










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.1 San José Jan./Mar. 2014
Awn length variation and its effect on dispersal unit burial of Trachypogon spicatus (Poaceae)
Efecto de la variación en longitud de la arista de la unidad dispersante de Trachypogon spicatus (Poaceae) en la profundidad de su entierro
Efecto de la variación en longitud de la arista de la unidad dispersante de Trachypogon spicatus (Poaceae) en la profundidad de su entierro
*Dirección para correspondencia:
Abstract
Trachypogon spicatus, formerly known as Trachypogon plumosus, is a dominant grass in some savannas of Northern South America. Its dispersal unit, like many other species of the Andropogoneae tribe, bears a hygroscopic awn which facilitates its establishment in favorable microsites. Some authors have previously proposed that there is a positive correlation between awn length and dispersal unit burial, and that this relationship increases the probability of seed survival in the event of a fire, since soil acts as insulator. In this study we experimentally tested this relationship for T. spicatus. A total of 192 diaspores were placed in randomized blocks, in aluminum trays filled with soil under greenhouse conditions. Diaspores were sprayed with water daily for a month to guarantee awn movement; on the last day of the experiment, they were sprayed with red aerosol paint to determine burial depth. The effects of awn length, presence of caryopses, and presence of a pivot for the passive segment of the awn on diaspore burial were evaluated. Germination viability was tested using a tetrazolium salt test for 35 caryopses. No significant differences in diaspore burial were observed between diaspores with and without caryopses (F(2,126)= 0.034, p=0.853). A positive correlation between awn length and diaspore burial was observed only if the passive awn lacked a pivot (r(66)=0.394, p<0.05). Diaspores whose awns had a pivot point achieved significantly deeper burial distances than their counterparts (F(2,126)=7.063, p=0.005). Viability test found that 0% of caryopses tested were able to germinate; this is possibly due to the time difference between sampling and testing. We considered the presence or absence of caryopsis as an important factor, since previous studies have not yet considered it and the high production of sterile diaspores in grasses. These results suggest that the physical mechanism behind T. spicatus diaspore burial is awn torque. This would explain why our results partially support the hypothesis previously proposed by other authors regarding the effect of awn length upon dispersal unit burial, but would also account for the differences observed in burial depth between diaspores with an available pivot point for the passive awn and diaspores lacking thereof. Rev. Biol. Trop. 62 (1): 321-326. Epub 2014 March 01.
Key words: hygroscopic awn, T. spicatus, diaspore burial, savanna, fire.
Resumen
Trachypogon spicatus es una gramínea dominante de algunas sabanas del norte de Suramérica. Su unidad dispersante presenta una arista higroscópica la cual facilita su inserción en irregularidades del suelo, facilitando su ubicación en microhábitats favorables para su establecimiento. Igualmente, se ha sugerido que existe una relación positiva entre la longitud de la arista y la capacidad de entierro de la unidad dispersante. Esta relación incrementa la probabilidad de supervivencia de la semilla ante el fuego al ser el suelo aislante de las altas temperaturas. Se colocaron 192 diásporas de T. spicatus, en bloques aleatorios, en bandejas de sustrato bajo condiciones de invernadero. Luego, se sometieron a cambios de humedad para garantizar el movimiento de la arista y pintaron con aerosol rojo el último día del experimento para determinar la profundidad de entierro. Se evaluó el efecto de la presencia de cariópsis, la longitud de la arista y la presencia de un punto de apoyo para la arista sobre la profundidad de entierro de la diáspora. Se realizó una prueba de sal de Tetrazolio para probar la viabilidad de germinación de 35 cariópsis. No se observaron diferencias significativas en la profundidad de entierro entre diásporas provistas y desprovistas de cariópsis (F(2,126)=0.034, p=0.853). La longitud de la arista parece guardar una relación positiva con la profundidad de entierro únicamente para aquellas unidades dispersantes cuyas aristas no cuentan con un punto de apoyo (r(66)=0.394, p<0.05). Por otro lado, aquellas diásporas cuyas aristas contaban con un punto de apoyo lograron profundidades de entierro significativamente mayores (F(2,126)=7.063, p=0.005). La prueba de viabilidad determinó que 0% de las cariópsis analizadas eran capaces de germinar, esto posiblemente sea debido a la diferencia entre el tiempo de recolección y de prueba. Consideramos que el factor presencia/ausencia de cariópsis era importante ya que había sido ignorado en estudios previos y por la alta producción de diásporas estériles en las gramíneas. Los resultados sugieren que el mecanismo físico que opera sobre la capacidad de entierro de la unidad dispersante de T. spicatus es el torque. Esto explica por qué solo se apoyaron parcialmente las hipótesis propuestas previamente por otros autores, y justifica las diferencias observadas en cuanto a la profundidad de entierro entre unidades dispersantes cuyas aristas contaban o carecían de un punto de apoyo.
Palabras clave: arista higroscópica, T. spicatus, diáspora, entierro, sabana, fuego.
Numerous grasses possess diaspores with specialized structures such as barbs, plumes or hygroscopic awns which aid their displacement and location on soil-surface irregularities (Peart, 1979; Peart, 1981; van der Pijl, 1982; Stamp, 1984; Cheplick, 1998; Garnier & Dajoz, 2000). This increases the probability of a seed being positioned in a microsite favorable for its establishment and germination, also known as a ‘safe-site’ (Harper, 1977; Fowler, 1988; Cheplick, 1998; Garnier & Dajoz, 2000; Schupp, Jordano & Gomez, 2010).
The hygroscopic awn of T. spicatus is composed of an active segment which is densely pubescent and contorted around its axis, and another passive segment which is smooth. Both segments are separated by a slight bend in the awn called the ‘knee’ (Peart, 1979). When the awn is moistened, the active segment unwinds along its axis until the passive segment touches the soil surface acting as a lever which allows the diaspore to move (Peart, 1979, 1981; Stamp, 1984; Fowler, 1988; Garnier & Dajoz, 2001; Wolgenmuth, 2009).
Garnier & Dajoz (2000) suggested that hygroscopic awns might be an adaptation in grasses native to fire-prone environments, such as savannas. They proposed that dispersal units (DUs) possessing longer awns achieve greater burial depth, thus increasing their probability of survival since soil shields the seed from high fire temperature.
However, there is no information regarding other factors that might affect diaspore burial, such as the presence or absence of caryopsis and/or the availability of a pivot point for the passive awn. On the other hand, very little is known about the physical mechanism behind awn movements and how it contributes to diaspore dispersal.
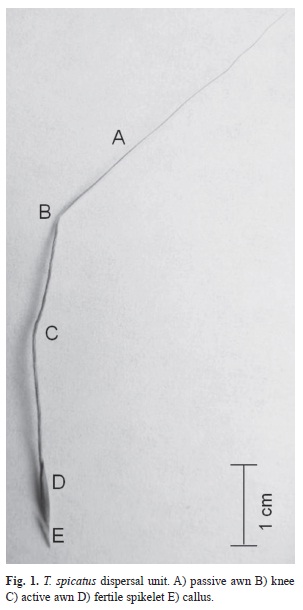
Trachypogon spicatus (L. f.) Kuntze is a perennial tussock grass dominant in the savannas of Northern South America (Luces de Febres, 1963; Baruch, Nassar & Bubis, 2004; Baruch, 2005a; Huber, 2007). Its dispersal unit has a hygroscopic awn (Fig. 1) which enables it to move and position itself in soil surface irregularities (Luces de Febres, 1963; Peart, 1979, 1981; Stamp, 1984; Fowler, 1988; Baskin & Baskin, 1998; Garnier & Dajoz, 2000; Garnier & Dajoz, 2001). Awn length in T. spicatus varies significantly within and among populations (Baruch, 2005b). This suggests that awn length is a trait subject to environmental selective pressures such as fire frequency and intensity. In populations where fire regimes are mild one would then expect higher awn length variability since selective pressures imposed by fires is fairly low and survival probability of any given awn length is high. On the other hand, as fire selective pressures become harsher; awn length variability would be diminished.
Here, we experimentally tested the effects of awn length, presence of caryopsis in the dispersal unit, and the availability of a pivot point on T. spicatus diaspore burial depth.
Materials and methods
Dispersal units of a T. spicatus population were collected on June 2009 in the vicinity of Puerto Ayacucho, Amazonas (5°41’35’’N- 67°34’59’’W). This population had both the longest awns and the highest variability of this trait (Baruch, 2005b). DUs were stored in paper bags at room temperature until the beginning of the experiment.
Germination viability was evaluated for 35 caryopses divided in five replica Petri dishes. These were soaked in water for 24 hours; afterwards, 2mL of tetrazolium salt (1%) with a Fig. 1. T. spicatus dispersal unit. A) passive awn B) knee C) active awn D) fertile spikelet E) callus. phosphate buffer were added to each Petri dish and placed in the dark for another 24 hours.
Diaspores were separated into those with and without caryopses (heavy and light). A total of 96 of each group were sown on a prepared substrate containing 80% sand (like the soil at Pto. Ayacucho) in aluminum trays of 16 x 23 x 5cm. DUs were placed on the substrate with the callus partially buried (following Peart, 1979) and arranged in 48 randomized blocks of four diaspores, each containing two heavy (with caryopsis) and two light (lacking caryopsis) diaspores. In order to avoid interference between awns and/or dislodged diaspores, these were separated by a grid of transparent acetate sheets (Peart 1979).
Substrate humidity was kept by piercing trays at the bottom and then submerging them, in groups of three, in a larger tray (32 x 52 x 10cm) filled with water, topped up when necessary (Peart, 1979). These larger trays were placed in a greenhouse with temperaturas varying between 15±2ºC and 29±3ºC (Bueno & Baruch, 2011) while photosynthetically active radiation was about 50% of that outdoors.
Dispersal units were sprayed with water daily at dusk, and were left to dry under direct sunlight during the day for 30 days. On the last day of the experiment, diaspores were sprayed with red aerosol paint so that the segment that remained exposed was painted. Diaspore burial was determined by subtracting painted length from total diaspore length (Garnier & Dajoz, 2000). DU lengths were measured with a digital caliper.
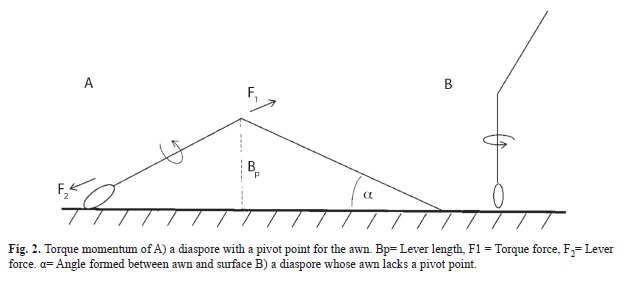
Awns were considered to possess a pivot if the passive awn touched any fixed surface (i.e. side of the tray, acetate sheet or soil surface) in a way that it might allow it to act as a lever (Fig. 2). The awn grants diaspores certain degree of mobility once placed on the soil (Peart, 1979). Because of this, we also counted the number of DUs that had moved at the end of the experiment.
Differences in awn length between heavy and light diaspores were determined with a Student’s T-test. The mobility of dispersal unit depending on caryopsis presence was analyzed with a Fisher’s exact test at the end of the experiment. The relationship between awn-length and dispersal unit burial depth was analyzed with a linear regression while the effect of presence/absence of caryopses and the availability of a pivot was determined with two-way ANOVAS. All statistical analyses were done with R V2.12 (R Development Core Team 2011).
Results
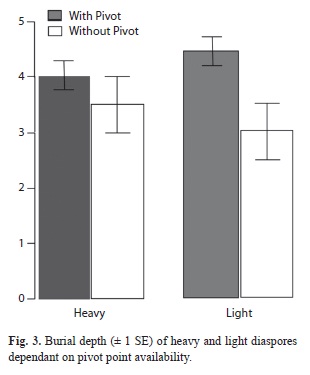
The tetrazolium test yielded a 0% germination viability of caryopses. Less than 2% of all examined DUs had caryopses (heavy). Their awns were significantly shorter than those belonging to light diaspores (t=-2.11, p=0.038). Dispersal units achieved burial depths that ranged from 0.10mm to 8.07mm which did not differ significantly between light and heavy diaspores (F(2,126)=0.034, p=0.853). The dispersal units that had an available pivot point achieved significantly greater burial depths regardless the presence of caryopses (F(2,126)=7.063, p=0.005) (Fig. 3).
At the end of the experiment 56.75% heavy and 75.0% light diaspores remained fixed on soil surface, indicating that diaspore weight did not affect its mobility (p=0.002). However, five heavy and six light DUs were able to re-insert themselves in the soil.
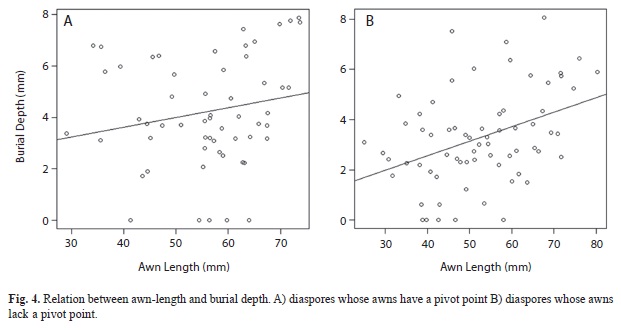
There was a positive relationship between awn-length and burial depth (Fig. 4). This relationship, however, was higher when the passive segment of the awn lacks a pivot point (r(66)=0.394, p<0.05) for DUs with awns lacking a pivot point and r(56)=0.196, p>0.05 for DUs with an available pivot.
Discussion
Seed burial depth is important for seed establishment and for the initial stages of plant growth. It offers fire and predator protection and improves germination. Therefore, traits that aid or promote seed burial such as a long hygroscopic awn should be favored by natural selection. However, this relationship is subjected to several variants such as the presence of caryopsis and the availability of a pivot.
The average awn length of T. spicatus dispersal units was longer in light diaspores. This might reflect a trade-off between energy and biomass allocated to awn length and that to caryopsis production. However, this tradeoff seems selectively illogical. Nevertheless, despite having generally longer awns, light diaspores did not achieve greater burial depths.
The reason why TZ test yielded 0% viability may be due to time difference between sampling (2009) and testing (2011). Seed and caryopsis viability tends to diminish significantly with time (Baruch com. pers.). We suggest that viability test be performed as soon as possible after sampling.
DU displacement was greater amongst heavy than light diaspores. However, we could not find a physical explanation to this difference which could be considered to be fortuitous due to the lack of air currents. Under natural conditions, we assume that wind would favor the movement of lighter dispersal units. The relationship between awn length and diaspore burial depth was independent of caryopsis presence. This might be because weight differences between heavy and light dispersal units are not enough to significantly affect burial depth. Although burial of diaspores lacking caryopses may not be ecologically relevant, since they are not able to yield offspring, this is a factor that has been ignored in previous studies (Peart, 1979; Peart, 1981; Garnier & Dajoz, 2000). We consider it should be contemplated due to the generally large production of light (sterile) dispersal units in grasses and its ecological implications since it involves energy input.
The tightly wound nature of the active awn grants the dispersal unit a certain amount of torque. Torque is defined as the tendency of a force to cause rotation around a given point (Resnick & Halliday, 1980; Sears, Zemansky, Young & Freedman, 2004). Tension naturally produced by this winding is released when the awn is wet, allowing it to rotate and expand.
DUs with a pivot achieved greater burial depths. Since the awn touches a fixed surface, it is able to act as a lever. In this case, torque would be defined by the following formula τ = F•b, and is directly proportional to the magnitude of the force (F) and lever length (b) (Resnick & Halliday, 1980; Sears et al., 2004). Lever length, however, depends on both passive and active awn lengths as well as the angle between the awn and soil surface. This explains why there was no significant correlation between awn length and burial depth in this case.
On the other hand, whenever awns are allowed to rotate freely, they are not able to act as levers. In this scenario, the awn rotates along an axis and torque; τ = I.α is proportional to its kinetic energy (moment of inertia, I) and angular acceleration (α).
Inertial momentum, defined as I = Σmiri2 is directly proportional to the distance of the body to the rotation axis (Resnick & Halliday, 1980; Sears et al., 2004). Hence, it is expected that diaspores bearing longer awns should achieve greater burial depths.
Previous research proposed that greater awn length results in deeper burial (Garnier & Dajoz, 2000). Nonetheless, our results partly support their hypothesis, particularly when diaspores lack a pivot for the passive awn.
Several considerations should be taken for future studies concerning the mechanics of the hygroscopic awn. Firstly, dispersal unit placement on substrate should be considered, especially regarding the position of the awn relative to soil surface and its ecological significance. On the other hand, it is necessary to consider the angle between the awn and soil surface since lever length is dependent on this. Though this was not contemplated in our experiment, it is considered that the effect of the angle between the awn and surface on burial depth should be studied in the future.
References
Baruch, Z., Nassar, J., & Bubis, J. (2004). Quantitative trait, genetic, environmental, and geographical distances among populations of the C4 grass Trachypogon plumosus in Neotropical savannas. Diversity and Distributions, 10, 283-292. [ Links ]
Baruch, Z. (2005a). Vegetation-environment relationships and classification of seasonal savannas in Venezuela. Flora, 200, 49-64. [ Links ]
Baruch, Z. (2005b). Trachypogon plumosus (POACEAE), un caso de alta diversidad interpoblacional en sabanas neotropicales. Interciencia, 30, 488-494. [ Links ]
Baskin, C., & Baskin, J. (1998). Seeds: ecology, biogeography and evolution of dormancy and germination. California: Academic Press. [ Links ]
Bueno, A., & Baruch, Z. (2011). Soil seed bank and the effect of needle litter layer on seedling emergence in a tropical pine plantation. Revista de Biología Tropical, 59(3), 1071-1079. [ Links ]
Cheplick, G. (1998). Seed dispersal and seedling establishment in grass populations. In G. Cheplick (Ed.), Population biology of grasses (pp. 84-106). London: Cambridge University Press. [ Links ]
Fowler, N. (1988). What is a safe site? Neighbor, litter, germination date, and patch effects. Ecology 69, 947-961. [ Links ]
Garnier, L., & Dajoz, I. (2000). Evolutionary significance of awn length variation in a clonal grass of fire-prone savannas. Ecology, 82, 1720-1733. [ Links ]
Garnier, L., & Dajoz, I. (2001). The influence of fire on demography of a dominant grass species of West African savannas, Hyparrhenia diplandra. Journal of Ecology, 89, 200-208. [ Links ]
Harper, J. L. (1977). Population biology of plants. London; New York, San Francisco: Academic Press. [ Links ]
Huber, O. (2007). Sabanas en los llanos venezolanos. In R. Duno de Stefano, G. Aymard, & O. Huber, (Eds.), Catálogo anotado e ilustrado de la flora vascular de los llanos de Venezuela. Caracas: FUDENA Fundación Polar. [ Links ]
Luces de Febres, Z. (1963). Las gramíneas del Distrito Federal. Caracas: Instituto Botánico, Dirección de Recursos Naturales Renovables, Ministerio de Agricultura y Cría. [ Links ]
Peart, M. (1979). Experiments on the biological significance on the morphology of seed-dispersal units in grasses. Journal of Ecology, 67, 843-863. [ Links ]
Peart, M. (1981). Further experiments on the biological significance of the morphology of seed-dispersal units in grasses. Journal of Ecology, 69, 425-436. [ Links ]
Resnick, R., & Halliday, D. (1980). Física: Parte 1. México D.F: John Wiley & Sons, Inc. [ Links ]
Stamp, N. E. (1984). Self-burial behaviour of Erodium cicutarium seeds. Journal of Ecology, 72, 611-620. [ Links ]
Schupp, E., Jordano, P., & Gomez, J. (2010). Seed dispersal effectiveness revisited: a conceptual review. New Phytologist, 188, 333-353. [ Links ]
Sears, F., Zemansky, M., Young, H., & Freedman, R. (2004). Física Universitaria. México D.F.: Pearson Educación. [ Links ]
Team, R. D. (2011). Retrieved from R: A Language and Environment for Statistical: http://www.R-project.org [ Links ]
van der Pijl, L. (1982). Principles of dispersal in higher plants. Berlin, Heidelberg, NewYork: Springer-Verlag. [ Links ]
Wolgenmuth, C. (2009). Plant biomechanics: Using shape to steal motion. Current Biology, 19(10), 409-410. [ Links ]
Baruch, Z. (2005a). Vegetation-environment relationships and classification of seasonal savannas in Venezuela. Flora, 200, 49-64. [ Links ]
Baruch, Z. (2005b). Trachypogon plumosus (POACEAE), un caso de alta diversidad interpoblacional en sabanas neotropicales. Interciencia, 30, 488-494. [ Links ]
Baskin, C., & Baskin, J. (1998). Seeds: ecology, biogeography and evolution of dormancy and germination. California: Academic Press. [ Links ]
Bueno, A., & Baruch, Z. (2011). Soil seed bank and the effect of needle litter layer on seedling emergence in a tropical pine plantation. Revista de Biología Tropical, 59(3), 1071-1079. [ Links ]
Cheplick, G. (1998). Seed dispersal and seedling establishment in grass populations. In G. Cheplick (Ed.), Population biology of grasses (pp. 84-106). London: Cambridge University Press. [ Links ]
Fowler, N. (1988). What is a safe site? Neighbor, litter, germination date, and patch effects. Ecology 69, 947-961. [ Links ]
Garnier, L., & Dajoz, I. (2000). Evolutionary significance of awn length variation in a clonal grass of fire-prone savannas. Ecology, 82, 1720-1733. [ Links ]
Garnier, L., & Dajoz, I. (2001). The influence of fire on demography of a dominant grass species of West African savannas, Hyparrhenia diplandra. Journal of Ecology, 89, 200-208. [ Links ]
Harper, J. L. (1977). Population biology of plants. London; New York, San Francisco: Academic Press. [ Links ]
Huber, O. (2007). Sabanas en los llanos venezolanos. In R. Duno de Stefano, G. Aymard, & O. Huber, (Eds.), Catálogo anotado e ilustrado de la flora vascular de los llanos de Venezuela. Caracas: FUDENA Fundación Polar. [ Links ]
Luces de Febres, Z. (1963). Las gramíneas del Distrito Federal. Caracas: Instituto Botánico, Dirección de Recursos Naturales Renovables, Ministerio de Agricultura y Cría. [ Links ]
Peart, M. (1979). Experiments on the biological significance on the morphology of seed-dispersal units in grasses. Journal of Ecology, 67, 843-863. [ Links ]
Peart, M. (1981). Further experiments on the biological significance of the morphology of seed-dispersal units in grasses. Journal of Ecology, 69, 425-436. [ Links ]
Resnick, R., & Halliday, D. (1980). Física: Parte 1. México D.F: John Wiley & Sons, Inc. [ Links ]
Stamp, N. E. (1984). Self-burial behaviour of Erodium cicutarium seeds. Journal of Ecology, 72, 611-620. [ Links ]
Schupp, E., Jordano, P., & Gomez, J. (2010). Seed dispersal effectiveness revisited: a conceptual review. New Phytologist, 188, 333-353. [ Links ]
Sears, F., Zemansky, M., Young, H., & Freedman, R. (2004). Física Universitaria. México D.F.: Pearson Educación. [ Links ]
Team, R. D. (2011). Retrieved from R: A Language and Environment for Statistical: http://www.R-project.org [ Links ]
van der Pijl, L. (1982). Principles of dispersal in higher plants. Berlin, Heidelberg, NewYork: Springer-Verlag. [ Links ]
Wolgenmuth, C. (2009). Plant biomechanics: Using shape to steal motion. Current Biology, 19(10), 409-410. [ Links ]
*Correspondencia a:
Erica E. Johnson. Laboratorio de Ecología Vegetal, Departamento de Estudios Ambientales. Universidad Simón Bolívar. Caracas, Venezuela; johnson.ojeda@gmail.com
Zdravko Baruch. Laboratorio de Ecología Vegetal, Departamento de Estudios Ambientales. Universidad Simón Bolívar. Caracas, Venezuela; zbaruc@usb.ve
1. Laboratorio de Ecología Vegetal, Departamento de Estudios Ambientales. Universidad Simón Bolívar. Caracas, Venezuela; johnson.ojeda@gmail.com
2. Laboratorio de Ecología Vegetal, Departamento de Estudios Ambientales. Universidad Simón Bolívar. Caracas, Venezuela; zbaruc@usb.ve
Received 12-iii-2013. Corrected 14-IX-2013. Accepted 15-X-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}