D. J. (2008). Foraging ecology of parrots in a modified landscape: Seasonal trends and introduced species. The Wilson Journal of Ornithology, 120, 353-365. [ Links ]
Quesada, M. & Stoner, K. E. (2003). Threats to the conservation of tropical dry forest in Costa Rica. In G. W. Frankie, A. Mata, & S. B. Vinso, (Eds.). Biodiversity Conservation in Costa Rica: Learning the lessons in the seasonal dry forest (pp. 266-280). Berkeley, CA, USA: University of California. [ Links ]
Rodriguez, L. F. (2006). Can invasive species facilitate native species? Evidence of how, when, and why these impacts occur. Biological Invasions, 8, 927-939. [ Links ]
Schlaepfer, M. A., Sax, D. F., & Olden, J. D. (2010). The potential conservation value of non-native species. Conservation Biology, 25, 428-437. [ Links ]
Sokal, R. R. & Rohlf, F. J. (1995). Biometry: The principles and practice of statistics in biological research. New York, USA: W. H. Freeman and Co. [ Links ]
Stern, M., Quesada, M., & Stoner, K. E. (2002). Changes in composition and structure of a tropical dry forest following intermittent cattle grazing. Revista de Biologia Tropical, 50(3/4), 1021-1034. [ Links ]
Thomson, L. A. J. & Evans, B. (2006). Terminalia catappa (tropical almond), ver. 2.2. Species Profiles for Pacific Island Agroforestry. Permanent Agriculture Resources (PAR). Hōlualoa, HI, USA. Retrieved from http://agroforestry.net/tti/T.catappa-tropical-almond.pdf [ Links ]
Tosi, J. (1969). Ecological map of Costa Rica. San José, Costa Rica: Tropical Science Center. [ Links ]
Trivedi, M. R., Cornejo, F. H., & Watkinson, A. R. (2004). Seed predation on Brazil nuts (Bertholletia excelsa) by macaws (Psittacidae) in Madre de Dios, Peru. Biotropica, 36, 118-122. [ Links ]
Vaughan, C., Nemeth, N. M., Cary, J., & Temple, S. (2005). Response of a scarlet macaw (Ara macao) population to conservation practices in Costa Rica. Bird Conservation International, 15, 119-130. [ Links ]
Vaughan, C., Nemeth. N., & Marineros. L. (2006). Scarlet Macaw, (Ara macao), diet in Central Pacific Costa Rica. Revista de Biología Tropical, 54, 919-926. [ Links ]
Villaseñor-Sanchez, E., Dirzo, I. R., & Renton, K. (2010). Importance of the lilac-crowned parrot in pre-dispersal seed predation of Astronium graveolens in a Mexican tropical dry forest. Journal of Tropical Ecology, 26, 227-236. [ Links ]
Vitousek, P. M., D’Antonio, C. M., Loope, L. L., & Westbrooks, R. (1996). Biological invasions as global environmental change. American Scientist, 84, 468-478. [ Links ]
Wiedenfeld, D. (1994). A new subspecies of scarlet macaw (Ara macao) and its status and conservation. Ornitologia Neotropical, 5, 99-104. [ Links ]
Zuchowski, W. (2007). Tropical plants of Costa Rica. Ithaca, New York, USA: Cornell University. [ Links ]
1. Biology Department, St. Olaf College, 1500 St. Olaf Ave, Northfield, MN, USA; henn.jonathan@gmail.com
2. Associated Colleges of the Midwest, Costa Rica Program, San Jose, Costa Rica; mmccoycolton@yahoo.com
3. Associated Colleges of the Midwest, Costa Rica Program, San Jose, Costa Rica; cvaughan@acm.edu
Received 20-iii-2014. Corrected 24-iv-2014. Accepted 26-v-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}