










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.1 San José Jan./Mar. 2014
Reproductive success and genetic diversity of Psychotria hastisepala (Rubiaceae), in fragmented Atlantic forest, Southeastearn Brazil
*Dirección para correspondencia:
Abstract
The impacts of forest fragmentation on both reproductive biology and genetic diversity of native plant species is hardly understood, despite some studies have analyzed this current worldwide problem. Since this constitutes one of the main threats to seasonal semi-deciduous forests in Southeastern Brazil, we investigated the reproductive success and the genetic diversity of a distylous, understory shrub (Psychotria hastisepala) within this context of forest fragmentation. For this study, a set of seven forest fragments of sizes ranging from 4.1 to 168.7 hectares were chosen. The intervenient matrix comprised pastures (25-50%), monocultures (33-50%) and rural roads and buildings (14-28.5%). Overall, 91 plants (54 for the short-styled morph and 37 for the long-styled morph; mean of 6.5 plants per fragment) were investigated. To evaluate reproductive success, we quantified fruit and seed production under natural pollination; to evaluate genetic diversity and population structure, we employed ISSR markers on genomic DNA. Plants with the short-styled morph exhibited a significantly higher reproductive success than those with the long-styled morph; there was no association between seed production and size of the forest fragment. Levels of genetic diversity were positively associated with the number of plants per fragment; but they were not related to flower morph. AMOVA showed that about 65% of the overall genetic variation was attributed to the differences between plants within fragments. The results suggested that populations of P. hastisepala were susceptible to decline owing to forest fragmentation. Rev. Biol. Trop. 62 (1): 309-319. Epub 2014 March 01.
Key words: conservation, distyly, habitat fragmentation, ISSR, Semideciduous Seasonal Forest.
Resumen
Los impactos de la fragmentación de los bosques tanto en la biología reproductiva como en la diversidad genética de las especies de plantas nativas apenas se entienden, a pesar de que algunos estudios han analizado este problema mundial. Dado que esto constituye una de las principales amenazas para los bosques semi-deciduos estacionales en el sureste de Brasil, se investigó el éxito reproductivo y la diversidad genética de un arbusto de sotobosque, Psychotria hastisepala, en un bosque fragmentado. Para este estudio, se seleccionaron siete fragmentos de tamaños que van desde 4.1 hasta 168.7 hectáreas. La matriz intervenida comprende pastos (25-50%), monocultivos (33-50%) y carreteras y edificios rurales (14-28.5%). En total, 91 plantas fueron investigadas (54 morfos florales brevistilo y 37 longistilo, con una media de 6.5 plantas por fragmento). Para evaluar el éxito reproductivo, se cuantificó la producción de frutos y semillas de plantas con polinización natural; para evaluar la diversidad genética y la estructura poblacional, se emplearon marcadores ISSR en el ADN genómico. Las plantas con morfo brevistilo exhibieron un éxito reproductivo significativamente más alto que las longistilo, no hubo asociación entre la producción de semillas y el tamaño del fragmento de bosque. Los niveles de diversidad genética se asociaron positivamente con el número de plantas por fragmento, pero que no estaban relacionados con los morfos florales. AMOVA mostró que alrededor del 65% de la variación genética en general se atribuyó a las diferencias entre plantas dentro de fragmentos. Los resultados sugieren que las poblaciones de P. hastisepala fueron susceptibles a disminuir debido a la fragmentación de los bosques.
Palabras clave: conservación, distilia, fragmentación del hábitat, ISSR, floresta estacional semideciduo.
The Atlantic Forest is one of the most threatened rainforests in Brazil. Currently, only about 8% of the forest’s original area remain intact, which represent little more than 1% of the entire Brazilian territory (Fundação S.O.S. Mata Atlântica, 2004). Impacts of forest fragmentation on both reproductive biology and genetic diversity of native plant species remains poorly understood, although some studies were undertaken (Tabarelli, Silva & Gascon, 2004; Goulart, Ribeiro & Louvato, 2005; Ramos & Santos, 2005; Lopes & Buzato, 2007).
Forest fragmentation leads to habitat loss, which in turn may lead to plant extinction (Taki, Kevan & Ascher, 2007). Moreover, forest management practices and land conversión to agriculture are factors that contribute significantly to extinctions at local scale given that they reduce both habitat quality (De Sanctis, Afo, Francesconi & Bruno, 2010) and population size of native species (Aizen & Feinsinger, 1994). When compared to continuous forests, isolated forest fragments harbor low levels of genetic diversity owing mostly to limited gene flow and genetic drift (Frankham, Ballou & Bruno, 2002). Longstanding isolation may increase endogamy (Ellstrand & Elam, 1993), reduce reproductive success (Steffan-Dewenter & Tscharntke, 1999; Cunningham, 2000; Wolf & Harrison, 2001; Donaldson, Nänni & Zachariades, 2002) and interrupt the gene flow (González-Astorga & Nuñez-Farfán, 2001).
Distylous plant species have two types of floral morphs, which are placed reciprocally between stigma and anthers on distinct individuals (Barrett, 1992): the long-styled morph (hereafter L morph) and the short-styled morph (hereafter S morph). Positioning anthers and stigmas at similar locations within a flower increases the precision of pollen transfer between morphs since both sexual organs contact similar parts of the pollinator (Ganders, 1979).
Distylous species exhibit heteromorphic incompatibility, therefore fruiting occurs exclusively when legitimate pollination (S x L and L x S) takes place (Barrett, 1992). Moreover, distylous species depend upon pollinators to accomplish legitimate pollination. Reduced pollination services, may appear if the distances among fragmented populations are greater than the foraging range of pollinators; if local pollinator populations declines significantly; or if long-distance pollinators avoid small plant populations (Kearns, Inouye & Waser, 1998). As a result of the deleterious effects of forest fragmentation, distylous species may undergo a reduction in reproductive success (Byers, 1995; Brys, Jacquemyn, Endels, Hermy & Blust, 2003; Ramos & Santos, 2005; Lopes & Buzato, 2007) and an overall decrease in genetic diversity (Rossi, Oliveira, Venturini & Silva, 2009). Conservation measures for distylous species will be more effective when genetic diversity within and among remnant populations are understood within the context of forest fragmentation.
In this study, we investigated the reproductive success and the genetic diversity of Psychotria hastisepala Müll. Arg., a plant species with a distylous system (Pereira, Vieira & Carvalho-Okano, 2006). This study addressed the following questions: a) Do floral morphs of P. hastisepala (S or L) differ on reproductive success? b) Is there any association between reproductive success rates and size of forest fragments? c) How is genetic diversity partitioned within and among forest fragments? d) Is reproductive success linked to genetic diversity? Our study took into consideration the context of recent human-led fragmentation which presently dominates the Atlantic Forest of Southeastern Brazil. Addressing these questions should shed light on causes that render populations of a distylous species, such as P. hastisepala, more susceptible to local extinction when exposed to a scenario of forest fragmentation. In addition, our study contributes to the elaboration of strategies to protect a threatened ecosystem that strongly requires conservation measures.
Materials and methods
Study species: Psychotria hastisepala is a distylous understory shrub; its distribution is restricted to Southeastern Brazil (Anderson, 1992). P. hastisepala exhibits both sexual and asexual reproduction; it form clumps of up to seven ramets (C.A. Silva, personal communication, March 18, 2008). The species is selfincompatible, both at plant and at morph levels (Pereira et al., 2006); therefore, reproduction takes place exclusively by intermorph pollination. Insects are the most likely pollinators (C.A. Silva, personal communication, March 18, 2008). The flowers are white, tubular, last for 24h and are reciprocally herkogamous; the stigma, lobes, and corolla are significantly larger in S than in L morphs (Pereira et al., 2006). Flowers are grouped in terminal inflorescences protected by paleaceous bracts that remain in the inflorescence until fruiting. The species produces drupaceous fruits, which exhibit a blue color when ripe and are bird-dispersed (sensu van der Pijl, 1982). A voucher specimen was deposited in the herbarium VIC (No. 26.964).
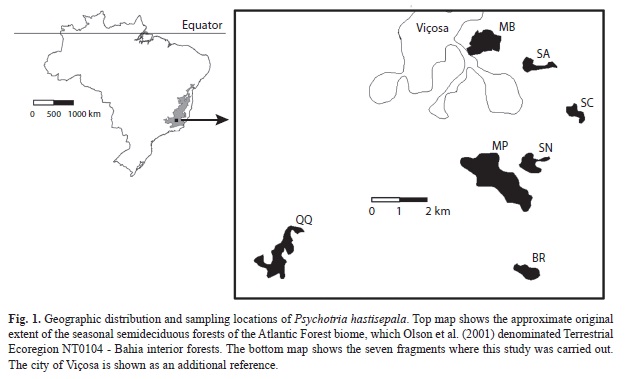
Study sites: Originally, continuous forests covered the study area (Rizzini, 1992). Nevertheless, the present-day vegetation is formed predominantly by patches of pastures of molasses grass (Melinis minutiflora Beauv.) intermingled with fragments of secondary forests at different succession stages. Primary vegetation is rare in this part of the country (less than 1%); most of the forest fragments are between 20 and 90 years old (Ribon, Simon & Mattos, 2003). Overall, the mean altitude is 690masl mean annual rainfall is 1 221mm, and the mean annual temperature ranges from 19 to 22ºC; the climate is strongly seasonal, with mild, rainy summers and dry winters (Marangon, Soares & Feliciano, 2003). Preliminary field surveys were carried out in 18 forest fragments and indicated that P. hastisepala occurred only in seven of them: Bárbara Rubim (BR) (20º49’53” S - 42º50’59” W), Sítio Cristais (SC) (20º46’47” S - 42º50’01” W), Sítio Agostinho (SA) (20º45’50” S - 42º50’18” W), Sítio Nico (SN) (20º47’37” S - 42º50’42” W), Mata Biologia (MB) (20º45’23” S - 42º52’19” W), Sítio Quim-Quim (QQ) (20º49’51” S - 42º55’50” W) e Mata do Paraíso (MP) (20º48’07”S - 42º51’29” W). For each of the seven forest fragment, we inspected visually the adjacent areas and recorded the surrounding vegetation cover (matrix); we also estimated the size of the fragment with the help of a global positioning system (GPS). The impact of anthropogenic changes was assessed using presence/absence field data, which recorded whether the surrounding areas were used for agricultural purpose, pastures or civil constructions (buildings and roads). Because P. hastisepala is associated with shady, forested areas, we evaluated habitat quality based on the presence or absence of understory favorable to the occurrence of the species.
Reproductive success: Overall, the number of plants of P. hastisepala per fragment was estimated to be less than 50. While choosing plants for this study, we applied a mínimum distance of 3m between specimens to minimize the likelihood of sampling clones.
To estimate reproductive success, we evaluated fruit and seed production under natural pollination. For each fragment, the number of study plants varied from one up to 10 per morph (mean=6.5). In total, we sampled 91 plants (54 for S morph and 37 for L morph). We choose randomly one to five inflorescences per plant (mean=4.7; total=425); fruits and sedes of each inflorescence were counted; the mean seed production per morph and the mean seed production per fragment was calculated.
DNA extraction and ISSR amplification: Total genomic DNA was extracted in accordance with the procedure described in Rossi et al. (2009) from leaf material (approximately 100mg) sampled from each of the 91 plants (54 for S morph and 37 for L morph) used for the studies of reproductive success. Genomic DNA from all plants was archived in our laboratory at the Federal University of Viçosa, Brazil.
For ISSR amplification, a total of 20 ISSR primers (UBC primer set #9, Biotechnology Laboratory, University of British Columbia, Canada) were evaluated using two randomly chosen plants of each morph per forest fragment. Ten primers produced strong bands and were evaluated further for ISSR polymorphism. Thereafter, amplification conditions, annealing temperatures and primer concentrations were optimized using the procedures described in Rossi et al. (2009).
Final PCR amplifications were carried out in a total volume of 20mL, consisting of 20ng DNA template, 2.0mL 10x PCR buffer (Phoneutria), 2.5mMMgCl2, 0.2mM of each dNTP, 2% formamide, 0.2mM primer, 0.75U Taq DNA polymerase (Phoneutria) and ultrapure water. PCR amplification was programmed on a GeneAmp PCR System 9700 thermal cycler (Applied Biosystems) under the following conditions: denaturation at 94°C for five minutes, 35 cycles of 45s denaturation at 94ºC, 45s annealing at 45 or 52ºC (according to the primer), 90s extension at 72ºC, followed by an additional seven min extension at 72ºC. Amplification products were separated via electrophoresis on 1.5% (w/v) agarose gels with 1x TBE buffer at constant voltage (110V) for approximately four hours. Negative controls, in which the DNA template was intentionally omitted, were also amplified. A typical gel carried two sets of 12 samples and each set was flanked on both sides by a 100bp DNA Ladder (Invitrogen). The gel was stained with ethidium bromide (0.6ng/mL) for about 30min and then photographed under UV light using a gel documentation system (Eagle Eye II, Stratagene). To ensure consistency during band size measurement, the images were grouped according to the ISSR primer and compared side-by-side on a computer screen.
Statistical analyses for reproductive success were run using general linear modeling one way ANOVA with Statistica (Stat Soft, 2002).
ISSR bands were treated as dominant genetic markers and scored as 1 (present) or 0 (absent) binary characters. Only polymorphic bands that could be unambiguously scored across all the surveyed plants were considered for further analyses. The resulting data was analyzed using POPGENE 1.31 (Yeh, Yang, & Boyle, 1999) to estimate the following three parameters of genetic diversity: P, percentage of polymorphism (P, %), Shannon diversity index (I) and Nei’s genetic diversity index HE, (Nei, 1973). Population differentiation was estimated by analysis of molecular variance (AMOVA, Excoffier, Smouse & Quattro, 1992) using Arlequin 3.01 (Excoffier, Laval, & Schneider, 2006). AMOVA examined how genetic diversity was partitioned within and among fragments at morph level. The significance of the genetic differentiation was tested with 1 000 permutations, where P denotes the probability of having a more extreme variance component than the observed values by chance alone. The Mantel test (Mantel, 1967) with 500 permutations as implemented in Genes (Cruz, 1997) was used to test the correlation between the total number of seed and the size of the fragment. Principal coordinate analysis (PCoA) was carried out at the plant level based on Nei and Li’s (1979) coefficient using NTSYSpc 2.2 (Rohlf, 2005).
Results
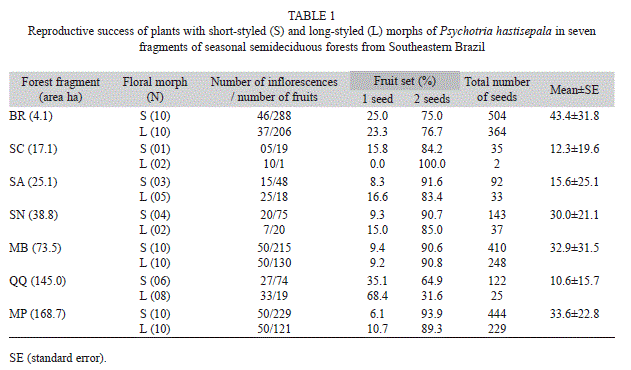
Study sites: The seven study fragments exhibited different shapes, with sizes ranging from 4.1 to 168.7 hectares (Table 1, Fig. 1). Matrices surrounding the fragments were mainly of agricultural origin: pastures (25- 50%), monocultures, such as eucalyptus, coffee or maize (33-50%), and rural constructions, such as roads and buildings (14-28.5%).
Reproductive success: Plants of the S morph produced significantly more fruits and more seeds (p<0.05) than plants of the L morph. In both morphs, there were significantly more fruits bearing two seeds than one seed (p<0.05); with fragment QQ being the only exception. In QQ, L-morph plants produced 68.4% of fruits bearing only one seed (Table 1). The Mantel test (r=-0.2057, p=0.6389) showed no correlation between the total number of seeds and the size of the fragment. For example, plants of the smallest forest fragment (BR, with only 4.1 hectares) produced about 43 seeds on average, while plants of the largest fragment (QQ, with 145 hectares) produced only about 11 seeds on average (Table 1).
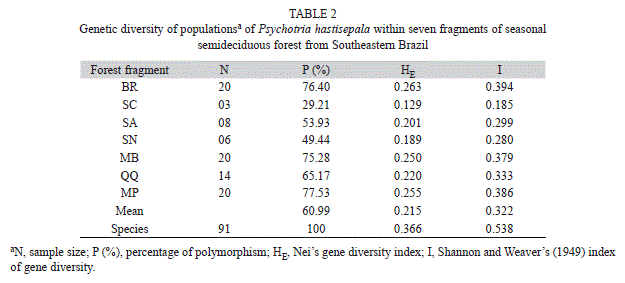
ISSR polymorphism and genetic diversity: We scored a total of 89 reproducible bands; most of which (98.88%) showed polymorphism. Among fragments, the percentage of polymorphic bands ranged from 29.21 to 77.53% (Table 2). Fragments with larger number of plants exhibited higher levels of genetic diversity, as estimated by Nei’s gene diversity index (HE) and Shannon and Weaver’s (1949) index (I) (Table 2); they also presented higher rates of reproductive success (Table 1). At morph level, Nei’s genetic diversity index (0.33 and 0.31 for B and L, respectively) and on Shannon’s index of diversity (0.49 and 0.46 for B and L, respectively) exhibited similar values.
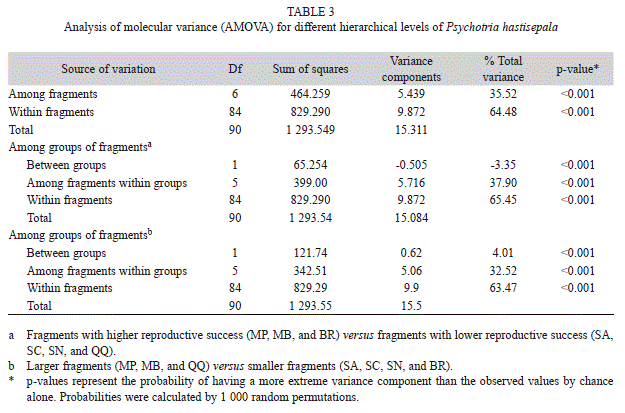
AMOVA showed that 64.5% of the total genetic diversity in plants was due to differences within fragments and 35.5% was explained by differences between fragments (Table 3). When fragments were separated into two groups of reproductive success (higher and lower), 37.9% of the total genetic diversity was caused by differences between fragments in each group, while 65.5% was due to differences among plants. The above results were confirmed when the fragments were grouped according to size: large fragments (MP, MB, and QQ) against small fragments (SA, SC, SN, and BR). The highest percentage of genetic variability (63.47%) was explained by variations among plants, independent of fragment size (Table 3).
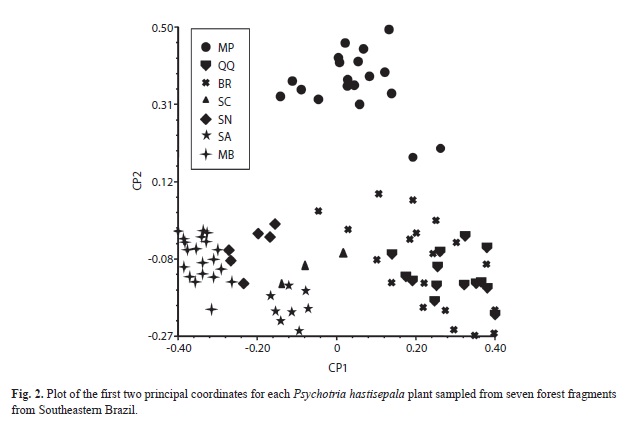
The principal coordinate analysis (Fig. 2) based on ISSR data grouped the 91 plants into two clusters, with no clear relationship between geographic distance among fragments and cluster composition. The first cluster contained plants from fragment MP exclusively. The second cluster brought together plants from fragments MB, SN, SA, QQ, BR, and SC; this cluster spread along the first coordinate (CP1). In the second cluster, there was a tendency of finding MB at one edge of the CP1, while BR and QQ occupied the opposite edge of the CP1. Pairwise Fst comparison indicated that fragments BR and QQ formed the pair that displayed the lowest level of genetic differentiation (0.1707). Nevertheless, BR differed from QQ in fragment size (4.1 x 145ha, respectively), number of study plants (20 x 14, respectively), and number of seeds produced (868 and 1547, respectively) (Table 1).
Discussion
The loss of suitable habitats accounts for the extinction of many plant species in the Neotropics (Fahrig, 2003; De Sanctis et al., 2010). Our data showed that there were instances in which small fragments harbored large populations of P. hastisepala, whereas large fragments contained a small number of plants. Moreover, the species was not present in 11 forest fragments (out of 18 we initially surveyed). Additional work is necessary to investigate further what ecological and genetic forces caused P. hastisepala to be absent from many fragments; population decline due to poor habitat quality cannot be rule out at the moment.
Our results clearly showed that reproductive success in P. hastisepala did not depend on the size of the forest fragment. Although BR was the smallest forest fragment, it displayed an understory that provided P. hastisepala with habitat quality such that the plants on this fragment produced the highest seed set per plant amongst the fragments we analyzed. In fragment BR, unlike other neighboring forest fragments, forest-disturbing activities that cause genetic erosion (Oliveira & Martins, 2002), such as cattle movement, cattle grazing, logging, and understory clearing, were not allowed.
The quality of the cover matrix and proximity among fragments may foster a higher rate of reproductive success in P. hastisepala. SN, for example, was a fragment that contained only six plants, which produced each over 30 seeds per plant. The fragment is located at a distance of about 250m from MP, the largest fragment in the study area. The short distance between fragments may allow pollinators to fly in and out of SN from MP, and therefore accomplish efficient pollinator services within SN. In other circumstances, where the distances are large, the cover matrix may work as a selective barrier and hamper the dispersal of plant pollinators, as they may not be able to cross it at a frequency efficient enough to accomplish pollination (Pires, Fernandez & Barros, 2006).
The higher reproductive success rate of the S morph over L morph of P. hastisepala may be related to the morphological dimorphisms found in distylous species (Pereira et al., 2006; Koch, Silva, & Silva, 2010). In the L morph, stigma exposure may favor geitonogamy and the low fruit set we uncovered may result from stigma clogging due to the deposition of incompatible pollen grains. In the distylous Turnera ulmifolia L. (Turneraceae), for example, occlusion of stigmas may result in lower fruit set in the L morph as compared to the S morph (Shore & Barrett, 1984).
Because P. hastisepala is a cross-pollinated species, its reproductive success depends on the availability of pollinators. Some studies have found a relationship between reproductive success and population size (Aizen & Feinsinger, 1994; Krauss, Hermanutz, Hopper, & Coates, 2007), suggesting that seed production may be lower in small fragments (Spears, 1987; Jennersten, 1988; Aguilar & Galeto, 2004) as fragments of small sizes may be less attractive to pollinating insects (Ohara, Tomimatsu, Takada, & Kawano, 2006). On the other hand, other studies suggested that floral resources for pollinators may not necessarily decrease linearly with the fragment size, because small fragments may be able to maintain or attract several pollinating insect species (Donaldson et al., 2002). Our results suggested that pollen deposition was effective for P. hastisepala even within fragments that harbor a relatively small number of plants.
Most of the genetic diversity in P. hastisepala was found within fragments. Similar findings were also reported for other tropical, cross-pollinating species such as Mabea fistulifera (Goulart et al., 2005) and Psychotria ipecacuanha (Rossi et al., 2009) and species with vegetative reproduction, such as Psammochloa villosa (Li & Ge, 2001). In general, high genetic diversity values within populations are expected in xenogamous species (Nybom & Bartish, 2000), while in autogamous species such as Coffea arabica (Rubiaceae) values of total diversity are relatively high and genetic diversity within populations low (Aga, Bekele, & Bryngelsson, 2005). High genetic diversity was reported for endemic species (Kang, Chang, & Kim, 2000; Gomes, Collevatti, Silveira, & Fernandes, 2004) whose genetic variability is maintained by their own particular reproductive mechanisms or strategies. Therefore, the greater genetic divergence within P. hastisepala populations may be explained by its self-incompatibility and vegetative reproduction, due to the maintenance of genetic variability by the clones in the latter case.
The absence of clear relationships between geographic and genetic distances between the forest fragments may also be due to other environmental factors. According to Schaal, Hayworth, Olsen, Rauscher, and Smith (1998), ecological factors, such as influence the genetic structure and inter-populational gene exchange. Apart from the ecological, historical factors may also explain the low genetic diversity observed in the forest fragments. In the past, the fragments may have been linked to each another over a long time, forming a continuous vegetation mass. Possibly the fragmentation was too recent to allow higher genetic divergence values.
In summary, our results showed that the reproductive success of P. hastisepala does not depend on the fragment size. Local environmental characteristics play a decisive role in the species’ maintenance. Lower genetic diversity values were observed in fragments with small populations of the species and with more marked habitat loss. All fragments analyzed in the present study are potentially vulnerable to a fast population decline, as a consequence of forest fragmentation.
Acknowledgments
Celice A. Silva received an DSc fellowship from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). Research fellowships from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) were granted to Milene F. Vieira and Luiz O. Oliveira (PQ 304153/2012-5). Financial support was provided by the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG). We are thankful to the administration staff of the Mata do Paraíso for allowing the field work.
References
Aga, E., Bekele, E., & Bryngelsson, E. (2005). Inter-simple sequence repeat (ISSR) variation in forest coffee trees (Coffea arabica L.) populations from Ethiopia. Genetica, 124, 213-221. http://link.springer.com/ [ Links ]
Aguilar, R., & Galetto, L. (2004). Effects of forest fragmentation on male and female reproductive success in Cestrum parqui (Solanaceae). Oecologia, 138, 513-520. http://link.springer.com/journal/442 [ Links ]
Aizen, M. A., & Feinsinger, P. (1994). Forest fragmentation, pollination, and plant reproduction in a Chaco dry forest, Argentina. Ecology, 75, 330-351. http://www.esajournals.org/loi/ecol [ Links ]
Andersson, L. A. (1992). A provisional checklist of Neotropical Rubiaceae. Scripta Botanica Belgica, 1, 1-119. http://www.scimagojr.com/ [ Links ]
Barrett, S. C. H. (1992). Heterostylous genetic polymorphisms: model systems for evolutionary analysis. In S. C. H. Barrett (Ed.), Evolution and function of heterostyly (pp. 1-29), Berlin: Springer-Verlag. [ Links ]
Brys, R., Jacquemyn, H., Endels, P., Hermy, M., & Blust, G. (2003). The relationship between reproductive success and demographic structure in remnant populations of Primula veris. Acta Oecologica, 24, 247-253. [ Links ]
Byers, D. L. (1995). Pollen quantity and quality as explanations for low seed set in small populations exemplified by Eupatorium (Asteraceae). American Journal of Botany, 82, 1000-1006. [ Links ]
Cruz, C. D. (1997). Programa genes: aplicativo computacional em genética e estatística. Viçosa, MG: Editora UFV. [ Links ]
Cunningham, S. A. (2000). Effects of habitat fragmentation on the reproductive ecology of four plant species in Mallee woodland. Conservation Biology, 14, 758-768. http://onlinelibrary.wiley.com/journal/ [ Links ]
De Sanctis, M., Afo, M., Attore, F., Francesconi, F., & Bruno, F. (2010). Effects of habitat configuration and quality on species richness and distribution in fragmented forest near Rome. Journal of Vegetation Science, 21, 55-65. [ Links ]
Donaldson, J., Nänni, I., & Zachariades, C. (2002). Effects of habitat fragmentation on pollinator diversity and plant reproductive success in Renosterveld shrublands of South Africa. Conservation Biology, 16, 1267-1276. [ Links ]
Ellstrand, N. C., & Elam, D. R. (1993). Population genetic consequences of small population size: implications for plant conservation. Annual Review of Ecology and Systematics, 24, 271-242. [ Links ]
Excoffier, L, Smouse, P. E., & Quattro, J. M. (1992). Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction sites. Genetics, 131, 479-491. [ Links ]
Excoffier, L., Laval, G., & Schneider, S. (2006). Arlequin v. 3.01. An integrated software package for population genetics data analysis. Computational and Molecular Population Genetics Lab., University of Berne, Berne. [ Links ]
Fahrig, L. (2003). Effects of habitat fragmentation on biodiversity. Annual Review of Ecology and Systematics, 34, 487-515. [ Links ]
Frakham, R., Ballou, J. D., & Briscoe, D. A. (2002). Introduction to conservation genetics. Cambridge: Cambridge University Press. [ Links ]
Fundação S.O.S Mata Atlântica http://www.sosmataatlantica.org.br. [ Links ]
Ganders, F. R. (1979). The biology of heterostyly. New Zealand Journal of Botany, 17, 607-635. http://www.researchgate.net/journal/0028-825X [ Links ]
Gomes, V., Collevatti, R. G., Silveira, F. A. O., & Fernandes, G. W. (2004). The distribution of genetic variability in Baccharis concinna (Asteraceae) an endemic, dioecious and threatened shrub of rupestrian fields of Brazil. Conservation Genetics, 5, 157-165. [ Links ]
González-Astorga, J., & Nuñez-Farfán, J. (2001). Effect of habitat fragmentation on the genetic structure of the narrow endemic Brongniartia vazquezii. Evolutionary Ecology Research, 3, 861-872. [ Links ]
Goulart, M. F., Ribeiro, S. P., & Lovato, M. B. (2005). Genetic, morphological and spatial characterization of two populations of Mabea fistulifera Mart. (Euphorbiaceae), in different successional stages. Brazilian Archives of Biology and Technology, 48, 275-284. [ Links ]
Jennersten, O. (1988). Pollination in Dianthus deltoids (Caryophyllaceae): effects of habitat fragmentation on visitation and seed set. Conservation Biology, 2, 359-366. [ Links ]
Kang, U., Chang, C. S., & Kim, Y. S. (2000). Genetic structure and conservation of the rare endemic Abeliophyllum distichum Nakai (Oleaceae) in Korea. Journal of Plant Research, 113, 127-138. [ Links ]
Kearns, C. A., Inouye, D. W., & Waser N. M. (1998). Endangered mutualisms: the conservation of plantpollinator interactions. Annual Review of Ecology and Systematics, 29, 83-112. [ Links ]
Koch, A. K., Silva, P. C., & Silva, C. A. (2010). Reproductive biology of Psychotria carthagenensis (Rubiaceae), a distylous species of riparian forest fragments, West Central Brazil. Rodriguesia, 61, 551-558. [ Links ]
Krauss, S. L., Hermanutz, L., Hopper, S. D., & Coates, D. J. (2007). Population size effects on seeds and seedlings from fragmented eucalypt populations: Implications for seed sourcing for ecological restoration. Australian Journal of Botany, 55, 390-399. [ Links ]
Li, A., & Ge, S. (2001). Genetic variation and clonal diversity of Psammochloa villosa (Poaceae) detected by ISSR markers. Annals of Botany, 87, 585-590. [ Links ]
Lopes, L. E., & Buzato, S. (2007). Variation in pollinator assemblages in a fragmented landscape and its effects on reproductive stages of a self-incompatible treelet, Psychotria suterella (Rubiaceae). Oecologia, 154, 305-314. [ Links ]
Mantel, N. (1967). The detection of disease clustering a generalized regression approach. Cancer Research, 27, 209-220. [ Links ]
Marangon, L. C., Soares, J. J., & Feliciano, A. (2003). Mata da Pedreira arboreal floristics, Viçosa, Minas Gerais, Brazil. Journal of Brazilian Forest Science, 27, 207-215. [ Links ]
Nei, M. (1973). Analysis of genetic diversity in subdivided populations (Population structure/genetic variability/heterozygosity/gene differentiation). Proceedings of the National Academy of Sciences of the United States of America, 70, 3321-3323. http://www.pnas.org/ [ Links ]
Nei, M., & Li, W. H. (1979). Mathematical model for studying genetic variation in terms of restriction endonucleases. Proceedings of the National Academy of Sciences of the United States of America, 76, 5269-5273. [ Links ]
Nybom, H., & Bartish, I. V. (2000). Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants. Perspectives in Plant Ecology, Evolution and Systematics, 3, 92-98. [ Links ]
Ohara, M., Tomimatsu, H., Takada, T., & Kawano, S. (2006). Importance of life history studies for conservation of fragmented populations: a case study of the understory herb, Trillium camschatcense. Plant Species Biology, 2, 11-12. [ Links ]
Oliveira, L. O., & Martins, E. R. (2002). A quantitative assessment of genetic erosion in ipecac (Psychotria ipecacuanha). Genetic Resources and Crop Evolution, 49, 607-617. [ Links ]
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, V. N., Underwood, E. C., Kassem, K. R. (2001). Terrestrial ecoregions of the world: A new map of life on earth. Bioscience, 51, 933-938. [ Links ]
Pereira, Z. V., Vieira, M. F., & Carvalho-Okano, R. M. (2006). Fenologia de floração, morfologia floral e sistema de incompatibilidade em espécies distílicas de Rubiaceae em fragmento florestal do sudeste brasileiro. Revista Brasileira de Botanica, 29, 471-480. [ Links ]
Pires, A. S., Fernandes, F. A. S., & Barros, C. S. (2006). Vivendo em um mundo em pedaços: efeitos da fragmentação florestal sobre comunidades e populações animais. In C. F. D. Rocha, H. G. Bergallo, M. Van Sluys, & M. A. S. Alves (Eds.), Biologia da conservação: essências (pp. 231-260) São Carlos, SP: Rima Editora. [ Links ]
Ramos, F. N., & Santos, F. A. M. (2005). Phenology of Psychotria tenuinervis (Rubiaceae) in Atlantic Forest fragments: fragment and habitat scales. Canadian Journal of Botany, 83, 1305-1316. [ Links ]
Ribon, R., Simon, J. E., & Mattos, G. T. (2003). Bird extinctions in Atlantic Forest fragments of the Viçosa region, Southeastern Brazil. Conservation Biology, 17, 1727-1839. [ Links ]
Rizzini, C. T. (1992). Tratado de fitogeografia do Brasil: aspectos ecológicos, sociológicos e florísticos. São Paulo, SP: Âmbito Cultural. [ Links ]
Rohlf, F. J. (2005). NTSYS-pc. Numerical taxonomy and multivariate analysis system, version 2.2. Exeter Software, Setauket. [ Links ]
Rossi, A. A. B., Oliveira, L. O., Venturini, B. A., & Silva, R. S. (2009). Genetic diversity and geographic differentiation of disjunct Atlantic and Amazonian populations of Psychotria ipecacuanha (Rubiaceae). Genetica, 136, 57-67. [ Links ]
Schaal, B. A., Hayworth, D. A., Olsen, K. M., Rauscher, J. T., & Smith, W. A. (1998). Phylogeographic studies in plants: problems and prospects. Molecular Ecology, 7, 465-474. [ Links ]
Shore, J. S., & Barrett, S. C. H. (1984). The effect of pollination intensity and incompatible pollen on seed set in Turnera ulmifolia (Turneraceae). Canadian Journal of Botany, 62, 1298-1303. [ Links ]
Spears, E. E. (1987). Island and mainland pollination ecology of Centrosema virginatum and Opuntia stricta. Journal of Ecology, 75, 351-362. [ Links ]
Stat Soft Inc. (2002). Statistica for Windows (Computer Program Manual). StatSoft Inc., Tulsa. [ Links ]
Steffan-Dewenter, I., & Tscharntke, T. (1199). Effects of habitat isolation on pollinator communities and seed set. Oecologia, 121, 78-88.
Tabarelli, M., Silva, J. M. C., & Gascon, C. (2004). Forest fragmentation, synergisms and the impoverishment of neotropical forests. Biodiversity and Conservation, 13, 1419-1425. [ Links ]
Taki, H., Kevan, P. G., & Ascher, J. S. (2007). Landscape effects of forest loss in a pollination system. Landscape Ecology, 22, 1575-1587. [ Links ]
Van der Pijl, L. (1982). Principles of dispersal in higher plants. New York: Springer-Verlag. [ Links ]
Wolf, A. T., & Harrison, S. P. (2001). Effects of hábitat size and patch isolation on reproductive success of the serpentine morning glory. Conservation Biology, 15, 111-121. [ Links ]
Yeh, F. C., Yang, R., & Boyle, T. (1999). Popgene. Microsoft Windows-based freeware for population genetic analysis. Release 1.31. Edmonton, University of Alberta. [ Links ]
Aguilar, R., & Galetto, L. (2004). Effects of forest fragmentation on male and female reproductive success in Cestrum parqui (Solanaceae). Oecologia, 138, 513-520. http://link.springer.com/journal/442 [ Links ]
Aizen, M. A., & Feinsinger, P. (1994). Forest fragmentation, pollination, and plant reproduction in a Chaco dry forest, Argentina. Ecology, 75, 330-351. http://www.esajournals.org/loi/ecol [ Links ]
Andersson, L. A. (1992). A provisional checklist of Neotropical Rubiaceae. Scripta Botanica Belgica, 1, 1-119. http://www.scimagojr.com/ [ Links ]
Barrett, S. C. H. (1992). Heterostylous genetic polymorphisms: model systems for evolutionary analysis. In S. C. H. Barrett (Ed.), Evolution and function of heterostyly (pp. 1-29), Berlin: Springer-Verlag. [ Links ]
Brys, R., Jacquemyn, H., Endels, P., Hermy, M., & Blust, G. (2003). The relationship between reproductive success and demographic structure in remnant populations of Primula veris. Acta Oecologica, 24, 247-253. [ Links ]
Byers, D. L. (1995). Pollen quantity and quality as explanations for low seed set in small populations exemplified by Eupatorium (Asteraceae). American Journal of Botany, 82, 1000-1006. [ Links ]
Cruz, C. D. (1997). Programa genes: aplicativo computacional em genética e estatística. Viçosa, MG: Editora UFV. [ Links ]
Cunningham, S. A. (2000). Effects of habitat fragmentation on the reproductive ecology of four plant species in Mallee woodland. Conservation Biology, 14, 758-768. http://onlinelibrary.wiley.com/journal/ [ Links ]
De Sanctis, M., Afo, M., Attore, F., Francesconi, F., & Bruno, F. (2010). Effects of habitat configuration and quality on species richness and distribution in fragmented forest near Rome. Journal of Vegetation Science, 21, 55-65. [ Links ]
Donaldson, J., Nänni, I., & Zachariades, C. (2002). Effects of habitat fragmentation on pollinator diversity and plant reproductive success in Renosterveld shrublands of South Africa. Conservation Biology, 16, 1267-1276. [ Links ]
Ellstrand, N. C., & Elam, D. R. (1993). Population genetic consequences of small population size: implications for plant conservation. Annual Review of Ecology and Systematics, 24, 271-242. [ Links ]
Excoffier, L, Smouse, P. E., & Quattro, J. M. (1992). Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction sites. Genetics, 131, 479-491. [ Links ]
Excoffier, L., Laval, G., & Schneider, S. (2006). Arlequin v. 3.01. An integrated software package for population genetics data analysis. Computational and Molecular Population Genetics Lab., University of Berne, Berne. [ Links ]
Fahrig, L. (2003). Effects of habitat fragmentation on biodiversity. Annual Review of Ecology and Systematics, 34, 487-515. [ Links ]
Frakham, R., Ballou, J. D., & Briscoe, D. A. (2002). Introduction to conservation genetics. Cambridge: Cambridge University Press. [ Links ]
Fundação S.O.S Mata Atlântica http://www.sosmataatlantica.org.br. [ Links ]
Ganders, F. R. (1979). The biology of heterostyly. New Zealand Journal of Botany, 17, 607-635. http://www.researchgate.net/journal/0028-825X [ Links ]
Gomes, V., Collevatti, R. G., Silveira, F. A. O., & Fernandes, G. W. (2004). The distribution of genetic variability in Baccharis concinna (Asteraceae) an endemic, dioecious and threatened shrub of rupestrian fields of Brazil. Conservation Genetics, 5, 157-165. [ Links ]
González-Astorga, J., & Nuñez-Farfán, J. (2001). Effect of habitat fragmentation on the genetic structure of the narrow endemic Brongniartia vazquezii. Evolutionary Ecology Research, 3, 861-872. [ Links ]
Goulart, M. F., Ribeiro, S. P., & Lovato, M. B. (2005). Genetic, morphological and spatial characterization of two populations of Mabea fistulifera Mart. (Euphorbiaceae), in different successional stages. Brazilian Archives of Biology and Technology, 48, 275-284. [ Links ]
Jennersten, O. (1988). Pollination in Dianthus deltoids (Caryophyllaceae): effects of habitat fragmentation on visitation and seed set. Conservation Biology, 2, 359-366. [ Links ]
Kang, U., Chang, C. S., & Kim, Y. S. (2000). Genetic structure and conservation of the rare endemic Abeliophyllum distichum Nakai (Oleaceae) in Korea. Journal of Plant Research, 113, 127-138. [ Links ]
Kearns, C. A., Inouye, D. W., & Waser N. M. (1998). Endangered mutualisms: the conservation of plantpollinator interactions. Annual Review of Ecology and Systematics, 29, 83-112. [ Links ]
Koch, A. K., Silva, P. C., & Silva, C. A. (2010). Reproductive biology of Psychotria carthagenensis (Rubiaceae), a distylous species of riparian forest fragments, West Central Brazil. Rodriguesia, 61, 551-558. [ Links ]
Krauss, S. L., Hermanutz, L., Hopper, S. D., & Coates, D. J. (2007). Population size effects on seeds and seedlings from fragmented eucalypt populations: Implications for seed sourcing for ecological restoration. Australian Journal of Botany, 55, 390-399. [ Links ]
Li, A., & Ge, S. (2001). Genetic variation and clonal diversity of Psammochloa villosa (Poaceae) detected by ISSR markers. Annals of Botany, 87, 585-590. [ Links ]
Lopes, L. E., & Buzato, S. (2007). Variation in pollinator assemblages in a fragmented landscape and its effects on reproductive stages of a self-incompatible treelet, Psychotria suterella (Rubiaceae). Oecologia, 154, 305-314. [ Links ]
Mantel, N. (1967). The detection of disease clustering a generalized regression approach. Cancer Research, 27, 209-220. [ Links ]
Marangon, L. C., Soares, J. J., & Feliciano, A. (2003). Mata da Pedreira arboreal floristics, Viçosa, Minas Gerais, Brazil. Journal of Brazilian Forest Science, 27, 207-215. [ Links ]
Nei, M. (1973). Analysis of genetic diversity in subdivided populations (Population structure/genetic variability/heterozygosity/gene differentiation). Proceedings of the National Academy of Sciences of the United States of America, 70, 3321-3323. http://www.pnas.org/ [ Links ]
Nei, M., & Li, W. H. (1979). Mathematical model for studying genetic variation in terms of restriction endonucleases. Proceedings of the National Academy of Sciences of the United States of America, 76, 5269-5273. [ Links ]
Nybom, H., & Bartish, I. V. (2000). Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants. Perspectives in Plant Ecology, Evolution and Systematics, 3, 92-98. [ Links ]
Ohara, M., Tomimatsu, H., Takada, T., & Kawano, S. (2006). Importance of life history studies for conservation of fragmented populations: a case study of the understory herb, Trillium camschatcense. Plant Species Biology, 2, 11-12. [ Links ]
Oliveira, L. O., & Martins, E. R. (2002). A quantitative assessment of genetic erosion in ipecac (Psychotria ipecacuanha). Genetic Resources and Crop Evolution, 49, 607-617. [ Links ]
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, V. N., Underwood, E. C., Kassem, K. R. (2001). Terrestrial ecoregions of the world: A new map of life on earth. Bioscience, 51, 933-938. [ Links ]
Pereira, Z. V., Vieira, M. F., & Carvalho-Okano, R. M. (2006). Fenologia de floração, morfologia floral e sistema de incompatibilidade em espécies distílicas de Rubiaceae em fragmento florestal do sudeste brasileiro. Revista Brasileira de Botanica, 29, 471-480. [ Links ]
Pires, A. S., Fernandes, F. A. S., & Barros, C. S. (2006). Vivendo em um mundo em pedaços: efeitos da fragmentação florestal sobre comunidades e populações animais. In C. F. D. Rocha, H. G. Bergallo, M. Van Sluys, & M. A. S. Alves (Eds.), Biologia da conservação: essências (pp. 231-260) São Carlos, SP: Rima Editora. [ Links ]
Ramos, F. N., & Santos, F. A. M. (2005). Phenology of Psychotria tenuinervis (Rubiaceae) in Atlantic Forest fragments: fragment and habitat scales. Canadian Journal of Botany, 83, 1305-1316. [ Links ]
Ribon, R., Simon, J. E., & Mattos, G. T. (2003). Bird extinctions in Atlantic Forest fragments of the Viçosa region, Southeastern Brazil. Conservation Biology, 17, 1727-1839. [ Links ]
Rizzini, C. T. (1992). Tratado de fitogeografia do Brasil: aspectos ecológicos, sociológicos e florísticos. São Paulo, SP: Âmbito Cultural. [ Links ]
Rohlf, F. J. (2005). NTSYS-pc. Numerical taxonomy and multivariate analysis system, version 2.2. Exeter Software, Setauket. [ Links ]
Rossi, A. A. B., Oliveira, L. O., Venturini, B. A., & Silva, R. S. (2009). Genetic diversity and geographic differentiation of disjunct Atlantic and Amazonian populations of Psychotria ipecacuanha (Rubiaceae). Genetica, 136, 57-67. [ Links ]
Schaal, B. A., Hayworth, D. A., Olsen, K. M., Rauscher, J. T., & Smith, W. A. (1998). Phylogeographic studies in plants: problems and prospects. Molecular Ecology, 7, 465-474. [ Links ]
Shore, J. S., & Barrett, S. C. H. (1984). The effect of pollination intensity and incompatible pollen on seed set in Turnera ulmifolia (Turneraceae). Canadian Journal of Botany, 62, 1298-1303. [ Links ]
Spears, E. E. (1987). Island and mainland pollination ecology of Centrosema virginatum and Opuntia stricta. Journal of Ecology, 75, 351-362. [ Links ]
Stat Soft Inc. (2002). Statistica for Windows (Computer Program Manual). StatSoft Inc., Tulsa. [ Links ]
Steffan-Dewenter, I., & Tscharntke, T. (1199). Effects of habitat isolation on pollinator communities and seed set. Oecologia, 121, 78-88.
Tabarelli, M., Silva, J. M. C., & Gascon, C. (2004). Forest fragmentation, synergisms and the impoverishment of neotropical forests. Biodiversity and Conservation, 13, 1419-1425. [ Links ]
Taki, H., Kevan, P. G., & Ascher, J. S. (2007). Landscape effects of forest loss in a pollination system. Landscape Ecology, 22, 1575-1587. [ Links ]
Van der Pijl, L. (1982). Principles of dispersal in higher plants. New York: Springer-Verlag. [ Links ]
Wolf, A. T., & Harrison, S. P. (2001). Effects of hábitat size and patch isolation on reproductive success of the serpentine morning glory. Conservation Biology, 15, 111-121. [ Links ]
Yeh, F. C., Yang, R., & Boyle, T. (1999). Popgene. Microsoft Windows-based freeware for population genetic analysis. Release 1.31. Edmonton, University of Alberta. [ Links ]
*Correspondencia a:
Celice A. Silva. Departamento de Ciências Biológicas, Centro de Estudos Pesquisa e Desenvolvimento Agro-ambiental (CPEDA), Universidade do Estado de Mato Grosso, campus de Tangará da Serra, Tangará da Serra, Mato Grosso, Brazil, CEP: 78300-000; celicealexandre645@hotmail.com
Milene F. Vieira. Departamento de Biologia Vegetal, Universidade Federal de Viçosa, Viçosa, Minas Gerais, Brazil; mfvieira@ufv.br
Rita M. de Carvalho-Okano. Departamento de Biologia Vegetal, Universidade Federal de Viçosa, Viçosa, Minas Gerais, Brazil; carvalho@ufv.br
Luiz O. de Oliveira. Departamento de Bioquímica e Biologia Molecular, Universidade Federal de Viçosa, Viçosa, Minas Gerais, Brazil; luiz.ufv@hotmail.com
1. Departamento de Ciências Biológicas, Centro de Estudos Pesquisa e Desenvolvimento Agro-ambiental (CPEDA), Universidade do Estado de Mato Grosso, campus de Tangará da Serra, Tangará da Serra, Mato Grosso, Brazil, CEP: 78300-000; celicealexandre645@hotmail.com
2. Departamento de Biologia Vegetal, Universidade Federal de Viçosa, Viçosa, Minas Gerais, Brazil; mfvieira@ufv.br
3. Departamento de Biologia Vegetal, Universidade Federal de Viçosa, Viçosa, Minas Gerais, Brazil; carvalho@ufv.br
4. Departamento de Bioquímica e Biologia Molecular, Universidade Federal de Viçosa, Viçosa, Minas Gerais, Brazil; luiz.ufv@hotmail.com
Received 07-i-2013. Corrected 20-VII-2013. Accepted 20-VIII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}