










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.1 San José Jan./Mar. 2014
In vitro propagation of Cyathea atrovirens (Cyatheaceae): spore storage and sterilization conditions
Propagación In vitro de Cyathea atrovirens (Cyatheaceae): spore storage and sterilization conditions
Propagación In vitro de Cyathea atrovirens (Cyatheaceae): spore storage and sterilization conditions
*Dirección para correspondencia:
Abstract
Cyathea atrovirens occurs in a wide range of habitats in Brazil, Paraguay, Uruguay and Argentina. In the Brazilian State of Rio Grande do Sul, this commonly found species is a target of intense exploitation, because of its ornamental characteristics. The in vitro cultura is an important tool for propagation which may contribute toward the reduction of extractivism. However, exogenous contamination of spores is an obstacle for the success of aseptic long-term cultures. This study evaluated the influence of different sterilization methods combined with storage conditions on the contamination of the in vitro cultures and the gametophytic development of C. atrovirens, in order to establish an efficient propagation protocol. Spores were obtained from plants collected in Novo Hamburgo, State of Rio Grande do Sul, Brazil. In the first experiment, spores stored at 7oC were surface sterilized with 0.5, 0.8 and 2% of sodium hypochlorite (NaClO) for 15 minutes and sown in Meyer’s culture medium. The cultures were maintained in a growth room at 26±1ºC for a 12-h photoperiod and photon flux density of 100μmol/m2/s provided by cool white fluorescent light. Contamination was assessed at 60 days, and gametophytic development was scored at 30, 60, 120 and 130 days of in vitro culture, analyzing 300 individuals for each treatment. There was no significant difference in culture contamination among the different sodium hypochlorite concentrations tested, and all treatments allowed for the development of cordiform gametophytes at 130 days of culture. In the second experiment, spores stored at 7 and -20°C were divided into two groups. Half of the spores were surface sterilized with 2% of NaClO for 15 minutes and the other half was not sterilized. All spores were sown in Meyer’s medium supplemented with one of the following antibiotics: nystatin, Micostatin® and actidione. The culture conditions and the procedures used for evaluating contamination and gametophytic development were the same described for the first experiment. No contamination was observed in spores stored at -20°C and treated with NaClO and actidione. In all treatments, cordiform gametophytes presenting antheridia were observed at 120 days. The percentages of these gametophytes increased from 120 to 130 days and no significant differences were observed among treatments. Archegonia were observed on cordiform gametophytes at 130 days. The findings provide data relevant to in vitro propagation procedures of this species, which may increase the availability of plants for ornamental purposes, therefore contributing to the reduction of the exploitation of endangered tree ferns species. Rev. Biol. Trop. 62 (1): 299-308. Epub 2014 March 01.
Key words: actidione, antibiotic, gametophyte, germination, in vitro culture, surface sterilization, tree fern.
Resumen
Cyathea atrovirens (Langsd. & Fisch.) Domin (Cyatheaceae) se presenta en una amplia gama de hábitats en Brasil, Paraguay, Uruguay y Argentina. Debido a sus características ornamentales, la especie es objeto de intensa explotación. El cultivo in vitro es una herramienta importante para la propagación lo que puede contribuir a la reducción del impacto de las actividades extractivas. Sin embargo, la contaminación exógena de esporas es un obstáculo para el éxito de cultivos asépticos a largo plazo. Este estudio evaluó la influencia de diferentes métodos de esterilización en combinación con las condiciones de almacenamiento sobre la contaminación de los cultivos in vitro y el desarrollo gametofítico de C. atrovirens. En el primer experimento, las esporas almacenadas a 7°C se esterilizaron superficialmente con 0.5, 0.8 y 2% de hipoclorito de sodio (NaClO) durante 15 minutos y se sembraron en medio de cultivo de Meyer. Aunque no hubo diferencia en la contaminación de lós cultivos entre las concentraciones de hipoclorito de sodio de las diferentes pruebas, en el tratamiento con 2% NaClO se observó un mayor porcentaje de gametofitos cordiformes a los 130 días. En el segundo experimento, las esporas almacenadas a 7 y -20°C fueron divididas en dos grupos. La mitad de las esporas se esterilizaron con 2% de NaClO durante 15 minutos y la otra mitad no fue esterilizada. Todas las esporas se sembraron en medio de Meyer suplementado con uno de los siguientes antibióticos: nistatina, Micostatin® o actidiona. No se observó contaminación de las esporas almacenadas a -20°C y tratadas con NaClO y actidiona. En todos los tratamientos, se observaron gametofitos cordiformes con anteridios y arquegonios. Los resultados proporcionan datos relevantes para la propagación in vitro de C. atrovirens, que pueden aumentar la disponibilidad de las plantas para fines ornamentales, contribuyendo así a la reducción de la exploracción de las especies de helechos arborescentes en peligro de extinción.
Palabras clave: actidiona, antibiótico, cultivo in vitro, esterilización superficial, gametófito, germinación, helecho arborescente.
Tree ferns are an important component of tropical rainforests and among them Cyatheaceae is a noteworthy family, being represented by approximately 170 species in the Neotropics (Tryon & Tryon, 1982) and 23 species in South and Southeastern regions of Brazil (Windisch & Santiago, 2013). Cyathea atrovirens (Langsd. & Fisch.) Domin (Cyatheaceae) occurs in a wide range of habitats in Brazil, Paraguay, Argentina (Ponce, 1996) and Uruguay (Marquez & Brussa, 2011). In the Brazilian State of Rio Grande do Sul, the species is commonly found in open or moderately shaded humid or swampy places and in areas largely affected by human action, such as roadsides (Lorscheitter, Ashraf, Windisch & Mosbrugger, 1999). Cyathea atrovirens forms 6m high caudices, with a sheath of adventitious roots at the persistent petiole bases. Petioles and leaves are, respectively, up to 1.10 and 3m long, and the laminas are bipinnate-pinnatifid to pinatilobate (Sehnem, 1978; Fernandes, 1997). The spores are tetrahedral-globose and present a triangular equatorial limb with prominent rounded angles and straight to slightly depressed sides (Lorscheitter et al., 1999).
Habitat destruction and fragmentation are the main factors related to the reduction of tree fern populations. In addition, tree ferns are exploited because of their gardening potential (Windisch, 2002). Leaves (Tryon & Tryon, 1982) and entire plants (Schmitt & Windisch, 2012) of C. atrovirens are used for ornamental purposes. Caudices of older plants, when presenting a sheath of adventitious roots at the base, are used to manufacture fiber handicrafts (Fernandes, 2000).
The in vitro culture is an important tool for plant propagation which may contribute toward the reduction of exploitation of natural populations (Caldecott, Jenkins, Johnson & Groombridge, 1996; Giudice, Giacosa, Luna, Yañez & de la Sota, 2011). Spore germination is considered the most efficient method of in vitro culture of ferns and lycophytes (Pence, 2008) and studies with different species can be cited (e.g. Kiss & Kiss, 1998; Bertrand, Albuerne, Fernandéz, González & Sánchez-Tamés, 1999; Fernandéz, Bertrand & Sánchez-Tamés, 1999; Kuriyama, Kobayashi & Maeda, 2004; Souza, Medeiros & Mendes, 2007).
In general, studies analyzing the viability of the spores and the initial ontogenetic development of gametophytes do not refer to contamination (Bertrand, Albuerne, Fernández, González & Sánchez-Tamés, 1999; Fernandéz et al., 1999; Chen, Cheng, Liu & Jiao, 2008), despite exogenous contamination of spores is an obstacle for the success of aseptic long-term cultures (Dyer, 1979; Simabukuro, Dyer & Felippe, 1998). The substances most commonly used for plant tissue surface sterilization are ethylic alcohol and compounds with chlorine, like calcium hypochlorite and sodium hypochlorite (Wu, Ping-Tin, Li-Ping & Long-Qing, 2009). Recently, the sodium dichloroisocyanurate was described as an efficient sterilizing agent for spores (Barnicoat, Cripps, Kendon & Sarasan, 2011). Addition of antimicrobial agents to the culture medium has also been tested. Among antibiotics that can be used to avoid the in vitro contamination of plants (Kyte & Kleyn, 1996), Micostatin® (Ranal, 1991), nystatin (Dyer, 1979; Simabukuro et al., 1998; Quintanilla, Amigo, Pangua & Pajaron, 2002) and streptomycin (Cox, Bhatia & Ashwath, 2003) were tested for ferns, although without total elimination of the microorganisms from the cultures. The treatment of plant tissues with sodium hypochlorite, followed by the addition of the fungicide Benlate® (Benomil - Dupont) to the culture medium has also been used for fern species (Brum & Randi, 2006; Begnini & Randi, 2009; Viviani & Randi, 2008; Santos, Lehmann, Santos & Randi, 2010). However, this fungicide had its manufacture discontinued since 2001 (Dupont, 2012). Unfortunately, most surface sterilization treatments may also reduce fern spore germination (Hamilton & Chaffin, 1998; Simabukuro et al., 1998).
Storage conditions under different temperatures have been tested focusing on the survival and germination of spores (Simabukuro et al., 1998; Rogge, Viana & Randi, 2000; Quintanilla et al., 2002; Begnini & Randi, 2009; Santos et al., 2010). The viability of spores from different species has been investigated after cold storage (for a revision, see Ranker & Haufler, 2008), and low temperature was considered as minimizing bacterial and fungal contamination without decreasing spore viability (Quintanilla et al., 2002).
The aim of this study was to test the influence of different sterilization methods combined with storage conditions on the contamination of the in vitro cultures and the gametophytic development of C. atrovirens, in order to contribute to the establishment of an efficient propagation protocol contributing to management programs of this species.
Material and methods
Fertile leaves of C. atrovirens were collected in the Parque Municipal Henrique Luis Roessler (29°40’54” S - 51º06’56” W, at 16.4m in elevation), which is a conservation area of the Rio dos Sinos basin in the municipality of Novo Hamburgo, State of Rio Grande do Sul, Brazil. The leaves were wrapped in smooth paper and kept at room temperatura to induce dehiscence of sporangia. Spores were filtered through lens cleaning tissue and stored. Two different experiments were carried out as follows.
Experiment I: In the first experiment, spore samples (~30mg) stored at 7°C for 30 days were distributed in microtubes of 1.5mL and surface sterilized in 1mL of sodium hypochlorite (NaClO) solution at concentrations of 0.5, 0.8 and 2.0% (v/v) for 15min. Spores immersed in 1mL of sterile distilled wáter instead of NaClO were used as control. After removing the supernatant, spores were rinsed in 1mL of sterile distilled water, and centrifuged at 2 000rpm for 3min. The washing and centrifugation steps were repeated three times. The spores were sown in petri dishes (9cm diameter, 10mg/dish) containing 30mL of Meyer’s medium (Meyer, Anderson & Swanson, 1955), supplemented with 0.25% (w/v) PhytagelTM, with pH adjusted to 6.0 before autoclaving. For each NaClO concentration tested and for the control, three repetitions were made. All procedures were carried out in a laminar hood. The cultures were maintained in a growth room at 26±1ºC for a 12-h photoperiod and photon flux density of 100μmol/m2/s provided by cool white fluorescent light. Once a month, 2mL of sterile distilled water (pH 6.0) were added to the culture medium in each dish, to stimulate the gametophytic development (Mendoza-Ruiz & Pérez-García, 2009).
To assess the contamination at 60 days of culture, a quantitative analysis was performed, using two printed paper grids, each containing 21 fields (each field of 1.5x1.5cm), a black one and a white one, for better visualization of fungi and bacteria. The grids were placed separately under each dish, covering the entire area, and the number of fields presenting macroscopically visible contamination was counted.
Gametophytic development was scored at 30, 60, 120 and 130 days of in vitro culture. Three slides (one slide per dish) from each treatment were analyzed, and 100 individuals (spores or young gametophytes) were counted on each slide (300 individuals per treatment). Individuals were classified according to their developmental stage (Rechenmacher, Schmitt & Droste, 2010) in the following classes: non-germinated spores (NG), gametophyte with chlorocyte and rhizoid (G1); filamentous gametophyte (G2); laminar gametophyte (G3) and cordiform gametophyte (G4). The criterion for germination was the emergence of the chlorocite or the rhizoid (Ranal, 1999).
Experiment II: In a second experiment, spore samples (~30 mg) stored at 7 and -20°C for 30 days were distributed in microtubes of 1.5mL. For each storage temperature, spores of three tubes were surface sterilized with 2% (v/v) NaClO for 15min, and spores of three tubes were immersed in sterile distilled wáter for the same time (control), according to the procedure described for the first experiment. The spores were sown in Petri dishes (9cm in diameter, 10 mg/dish), with 30mL of Meyer’s medium (pH 6.0 before autoclaving), supplemented with one of the following antibiotics: (a) 1mL of nystatin (Sigma-Aldrich) 10 000U/ mL, (b) 1mL of Micostatin® (Bristol-Myers Squibb), a commercial antibiotic containing 100 000U/mL of nystatin, and (c) 0.5g/L of actidione (Fluka, Sigma-Aldrich). The antibiotics were added to the medium after autoclaving and partial cooling. For each treatment, that combined a different antibiotic and storage temperature with or without previous surface sterilization, three dishes were used, totalizing 36 dishes. The culture conditions and the procedures used for evaluating contamination and gametophytic development were the same described for the first experiment.
The statistical analyses of the data were performed using the SPSS 17.0 and the BioEstat 5.0 softwares. The Shapiro-Wilk and Levene tests, respectively for goodness of fit for normality and for the homogeneity of variance (0.05) were applied before the analysis. Data related to contaminated fields in the first experiment, as well as data of gametophytic development in both experiments were submitted to the nonparametric Kruskal-Wallis test followed by the Student-Newman-Keuls test, at 0.05 significance. Data of gametophytic development in the second experiment were submitted to analysis of variance (ANOVA) and the pairwise comparison between means was performed by the parametric Tukey test at 0.05 significance (Zar, 1999).
Results
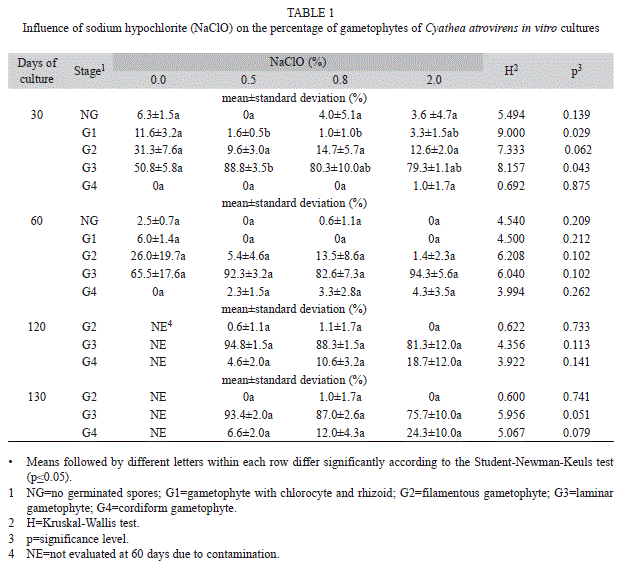
Experiment I: In the first experiment, contamination was observed in all cultures. A minor number of contaminated fields were visualized in cultures when spores were treated with 2.0% of NaClO (9.6) compared to 0% (21.0), 0.5% (14.0) and 0.8% of NaClO (18.6), although these differences were no statistically significant (Kruskal-Wallis, H=5.0600, p=0.1675). Germination and gametophytic development were observed in cultures from the three NaClO concentrations tested (Table 1). At 30 days of culture, most gametophytes were on laminar stage and cordiform gametophytes only have been observed in treatments with 2.0% of NaClO (Table 1). At 60 days, gametophytes in the first stage (with chlorocytes and rhizoids) were present only in the treatment without NaClO and most gametophytes were in laminar stage. In the following observations, there was an increase in the percentage of cordiform gametophytes in all treatments with NaClO. The highest percentage of this type of gametophytes was observed after treatment with 2% of NaClO, although without significant difference to the other treatments with this sterilizing agent (Table 1). Antheridia were observed in the control and in treatments with 0.5 and 2.0% of NaClO at 60 days, and also in treatment with 0.8% at 120 days. Archegonia were seen only in the treatment with 2% NaClO, at 120 days of cultivation.
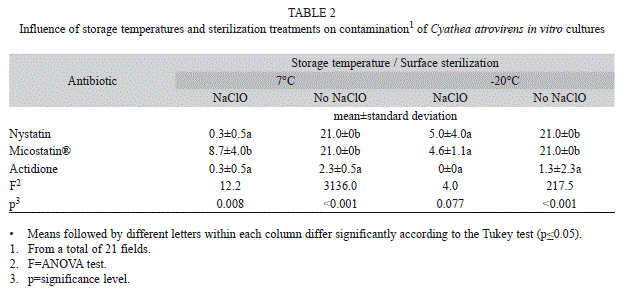
Although there was no statistically difference in culture contamination among the different sodium hypochlorite concentrations tested in the first experiment, the treatment with 2% NaClO allowed for numerically lower contamination and higher percentage of cordiform gametophytes at 130 days of culture. Therefore, this treatment was used in the second experiment. The lowest mean numbers of contaminated fields were observed when spores were surface sterilized with NaClO combined with the addition of antibiotics to the culture medium (Table 2). For the spores stored at 7°C and previously surface sterilized, the use of actidione did not differ significantly from the use of nystatin, which led to significantly less contamination than using Micostatin®. When there was no prior sterilization of spores stored at 7°C, the use of actidione provided significantly less contaminated fields in relation to the use of nystatin and Micostatin®. For spores stored at -20°C and previously surface sterilized, no significant difference among the antibiotics tested was verified. Contamination was low in treatments with nystatin and Micostatin ®, and the use of actidione eliminated contamination totally. For spores stored at -20°C, without prior sterilization with NaClO, the use of actidione showed the lowest mean contamination values, with significant difference compared to the other antibiotics tested (Table 2).
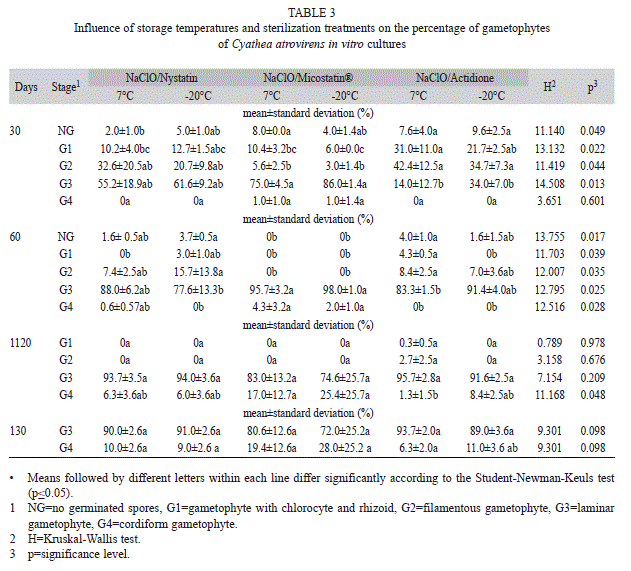
Experiment II: In the second experiment, the development of gametophytes was studied when subjected to antibiotic treatments and previous sterilization of spores with NaClO. At 30 days, the percentages of laminar gametophytes were significantly higher in treatments with Micostatin® than in treatments with actidione, and cordiform gametophytes could already be observed in treatments with this antibiotic (Table 3). At 60 days, most gametophytes were in laminar stage, and cordiform gametophytes were registered in treatments with nystatin and Micostatin®. There was an increase in the number of laminar gametophytes from 60 to 120 days of culture in the treatments with nystatin and actidione. In the presence of Micostatin®, the number of laminar gametophytes decreased, because some of them have already developed into cordiform gametophytes. However, no statistical differences were found on laminar gametophyte percentages among treatments. In all treatments, cordiform gametophytes presenting antheridia were observed at 120 days. At 130 days, no filamentous gametophytes were found, and in all treatments most gametophytes were of the laminar type. The percentages of cordiform gametophytes increased from 120 to 130 days and no significant differences were observed among treatments (Table 3). Archegonia were observed on cordiform gametophytes at 130 days.
Discussion
Although without significant statistical difference among treatments, the contamination of cultures coming from spores sterilized with 2% NaClO was about 50% lower tan the contamination of cultures from spores not treated with this substance. Due to exogenous contamination, surface sterilization of spores is the first step in an aseptic culture of ferns (Dyer, 1979). Cultures initiated with unsterilized biological material tend to contamínate after 10 days, as noted by Camloh (1999) for Platycerium bifurcatum (Cav.) C. Chr. And Rechenmacher et al. (2010) for C. atrovirens. On the other hand, although surface sterilizing treatments are commonly referred to as drastically reducing germination (Simabukuro et al., 1998), this did not occur in the present work.
The reduction of microorganisms of in vitro cultures observed in the present study for spores sterilized with NaClO, also was recorded for other tree ferns. Low fungal contamination and the elimination of bacterial contamination on Cyathea delgadii Sternb. (Cyatheaceae) were observed by Simabukuro et al. (1998) when using NaClO concentrations of 1.0, 3.0 and 5.0%. Souza et al. (2007) reported contamination by fungi and bacteria in about 50% of the culture dishes when spores of Dicksonia sellowiana Hook. (Dicksoniaceae) where sterilized in 2% of NaClO.
The surface sterilization of spores with NaClO associated with the addition of the fungicide Benlate® (benomyl) to the nutrient medium has been used as an alternative to eliminate contamination of fern cultures in Brazil (Renner & Randi, 2004; Brum & Randi, 2006; Begnini & Randi, 2009; Viviani & Randi, 2008; Santos et al., 2010). However, this fungicide is not a viable alternative, since it is no longer available in the market of this country. Nystatin is the active ingredient of the commercial antibiotic Micostatin®. In the present study, nystatin allowed less contamination than Micostatin®, when combined with the use of NaClO in spores stored at 7°C, although a 10-fold higher concentration of nystatin is present in 1mL of Micostatin® compared with the use of pure nystatin. Ranal (1991, 1999) did not found contamination by fungi in cultures of different fern species from spores stored at 4°C and developed in medium with Micostatin® (E.R. Squibb 10 000U/mL). On the other hand, according to Kozai (1991) and Souza et al. (2007), the presence of sucrose among the inactive ingredients of Micostatin® may favor the development of fungi and bacteria and thereby increase the rate of contamination.
Nystatin was previously used for spore surface sterilization. Filippini, Duz & Randi (1999) cited the use of NaClO and nystatin on in vitro culture of D. sellowiana. Quintanilla et al. (2002) considered the use of 100U/mL of nystatin efficient to reduce fungus and bacteria in wet storage of spores of five threatened fern species: Culcita macrocarpa C. Presl (Culcitaceae), Dryopteris aemula (Aiton) O. Kuntze (Dryopteridaceae), D. corleyi Fraser-Jenkins (Dryopteridaceae), D. guanchica Gibby and Jermy (Dryopteridaceae) and Woodwardia radicans (L.) Sm. (Blechnaceae). However, in these studies, no numerical data of contamination were presented. Simabukuro et al. (1998) observed that the best treatment for elimination of bacteria and fungi, in C. delgadii spore cultures, was the surface sterilization with 0.5% of calcium hypochlorite for two minutes and addition of 100U/mL of nystatin to the medium. However, contamination by fungi and bacteria persisted, with 18.3 and 48.3% of contaminated points, respectively, and only 52.7% of the spores germinated.
Actidione is an antibiotic indicated as inhibitor of bacteria and fungi (Panthier, Diem & Dommergues, 1979). In the present study, the use of actidione without previous surface sterilization of spores stored at 7 and -20oC led to significantly less contamination than the other antibiotics tested. The associated use of this antibiotic with previous surface sterilization of spores stored at -20°C allowed the total decontamination of the cultures. Rojas & Rondón (1995) showed in vitro growth inhibition of Fusarium decemcellulare Brick using actidione associated with Benlate, and suggested the use of these substances in Mangifera indica L. (Anacardiaceae) field cultures to control this harmful fungus.
Although sterilizing agents are used for in vitro culture of Cyatheaceae, there were no found reports in literature concerning the qualitative and quantitative influence of them on the development of gametophytes. In the present study, the data at 30 days of culture suggest that the gametophytic development has been delayed in treatments with actidione compared to the treatments with Micostatin®, in which higher percentages of laminar gametophytes and even cordiform gametophytes were observed. This delay was not maintained during the experiment, and there were no significant differences between treatments at the end of the experiment. Furthermore, the presence of reproductive structures at 120 and 130 days corroborate with the findings of Rechenmacher et al. (2010) for C. atrovirens cultures without using sterilizing agents, suggesting that the antimicrobial action of these agents does not seem to influence negatively the development of gametophytes. The increasing number of reproductive structures observed during the evaluations suggests the continuity of the developmental process after the study period of 130 days.
The surface sterilization of spores stored at -20°C associated with the use of actidione in the culture medium may be an efficient method for an aseptic in vitro culture of C. atrovirens, since it showed to eliminate the contamination and to allow the development of gametophytes with reproductive structures. Moreover, the findings provide data relevant to in vitro propagation procedures of this species, which may increase the availability of plants for ornamental purposes, therefore contributing to the reduction of the exploitation of endangered tree ferns species.
Acknowledgments
The authors are thankful to the Universidade Feevale for research support.
References
Barnicoat, H., Cripps, R., Kendon, J., & Sarasan, V. (2011). Conservation in vitro of rare and threatened ferns – case studies of biodiversity hotspot and island species. In Vitro Cellular and Developmental Biology, 47, 37-45. [ Links ]
Begnini, R. M., & Randi, A. M. (2009). Viabilidade de esporos de Dicksonia sellowiana Hook. (Cyatheales, Dicksoniaceae) e Rumohra adiantiformis (Forst.) Ching (Polypodiales, Dryopteridaceae) armazenados sob refrigeração. Insula, 38, 15-27. [ Links ]
Bertrand, A. M., Albuerne, M. A., Fernández, A., González, A., & Sánchez-Tamés, R. (1999). In vitro organogénesis of Polypodium cambricum. Plant Cell, Tissue and Organ Culture, 57, 65-69. [ Links ]
Brum, F. M. R., & Randi, A. M. (2006). Germination of spores and growth of gametophytes and sporophytes of Rumohra adiantiformis (Forst.) Ching (Dryopteridaceae) after spore cryogenic storage. Revista Brasileira de Botânica, 29, 489-495. [ Links ]
Caldecott, J. O., Jenkins, M. D., Johnson, T. H., & Groombridge, B. (1996). Priorities for conserving global species richness and endemism. Biodiversity Conservation, 5, 699-727. [ Links ]
Camloh, M. (1999). Spore age and sterilization affect germination and early gametophyte development of Platycerium bifurcatum. American Fern Journal, 89, 124-132. [ Links ]
Chen, G., Cheng, X., Liu, B., & Jiao, Y. (2008). Comparative studies on gametophyte morphology and development of seven species of Cyatheaceae. American Fern Journal, 98, 83-95. [ Links ]
Cox, J., Bhatia, P., & Ashwath, N. (2003). In vitro spore germination of the fern Schizaea dichotoma. Scientia Horticulturae, 97, 369-378. [ Links ]
Dupont. (2012). Benlate®: Fungicide. Retrieved from http://www2.dupont.com/Heritage/en_US/related_topics/benlate.html [ Links ]
Dyer, A. F. (1979). The culture of fern gametophytes for experimental investigation. In A. F. Dyer (Ed.), The experimental biology of ferns (pp. 253-305). London: Academic Press. [ Links ]
Fernandes, I. (1997). Taxonomia e fitogeografia de Cyatheaceae e Dicksoniaceae nas regiões sul e sudeste do Brasil. Tese de Doutorado, Universidade de São Paulo, São Paulo, Brazil. [ Links ]
Fernandes, I. (2000). Taxonomia dos representantes de Dicksoniaceae no Brasil. Pesquisas, 50, 5-26. [ Links ]
Fernandéz, H., Bertrand, A. M. & Sánchez-Tamés, R. (1999). Biological and nutritional aspects involved in fern multiplication. Plant Cell, Tissue and Organ Culture, 56, 211-214. [ Links ]
Filippini, E. C. P., Duz, S. R., & Randi, A. M. (1999). Light and storage in the germination of spores of Dicksonia selowiana (Presl.) Hook., Dicksoniaceae. Revista Brasileira de Botânica, 22, 21-26. [ Links ]
Giudice, G. E., Giacosa, J. P. R., Luna, M. L., Yañez, A., & de la Sota, E. R. (2011). Diversidad de helechos y licófitas de La Reserva Natural Punta Lara, Buenos Aires, Argentina. Revista de Biologia Tropical, 59, 1037-1046. [ Links ]
Hamilton, R., & Chaffin, C. (1998). The effect of surface sterilization on cultures of Ceratopteris richardii gametophytes. American Fern Journal, 88, 81-85. [ Links ]
Kozai, T. (1991). Photoautotrophic micropropagation. In Vitro Cellular and Developmental Biology, 27, 47-51. [ Links ]
Kiss, H. G., & Kiss, J. Z. (1998). Spore germination in populations of Schizaea pusilla from New Jersey and Nova Scotia. International Journal of Plant Science, 159, 848-852. [ Links ]
Kyte, L., & Kleyn, J. (1996). Plants from test tubes: an introduction to micropropagation. Portland: Timber Press. [ Links ]
Kuriyama, A., Kobayashi, T., & Maeda, M. (2004). Production of sporophytic plants of Cyathea lepifera, a tree fern, from in vitro cultured gametophyte. Journal of the Japanese Society of Horticulture Science, 73, 140-142. [ Links ]
Lorscheitter, M. L., Ashraf, A. R., Windisch, P. G., & Mosbrugger, V. (1999). Pteridophyte spores of Rio Grande do Sul flora, Brazil. Part II. Palaeontographica, 251, 71-235. [ Links ]
Marquez, G. J., & Brussa, C. A. (2011). First record of Cyatheaceae in Uruguay. American Fern Journal, 101, 205-205. [ Links ]
Mendoza-Ruiz, A., & Pérez-García, B. (2009). Morphogenesis of the gametophytes of eight mexican species of Blechnum (Blechnaceae). Acta Botanica Mexicana, 88, 59-72. [ Links ]
Meyer, B. S., Anderson, D. B., & Swanson, C. A. (1955). Laboratory Plant Physiology. New York: Van Nostrand. [ Links ]
Panthier, J. J., Diem, H. G., & Dommergues, Y. (1979). Rapid method to enumerate and isolate soil actinomycetes antagonistic towards rhizobia. Soil Biology & Biochemistry, 11, 443-445. [ Links ]
Pence, V. C. (2008). In vitro collecting for ex situ conservation. In T. A. Ranker, & C. Haufler (Ed.), Biology and Evolution of Ferns and Lycophytes (pp. 417-461). New York: Cambridge University Press. [ Links ]
Ponce, M. (1996). Pteridophyta. In F. O. Zuloaga, & O. Morrone (Eds.), Catálogo de las plantas vasculares de Argentina I: Pteridophyta, Gymnospermae y Angiospermae (Monocotyledoneae). Monographs in Systematic Botany from the Missouri Botanical Garden 60, 1-79. [ Links ]
Quintanilla, L. G., Amigo, J., Pangua, E., & Pajaron, S. (2002). Effect of storage method on spore viability in five globally threatened fern species. Annals of Botany, 90, 461-467. [ Links ]
Ranal, M. (1991). Germination of Polypodium hirsutissimum spores and antheridia formation in darkness. Revista Brasileira de Biologia, 51, 675-679. [ Links ]
Ranal, M. (1999). Effects of temperature on spore germination in some fern species from semideciduous mesophytic Forest. American Fern Journal, 89, 149-158. [ Links ]
Ranker, T. A., & Haufler, C. H. (2008). Biology and Evolution of Ferns and Lycophytes. New York: Cambridge University Press. [ Links ]
Rechenmacher, C., Schmitt, J. L. & Droste, A. (2010). Spore germination and gametophyte development of Cyathea atrovirens (Langsd. & Fisch.) Domin (Cyatheaceae) under different pH conditions. Brazilian Journal of Biology, 70, 1155-1160. [ Links ]
Renner, G. D. R., & Randi, A. M. (2004). Effects of sucrose and irradiance on germination and early gametophyte growth of the endangered tree fern Dicksonia sellowiana Hook. (Dicksoniaceae). Acta Botanica Brasilica, 18, 375-380. [ Links ]
Rogge, G. D., Viana, A. M., & Randi, A. M. (2000). Cryopreservation of spores of Dicksonia sellowiana, an endangered tree fern indigenous to South and Central America. Cryoletters, 21, 223-230. [ Links ]
Rojas, T., & Rondón, A. J. (1995). Control químico in vitro de Fusarium decemcellulare Brick aislado de mango. Agricultura Tropical, 45, 417-428. [ Links ]
Santos, E. P. G., Lehmann, D. R. M., Santos, M., & Randi, A. M. (2010). Spore germination of Gleichenella pectinata (Willd.) Ching (Polypodiopsida-Gleicheniaceae) at different temperatures, levels of light and pH. Brazilian Archives of Biology and Technology, 53, 1309-1318. [ Links ]
Schmitt, J. L., & Windisch, P. G. (2012). Caudex growth and phenology of Cyathea atrovirens (Langsd. & Fisch.) Domin (Cyatheaceae) in secondary forest, southern Brazil. Brazilian Journal of Biology, 72, 397-405. [ Links ]
Sehnem, A. (1978). Ciateáceas: Flora Ilustrada Catarinense. Itajaí: Herbário Barbosa Rodrigues. [ Links ]
Simabukuro, E. A., Dyer, A. F., & Felippe, G. M. (1998). The effect of sterilization and storage conditions on the viability of the spores of Cyathea delgadii Sternb. American Fern Journal, 88, 72-80. [ Links ]
Souza, R. N., Medeiros, M. B., & Mendes, R. A. (2007). Micropropagação e conservação do germoplasma de Dicksonia sellowiana (Hook.) (Dicksoniaceae) visando o repovoamento da área de influência do aproveitamento hidrelétrico Barra Grande, SC, RS. Anais do VIII Congresso de Ecologia do Brasil, Caxambu, Brazil. [ Links ]
Tryon, R. M., & Tryon, A. F. (1982). Ferns and allied plants with special reference to Tropical America. New York: Springer Verlag. [ Links ]
Viviani, D., & Randi, A. M. (2008). Effects of pH, temperature and light intensity on spore germination and growth analysis of young sporophytes of Polypodium lepidopterist (Langsd. & Fisch.) Kunze (Pteridophyta, Polypodiaceae). Rodriguésia, 59, 435-444. [ Links ]
Windisch, P. G. (2002). Fern conservation in Brazil. Fern Gazette, 16, 295-300. [ Links ]
Windisch, P. G., & Santiago, A. C. P. (2013). Cyatheaceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. Retrieved from http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB90850 [ Links ]
Wu, H., Ping-Tin, C., Li-Ping, Y., & Long-Qing, C. (2009). An efficient method for surface sterilization and sowing fern spores in vitro. American Fern Journal, 99, 226-230. [ Links ]
Zar, J. H. (1999). Biostatistical analyses. 4 ed. Upper Saddle River: Prentice-Hall. [ Links ]
Begnini, R. M., & Randi, A. M. (2009). Viabilidade de esporos de Dicksonia sellowiana Hook. (Cyatheales, Dicksoniaceae) e Rumohra adiantiformis (Forst.) Ching (Polypodiales, Dryopteridaceae) armazenados sob refrigeração. Insula, 38, 15-27. [ Links ]
Bertrand, A. M., Albuerne, M. A., Fernández, A., González, A., & Sánchez-Tamés, R. (1999). In vitro organogénesis of Polypodium cambricum. Plant Cell, Tissue and Organ Culture, 57, 65-69. [ Links ]
Brum, F. M. R., & Randi, A. M. (2006). Germination of spores and growth of gametophytes and sporophytes of Rumohra adiantiformis (Forst.) Ching (Dryopteridaceae) after spore cryogenic storage. Revista Brasileira de Botânica, 29, 489-495. [ Links ]
Caldecott, J. O., Jenkins, M. D., Johnson, T. H., & Groombridge, B. (1996). Priorities for conserving global species richness and endemism. Biodiversity Conservation, 5, 699-727. [ Links ]
Camloh, M. (1999). Spore age and sterilization affect germination and early gametophyte development of Platycerium bifurcatum. American Fern Journal, 89, 124-132. [ Links ]
Chen, G., Cheng, X., Liu, B., & Jiao, Y. (2008). Comparative studies on gametophyte morphology and development of seven species of Cyatheaceae. American Fern Journal, 98, 83-95. [ Links ]
Cox, J., Bhatia, P., & Ashwath, N. (2003). In vitro spore germination of the fern Schizaea dichotoma. Scientia Horticulturae, 97, 369-378. [ Links ]
Dupont. (2012). Benlate®: Fungicide. Retrieved from http://www2.dupont.com/Heritage/en_US/related_topics/benlate.html [ Links ]
Dyer, A. F. (1979). The culture of fern gametophytes for experimental investigation. In A. F. Dyer (Ed.), The experimental biology of ferns (pp. 253-305). London: Academic Press. [ Links ]
Fernandes, I. (1997). Taxonomia e fitogeografia de Cyatheaceae e Dicksoniaceae nas regiões sul e sudeste do Brasil. Tese de Doutorado, Universidade de São Paulo, São Paulo, Brazil. [ Links ]
Fernandes, I. (2000). Taxonomia dos representantes de Dicksoniaceae no Brasil. Pesquisas, 50, 5-26. [ Links ]
Fernandéz, H., Bertrand, A. M. & Sánchez-Tamés, R. (1999). Biological and nutritional aspects involved in fern multiplication. Plant Cell, Tissue and Organ Culture, 56, 211-214. [ Links ]
Filippini, E. C. P., Duz, S. R., & Randi, A. M. (1999). Light and storage in the germination of spores of Dicksonia selowiana (Presl.) Hook., Dicksoniaceae. Revista Brasileira de Botânica, 22, 21-26. [ Links ]
Giudice, G. E., Giacosa, J. P. R., Luna, M. L., Yañez, A., & de la Sota, E. R. (2011). Diversidad de helechos y licófitas de La Reserva Natural Punta Lara, Buenos Aires, Argentina. Revista de Biologia Tropical, 59, 1037-1046. [ Links ]
Hamilton, R., & Chaffin, C. (1998). The effect of surface sterilization on cultures of Ceratopteris richardii gametophytes. American Fern Journal, 88, 81-85. [ Links ]
Kozai, T. (1991). Photoautotrophic micropropagation. In Vitro Cellular and Developmental Biology, 27, 47-51. [ Links ]
Kiss, H. G., & Kiss, J. Z. (1998). Spore germination in populations of Schizaea pusilla from New Jersey and Nova Scotia. International Journal of Plant Science, 159, 848-852. [ Links ]
Kyte, L., & Kleyn, J. (1996). Plants from test tubes: an introduction to micropropagation. Portland: Timber Press. [ Links ]
Kuriyama, A., Kobayashi, T., & Maeda, M. (2004). Production of sporophytic plants of Cyathea lepifera, a tree fern, from in vitro cultured gametophyte. Journal of the Japanese Society of Horticulture Science, 73, 140-142. [ Links ]
Lorscheitter, M. L., Ashraf, A. R., Windisch, P. G., & Mosbrugger, V. (1999). Pteridophyte spores of Rio Grande do Sul flora, Brazil. Part II. Palaeontographica, 251, 71-235. [ Links ]
Marquez, G. J., & Brussa, C. A. (2011). First record of Cyatheaceae in Uruguay. American Fern Journal, 101, 205-205. [ Links ]
Mendoza-Ruiz, A., & Pérez-García, B. (2009). Morphogenesis of the gametophytes of eight mexican species of Blechnum (Blechnaceae). Acta Botanica Mexicana, 88, 59-72. [ Links ]
Meyer, B. S., Anderson, D. B., & Swanson, C. A. (1955). Laboratory Plant Physiology. New York: Van Nostrand. [ Links ]
Panthier, J. J., Diem, H. G., & Dommergues, Y. (1979). Rapid method to enumerate and isolate soil actinomycetes antagonistic towards rhizobia. Soil Biology & Biochemistry, 11, 443-445. [ Links ]
Pence, V. C. (2008). In vitro collecting for ex situ conservation. In T. A. Ranker, & C. Haufler (Ed.), Biology and Evolution of Ferns and Lycophytes (pp. 417-461). New York: Cambridge University Press. [ Links ]
Ponce, M. (1996). Pteridophyta. In F. O. Zuloaga, & O. Morrone (Eds.), Catálogo de las plantas vasculares de Argentina I: Pteridophyta, Gymnospermae y Angiospermae (Monocotyledoneae). Monographs in Systematic Botany from the Missouri Botanical Garden 60, 1-79. [ Links ]
Quintanilla, L. G., Amigo, J., Pangua, E., & Pajaron, S. (2002). Effect of storage method on spore viability in five globally threatened fern species. Annals of Botany, 90, 461-467. [ Links ]
Ranal, M. (1991). Germination of Polypodium hirsutissimum spores and antheridia formation in darkness. Revista Brasileira de Biologia, 51, 675-679. [ Links ]
Ranal, M. (1999). Effects of temperature on spore germination in some fern species from semideciduous mesophytic Forest. American Fern Journal, 89, 149-158. [ Links ]
Ranker, T. A., & Haufler, C. H. (2008). Biology and Evolution of Ferns and Lycophytes. New York: Cambridge University Press. [ Links ]
Rechenmacher, C., Schmitt, J. L. & Droste, A. (2010). Spore germination and gametophyte development of Cyathea atrovirens (Langsd. & Fisch.) Domin (Cyatheaceae) under different pH conditions. Brazilian Journal of Biology, 70, 1155-1160. [ Links ]
Renner, G. D. R., & Randi, A. M. (2004). Effects of sucrose and irradiance on germination and early gametophyte growth of the endangered tree fern Dicksonia sellowiana Hook. (Dicksoniaceae). Acta Botanica Brasilica, 18, 375-380. [ Links ]
Rogge, G. D., Viana, A. M., & Randi, A. M. (2000). Cryopreservation of spores of Dicksonia sellowiana, an endangered tree fern indigenous to South and Central America. Cryoletters, 21, 223-230. [ Links ]
Rojas, T., & Rondón, A. J. (1995). Control químico in vitro de Fusarium decemcellulare Brick aislado de mango. Agricultura Tropical, 45, 417-428. [ Links ]
Santos, E. P. G., Lehmann, D. R. M., Santos, M., & Randi, A. M. (2010). Spore germination of Gleichenella pectinata (Willd.) Ching (Polypodiopsida-Gleicheniaceae) at different temperatures, levels of light and pH. Brazilian Archives of Biology and Technology, 53, 1309-1318. [ Links ]
Schmitt, J. L., & Windisch, P. G. (2012). Caudex growth and phenology of Cyathea atrovirens (Langsd. & Fisch.) Domin (Cyatheaceae) in secondary forest, southern Brazil. Brazilian Journal of Biology, 72, 397-405. [ Links ]
Sehnem, A. (1978). Ciateáceas: Flora Ilustrada Catarinense. Itajaí: Herbário Barbosa Rodrigues. [ Links ]
Simabukuro, E. A., Dyer, A. F., & Felippe, G. M. (1998). The effect of sterilization and storage conditions on the viability of the spores of Cyathea delgadii Sternb. American Fern Journal, 88, 72-80. [ Links ]
Souza, R. N., Medeiros, M. B., & Mendes, R. A. (2007). Micropropagação e conservação do germoplasma de Dicksonia sellowiana (Hook.) (Dicksoniaceae) visando o repovoamento da área de influência do aproveitamento hidrelétrico Barra Grande, SC, RS. Anais do VIII Congresso de Ecologia do Brasil, Caxambu, Brazil. [ Links ]
Tryon, R. M., & Tryon, A. F. (1982). Ferns and allied plants with special reference to Tropical America. New York: Springer Verlag. [ Links ]
Viviani, D., & Randi, A. M. (2008). Effects of pH, temperature and light intensity on spore germination and growth analysis of young sporophytes of Polypodium lepidopterist (Langsd. & Fisch.) Kunze (Pteridophyta, Polypodiaceae). Rodriguésia, 59, 435-444. [ Links ]
Windisch, P. G. (2002). Fern conservation in Brazil. Fern Gazette, 16, 295-300. [ Links ]
Windisch, P. G., & Santiago, A. C. P. (2013). Cyatheaceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. Retrieved from http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB90850 [ Links ]
Wu, H., Ping-Tin, C., Li-Ping, Y., & Long-Qing, C. (2009). An efficient method for surface sterilization and sowing fern spores in vitro. American Fern Journal, 99, 226-230. [ Links ]
Zar, J. H. (1999). Biostatistical analyses. 4 ed. Upper Saddle River: Prentice-Hall. [ Links ]
*Correspondencia a:
Isabel Beatriz de Vargas. Laboratório de Biotecnologia Vegetal, Universidade Feevale, Rodovia RS 239, 2755, CEP 93352-000, Novo Hamburgo-RS, Brazil; isabelbeatriz@bol.com.br
Annette Droste. Laboratório de Biotecnologia Vegetal, Universidade Feevale, Rodovia RS 239, 2755, CEP 93352-000, Novo Hamburgo-RS, Brazil.
Programa de Pós-Graduação em Qualidade Ambiental, Universidade Feevale, Rodovia RS 239, 2755, CEP 93352-000, Novo Hamburgo-RS, Brazil; annette@feevale.br
1. Laboratório de Biotecnologia Vegetal, Universidade Feevale, Rodovia RS 239, 2755, CEP 93352-000, Novo Hamburgo-RS, Brazil; isabelbeatriz@bol.com.br
2. Laboratório de Biotecnologia Vegetal, Universidade Feevale, Rodovia RS 239, 2755, CEP 93352-000, Novo Hamburgo-RS, Brazil.
Programa de Pós-Graduação em Qualidade Ambiental, Universidade Feevale, Rodovia RS 239, 2755, CEP 93352-000, Novo Hamburgo-RS, Brazil; annette@feevale.br
Received 05-XII-2012. Corrected 20-VII-2013. Accepted 30-VIII-2013.


{kind=link}
{kind=link}
{kind=link}